Abstract: To study the effects of cooling conditions in the root zone on frost injury in grapevine leaves, we used 1-year-old Merlot grapevine (Vitis vinifera) seedlings. The root zone was cooled regularly or buffered. All seedlings were treated in the simulated frost condition, then the frost index in leaves was calculated, and the chlorophyll fluorescence parameters were analyzed to reflect the change of PSII activity. Root temperature cooled regularly conferred severe frost damage in leaves, with a frost index of 74.36%. Buffered cooling alleviated frost injury to 53.29%, with a frost index of only 21.07%. Buffered cooling in the root zone improved qP and Fvʹ/Fmʹ in leaves during the recovery period, sped up the recovery of photochemical activity in PSII (ΦPSII), improved the ability of heat dissipation (NPQ), and relieved photo- inhibition. Buffered cooling in the root zone was conducive for leaves to recover after frost.
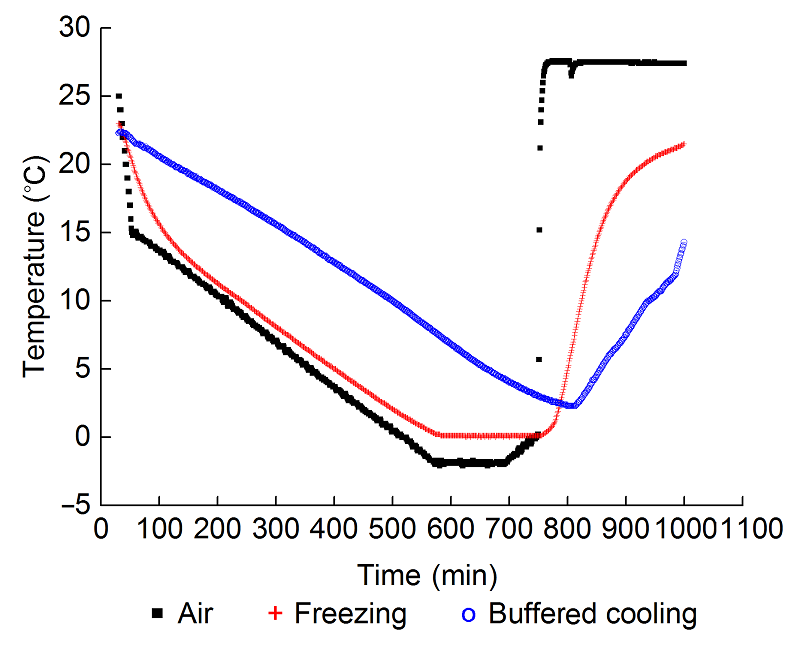
2 结果与讨论2.1 霜冻过程中气温和土壤温度的变化图1显示了霜冻及恢复过程中气温和根区土壤温度的变化。在降温阶段, 土壤温度会随着气温的降低而降低, 不同根区温度下降曲线基本平行, 说明二者的降温速度相近。当气温达到最低(-2°C)时, 冻根组的土壤温度也基本同步达到最低(-0.2°C), 而根区缓冲降温组的土壤温度在此时继续降低。低温维持阶段, 气温维持在-2°C, 冻根组的土壤温度维持在-0.2°C, 根区缓冲降温组的土壤温度继续降低。霜冻结束后, 将幼苗立即转移到25°C培养箱中恢复, 冻根组的土壤温度并不立即上升, 而是在最低温度附近维持了近20分钟才开始逐步升温, 冻根组的最低温从开始达到最低到开始回升共维持了174分钟; 根区缓冲降温组的土壤温度在此阶段进一步降低, 并在霜冻处理结束后1小时左右达到最低值(2.3°C), 最低温一共维持了15分钟, 之后逐渐回温。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_1.png<b>图1</b> 葡萄幼苗霜冻处理过程中气温和根区土壤温度的变化<br/>Air: 霜箱中空气温度; Freezing: 冻根处理; Buffered cooling: 根区缓冲降温<br/><b>Figure 1</b> Changes in temperatures of air and soil in the root zone during freezing treatment of grape leaves<br/>Air: Temperature in the freezer; Freezing: Roots were frozen during the period of cooling; Buffered cooling: Temperature in the root zone was buffered during the period of cooling Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_1.png<b>图1</b> 葡萄幼苗霜冻处理过程中气温和根区土壤温度的变化<br/>Air: 霜箱中空气温度; Freezing: 冻根处理; Buffered cooling: 根区缓冲降温<br/><b>Figure 1</b> Changes in temperatures of air and soil in the root zone during freezing treatment of grape leaves<br/>Air: Temperature in the freezer; Freezing: Roots were frozen during the period of cooling; Buffered cooling: Temperature in the root zone was buffered during the period of cooling
图1 葡萄幼苗霜冻处理过程中气温和根区土壤温度的变化 Air: 霜箱中空气温度; Freezing: 冻根处理; Buffered cooling: 根区缓冲降温 Figure 1 Changes in temperatures of air and soil in the root zone during freezing treatment of grape leaves Air: Temperature in the freezer; Freezing: Roots were frozen during the period of cooling; Buffered cooling: Temperature in the root zone was buffered during the period of cooling
2.2 根区缓冲降温对葡萄幼苗叶片冻害的影响在霜冻恢复过程结束后, 我们对2组处理的叶片冻害指数进行了统计分析。由表1可知, -2°C的低温霜冻对美乐葡萄幼苗叶片造成了不同程度的伤害。对根区进行不同的温度处理, 叶片的受冻程度不同。冻根组霜冻指数为74.36%, 而根区缓冲降温组叶片霜冻指数为21.07%, 比冻根组降低了53.29%, 且差异达到极显著水平(α=0.01)。 表1 Table 1 表1 表1 根区降温条件对葡萄叶片霜冻指数的影响 Table 1 Effects of temperature dropping down conditions in root zone on frost index of grape leaves
Number of frozen leaves
Number of leaves
Frost index (%)
Freezing
4.48±2.50
6.04±0.98
74.36±8.13
Buffered
1.56±0.49
7.78±2.01
21.07±6.61**
Index of frost was calculated as the ratio of leaves shown sign of frost injury. Data was expressed as means±SD, n=30. Difference in indexes of frost resulted from temperature dropping down conditions in root zone was compared by t-test, ** indicate significant difference at 0.01 level. 霜冻指数以表现冻害的叶片数与植株上叶片总数的比例表示, 结果以平均值±标准差表示, n=30。采用t-检验对两种根系降温方式下霜冻指数的平均值进行了比较, ** 代表在α=0.01水平下差异显著。
表1 根区降温条件对葡萄叶片霜冻指数的影响 Table 1 Effects of temperature dropping down conditions in root zone on frost index of grape leaves
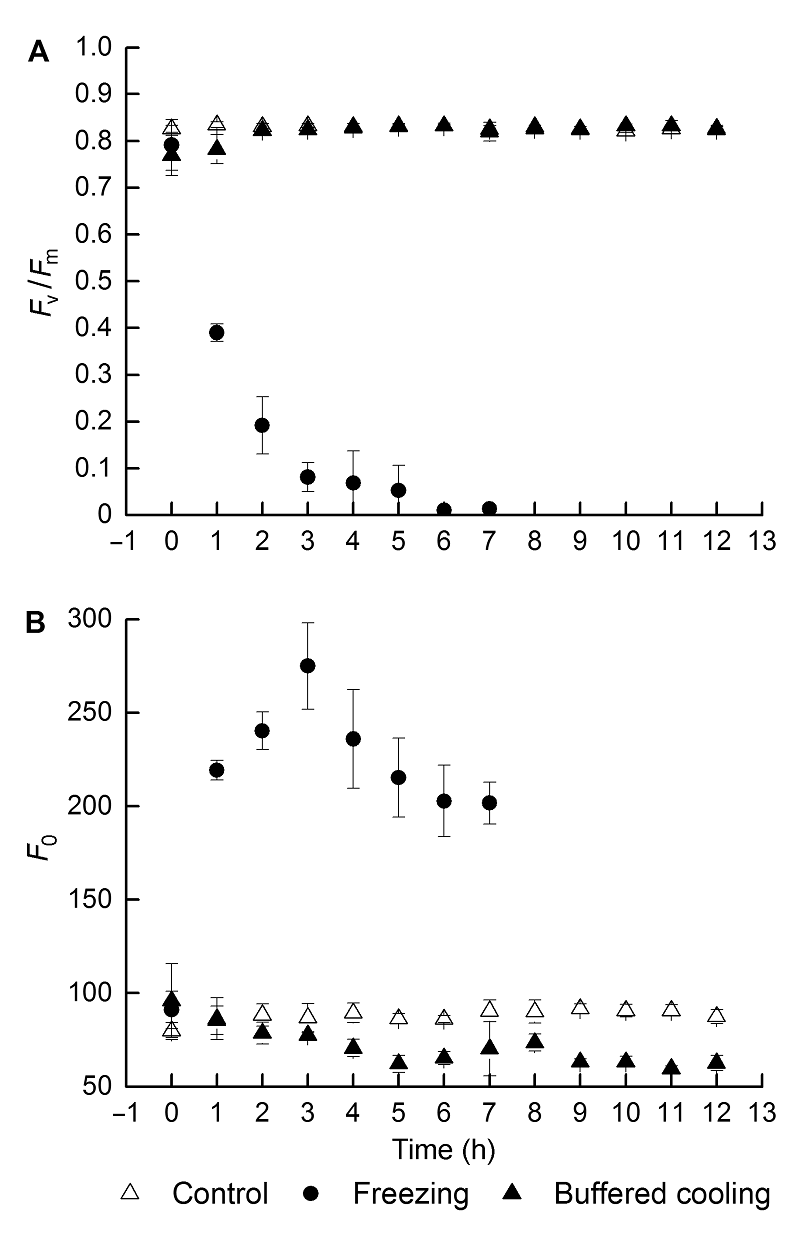
2.3 根区缓冲降温对葡萄幼苗叶片霜冻后恢复过程中叶片光抑制的影响由图2可知, 与对照相比, 霜冻处理造成叶片Fv/Fm下降, 且不同处理对Fv/Fm的下降幅度及恢复过程中Fv/Fm的变化有不同的影响。霜冻处理后, 冻根组的Fv/Fm下降到0.39, 在随后的恢复过程中, Fv/Fm持续下降, F0大幅度上升, 表明霜冻造成了PSII反应中心的不可逆失活。相比之下, 根区缓冲降温组经过霜冻处理后, Fv/Fm有小幅降低, 但并未达到受抑制的程度, 在恢复的第2小时, 基本达到霜冻处理前的水平, F0在霜冻后及恢复过程中基本处于下降的趋势, 表明PSII反应中心启动了保护性的能量耗散以避免霜冻后光抑制的发生。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_2.png<b>图2</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>F</i><sub>v</sub>/<i>F</i><sub>m </sub>(A)和<i>F</i><sub>0 </sub>(B)的影响(平均值±标准差, <i>n</i>=30)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率; <i>F</i><sub>0</sub>: 暗适应后的初始荧光强度<br/><b>Figure 2</b> Effects of delaying root zone temperature dropping down on <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub> (A) and <i>F</i><sub>0</sub> (B) of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximum quantum yield of PSII; <i>F</i><sub>0</sub>: Dark fluorescence yield Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_2.png<b>图2</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>F</i><sub>v</sub>/<i>F</i><sub>m </sub>(A)和<i>F</i><sub>0 </sub>(B)的影响(平均值±标准差, <i>n</i>=30)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率; <i>F</i><sub>0</sub>: 暗适应后的初始荧光强度<br/><b>Figure 2</b> Effects of delaying root zone temperature dropping down on <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub> (A) and <i>F</i><sub>0</sub> (B) of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximum quantum yield of PSII; <i>F</i><sub>0</sub>: Dark fluorescence yield
图2 根区缓冲降温对葡萄叶片霜冻后恢复过程中Fv/Fm (A)和F0 (B)的影响(平均值±标准差, n=30) Fv/Fm: 最大光化学效率; F0: 暗适应后的初始荧光强度 Figure 2 Effects of delaying root zone temperature dropping down on Fv/Fm (A) and F0 (B) of grape leaves after frost, during the period of recovery (means±SD, n=30) Fv/Fm: Maximum quantum yield of PSII; F0: Dark fluorescence yield
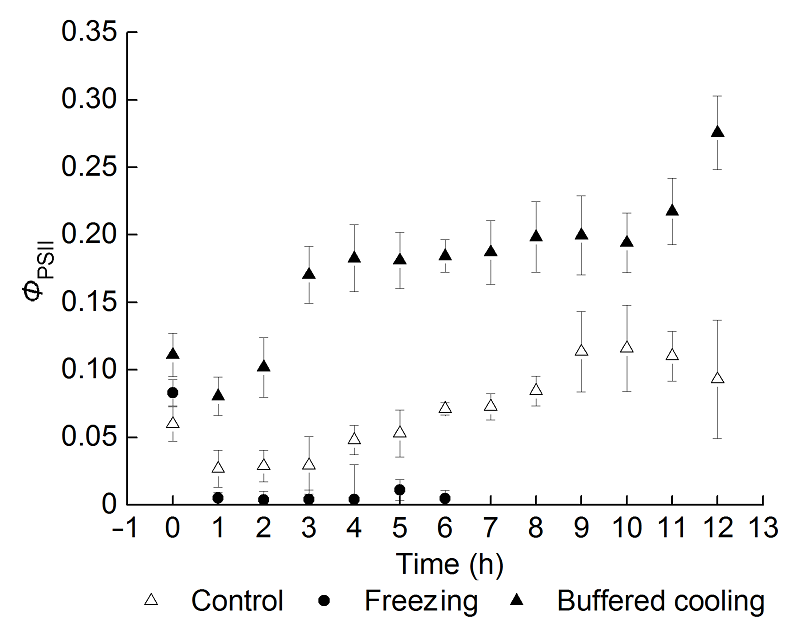
2.4 根区缓冲降温对葡萄幼苗叶片霜冻后恢复过程中ΦPSII的影响霜冻处理会造成ΦPSII的降低(图3)。正常条件下(对照组), 经过一夜的黑暗处理后ΦPSII有所下降, 但是进入恢复阶段后随着恢复时间的延长, ΦPSII逐渐恢复到处理前的水平并呈增加的趋势。霜冻后冻根组ΦPSII下降到很低的水平, 且随着恢复时间的延长不能恢复。根区缓冲降温处理的ΦPSII变化趋势与对照相似, 但在恢复期的第1小时就可以恢复到正常水平, 并且随着恢复时间的延长ΦPSII还显著高于正常水平。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_3.png<b>图3</b> 根区缓冲降温对霜冻后葡萄叶片恢复过程中<i>Φ</i><sub>PSII</sub>的影响(平均值±标准差, <i>n</i>=30)<br/><i>Φ</i><sub>PSII</sub>: PSII光合电子传递量子效率<br/><b>Figure 3</b> Effects of delaying root area temperature dropping down on <i>Φ</i><sub>PSII </sub>of grape leaves after frost, and during the period of recovery (means±SD, <i>n</i>=30)<br/><i>Φ</i><sub>PSII</sub>: Photochemical yield of photosystem II Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_3.png<b>图3</b> 根区缓冲降温对霜冻后葡萄叶片恢复过程中<i>Φ</i><sub>PSII</sub>的影响(平均值±标准差, <i>n</i>=30)<br/><i>Φ</i><sub>PSII</sub>: PSII光合电子传递量子效率<br/><b>Figure 3</b> Effects of delaying root area temperature dropping down on <i>Φ</i><sub>PSII </sub>of grape leaves after frost, and during the period of recovery (means±SD, <i>n</i>=30)<br/><i>Φ</i><sub>PSII</sub>: Photochemical yield of photosystem II
图3 根区缓冲降温对霜冻后葡萄叶片恢复过程中ΦPSII的影响(平均值±标准差, n=30) ΦPSII: PSII光合电子传递量子效率 Figure 3 Effects of delaying root area temperature dropping down on ΦPSII of grape leaves after frost, and during the period of recovery (means±SD, n=30) ΦPSII: Photochemical yield of photosystem II
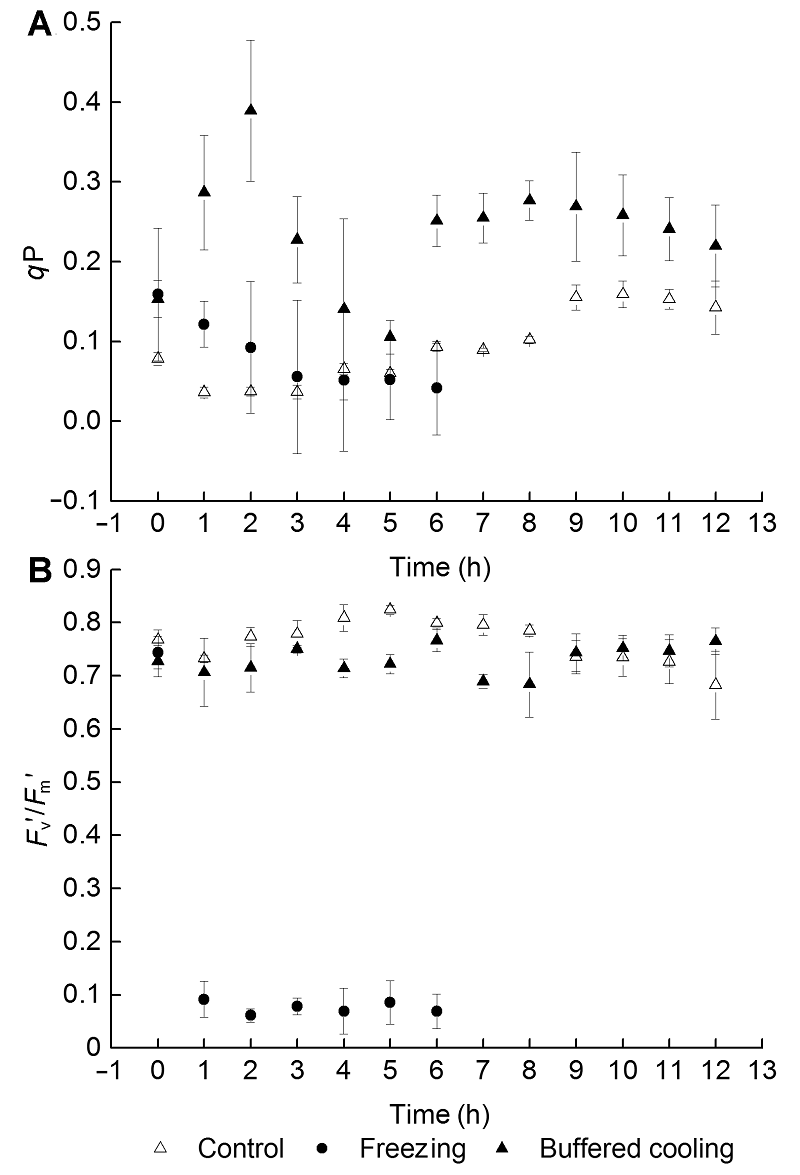
2.5 根区缓冲降温对葡萄幼苗叶片霜冻后恢复过程中qP和Fvʹ/Fmʹ的影响霜冻后, 冻根处理的qP降低, 且在恢复过程中持续降低。根区缓冲降温组的qP先迅速升高, 后大幅下降, 但没有显著低于正常水平, 之后又恢复到较高水平。根系受冻也造成Fvʹ/Fmʹ的大幅度降低且不可恢复。根区缓冲降温组的Fvʹ/Fmʹ处于波动状态, 但可维持在正常水平, 在恢复期的前9个小时内低于对照, 从第10小时开始高于对照(图4)。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_4.png<b>图4</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>q</i>P (A)和<i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i> (B)的影响(平均值±标准差, <i>n</i>=30)<br/><i>q</i>P: 光化学淬灭系数; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: 无线色素转化效率<br/><b>Figure 4</b> Effects of delaying root area temperature dropping down on <i>q</i>P (A) and <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i> (B) of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/><i>q</i>P: Coefficient of photochemical quenching; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: Quantum efficiency of open PSII reaction centers Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_4.png<b>图4</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>q</i>P (A)和<i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i> (B)的影响(平均值±标准差, <i>n</i>=30)<br/><i>q</i>P: 光化学淬灭系数; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: 无线色素转化效率<br/><b>Figure 4</b> Effects of delaying root area temperature dropping down on <i>q</i>P (A) and <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i> (B) of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/><i>q</i>P: Coefficient of photochemical quenching; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: Quantum efficiency of open PSII reaction centers
图4 根区缓冲降温对葡萄叶片霜冻后恢复过程中qP (A)和Fvʹ/Fmʹ (B)的影响(平均值±标准差, n=30) qP: 光化学淬灭系数; Fvʹ/Fmʹ: 无线色素转化效率 Figure 4 Effects of delaying root area temperature dropping down on qP (A) and Fvʹ/Fmʹ (B) of grape leaves after frost, during the period of recovery (means±SD, n=30) qP: Coefficient of photochemical quenching; Fvʹ/Fmʹ: Quantum efficiency of open PSII reaction centers
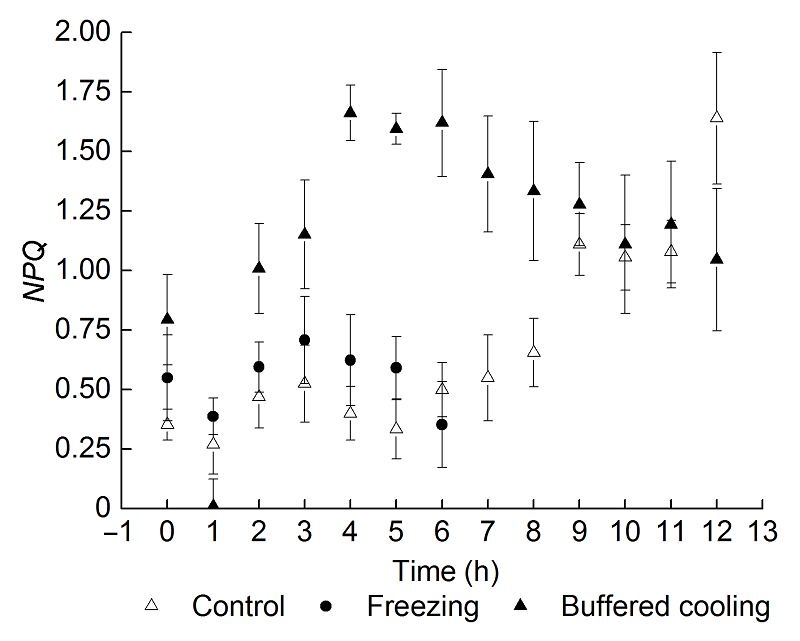
2.6 根区缓冲降温对葡萄幼苗叶片霜冻后恢复过程中热耗散的影响图5显示, 与对照相比, 2种根系处理方式的NPQ在霜冻后均显著降低, 且根区缓冲降温组的NPQ降低到几乎测不出来的程度。在恢复期, 冻根组的NPQ在恢复的第3小时达到最大, 较正常水平略有提高, 之后降低; 根区缓冲降温组的NPQ在恢复的第4小时达到最大, 且极显著高于正常水平, 之后逐渐降低, 但仍显著高于正常值。 图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_5.png<b>图5</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>NPQ</i>的影响(平均值±标准差, <i>n</i>=30)<br/>NPQ: 非光化学淬灭系数<br/><b>Figure 5</b> Effects of delaying root area temperature dropping down on <i>NPQ</i> of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/>NPQ: Non-photochemical quenching Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-290/img_5.png<b>图5</b> 根区缓冲降温对葡萄叶片霜冻后恢复过程中<i>NPQ</i>的影响(平均值±标准差, <i>n</i>=30)<br/>NPQ: 非光化学淬灭系数<br/><b>Figure 5</b> Effects of delaying root area temperature dropping down on <i>NPQ</i> of grape leaves after frost, during the period of recovery (means±SD, <i>n</i>=30)<br/>NPQ: Non-photochemical quenching
图5 根区缓冲降温对葡萄叶片霜冻后恢复过程中NPQ的影响(平均值±标准差, n=30) NPQ: 非光化学淬灭系数 Figure 5 Effects of delaying root area temperature dropping down on NPQ of grape leaves after frost, during the period of recovery (means±SD, n=30) NPQ: Non-photochemical quenching
2.7 讨论由于昼夜交替, 气温和土壤温度都处于波动状态, 且气温和土壤温度的变化并不同步, 这就使植物的叶片和根系分别处于不同的温度环境中。在正常情况下, 植物可以适应这种地上-地下的温度差异, 并使其各自以及整体的功能处于相对平衡状态。但极端的降温天气会打破这一平衡。因此有理由推测, 这种整体平衡性被打破可能会改变局部器官甚至是植物整体对环境的响应。前人的研究从侧面证实了这一推测(Huang et al., 1989; Zhou et al., 2007)。我们前期的研究表明, 不同的根系低温处理会对叶片冻害以及PSII电子传递链的供体侧、受体侧以及反应中心有不同的效应, 但缺乏定量的评估。因此, 本研究进一步对根系低温环境对叶片霜冻害的影响进行了评估, 发现当霜冻发生时, 地上部叶片和地下部根系所处的温度环境都会发生剧烈的变化(图1), 叶片和根系分别处于不同的低温胁迫状态下, 根区缓冲降温会减轻叶片的冻害程度, 使霜冻指数显著降低(表1)。实验结果进一步证明根系确实参与调节叶片对低温的响应能力。 已有研究表明, 低温会造成PSII反应中心的光损伤(Mauro et al.,1997; Heber et al., 2001; Velikova et al., 2005; Bertamini et al., 2006)。也有研究证实, 遇夜间低温后, 白天的光照会造成光抑制(胡文海等, 2005)。但根系能否调节霜冻后PSII反应中心的光抑制还未见报道。 Fv/Fm是检测和衡量光抑制程度的敏感指标(Nikiforou et al., 2011)。Fv/Fm下降到0.8以下即表明发生了显著的光抑制(Johnson et al.,1993)。本研究显示, 根区缓冲降温使Fv/Fm的降低程度减小, 并且有利于Fv/Fm的光下恢复(图2); 而冻根处理不仅会造成Fv/Fm的显著下降, 还会造成光下恢复时强烈的光抑制(图2)。对F0的进一步分析发现, 根区缓冲降温使F0在光下恢复过程中不断降低, 表明根区缓冲降温处理使PSII反应中心启动了保护性的散热机制, 以减少过剩光能对反应中心的伤害。而冻根处理的F0在光下恢复阶段处于较高的水平, 表明霜冻已经对PSII反应中心造成了不可逆的损伤, 照光后会加剧损伤。 霜冻会造成葡萄叶片实际光化学效率ΦPSII的显著下降, 但根区缓冲降温会使温度下降的程度减小, 并有利于光下的恢复(图3)。这表明根区缓冲降温增强了PSII反应中心可逆失活以减轻光抑制的能力。 qP和Fv'/Fm'共同决定ΦPSII值的大小(Demmig- Adams et al., 1996), qP反映了开放的PSII反应中心数目(Maxwell and Johnson, 2000), 而Fv'/Fm'则可以衡量开放的PSII反应中心的激发能捕获效率。本研究表明, 缓冲根区温度的降低使霜冻后叶片开放的PSII反应中心数目有所增加, 而反应中心的激发能捕获效率有所下降; 在恢复阶段, 由于光照增强, 开放的PSII反应中心数目减少, 而反应中心的激发能捕获效率有所升高, 使ΦPSII维持稳定且保持较高水平。冻根处理在霜冻后及光下恢复阶段, qP和Fv'/Fm'都大幅降低, 最终导致ΦPSII无法恢复。 热耗散是叶片减轻光抑制的有效途径(郭连旺和沈允钢, 1996)。本研究表明, 霜冻处理导致叶片热耗散能力显著降低。根区缓冲降温处理导致叶片热耗散能力更加降低, 但是光照后热耗散能力会有所回升, 随着光照时间的延长, NPQ会增加到较高的水平。冻根处理也会导致恢复期NPQ的先升高后降低, 但是其值比较低, 表明叶片的热耗散能力也受到了破坏, 导致光抑制的产生, 进一步对光反映中心造成破坏。 综上所述, 根系温度条件参与调节叶片对霜冻的反应, 进一步影响叶片在霜冻后光下的恢复, 根区缓冲降温有利于减轻美乐葡萄叶片的冻害程度。
The authors have declared that no competing interests exist.
BertaminiM, MuthuchelianK, RubiniggM, ZorerR, VelascoR, NedunchezhianN (2006). Low-night temperature increased the photo-inhibition of photosynthesis in grapevine (Vitis vinifera L. cv. Riesling) leaves. Environ Exp Bot 57, 25-31. [本文引用: 1]
[7]
Demmig-AdamsB, Adams IIIWW, BarkerDH (1996). Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation.Physiol Plant 98, 253-264. [本文引用: 1]
[8]
FengYL, CaoKF, FengZL (2002). Thermal dissipation, leaf rolling and inactivation of PSII reaction centers in Amomum villosum in diurnal course. J Tropic Ecol 18, 865-876.
[9]
HeberU, BukhovNG, ShuvalovVA, KobayashiY, LangeOL (2001). Protection of the photosynthetic apparatus against damage by excessive illumination in homoiohydric leaves and poikilohydric mosses and lichens.J Exp Bot 52, 1999-2006. [本文引用: 1]
[10]
HuangLK, WongSC, TerashimaI, ZhangX, LinDX, OsmondCB (1989). Chilling injury in mature leaves of rice. I. Varietal differences in the effects of chilling on canopy photosynthesis under simulated ‘dry cold dew wind’ conditions experienced in south-east China.Aust J Plant Physiol 16, 321-337. [本文引用: 2]
[11]
JohnsonGN, YoungAJ, ScholesJD (1993). The dissipation of excess excitation energy in British plant species.Plant Cell Environ 16, 673-679. [本文引用: 1]
[12]
MauroS, DaineseP, LannoyeR, BassiR (1997). Cold- resistant and cold-sensitive maize lines differ in the phosphorylation of the photosystem II subunit, CP29. Plant Physiol 115, 171-180. [本文引用: 1]
MolitorD, CaffarraA, SinigojP (2014). Late frost damage risk for viticulture under future climate conditions: a case study for the Luxembourgish winegrowing region.Aust J Grape Wine Res 20, 160-168. [本文引用: 1]
[15]
NikiforouC, NikolopoulosD, ManetasY (2011). The winter-red-leaf syndrome in Pistacia lentiscus: evidence that the anthocyanic phenotype suffers from nitrogen deficiency, low carboxylation efficiency and high risk of photo- inhibition. J Plant Physiol 168, 2184-2187. [本文引用: 1]
[16]
SonyaL (2014). Frost: Canberra sees the silver lining in damaging frosts.Wine Viticult J 29, 48. [本文引用: 1]
[17]
SugiuraT, SumidaH, YokoyamaS (2012). Overview of recent effects of global warming on agricultural production in Japan.Jpn Agr Res Q 46, 7-13. [本文引用: 1]
[18]
VelikovaV, PinelliP, LoretoF (2005). Consequences of inhibition of isoprene synthesis in Phragmites australis leaves exposed to elevated temperatures. Agr Ecosyst Environ 106, 209-217. [本文引用: 1]
[19]
ZhouY, HuangL, ZhangY, ShiK, YuJ, NoguésS (2007). Chill-induced decrease in capacity of RuBP carboxylation and associated H2O2 accumulation in cucumber leaves are alleviated by grafting onto figleaf gourd.Ann Bot 100, 839-848. [本文引用: 2]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}