Abstract: We investigated the effects of amino acid supplements on thallus growth, superoxide dismutase (SOD) activity and huperzine-A accumulation in Huperzia serrata by adding aspartic acid, lysine, and tryptophan at different concentrations into the medium. With aspartic acid, the dry weight of the thallus increased by 18.50%-25.75% and the huperzine-A accumulation was significantly higher than in the control. The optimal concentration of aspartic acid was 0.5 mmol·L-1. The relative growth of thallus was significantly higher with 1 mmol·L-1 tryptophan than in the control, but the huperzine-A accumulation was slightly lower than for the control. The relative growth rate, dry weight and huperzine-A accumulation was lower for thallus cultured with lysine than the control. The SOD activity of the thallus cultured with aspartic acid was increased properly, and the huperzine-A accumulation of the thallus was increased. However, huperzine-A accumulation of the thallus was suppressed when SOD activity remained high in the long-term.
Key words:Huperziaserrata ; huperzine A ; precursor ; superoxide dismutase
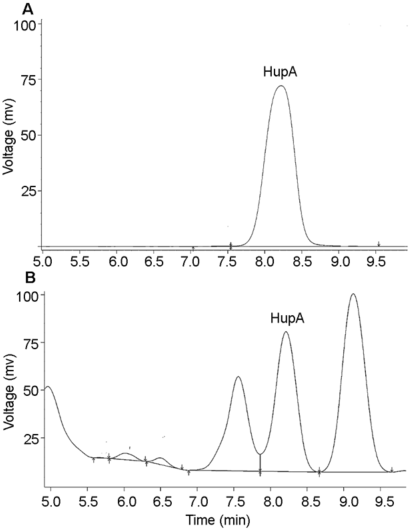
图1 石杉碱甲标准品(A)及蛇足石杉离体叶状体培养物提取物(B)的HPLC (高效液相色谱)色谱图 Figure 1 The HPLC (high performance liquid chromatography fluorescence) test of huperzine A standard (A) and extract of Huperzia serrata thallus in vitro culture (B)
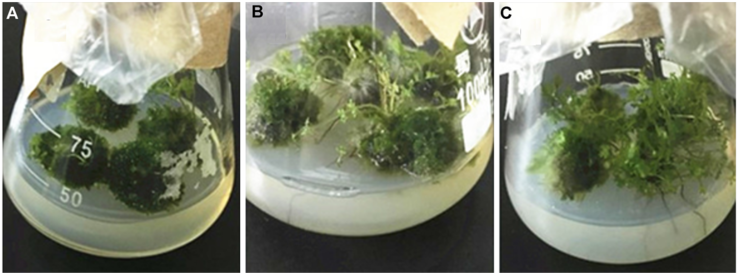
2 结果与讨论2.1 添加3种氨基酸对蛇足石杉叶状体生长增殖及干物质量的影响表1为3种氨基酸对蛇足石杉叶状体增殖率及干物质量的影响。在培养基中添加1 mmol·L-1 Trp后, 叶状体增殖率较对照提高1.24倍, 达显著水平; 而添加2 mmol·L-1 Trp或0.25 mmol·L-1 Lys的叶状体增殖率则明显低于对照; 其它各浓度氨基酸处理的叶状体增殖率与对照相比均未达到显著水平, 说明它们对叶状体生长增殖(鲜重)的影响不大(表1; 图2A-C)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_2.png<b>图2</b> 添加不同氨基酸培养的蛇足石杉叶状体<br/>(A) 添加天冬氨酸(Asp)培养的蛇足石杉叶状体; (B) 添加色氨酸(Trp)培养的蛇足石杉叶状体; (C) 添加赖氨酸(Lys)培养的蛇足石杉叶状体<br/><b>Figure 2</b> The thallus of <i>Huperzia serrata </i>supplemented with different amino acids<br/>(A) The thallus of <i>H. serrata </i>supplemented with Asp; (B) The thallus of <i>H. serrata </i>supplemented with Trp; (C) The thallus of <i>H. serrata </i>supplemented with Lys Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_2.png<b>图2</b> 添加不同氨基酸培养的蛇足石杉叶状体<br/>(A) 添加天冬氨酸(Asp)培养的蛇足石杉叶状体; (B) 添加色氨酸(Trp)培养的蛇足石杉叶状体; (C) 添加赖氨酸(Lys)培养的蛇足石杉叶状体<br/><b>Figure 2</b> The thallus of <i>Huperzia serrata </i>supplemented with different amino acids<br/>(A) The thallus of <i>H. serrata </i>supplemented with Asp; (B) The thallus of <i>H. serrata </i>supplemented with Trp; (C) The thallus of <i>H. serrata </i>supplemented with Lys
图2 添加不同氨基酸培养的蛇足石杉叶状体 (A) 添加天冬氨酸(Asp)培养的蛇足石杉叶状体; (B) 添加色氨酸(Trp)培养的蛇足石杉叶状体; (C) 添加赖氨酸(Lys)培养的蛇足石杉叶状体 Figure 2 The thallus of Huperzia serrata supplemented with different amino acids (A) The thallus of H. serrata supplemented with Asp; (B) The thallus of H. serrata supplemented with Trp; (C) The thallus of H. serrata supplemented with Lys
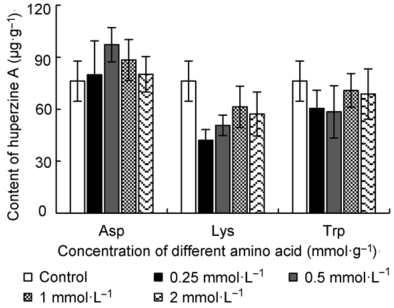
2.2 添加3种氨基酸对蛇足石杉叶状体积累石杉碱甲的影响采用HPLC检测不同浓度的3种氨基酸培养的蛇足石杉叶状体中的石杉碱甲含量(图3), 不同浓度Asp处理的叶状体石杉碱甲含量均高于对照(对照为76.25 µg·g-1), 其中以0.5 mmol·L-1 Asp培养效果最佳, 其含量高达97.23 µg·g-1, 比对照提高1.28倍; 用不同浓度的Trp处理, 除1 mmol·L-1 Trp培养叶状体石杉碱甲含量(70.801 µg·g-1)与对照相近, 其余浓度处理的叶状体中石杉碱甲含量均低于对照; 不同浓度Lys处理的叶状体中石杉碱甲含量均明显低于对照, 仅为(42.08-61.398) µg·g-1, 表明添加Lys抑制叶状体石杉碱甲的积累。 表1 Table 1 表1 表1 添加不同浓度的天冬氨酸(Asp)、赖氨酸(Lys)和色氨酸(Trp)的培养基对蛇足石杉叶状体生物量的影响 Table 1 Effect of different concentrations of Asp, Lys or Trp on the biomass of thallus in Huperzia serrata
Concentration of different amino acid (mmol·L-1)
Relative growth rate (%)
Dry weight (g·L-1)
Water/control
1313.05±21 bc
7.61680
Asp 0.25
1189.68±19 bc
9.30778
Asp 0.50
1337.76±29 bc
9.02623
Asp 1.00
1217.20±25 bc
9.57869
Asp 2.00
1211.28±32 bc
7.92295
Lys 0.25
1108.18±27 c
7.57992
Lys 0.50
1222.61±31 bc
7.40164
Lys 1.00
1328.25±30 bc
7.58278
Lys 2.00
1231.90±35 bc
7.32459
Trp 0.25
1311.53±33 bc
7.80328
Trp 0.50
1460.42±24 ab
8.01024
Trp 1.00
1647.51±37 a
9.35860
Trp 2.00
1050.53±17 c
6.66844
Different lowercase letters indicate significant difference at 0.05 level by Duncan’s new multiple range test. 不同小写字母表示Duncan新复极差法在0.05水平上差异显著。
表1 添加不同浓度的天冬氨酸(Asp)、赖氨酸(Lys)和色氨酸(Trp)的培养基对蛇足石杉叶状体生物量的影响 Table 1 Effect of different concentrations of Asp, Lys or Trp on the biomass of thallus in Huperzia serrata
图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_3.png<b>图3</b> 添加不同浓度的天冬氨酸(Asp)、赖氨酸(Lys)和色氨酸(Trp)对蛇足石杉叶状体中石杉碱甲含量的影响<br/><b>Figure 3</b> Effect of different concentrations of Asp, Lys and Trp on the content of huperzine A in thallus of <i>Huperzia serrata</i> Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_3.png<b>图3</b> 添加不同浓度的天冬氨酸(Asp)、赖氨酸(Lys)和色氨酸(Trp)对蛇足石杉叶状体中石杉碱甲含量的影响<br/><b>Figure 3</b> Effect of different concentrations of Asp, Lys and Trp on the content of huperzine A in thallus of <i>Huperzia serrata</i>
图3 添加不同浓度的天冬氨酸(Asp)、赖氨酸(Lys)和色氨酸(Trp)对蛇足石杉叶状体中石杉碱甲含量的影响 Figure 3 Effect of different concentrations of Asp, Lys and Trp on the content of huperzine A in thallus of Huperzia serrata
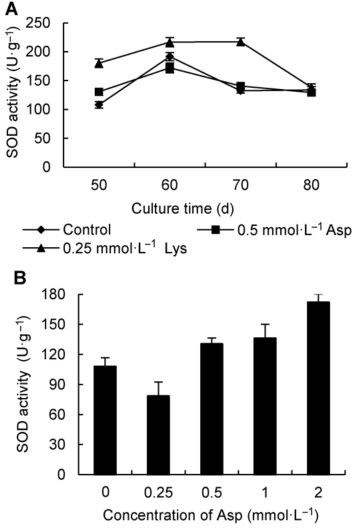
2.3 添加Asp与Lys对蛇足石杉叶状体中SOD酶活性的影响为探索不同氨基酸促进或抑制叶状体积累石杉碱甲的生化差别, 我们选择添加0.5 mmol·L-1 Asp及0.25 mmol·L-1 Lys培养的叶状体与对照叶状体, 分别检测其生长过程中SOD酶活性的变化趋势(图4A), 自添加Asp至叶状体收获(即培养50-80天), SOD酶活呈先增强后减弱的趋势(与对照SOD酶活性变化特征相似, 但在培养50天时SOD酶活性较高于对照)。其中, 培养60天的SOD酶活性最高(171.99 U·g-1)。添加0.25 mmol·L-1 Lys培养的叶状体在50、60及70天SOD酶活性均明显高于对照, 培养80天其活性与对照接近。以上研究结果表明, 添加0.5 mmol·L-1 Asp促进叶状体积累石杉碱甲与添加0.25 mmol·L-1 Lys抑制叶状体积累石杉碱甲的SOD酶活性有明显差异。 为探明添加不同浓度Asp促进叶状体积累石杉碱甲的SOD酶活性特点, 我们检测了不同浓度Asp培养的蛇足石杉叶状体培养50天的SOD酶活性(图4B)。结果表明, 添加0.25 mmol·L-1 Asp培养物SOD酶活性低于对照, 该处理积累石杉碱甲增效也不明显(图3), 其它处理的SOD酶活性均高于对照, 且培养物SOD酶活性随Asp添加浓度的升高而增强。以2 mmol·L-1 Asp培养的叶状体SOD酶活性最高, 但该处理积累石杉碱甲含量相对于0.5 mmol·L-1和1.0 mmol·L-1 Asp趋于下降(图3)。该结果进一步表明, 培养蛇足石杉叶状体时添加一定浓度的Asp, 培养50天适当提高SOD酶活性与石杉碱甲积累增加有关, 而当叶状体中SOD酶活性继续升高, 石杉碱甲积累量会受影响。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_4.png<b>图4</b> 添加不同氨基酸培养蛇足石杉叶状体的SOD酶活性特征<br/>(A) 添加0.5 mmol·L<sup>-1</sup>天冬氨酸(Asp)和0.25 mmol·L<sup>-1</sup>赖氨酸(Lys)培养叶状体SOD酶活性变化特征, (B) 添加不同浓度Asp培养50天叶状体SOD酶活性特征<br/><b>Figure 4</b> The change features of SOD activity for <i>Huperzia serrata</i> thallus supplemented with different amino acids<br/>(A) The change features of SOD activity for <i>H. serrata</i> thallus supplemented with 0.5 mmol·L<sup>-1</sup> Asp and 0.25 mmol·L<sup>-1 </sup>Lys, respectively; (B) The SOD activity of <i>H. serrata</i> thallus supplemented with different concentrations of Asp on 50 days Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-218/img_4.png<b>图4</b> 添加不同氨基酸培养蛇足石杉叶状体的SOD酶活性特征<br/>(A) 添加0.5 mmol·L<sup>-1</sup>天冬氨酸(Asp)和0.25 mmol·L<sup>-1</sup>赖氨酸(Lys)培养叶状体SOD酶活性变化特征, (B) 添加不同浓度Asp培养50天叶状体SOD酶活性特征<br/><b>Figure 4</b> The change features of SOD activity for <i>Huperzia serrata</i> thallus supplemented with different amino acids<br/>(A) The change features of SOD activity for <i>H. serrata</i> thallus supplemented with 0.5 mmol·L<sup>-1</sup> Asp and 0.25 mmol·L<sup>-1 </sup>Lys, respectively; (B) The SOD activity of <i>H. serrata</i> thallus supplemented with different concentrations of Asp on 50 days
图4 添加不同氨基酸培养蛇足石杉叶状体的SOD酶活性特征 (A) 添加0.5 mmol·L-1天冬氨酸(Asp)和0.25 mmol·L-1赖氨酸(Lys)培养叶状体SOD酶活性变化特征, (B) 添加不同浓度Asp培养50天叶状体SOD酶活性特征 Figure 4 The change features of SOD activity for Huperzia serrata thallus supplemented with different amino acids (A) The change features of SOD activity for H. serrata thallus supplemented with 0.5 mmol·L-1 Asp and 0.25 mmol·L-1 Lys, respectively; (B) The SOD activity of H. serrata thallus supplemented with different concentrations of Asp on 50 days
BeauchampC, FridovichI (1971). Superoxide dismutase: improved assays and an assay applicable to acrylamide gels.Anal Biochem 44, 276-287. [本文引用: 1]
[20]
HemscheidtT (2000). Tropane and Related Alkaloids. Biosynthesis. Vol. 209. Berlin: Springer. 175-206. [本文引用: 2]
[21]
ShuS, ZhaoX, WangW, ZhangG, CosoveanuA (2014). Identification of a novel endophytic fungus fromHuperzia serrata, which produces huperzine A. World J Microb Biot 30, 3101-3109. [本文引用: 1]
[22]
WuJ, GeX (2004). Oxidative burst, jasmonic acid biosynthesis, and taxol production induced by low-energy ultrasound inTaxus chinensis, cell suspension cultures. Biotechnol Bioeng 85, 714-721. [本文引用: 1]
[23]
ZhangG, WangW, ZhangX, XiaQ, ZhaoX, AhnY (2015). De novo RNA sequencing and transcriptome analysis ofColletotrichum gloeosporioides es026 reveal genes related to biosynthesis of huperzine A. PLoS One 10, e0120809. [本文引用: 1]
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}