李捷, 陈莹莹, 乔福云, 郅堤港, 郭正刚
, * 兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
Effects of disturbance by plateau pika on the β diversity of an alpine meadow Jie LI, Ying-Ying CHEN, Fu-Yun QIAO, Di-Gang ZHI, Zheng-Gang GUO
, * State Key Laboratory of Grassland Agro-ecosystems, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020, China
通讯作者: *(
guozhg@lzu.edu.cn )
编委: 张大勇
责任编辑: 赵航
收稿日期: 2020-08-10
接受日期: 2020-12-14
基金资助: 国家自然科学基金 (31172258 )西藏自治区牧草种质改良与利用重大专项 (XZ201901NA03 )
Corresponding authors: *(
guozhg@lzu.edu.cn )
Received: 2020-08-10
Accepted: 2020-12-14
Fund supported: the National Natural Science Foundation of China (31172258 )the Major Special Project for Improvement and Utilization of Forage Germplasm in Tibetan Autonomous Region (XZ201901NA03 )
摘要 β多样性反映生物群落沿某一环境梯度的物种周转速率, 该研究尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程。该研究利用野外随机样地的采集数据, 分析了高原鼠兔(
Ochotona curzoniae )不同干扰强度下Whittaker指数的变化特征, 并利用群落二元丰富度的方差分解法, 确定了单个物种(SCBD)和单个干扰位点(LCBD)对β多样性的贡献。主要结果: 随高原鼠兔干扰强度增加, 植物群落内物种周转速率呈先增加后降低的趋势; 占据位点数居中的物种对区域内的β多样性贡献较大, 其中冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )、小花草玉梅(
Anemone rivularis var.
flore-minore )等单个物种对整个区域内β多样性的贡献最为突出; 整个区域内干扰位点T
0 (高原鼠兔干扰强度为0)对区域β多样性贡献值最大, LCBD值和该位点的群落丰富度呈显著负相关关系, 但与高原鼠兔干扰强度无显著关联。说明重点保护LCBD值高的干扰位点所在的高寒草甸, 以及SCBD值较高的冰草、臭蒿、小花草玉梅, 对保护高原鼠兔存在时高寒草甸植物群落多样性具有重要意义。
关键词: 高原鼠兔 ;
高寒草甸 ;
β多样性 ;
干扰 ;
相关性 Abstract Aims β diversity reflects species turnover rate across environmental gradients, and this study attempts to use β diversity to reveal relevant ecological processes underlying the changes in plant community composition along a disturbance gradient induced by small burrowing herbivores.
Methods This study conducted a field survey at Gangcha County in the Qingzang Plateau to determine the effect of disturbance intensities of plateau pika (
Ochotona curzoniae ) on the β diversity of an alpine meadow measured by Whittaker’s index. Then a variance decomposition was used to measure the contribution of individual species to overall β diversity (SCBD) and single interference sites to overall β diversity (LCBD) according presence- absence community matrix.
Important findings The turnover rate of species in the plant community first increased and then decreased with the increase of the disturbance intensities induced by plateau pika. Species with intermediate site occupancy had a greater contribution to β diversity, in which
Agropyron cristatum, Artemisia hedinii and
Anemone rivular var.
flore-minore were the single-species plants that contributed the most to the β diversity of the study regions. The disturbance plot T
0 with the absence of plateau pika had the greatest contribution to the β diversity of whole study region. LCBD of individual site was negetively related to species richness of that individual site, and had no significant correlation with the disturbed intensity of plateau pika. These results indicate that alpine meadows distributed in high LCBD and
Agropyron cristatum, Artemisia hedinii and
Anemone rivularis var.
flore-minore with high SCBD should be protected to conserve plant diversity when the plateau pikas are present.
Keywords: plateau pika ;
alpine meadow ;
β diversity ;
disturbance ;
correlation PDF (597KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 李捷, 陈莹莹, 乔福云, 郅堤港, 郭正刚. 高原鼠兔干扰对高寒草甸β多样性的影响. 植物生态学报, 2021, 45(5): 476-486. DOI:
10.17521/cjpe.2020.0274 LI Jie, CHEN Ying-Ying, QIAO Fu-Yun, ZHI Di-Gang, GUO Zheng-Gang.
Effects of disturbance by plateau pika on the β diversity of an alpine meadow .
Chinese Journal of Plant Ecology , 2021, 45(5): 476-486. DOI:
10.17521/cjpe.2020.0274 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 )。植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 )。β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 )。大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献。
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性。
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持。然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注。
草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 )。已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 )。除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 )。小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 )。已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据。
高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 )。高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 )。虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据。因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据。
1 材料和方法 1.1 研究地区概况 研究地区位于青藏高原青海湖北岸, 行政区划隶属于青海省刚察县(99.33°-100.62° E, 36.97°- 38.07° N), 海拔3 300-3 800 m。气候属高原大陆性气候, 年降水量370 mm, 年蒸发量1 500-1 847 mm。1月平均气温-17.5 ℃, 7月平均气温11 ℃, 年平均气温-0.6 ℃左右。年日照时间为3 037 h, 年内霜期大于270天, 全年无绝对无霜期。土壤为亚高山草甸土。天然草地面积为71.2万hm
2 , 占全县总土地面积的73.8% (
索南拉毛, 2015 )。草地类型以高寒草甸为主, 优势种以莎草科的植物为主。高寒草甸虽然是优良放牧地, 但因其草层低矮而成为高原鼠兔的适宜分布区。
1.2 试验设计和调查 调查样区在刚察县哈尔盖镇的冷季放牧高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 主要伴生植物为莎草科的矮生嵩草(
Kobresia humilis )、禾本科的高原早熟禾(
Poa alpigena )和紫羊茅(
Festuca rubra )、菊科的美丽风毛菊(
Saussurea pulchra )和乳白香青(
Anaphalis lactea )、蔷薇科的莓叶委陵菜(
Potentilla fragarioides )等, 草层低矮、植株密聚。冷季放牧高寒草甸采用全年围封, 暖季(牧草生长季)不放牧, 冷季(牧草休眠期)打开围栏放牧, 因此从高寒草甸返青到植物枯黄期, 均无放牧动物干扰。野外调查时间为2019年7月底牧草生长旺盛时期, 该时期植物生物量最大, 高原鼠兔干扰也最为严重(
周雪荣等, 2010 )。首先, 选择地势相对平坦一致、草地类型一致的高寒草甸区, 确定是否有高原鼠兔干扰存在。高原鼠兔干扰的标准为高寒草甸内目视有高原鼠兔出没和有效洞口存在, 若存在高原鼠兔干扰, 则设置样地, 样地面积大小为35 m × 35 m, 接近高原鼠兔的巢域尺度(
Pang et al ., 2020 )。选定第1个样地后, 驱车继续前行距离超过5 km后, 选定第2样地。连续选定10个有高原鼠兔干扰的样地后, 在距离第10个样地不到5 km的范围内, 设置一个没有高原鼠兔干扰的样地, 其面积、地形地貌和植被类型均与所有干扰样地一致。高原鼠兔营家族式生活, 分布具有领域性, 因此高原鼠兔整体分布状态呈镶嵌分布, 因此, 有些高寒草甸虽然与干扰样地地貌、地形和植被类型一致, 调查时依然没有高原鼠兔干扰存在, 由于高原鼠兔种群扩散是一个逐渐的过程, 因此没有高原鼠兔干扰的样地仅是高原鼠兔潜在的适宜分布区, 因此设置非干扰样地(0干扰强度)是能够实现的。然后, 采用3天堵洞法测定每个干扰样地的有效洞口数。由于沿着公路布设样地, 而公路的走向并非直线, 这样客观上形成了一个样地群。以干扰样地内有效洞口密度测度该样地的干扰强度, 且将其视为一个位点, 其空间位置和干扰程度均存在差异。因未干扰样地没有高原鼠兔存在, 标记为T
0 , 然后根据每个干扰样地内的有效洞口密度, 由低到高标记为T
1 、T
2 、T
3 、T
4 、T
5 、T
6 、T
7 、T
8 、T
9 、T
10 , 共计11个位点(
图1 )(
Anderson et al ., 2011 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1高原鼠兔干扰下高寒草甸不同位点间的物种周转示意图。 Fig. 1Species turnover between different sites along the gradient of disturbance induced by plateau pika in an alpine meadow. 样地内高原鼠兔有效洞口密度调查完成的次日, 开始样方调查。每个样地内(干扰和未干扰)按“W”形设置5个样方, 样方面积为1 m × 1 m, 样方间距为8 m, 样方位于植被覆盖区, 避开洞口1 m。然后记录每个样方内出现的所有物种数, 确定样地累计出现物种数以及样地物种丰富度。共计调查55个样方。
1.3 指标计算 植物群落间β多样性的测度指标有多种, 本研究采用常用的Whittaker指数测定β多样性(
马克平等, 1995 ), 其计算公式为${\beta _{\rm{W}}} = (S{\rm{/}}X) - 1$, 式中:
S 为干扰样地内的累计出现物种数,
X 为样地内物种丰富度, 即各样方物种数的平均值。
采用植物群落二元丰富度的方差分解法分析单个物种和物种组合(位点)对调查区域内β多样性的贡献(
Legendre, 2007 ;
Legendre & de Cáceres, 2013 )。即将区域内不同物种在每个位点的分布数据表, 视为一系列环境变量(干扰强度)和空间变量(样地间距离)的函数, 从而将β多样性分解为单个位点(特定干扰强度下的物种组合)对β多样性的贡献(LCBD)和单个物种对β多样性的贡献(SCBD), 进而确定单个位点和单一物种对整个高原鼠兔干扰区域内β多样性的影响。
所有干扰位点植物群落的二元丰富度构成的矩 阵
VAR (
Y )具有
n 个干扰位点(行)和
p 个物种(列), 每个值用
yij 表示, ¯
yj 表示第
j 个物种二元丰富度的平均值, 在每个干扰位点, 某物种出现用“1”表示, 未出现则用“0”表示(
Legendre et al ., 2005 )。计算公式为:
(1) ${\beta _T} = VAR(Y) = \frac{{S{S_T}}}{{n - 1}} = \frac{{\sum\limits_{i = 1}^n {\sum\limits_{j = 1}^p {{{({y_{ij}} - {{\bar y}_j})}^2}} } }}{{n - 1}}$式中,
βT 为调查区域的β多样性大小,
SST 为矩阵$VAR(Y)$总的离均差平方和。
设$S{S_j}$和$S{S_j}$分别为物种
j 的方差和第
i 个位点的方差, 则第$j$个物种对β多样性的贡献$SCB{D_j}$和第
i 个位点对β多样性的贡献
LCBDi 分别表示为:
(2) $SCB{D_j} = S{S_j}{\rm{/}}S{S_T}$(3) $LCB{D_i} = S{S_i}{\rm{/}}S{S_T}$1.4 数据处理 采用Levene’s test检测每个样地物种数据的方差齐性, 若不符合正态分布, 则对数据进行对数转换, 使其满足正态分布。β多样性指数与有效洞口密度的相关性用Pearson相关系数衡量, 若显著相关, 则进行回归分析。Mantel检验可以检测群落变异与干扰梯度的变异是否具有相关性, 因此以样方丰富度差异衡量群落的结构变化, 以每个样地对应的有效洞口密度作为环境变量, 采用Mantel检验分析环境变化与群落异质性的关系, 设置999次随机排列重复得到相关性(
r )以及显著性, 若显著相关, 则进行回归分析。采用方差分解法确定SCBD和LCBD值, 分析过程采用R语言adespatial软件包完成: 首先采用“hellinger”转换, 使植物群落的二元丰富度表格具有Euclidean距离, 然后通过beta.div函数获取数据表的总离差平方和, 从而计算出SCBD和LCBD值。通过置换检验(Permutation text), 分析各个位点间的LCBD值是否有显著差异。以物种占据位点数为自变量, SCBD值为因变量, 利用Pearson相关法分析物种占据位点数和SCBD值的关系。为确定物种丰富度和干扰程度对LCBD的相对作用, 分别以物种丰富度、干扰强度为自变量, LCBD值为因变量, 采用Pearson相关法进行相关分析。以上数据分析和绘图均在R语言(4.0.1)环境下完成。
2 结果 2.1 高原鼠兔干扰区域内不同位点植物群落物种组成 不同干扰位点下(干扰强度)高原鼠兔有效洞口数量变化较大(
表1 ), 高原鼠兔干扰区域内有效洞口密度最低为335个·hm
-2 , 最高达1 633个·hm
-2 。不同干扰位点植物群落内累计物种数变化较大, 最小为21种, 最大为39种。物种丰富度随干扰强度增加表现为先增加后降低趋势, 物种数最低的位点是T
0 (干扰强度为0)和T
10 (干扰强度为1 633个·hm
-2 ), 最大的位点是T
4 (干扰强度为710个·hm
-2 )。
Table 1 表1 表1 高原鼠兔不同干扰位点下高寒草甸植物群落内累计物种数和物种丰富度
Table 1
Total species number and richness of plant communities under different disturbance by plateau pika in an alpine meadow 位点 有效洞口密度-2 ) 物种数1) 1) 丰富度2) 2) T0 0 22 20.2 T1 335 26 20.4 T2 417 30 20.6 T3 555 34 22.4 T4 710 39 23.4 T5 759 36 23.0 T6 906 31 19.6 T7 1 004 29 19.4 T8 1 186 27 17.6 T9 1 360 24 15.8 T10 1 633 21 13.8
1), 某一位点下所有样方的累计物种数; 2), 某一位点下各个样方内出现物种数的平均值。 1), total species of each site; 2), the average number of species in each site. 新窗口打开 |
下载CSV 11个干扰位点中共出现植物种数47种(
表2 ), 隶属于17个科。每个物种的分布位点存在差异, 有些物种分布位点多, 范围广, 譬如莎草科高山嵩草跨越11个位点; 有些物种分布的位点数少, 范围小, 如豆科的多枝黄耆(
Astragalus polycladus )分布于8个干扰位点, 而大戟科的地锦(
Euphorbia humifusa )仅分布于T
1 一个位点。
Table 2 表2 表2 物种占据的位点数
Table 2
Appearance site numbers of each species 物种 Species 位点数 No. of site 物种 Species 位点数 No. of site SP1 高山嵩草 Kobresia pygmaea 11 SP25 圆穗蓼 Polygonum macrophyllum 10 SP2 青藏薹草 Carex moorcroftii 11 SP26 西伯利亚蓼 Polygonum sibiricum 3 SP3 高山豆 Tibetia himalaica 11 SP27 酸模 Rumex acetosa 5 SP4 多枝黄耆 Astragalus polycladus 8 SP28 珠芽蓼 Polygonum viviparum 4 SP5 黄花棘豆 Oxytropis ochrocephala 11 SP29 多裂委陵菜 Potentilla multifida 9 SP6 披针叶黄华 Thermopsis lanceolata 4 SP30 二裂委陵菜 Potentilla bifurca 7 SP7 针茅 Stipa capillata 6 SP31 莓叶委陵菜 Potentilla fragarioides 11 SP8 紫羊茅 Festuca rubra 11 SP32 鹅绒委陵菜 Potentilla anserina 5 SP9 垂穗披碱草 Elymus nutans 4 SP33 獐牙菜 Swertia bimaculata 7 SP10 高原早熟禾 Poa alpigena 11 SP34 达乌里秦艽 Gentiana dahurica 9 SP11 冰草 Agropyron cristatum 5 SP35 刺芒龙胆 Gentiana aristata 11 SP12 美丽风毛菊 Saussurea pulchra 11 SP36 矮生嵩草 Kobresia humilis 8 SP13 矮火绒草 Leontopodium nanum 8 SP37 肉果草 Lancea tibetica 10 SP14 蒲公英 Taraxacum mongolicum 8 SP38 婆婆纳 Veronica didyma 5 SP15 甘肃马先蒿 Pedicularis kansuensis 8 SP39 打碗花 Calystegia hederacea 6 SP16 紫苑 Aster tataricus 4 SP40 车前 Plantago asiatica 9 SP17 臭蒿 Artemisia hedinii 6 SP41 狼毒 Euphorbia fischeriana 7 SP18 乳白香青 Anaphalis lactea 9 SP42 微孔草 Microula sikkimensis 3 SP19 细叶亚菊 Ajania tenuifolia 8 SP43 长茎藁本 Ligusticum thomsonii 3 SP20 唐松草 Thalictrum aquilegifolium 10 SP44 地锦 Euphorbia humifusa 1 SP21 钝裂银莲花 Anemone obtusiloba 6 SP45 鸢尾 Iris tectorum 3 SP22 高原毛莨 Ranunculus tanguticus 4 SP46 女娄菜 Silene aprica 4 SP23 疏齿银莲花 Anemone obtusiloba 2 SP47 细果角茴香 Hypecoum leptocarpum 2 SP24 小花草玉梅 Anemone rivularis var. flore-minore 4
新窗口打开 |
下载CSV 2.2 高原鼠兔干扰对群落间β多样性的影响 高原鼠兔干扰下, 植物群落β多样性与干扰位点有效洞口密度显著相关(
p < 0.05)。当高原鼠兔有效洞口密度从0增加到1 633个·hm
-2 时(干扰位点从T
0 到T
10 ), 群落β多样性指数表现为先升高后缓慢降低的过程, 有效洞口密度为710个·hm
-2 (干扰位点为T
4 )时β多样性指数最大(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同干扰位点有效洞口密度下β多样性的变化。阴影部分表示95%的置信区间。 Fig. 2Variation of the β diversity of the plateau pika under different disturbance intensities. The shaded part indicates the 95% confidence interval. Mantel检验表明高原鼠兔不同干扰位点的群落结构差异与环境差异显著相关(
r = 0.552 5,
p = 0.004)。群落矩阵和环境矩阵距离的散点图回归拟合结果表明, 随着高原鼠兔干扰下环境差异的增加, 植物群落的异质性整体表现为增加趋势(
图3 )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3高原鼠兔干扰梯度下群落差异与环境差异的关系。 Fig. 3Relationship between community deviation and environment deviation along the gradient of disturbance of plateau pika. 2.3 高原鼠兔干扰下单一物种对β多样性的贡献 统计和计算结果表明, 调查区域内植物种类
SST , 即总的离均差平方和为2.795 5,
βT 为0.279 6。SCBD平均值为0.021 3, 47个物种中有28个物种超过平均值(
图4 )。SCBD最小值为0.001 3, 主要源自广泛分布的高山嵩草、高山豆(
Tibetia himalaica )、黄花棘豆(
Oxytropis ochrocephala )、紫羊茅、高原早熟禾、美丽风毛菊、莓叶委陵菜等物种的贡献; SCBD最大值为0.033 6, 由冰草(
Agropyron cristatum )贡献, 其次是臭蒿(
Artemisia hedinii )、小花草玉梅(
Anemone rivularis var.
flore-minore )、獐牙菜(
Swertia bimaculata )、婆婆纳(
Veronica didyma )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4单一物种对高寒草甸β多样性的贡献(SCBD)值(A)及其分布(B)。物种同表2。 Fig. 4Value (A) and distribution (B) of the individual species contribution to beta diversity (SCBD) in an alpine meadow. See Table 2 for species. 单一物种对β多样性的贡献和该物种在调查区内所占有的干扰位点数表现出了高度关联性(
p < 0.001)。SCBD值和物种占据的位点数表现为开口向下的抛物线关系(
图5 ), 占据位点数为中等水平的物种对整个调查区域的β多样性贡献较大, 例如臭蒿、针茅(
Stipa sp.)、冰草等, 而占据位点数较多或较少的物种, 对整个调查区内的β多样性贡献较低, 例如高山嵩草、黄花棘豆、高山豆等。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5单一物种对高寒草甸β多样性的贡献(SCBD)值与物种占据位点数关系。阴影部分表示95%的置信区间。 Fig. 5Relationship between species contribution to beta diversity (SCBD) values and the numbers of sites occupied by species in an alpine meadow. The shaded part indicates the 95% confidence interval. 2.4 高原鼠兔干扰下单一位点对β多样性的贡献 不同位点对区域内β多样性的贡献存在差异, 所有干扰位点的平均贡献度为0.091 (
图6 ), 其中4个位点的LCBD值超过平均值, 依次为T
0 、T
1 、T
6 、T
10 。LCBD最大值为0.131 7, 源自位点T
0 , LCBD值最小为0.064 3, 属于位点T
5 。置换检验表明, 干扰位点T
0 (
p = 0.007)、T
1 (
p = 0.048)、T
10 (
p = 0.04)的LCBD值分别与其他干扰位点具有显著差异, 其他各干扰位点的LCBD值之间并不存在显著差异。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6单一位点对高寒草甸β多样性贡献(LCBD)值(A)及其分布(B)。 Fig. 6Value (A) and distribution (B) of the local contribution to beta diversity (LCBD) for each distribution site in an alpine meadow. 高原鼠兔干扰下, LCBD值与物种丰富度呈显著负相关关系(
p < 0.001), 即随着位点物种丰富度的增加, LCBD值表现为逐渐降低的变化过程。各位点的有效洞口数与LCBD值之间没有明显的拟合关系(
p > 0.05)(
图7 )。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7单一位点对高寒草甸β多样性贡献(LCBD)值与物种丰富度(A)和有效洞口密度(B)的关系。阴影部分表示95%的置信区间。 Fig. 7Relationship between the local contribution to beta diversity (LCBD) with species richness (A) and active burrow entrances (B) in an alpine meadow. The shaded part indicates the 95% confidence interval. 3 讨论 自
Legendre和de Cáceres (2013) 提出通过群落二元丰富度的方差分解来估计个别物种和物种组合对β多样性贡献的研究方法后, β多样性的研究逐渐从群落间的物种周转率向物种和物种组合对一定空间范围内β多样性的相对重要性转变, 从而根据β多样性的变化原因, 提出更具有针对性的生物多样性保护策略(
Qiao et al ., 2015 ;
Wang et al ., 2020 )。本研究在分析高原鼠兔干扰下群落间β多样性变化的基础上, 采用群落二元丰富度数据的方差分解法分析了高原鼠兔干扰区域内, 单个物种和单个位点(物种组合)对植物群落β多样性的相对重要性, 以期确定单个物种、单一位点, 或两者结合对植物群落β多样性保护的意义。
高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 )。研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化。高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加。然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数。
整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大。而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 )。同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因。本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异。
干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 )。本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 )。其中T
0 、T
1 、T
10 位点对β多样性的贡献最大。T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特。T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性。T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性。位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小。在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异。因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 )。研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 )。本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 )。尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系。因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点。而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合。
本研究表明, β多样性能够度量高原鼠兔干扰对高寒草甸总体多样性的影响, 更能量化单个物种组成和物种组合对植物群落物种组成差异的影响, 主要是β多样性能够评价高原鼠兔干扰对高寒草地植物多样性的影响从样方尺度向较大空间尺度的转变, 从而有助于从位点和单个物种的视角, 提出保护高寒草甸植物多样性的策略。高原鼠兔干扰通过形成洞口、裸斑、重新分配土壤、选择性采食、与土壤动物互作等方式, 增加了高寒草甸的异质性, 改变了植物群落间的物种周转速率。优势种对区域β多样性的贡献较低, 而杂类草对区域β多样性的贡献较大。若要维持整个区域的β多样性, 应该重点保护LCBD值较高的T
0 、T
1 、T
10 位点, 因为这3个位点的物种组成具有较高的独特性, 对高原鼠兔干扰区域β多样性贡献更高, 虽然其物种丰富度较低, 但具有较高的物种保护或恢复价值。保护独特性较高位点的同时, 还需要保护SCBD值高的单个物种, 如冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳等。通过采用植物群落二元丰富度的方差分解法分析高原鼠兔干扰对高寒草甸植物群落β多样性的影响, 明确了需要保护的位点(物种组合)和单个物种, 一定程度上证实了该方法能够用于草地植物群落β多样性的研究, 提出了高原鼠兔干扰下植物群落多样性的保护需要采用位点和物种结合的策略。
[1] Anderson MJ Crist TO Chase JM Vellend M Inouye BD Freestone AL Sanders NJ Cornell HV Comita LS Davies KF Harrison SP Kraft NJB Stegen JC Swenson NG 2011 ).Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologistEcology Letters ,14, 19 -28 . DOI:
10.1111/j.1461-0248.2010.01552.x PMID:
21070562 [本文引用: 2] A recent increase in studies of β diversity has yielded a confusing array of concepts, measures and methods. Here, we provide a roadmap of the most widely used and ecologically relevant approaches for analysis through a series of mission statements. We distinguish two types of β diversity: directional turnover along a gradient vs. non-directional variation. Different measures emphasize different properties of ecological data. Such properties include the degree of emphasis on presence/absence vs. relative abundance information and the inclusion vs. exclusion of joint absences. Judicious use of multiple measures in concert can uncover the underlying nature of patterns in β diversity for a given dataset. A case study of Indonesian coral assemblages shows the utility of a multi-faceted approach. We advocate careful consideration of relevant questions, matched by appropriate analyses. The rigorous application of null models will also help to reveal potential processes driving observed patterns in β diversity.© 2010 Blackwell Publishing Ltd/CNRS.
[2] Archidona-Yuste A Wiegand T Castillo P Navas-Cortés JA 2020 ).Spatial structure and soil properties shape local community structure of plant-parasitic nematodes in cultivated olive trees in southern SpainAgriculture, Ecosystems & Environment ,287, 106688 . DOI:10.1016/j.agee.?2019.?106688 . DOI:
10.1016/j.agee.?2019.?106688 URL [本文引用: 1] [3] Bagchi S Namgail T Ritchie ME 2006 ).Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of Trans-HimalayaBiological Conservation ,127, 438 -442 . DOI:
10.1016/j.biocon.2005.09.003 URL [本文引用: 3] [4] Bakker ES Ritchie ME Olff H Milchunas DG Knops JMH 2006 ).Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore sizeEcology Letters ,9, 780 -788 . DOI:
10.1111/ele.2006.9.issue-7 URL [本文引用: 1] [5] Bangert RK Slobodchikoff CN 2006 ).Conservation of prairie dog ecosystem engineering may support arthropod beta and gamma diversityJournal of Arid Environments ,67, 100 -115 . [本文引用: 1] [6] da Silva PG Hernández MIM Heino J 2018 ).Disentangling the correlates of species and site contributions to beta diversity in dung beetle assemblagesDiversity and Distributions ,24, 1674 -1686 . DOI:
10.1111/ddi.2018.24.issue-11 URL [本文引用: 1] [7] Gaston KJ Borges PAV He F Gaspar C 2006 ).Abundance, spatial variance and occupancy: arthropod species distribution in the AzoresJournal of Animal Ecology ,75, 646 -656 . DOI:
10.1111/jae.2006.75.issue-3 URL [本文引用: 1] [8] Gibson DJ 1989 ).Effects of animal disturbance on tallgrass prairie vegetationAmerican Midland Naturalist ,121, 144 -154 . DOI:
10.2307/2425665 URL [本文引用: 1] [9] Guo ZG Li XF Liu XY Zhou XR 2012 ).Response of alpine meadow communities to burrow density changes of plateau pika (Ochotona curzoniae ) in the Qinghai-Tibet PlateauActa Ecologica Sinica 44 -49 . DOI:
10.1016/j.chnaes.2011.12.002 URL [本文引用: 1] [10] Hagenah N Bennett NC 2013 ).Mole rats act as ecosystem engineers within a biodiversity hotspot, the Cape FynbosJournal of Zoology ,289, 19 -26 . DOI:
10.1111/jzo.2013.289.issue-1 URL [本文引用: 1] [11] Jia TT Mao L Guo ZG 2014 ).Effect of available burrow densities of plateau pika (Ochotona curzoniae ) on plant niche of alpine meadow communities in the Qinghai-Tibet PlateauActa Ecologica Sinica 869 -877 . [本文引用: 1] [贾婷婷 ,毛亮 ,郭正刚 (2014 ).高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响生态学报 ,34, 869 -877 .] [本文引用: 1] [12] Jones CC Halpern CB Niederer J 2008 ).Plant succession on gopher mounds in western Cascade meadows: consequences for species diversity and heterogeneityThe American Midland Naturalist ,159, 275 -286 . DOI:
10.1674/0003-0031(2008)159[275:PSOGMI]2.0.CO;2 URL [本文引用: 1] [13] Legendre P 2007 ).Studying beta diversity: ecological variation partitioning by multiple regression and canonical analysisJournal of Plant Ecology (Chinese Version )976 -981 . [本文引用: 1] [Legendre P 2007 ).β-多样性的研究: 应用多元回归和典范分析研究生态方差的分解植物生态学报 ,31, 976 -981 .] [本文引用: 1] [14] Legendre P Borcard D Peres-Neto PR 2005 ).Analyzing beta diversity: partitioning the spatial variation of community composition dataEcological Monographs ,75, 435 -450 . DOI:
10.1890/05-0549 URL [本文引用: 1] [15] Legendre P de Cáceres M 2013 ).Beta diversity as the variance of community data: dissimilarity coefficients and partitioningEcology Letters ,16, 951 -963 . DOI:
10.1111/ele.12141 PMID:
23809147 [本文引用: 4] Beta diversity can be measured in different ways. Among these, the total variance of the community data table Y can be used as an estimate of beta diversity. We show how the total variance of Y can be calculated either directly or through a dissimilarity matrix obtained using any dissimilarity index deemed appropriate for pairwise comparisons of community composition data. We addressed the question of which index to use by coding 16 indices using 14 properties that are necessary for beta assessment, comparability among data sets, sampling issues and ordination. Our comparison analysis classified the coefficients under study into five types, three of which are appropriate for beta diversity assessment. Our approach links the concept of beta diversity with the analysis of community data by commonly used methods like ordination and anova. Total beta can be partitioned into Species Contributions (SCBD: degree of variation of individual species across the study area) and Local Contributions (LCBD: comparative indicators of the ecological uniqueness of the sites) to Beta Diversity. Moreover, total beta can be broken up into within- and among-group components by manova, into orthogonal axes by ordination, into spatial scales by eigenfunction analysis or among explanatory data sets by variation partitioning. © 2013 John Wiley & Sons Ltd/CNRS.
[16] Levin SA 1992 ).The problem of pattern and scale in ecology: the Robert H. MacArthur award lectureEcology ,73, 1943 -1967 . DOI:
10.2307/1941447 URL [本文引用: 1] [17] Li H Wu SK 1985 ).The floristic structure of Qinghai-Xizang Plateau subkingdom in XizangScientia Geographica Sinica ,5(1 ), 20 -29 . [本文引用: 1] [李恒 ,武素功 (1985 ).青藏高原植物亚区(西藏部分)的区系结构地理科学 ,5(1 ), 20 -29 .] [本文引用: 1] [18] Lindtner P Svitok M Ujházy K Kubovčík V 2020 ).Disturbances by the European ground squirrel enhance diversity and spatial heterogeneity of plant communities in temperate grasslandBiodiversity and Conservation ,29, 853 -867 . DOI:
10.1007/s10531-019-01914-5 [本文引用: 2] Fossorial rodents are recognized as diversity drivers in grassland ecosystems and are therefore considered ecosystem engineers and keystone species. However, there is a lack of evidence regarding this function for species in temperate grasslands, especially one of the most threatened, the European ground squirrel. We examined the effect of the European ground squirrel on plant species composition and diversity along the disturbance gradient mediated by their different population density and subsequently different disturbance intensity. We evaluated the effect of ground squirrel disturbance on plant diversity patterns in two plant communities with different species richness to determine whether the same pattern exists in contrasting habitats. In each plant community, we established transect of 25 x 250 m composed of 10 quadrats with different disturbance intensities of the ground squirrels. Vascular plant species were recorded in 320 plots of 1 m(2). The distribution of individual species along the ground squirrel-mediated disturbance gradient was analysed using redundancy analysis. Diversity measures were calculated and modelled as a function of disturbances using generalized additive models. We observed significant compositional changes in plant communities accompanied by a reduction in dominant graminoid cover and a non-decreasing trend in forb cover along the disturbance gradient in both types of communities. We found that increasing disturbance activity leads to an increase in diversity at coarse spatial scale (625 m(2)) and spatial heterogeneity in species composition of both species-poor and species-rich plant community. The fine-scale (1 m(2)) diversity increased significantly only in species-poor community. Our results demonstrate that the European ground squirrel can be deservedly labelled as an important ecosystem engineer and keystone species promoting the diversity and heterogeneity of European temperate grasslands.
[19] Liu W Zhang Y Wang X Zhao JZ Xu QM Zhou L 2008 ).Food selection by plateau pikas in different habitats during plant growing seasonActa Theriologica Sinica ,28, 358 -366 . [本文引用: 2] [刘伟 ,张毓 ,王溪 ,赵建中 ,许庆民 ,周立 (2008 ).植物生长季节不同栖息地高原鼠兔的食物选择兽类学报 ,28, 358 -366 .] [本文引用: 2] [20] Liu XQ Zhang X Zhang LF Li YN Zhao L Xu SX Li HQ Ma RR Niu B Gao YB Gu S 2016 ).Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet PlateauActa Ecologica Sinica ,36, 5150 -5162 . [本文引用: 1] [刘晓琴 ,张翔 ,张立锋 ,李英年 ,赵亮 ,徐世晓 ,李红琴 ,马荣荣 ,牛犇 ,高玉葆 ,古松 (2016 ).封育年限对高寒草甸群落组分和物种多样性的影响生态学报 ,36, 5150 -5162 .] [本文引用: 1] [21] Liu YS Fan JW Harris W Shao QQ Zhou YC Wang N Li YZ 2013 ).Effects of plateau pika (Ochotona curzoniae ) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, ChinaPlant and Soil 491 -504 . DOI:
10.1007/s11104-012-1442-x URL [本文引用: 2] [22] Luo YY Meng QT Zhang JH Zhao XY Qin Y 2014 ).Species diversity and biomass in relation to soil properties of alpine meadows in the eastern Tibetan Plateau in different degradation stagesJournal of Glaciology and Geocryology ,36, 1298 -1305 . [本文引用: 1] [罗亚勇 ,孟庆涛 ,张静辉 ,赵学勇 ,秦彧 (2014 ).青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系冰川冻土 ,36, 1298 -1305 .] [本文引用: 1] [23] Ma KP Liu CR Liu YM 1995 ).Biological community diversity measure II: beta diversity measure methodChinese Biodiversity ,3, 38 -43 . [本文引用: 1] [马克平 ,刘灿然 ,刘玉明 (1995 ).生物群落多样性的测度方法II: β多样性的测度方法生物多样性 ,3, 38 -43 .] [本文引用: 1] [24] Maloufi S Catherine A Mouillot D Louvard C Couté A Bernard C Troussellier M 2016 ).Environmental heterogeneity among lakes promotes hyper β-diversity across phytoplankton communitiesFreshwater Biology ,61, 633 -645 . DOI:
10.1111/fwb.12731 URL [本文引用: 1] [25] Niu YF 1999 ).Research on the ecological environment of the Qinghai-Tibet PlateauProgress in Geography ,18, 164 -171 . [本文引用: 1] [牛亚菲 (1999 ).青藏高原生态环境问题研究地理科学进展 ,18, 164 -171 .] [本文引用: 1] [26] Pang XP Guo ZG 2017 ).Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau, ChinaThe Rangeland Journal ,39, 133 -144 . DOI:
10.1071/RJ16093 URL [本文引用: 1] [27] Pang XP Guo ZG 2018 ).Effects of plateau pika disturbance levels on the plant diversity and biomass of an alpine meadowGrassland Science ,64, 159 -166 . DOI:
10.1111/grs.2018.64.issue-3 URL [本文引用: 2] [28] Pang XP Jia TT Li QQ Luo MW Xiao Y Zhao X Guo ZG 2015 a).Effect of available burrow densities of plateau pika (Ochotona curzoniae ) on characteristics and distribution pattern of Kobresia pygmaea communityActa Ecologica Sinica 873 -884 . [本文引用: 2] [庞晓攀 ,贾婷婷 ,李倩倩 ,雒眀伟 ,肖玉 ,赵旭 ,郭正刚 (2015 a).高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响生态学报 ,35, 873 -884 .] [本文引用: 2] [29] Pang XP Wang Q Jia TT Li QQ Guo ZG 2015 b).Effect of burrow entrance densities of plateau pika (Ochotona curzoniae ) on interspecific association in Kobresia pygmaea meadowActa Prataculturae Sinica 224 -230 . [本文引用: 1] [庞晓攀 ,王倩 ,贾婷婷 ,李倩倩 ,郭正刚 (2015 b).高原鼠兔有效洞口数密度对高山嵩草草甸植物种间联结性的影响草业学报 ,24, 224 -230 .] [本文引用: 1] [30] Pang XP Wang Q Zhang J Xu HP Zhang WN Wang J Guo ZG 2020 ).Responses of soil inorganic and organic carbon stocks of alpine meadows to the disturbance by plateau pikasEuropean Journal of Soil Science ,71, 706 -715 . DOI:
10.1111/ejss.v71.4 URL [本文引用: 1] [31] Qiao XJ Li QX Jiang QH Lu JM Franklin S Tang ZY Wang QG Zhang JX Lu ZJ Bao DC Guo YL Liu HB Xu YZ Jiang MX 2015 ).Beta diversity determinants in Badagongshan, a subtropical forest in central ChinaScientific Reports ,5, 17043 . DOI:10.1038/srep17043 . DOI:
10.1038/srep17043 URL [本文引用: 2] [32] Seabloom EW Bjørnstad ON Bolker BM Reichman OJ 2005 ).Spatial signature of environmental heterogeneity, dispersal, and competition in successional grasslandsEcological Monographs ,75, 199 -214 . DOI:
10.1890/03-0841 URL [本文引用: 1] [33] Smith AT Badingqiuying Wilson MC Hogan BW 2019 ).Functional-trait ecology of the plateau pika Ochotona curzoniae in the Qinghai-Tibetan Plateau ecosystemIntegrative Zoology, 87 -103 . DOI:
10.1111/inz2.2019.14.issue-1 URL [本文引用: 1] [34] Suonan LM 2015 ).Main types of natural and degradation situation in Gangcha CountyQinghai Prataculture ,24(3 ), 53 -56 . [本文引用: 1] [索南拉毛 (2015 ).刚察县天然草地主要类型及退化状况青海草业 ,24(3 ), 53 -56 .] [本文引用: 1] [35] Szabó B Lengyel E Padisák J Stenger-Kovács C 2019 ).Benthic diatom metacommunity across small freshwater lakes: driving mechanisms, β-diversity and ecological uniquenessHydrobiologia ,828, 183 -198 . DOI:
10.1007/s10750-018-3811-9 URL [本文引用: 2] [36] Tan LZ Fan CY Zhang CY Zhao XH 2019 ).Understanding and protecting forest biodiversity in relation to species and local contributions to beta diversityEuropean Journal of Forest Research ,138, 1005 -1013 . DOI:
10.1007/s10342-019-01220-3 URL [本文引用: 2] [37] Wang J Legendre P Soininen J Yeh CF Graham E Stegen J Casamayor EO Zhou JZ Shen J Pan FY 2020 ).Temperature drives local contributions to beta diversity in mountain streams: stochastic and deterministic processesGlobal Ecology and Biogeography ,29, 420 -432 . DOI:
10.1111/geb.v29.3 URL [本文引用: 1] [38] Wang KX Shen Y Li Y Ma HB 2019 ).Effects of grazing systems on the diversity and stability of plant communities in desert steppeHeilongjiang Animal Science and Veterinary Medicine ,8, 103 -108 . [本文引用: 1] [王科鑫 ,沈艳 ,李阳 ,马红彬 (2019 ).放牧制度对荒漠草原植物群落多样性及稳定性的影响黑龙江畜牧兽医 ,8, 103 -108 .] [本文引用: 1] [39] Wen L Dong SK Zhu L Shi JJ Liu DM Wang YL Ma YS 2011 ).The effect of natural factors and disturbance intensity on spacial heterogeneity of plant diversity in alpine meadowActa Ecologica Sinica ,31, 1844 -1854 . [本文引用: 1] [温璐 ,董世魁 ,朱磊 ,施建军 ,刘德梅 ,王彦龙 ,马玉寿 (2011 ).环境因子和干扰强度对高寒草甸植物多样性空间分异的影响生态学报 ,31, 1844 -1854 .] [本文引用: 1] [40] Whittaker RH 1960 ).Vegetation of the Siskiyou Mountains, Oregon and CaliforniaEcological Monographs ,30, 279 -338 . DOI:
10.2307/1943563 URL [本文引用: 1] [41] Xu HP Yu C Shu CC Jin SH Pang XP Guo ZG 2019 ).The effect of plateau pika disturbance on plant community diversity and stability in an alpine meadowActa Prataculturae Sinica ,28, 90 -99 . [本文引用: 1] [徐海鹏 ,于成 ,舒朝成 ,金少红 ,庞晓攀 ,郭正刚 (2019 ).高原鼠兔干扰对高寒草甸植物群落多样性和稳定性的影响草业学报 ,28, 90 -99 .] [本文引用: 1] [42] Yang J Chu PF Chen DM Wang MJ Bai YF 2014 ).Mechanisms underlying the impacts of grazing on plant α, β and γ diversity in a typical steppe of the Inner Mongolia grasslandChinese Journal of Plant Ecology ,38, 188 -200 . DOI:
10.3724/SP.J.1258.2014.00017 URL [本文引用: 1] [杨婧 ,褚鹏飞 ,陈迪马 ,王明玖 ,白永飞 (2014 ).放牧对内蒙古典型草原α、β和γ多样性的影响机制植物生态学报 ,38, 188 -200 .] [本文引用: 1] [43] Yang LM Li JD Yang YF 1999 ).β-diversity of grassland communities along gradient of grazing disturbanceChinese Journal of Applied Ecology ,10, 442 -446 . [本文引用: 1] [杨利民 ,李建东 ,杨允菲 (1999 ).草地群落放牧干扰梯度β多样性研究应用生态学报 ,10, 442 -446 .] [本文引用: 1] [44] Ye GH Chu B Zhou R Zhang FY Hua XZ Hua R Wang T Hua LM 2019 ).Impact of plateau pika (Ochotona curzoniae ) disturbance on larger soil animal fauna in alpine meadowJournal of Gansu Agricultural University 150 -158 . [本文引用: 1] [叶国辉 ,楚彬 ,周睿 ,张飞宇 ,华铣泽 ,花蕊 ,王婷 ,花立民 (2019 ).高原鼠兔干扰对高寒草甸大型土壤动物类群的影响甘肃农业大学学报 ,54, 150 -158 .] [本文引用: 1] [45] Zhang WN Wang Q Zhang J Pang XP Xu HP Wang J Guo ZG 2020 ).Clipping by plateau pikas and impacts to plant communityRangeland Ecology & Management ,73, 368 -374 . DOI:
10.1016/j.rama.2020.01.010 URL [本文引用: 1] [46] Zhou XR Guo ZG Guo XH 2010 ).The role of plateau pika and plateau zokor in alpine meadowPratacultural Science ,27(5 ), 38 -44 . [本文引用: 2] [周雪荣 ,郭正刚 ,郭兴华 (2010 ).高原鼠兔和高原鼢鼠在高寒草甸中的作用草业科学 ,27(5 ), 38 -44 .] [本文引用: 2] Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist
2
2011
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
... 调查样区在刚察县哈尔盖镇的冷季放牧高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 主要伴生植物为莎草科的矮生嵩草(
Kobresia humilis )、禾本科的高原早熟禾(
Poa alpigena )和紫羊茅(
Festuca rubra )、菊科的美丽风毛菊(
Saussurea pulchra )和乳白香青(
Anaphalis lactea )、蔷薇科的莓叶委陵菜(
Potentilla fragarioides )等, 草层低矮、植株密聚.冷季放牧高寒草甸采用全年围封, 暖季(牧草生长季)不放牧, 冷季(牧草休眠期)打开围栏放牧, 因此从高寒草甸返青到植物枯黄期, 均无放牧动物干扰.野外调查时间为2019年7月底牧草生长旺盛时期, 该时期植物生物量最大, 高原鼠兔干扰也最为严重(
周雪荣等, 2010 ).首先, 选择地势相对平坦一致、草地类型一致的高寒草甸区, 确定是否有高原鼠兔干扰存在.高原鼠兔干扰的标准为高寒草甸内目视有高原鼠兔出没和有效洞口存在, 若存在高原鼠兔干扰, 则设置样地, 样地面积大小为35 m × 35 m, 接近高原鼠兔的巢域尺度(
Pang et al ., 2020 ).选定第1个样地后, 驱车继续前行距离超过5 km后, 选定第2样地.连续选定10个有高原鼠兔干扰的样地后, 在距离第10个样地不到5 km的范围内, 设置一个没有高原鼠兔干扰的样地, 其面积、地形地貌和植被类型均与所有干扰样地一致.高原鼠兔营家族式生活, 分布具有领域性, 因此高原鼠兔整体分布状态呈镶嵌分布, 因此, 有些高寒草甸虽然与干扰样地地貌、地形和植被类型一致, 调查时依然没有高原鼠兔干扰存在, 由于高原鼠兔种群扩散是一个逐渐的过程, 因此没有高原鼠兔干扰的样地仅是高原鼠兔潜在的适宜分布区, 因此设置非干扰样地(0干扰强度)是能够实现的.然后, 采用3天堵洞法测定每个干扰样地的有效洞口数.由于沿着公路布设样地, 而公路的走向并非直线, 这样客观上形成了一个样地群.以干扰样地内有效洞口密度测度该样地的干扰强度, 且将其视为一个位点, 其空间位置和干扰程度均存在差异.因未干扰样地没有高原鼠兔存在, 标记为T
0 , 然后根据每个干扰样地内的有效洞口密度, 由低到高标记为T
1 、T
2 、T
3 、T
4 、T
5 、T
6 、T
7 、T
8 、T
9 、T
10 , 共计11个位点(
图1 )(
Anderson et al ., 2011 ). ...
Spatial structure and soil properties shape local community structure of plant-parasitic nematodes in cultivated olive trees in southern Spain
1
2020
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of Trans-Himalaya
3
2006
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size
1
2006
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
Conservation of prairie dog ecosystem engineering may support arthropod beta and gamma diversity
1
2006
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Disentangling the correlates of species and site contributions to beta diversity in dung beetle assemblages
1
2018
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Abundance, spatial variance and occupancy: arthropod species distribution in the Azores
1
2006
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
Effects of animal disturbance on tallgrass prairie vegetation
1
1989
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
Response of alpine meadow communities to burrow density changes of plateau pika (
Ochotona curzoniae ) in the Qinghai-Tibet Plateau
1
2012
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
Mole rats act as ecosystem engineers within a biodiversity hotspot, the Cape Fynbos
1
2013
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响
1
2014
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响
1
2014
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
Plant succession on gopher mounds in western Cascade meadows: consequences for species diversity and heterogeneity
1
2008
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
β-多样性的研究: 应用多元回归和典范分析研究生态方差的分解
1
2007
... 采用植物群落二元丰富度的方差分解法分析单个物种和物种组合(位点)对调查区域内β多样性的贡献(
Legendre, 2007 ;
Legendre & de Cáceres, 2013 ).即将区域内不同物种在每个位点的分布数据表, 视为一系列环境变量(干扰强度)和空间变量(样地间距离)的函数, 从而将β多样性分解为单个位点(特定干扰强度下的物种组合)对β多样性的贡献(LCBD)和单个物种对β多样性的贡献(SCBD), 进而确定单个位点和单一物种对整个高原鼠兔干扰区域内β多样性的影响. ...
β-多样性的研究: 应用多元回归和典范分析研究生态方差的分解
1
2007
... 采用植物群落二元丰富度的方差分解法分析单个物种和物种组合(位点)对调查区域内β多样性的贡献(
Legendre, 2007 ;
Legendre & de Cáceres, 2013 ).即将区域内不同物种在每个位点的分布数据表, 视为一系列环境变量(干扰强度)和空间变量(样地间距离)的函数, 从而将β多样性分解为单个位点(特定干扰强度下的物种组合)对β多样性的贡献(LCBD)和单个物种对β多样性的贡献(SCBD), 进而确定单个位点和单一物种对整个高原鼠兔干扰区域内β多样性的影响. ...
Analyzing beta diversity: partitioning the spatial variation of community composition data
1
2005
... 所有干扰位点植物群落的二元丰富度构成的矩 阵
VAR (
Y )具有
n 个干扰位点(行)和
p 个物种(列), 每个值用
yij 表示, ¯
yj 表示第
j 个物种二元丰富度的平均值, 在每个干扰位点, 某物种出现用“1”表示, 未出现则用“0”表示(
Legendre et al ., 2005 ).计算公式为: ...
Beta diversity as the variance of community data: dissimilarity coefficients and partitioning
4
2013
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
... 采用植物群落二元丰富度的方差分解法分析单个物种和物种组合(位点)对调查区域内β多样性的贡献(
Legendre, 2007 ;
Legendre & de Cáceres, 2013 ).即将区域内不同物种在每个位点的分布数据表, 视为一系列环境变量(干扰强度)和空间变量(样地间距离)的函数, 从而将β多样性分解为单个位点(特定干扰强度下的物种组合)对β多样性的贡献(LCBD)和单个物种对β多样性的贡献(SCBD), 进而确定单个位点和单一物种对整个高原鼠兔干扰区域内β多样性的影响. ...
... 自
Legendre和de Cáceres (2013) 提出通过群落二元丰富度的方差分解来估计个别物种和物种组合对β多样性贡献的研究方法后, β多样性的研究逐渐从群落间的物种周转率向物种和物种组合对一定空间范围内β多样性的相对重要性转变, 从而根据β多样性的变化原因, 提出更具有针对性的生物多样性保护策略(
Qiao et al ., 2015 ;
Wang et al ., 2020 ).本研究在分析高原鼠兔干扰下群落间β多样性变化的基础上, 采用群落二元丰富度数据的方差分解法分析了高原鼠兔干扰区域内, 单个物种和单个位点(物种组合)对植物群落β多样性的相对重要性, 以期确定单个物种、单一位点, 或两者结合对植物群落β多样性保护的意义. ...
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture
1
1992
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
青藏高原植物亚区(西藏部分)的区系结构
1
1985
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
青藏高原植物亚区(西藏部分)的区系结构
1
1985
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
Disturbances by the European ground squirrel enhance diversity and spatial heterogeneity of plant communities in temperate grassland
2
2020
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
植物生长季节不同栖息地高原鼠兔的食物选择
2
2008
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
植物生长季节不同栖息地高原鼠兔的食物选择
2
2008
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
封育年限对高寒草甸群落组分和物种多样性的影响
1
2016
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
封育年限对高寒草甸群落组分和物种多样性的影响
1
2016
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
Effects of plateau pika (
Ochotona curzoniae ) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, China
2
2013
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系
1
2014
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系
1
2014
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
生物群落多样性的测度方法II: β多样性的测度方法
1
1995
... 植物群落间β多样性的测度指标有多种, 本研究采用常用的Whittaker指数测定β多样性(
马克平等, 1995 ), 其计算公式为${\beta _{\rm{W}}} = (S{\rm{/}}X) - 1$, 式中:
S 为干扰样地内的累计出现物种数,
X 为样地内物种丰富度, 即各样方物种数的平均值. ...
生物群落多样性的测度方法II: β多样性的测度方法
1
1995
... 植物群落间β多样性的测度指标有多种, 本研究采用常用的Whittaker指数测定β多样性(
马克平等, 1995 ), 其计算公式为${\beta _{\rm{W}}} = (S{\rm{/}}X) - 1$, 式中:
S 为干扰样地内的累计出现物种数,
X 为样地内物种丰富度, 即各样方物种数的平均值. ...
Environmental heterogeneity among lakes promotes hyper β-diversity across phytoplankton communities
1
2016
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
青藏高原生态环境问题研究
1
1999
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
青藏高原生态环境问题研究
1
1999
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau, China
1
2017
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
Effects of plateau pika disturbance levels on the plant diversity and biomass of an alpine meadow
2
2018
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响
2
2015
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
... ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响
2
2015
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
... ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
高原鼠兔有效洞口数密度对高山嵩草草甸植物种间联结性的影响
1
2015
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
高原鼠兔有效洞口数密度对高山嵩草草甸植物种间联结性的影响
1
2015
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Responses of soil inorganic and organic carbon stocks of alpine meadows to the disturbance by plateau pikas
1
2020
... 调查样区在刚察县哈尔盖镇的冷季放牧高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 主要伴生植物为莎草科的矮生嵩草(
Kobresia humilis )、禾本科的高原早熟禾(
Poa alpigena )和紫羊茅(
Festuca rubra )、菊科的美丽风毛菊(
Saussurea pulchra )和乳白香青(
Anaphalis lactea )、蔷薇科的莓叶委陵菜(
Potentilla fragarioides )等, 草层低矮、植株密聚.冷季放牧高寒草甸采用全年围封, 暖季(牧草生长季)不放牧, 冷季(牧草休眠期)打开围栏放牧, 因此从高寒草甸返青到植物枯黄期, 均无放牧动物干扰.野外调查时间为2019年7月底牧草生长旺盛时期, 该时期植物生物量最大, 高原鼠兔干扰也最为严重(
周雪荣等, 2010 ).首先, 选择地势相对平坦一致、草地类型一致的高寒草甸区, 确定是否有高原鼠兔干扰存在.高原鼠兔干扰的标准为高寒草甸内目视有高原鼠兔出没和有效洞口存在, 若存在高原鼠兔干扰, 则设置样地, 样地面积大小为35 m × 35 m, 接近高原鼠兔的巢域尺度(
Pang et al ., 2020 ).选定第1个样地后, 驱车继续前行距离超过5 km后, 选定第2样地.连续选定10个有高原鼠兔干扰的样地后, 在距离第10个样地不到5 km的范围内, 设置一个没有高原鼠兔干扰的样地, 其面积、地形地貌和植被类型均与所有干扰样地一致.高原鼠兔营家族式生活, 分布具有领域性, 因此高原鼠兔整体分布状态呈镶嵌分布, 因此, 有些高寒草甸虽然与干扰样地地貌、地形和植被类型一致, 调查时依然没有高原鼠兔干扰存在, 由于高原鼠兔种群扩散是一个逐渐的过程, 因此没有高原鼠兔干扰的样地仅是高原鼠兔潜在的适宜分布区, 因此设置非干扰样地(0干扰强度)是能够实现的.然后, 采用3天堵洞法测定每个干扰样地的有效洞口数.由于沿着公路布设样地, 而公路的走向并非直线, 这样客观上形成了一个样地群.以干扰样地内有效洞口密度测度该样地的干扰强度, 且将其视为一个位点, 其空间位置和干扰程度均存在差异.因未干扰样地没有高原鼠兔存在, 标记为T
0 , 然后根据每个干扰样地内的有效洞口密度, 由低到高标记为T
1 、T
2 、T
3 、T
4 、T
5 、T
6 、T
7 、T
8 、T
9 、T
10 , 共计11个位点(
图1 )(
Anderson et al ., 2011 ). ...
Beta diversity determinants in Badagongshan, a subtropical forest in central China
2
2015
... 自
Legendre和de Cáceres (2013) 提出通过群落二元丰富度的方差分解来估计个别物种和物种组合对β多样性贡献的研究方法后, β多样性的研究逐渐从群落间的物种周转率向物种和物种组合对一定空间范围内β多样性的相对重要性转变, 从而根据β多样性的变化原因, 提出更具有针对性的生物多样性保护策略(
Qiao et al ., 2015 ;
Wang et al ., 2020 ).本研究在分析高原鼠兔干扰下群落间β多样性变化的基础上, 采用群落二元丰富度数据的方差分解法分析了高原鼠兔干扰区域内, 单个物种和单个位点(物种组合)对植物群落β多样性的相对重要性, 以期确定单个物种、单一位点, 或两者结合对植物群落β多样性保护的意义. ...
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Spatial signature of environmental heterogeneity, dispersal, and competition in successional grasslands
1
2005
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
Functional-trait ecology of the plateau pika
Ochotona curzoniae in the Qinghai-Tibetan Plateau ecosystem
1
2019
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
刚察县天然草地主要类型及退化状况
1
2015
... 研究地区位于青藏高原青海湖北岸, 行政区划隶属于青海省刚察县(99.33°-100.62° E, 36.97°- 38.07° N), 海拔3 300-3 800 m.气候属高原大陆性气候, 年降水量370 mm, 年蒸发量1 500-1 847 mm.1月平均气温-17.5 ℃, 7月平均气温11 ℃, 年平均气温-0.6 ℃左右.年日照时间为3 037 h, 年内霜期大于270天, 全年无绝对无霜期.土壤为亚高山草甸土.天然草地面积为71.2万hm
2 , 占全县总土地面积的73.8% (
索南拉毛, 2015 ).草地类型以高寒草甸为主, 优势种以莎草科的植物为主.高寒草甸虽然是优良放牧地, 但因其草层低矮而成为高原鼠兔的适宜分布区. ...
刚察县天然草地主要类型及退化状况
1
2015
... 研究地区位于青藏高原青海湖北岸, 行政区划隶属于青海省刚察县(99.33°-100.62° E, 36.97°- 38.07° N), 海拔3 300-3 800 m.气候属高原大陆性气候, 年降水量370 mm, 年蒸发量1 500-1 847 mm.1月平均气温-17.5 ℃, 7月平均气温11 ℃, 年平均气温-0.6 ℃左右.年日照时间为3 037 h, 年内霜期大于270天, 全年无绝对无霜期.土壤为亚高山草甸土.天然草地面积为71.2万hm
2 , 占全县总土地面积的73.8% (
索南拉毛, 2015 ).草地类型以高寒草甸为主, 优势种以莎草科的植物为主.高寒草甸虽然是优良放牧地, 但因其草层低矮而成为高原鼠兔的适宜分布区. ...
Benthic diatom metacommunity across small freshwater lakes: driving mechanisms, β-diversity and ecological uniqueness
2
2019
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
... 整个调查区域内不同物种对高寒草甸β多样性的相对重要性存在差异, 其中冰草、臭蒿、小花草玉梅、獐牙菜、婆婆纳的SCBD值相对较大, 说明其对区域内β多样性的重要性贡献相对突出, 主要是这些植物大多数为杂类草, 是r策略者, 生态位宽度较小, 易受时间和空间尺度上资源的限制, 当高原鼠兔干扰后, 这些植物在群落内的受抑制程度明显减弱, 植物种群的空间分布特征从聚集分布转变为随机分布(
庞晓攀等, 2015a ), 生态位宽度增加, 说明高原鼠兔干扰为臭蒿、小花草玉梅、獐牙菜、婆婆纳等创造了良好的生长环境(
贾婷婷等, 2014 ), 从而增加了这些植物在构建植物群落β多样性中的分量, 因此, 在较大的空间尺度上, 高原鼠兔对β多样性的贡献较大.而高山嵩草、高山豆、黄花棘豆、紫羊茅、高原早熟禾等物种的SCBD值较小, 说明莎草科、豆科和禾本科植物对β多样性的贡献普遍较低, 主要原因是这些物种生态宽度较大, 分布范围跨越多个高原鼠兔干扰位点, 因此, 它们虽然在维持植物群落的特征时具有关键作用, 但对植物群落β多样性的贡献相对较小(
Archidona-Yuste et al ., 2020 ).同时, 高原鼠兔喜食植物随着干扰梯度存在一定的分异, 当高原鼠兔食物资源丰富时, 其喜食单子叶植物, 而当高原鼠兔干扰强度很大时, 其喜食植物的资源数量降低, 此时高原鼠兔亦喜食双子叶植物, 譬如杂类草矮火绒草(
Leontopodium nanum )等(
刘伟等, 2008 ), 因此, 高原鼠兔不同干扰程度下, 其食物资源的变化也可能是导致不同物种对β多样性贡献存在差异的原因.本研究还发现, 整个调查区内, 一个物种占据的干扰位点数过多或过少, 其对植物群落β多样性的贡献相对较小, 而一个物种占据的高原鼠兔干扰位点数处于中等水平时, 其对区域β多样性贡献较大, 说明物种的SCBD值与该物种的位点占有率显著相关, 可能的原因是占据中等位点数的物种对高原鼠兔干扰强度具有不同的环境偏好, 这导致物种在不同干扰样地间的占有率差异较大(
Gaston et al ., 2006 ), 因此具有较高的SCBD值, 这与地点对喀尔巴阡盆地淡水湖硅藻群落β多样性贡献的研究结果(
Szabó et al ., 2019 )类似, 说明小型草食动物干扰会改变草地植物群落的生境, 群落内不同植物因适应改变后的生境而进行自我调整(
Bagchi et al ., 2006 ), 植物群落自我调整的过程中, 不同物种的适应性存在明显分异(
庞晓攀等, 2015a ), 从而会改变某一单一植物对高原鼠兔干扰区域β多样性的贡献, 客观上导致不同物种对植物群落β多样性贡献存在分异. ...
Understanding and protecting forest biodiversity in relation to species and local contributions to beta diversity
2
2019
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Temperature drives local contributions to beta diversity in mountain streams: stochastic and deterministic processes
1
2020
... 自
Legendre和de Cáceres (2013) 提出通过群落二元丰富度的方差分解来估计个别物种和物种组合对β多样性贡献的研究方法后, β多样性的研究逐渐从群落间的物种周转率向物种和物种组合对一定空间范围内β多样性的相对重要性转变, 从而根据β多样性的变化原因, 提出更具有针对性的生物多样性保护策略(
Qiao et al ., 2015 ;
Wang et al ., 2020 ).本研究在分析高原鼠兔干扰下群落间β多样性变化的基础上, 采用群落二元丰富度数据的方差分解法分析了高原鼠兔干扰区域内, 单个物种和单个位点(物种组合)对植物群落β多样性的相对重要性, 以期确定单个物种、单一位点, 或两者结合对植物群落β多样性保护的意义. ...
放牧制度对荒漠草原植物群落多样性及稳定性的影响
1
2019
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
放牧制度对荒漠草原植物群落多样性及稳定性的影响
1
2019
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
环境因子和干扰强度对高寒草甸植物多样性空间分异的影响
1
2011
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
环境因子和干扰强度对高寒草甸植物多样性空间分异的影响
1
2011
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
Vegetation of the Siskiyou Mountains, Oregon and California
1
1960
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
高原鼠兔干扰对高寒草甸植物群落多样性和稳定性的影响
1
2019
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
高原鼠兔干扰对高寒草甸植物群落多样性和稳定性的影响
1
2019
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
放牧对内蒙古典型草原α、β和γ多样性的影响机制
1
2014
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
放牧对内蒙古典型草原α、β和γ多样性的影响机制
1
2014
... 草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子, 当其干扰强度变化时, 必然会在草地生境内形成不同的环境梯度(
杨婧等, 2014 ).已有研究表明, 大型草食动物围栏区草地的β多样性指数显著高于其他放牧状态下草地的β多样性(
王科鑫等, 2019 ).除大型草食动物之外, 小型草食动物也是草地生态系统不可缺少的组分, 其往往通过多种途径干扰草地(
Bagchi et al ., 2006 ).小型草食动物扰动在空间和时间上往往是分散的, 易造成植被状态的“时空异质性”, 从而在较大的空间尺度上影响群落结构(
Levin, 1992 ).已有研究表明, 欧洲地松鼠(
Spermophilus citellus )干扰会增加物种贫乏的温带草原植物群落的Whittaker指数, 从而影响β多样性(
Lindtner et al ., 2020 ), 这意味着小型草食动物可以通过改变较小尺度的生境而影响大尺度植物群落的β多样性(
Gibson, 1989 ;
Seabloom et al ., 2005 ), 然而单个物种或物种组合是如何影响小型草食动物干扰与草地植物群落β多样性的关系, 尚需科学研究提供证据. ...
草地群落放牧干扰梯度β多样性研究
1
1999
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
草地群落放牧干扰梯度β多样性研究
1
1999
... 青藏高原高寒草甸是维持泛北极植物区生物多样性和畜牧业生产的基础(
李恒和武素功, 1985 ), 然而严酷脆弱的自然环境和较短的生长周期, 致使植物多样性对外界干扰的响应十分敏感(
牛亚菲, 1999 ).植物多样性对外界干扰的响应结果, 往往引起植物群落结构和功能的改变, 进而能够揭示区域内高寒草甸的生态过程(
罗亚勇等, 2014 ).β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率(
Whittaker, 1960 ), 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映植物群落的抗干扰能力(
杨利民等, 1999 ), 从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程(
Anderson et al ., 2011 ).大多数植物群落的β多样性研究往往采用Whittaker指数反映植物群落间β多样性的变化(
温璐等, 2011 ;
刘晓琴等, 2016 ), 虽然能够说明物种在不同环境梯度之间的替代程度, 但无法确定单一物种和物种组合(位点)对较大空间尺度范围内群落β多样性的贡献.
Legendre和de Cáceres (2013) 提出了基于群落二元丰富度的方差分解来测度单个物种或物种组合对区域内β多样性贡献的相对重要性,
Tan等(2019) 采用此方法分析了单个物种和物种组合对森林群落β多样性的贡献, 提出保护贡献度高的物种更有利于维持森林植物群落的多样性.
Szabó等(2019) 分析了单个硅藻物种和不同地点硅藻组合对淡水湖硅藻群落β多样性的贡献, 提出保护贡献度高的物种, 改善贡献度低的物种组合, 可实现区域内生物多样性的维持.然而通过二元群落丰富度数据的方差分解分析草地植物群落的β多样性变化, 并没有引起足够的关注. ...
高原鼠兔干扰对高寒草甸大型土壤动物类群的影响
1
2019
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
高原鼠兔干扰对高寒草甸大型土壤动物类群的影响
1
2019
... 干扰位点对总体β多样性的贡献, 可以确定生态独特性高的位点, 以及物种丰富度较高或较低的位点, 从而判断具有高保存价值的位点(
Legendre & de Cáceres, 2013 ).本研究结果表明, 不同位点对区域β多样性的相对重要性亦存在差异, 说明小型草食动物不同扰动强度下的植物群落在高寒草甸发育过程中的零散性分布特征在空间尺度上对β多样性有积极的影响(
Jones et al ., 2008 ).其中T
0 、T
1 、T
10 位点对β多样性的贡献最大.T
0 为没有干扰的位点, 其植物群落物种组成经过长期的演化, 形成相对稳定的种间关系(
庞晓攀等, 2015b ), 而干扰位点内常见和稀有种的共存提供了多样化生境而存在植物群落组分重组现象(
Hagenah & Bennett, 2013 ), 因此, T
0 的植物群落在调查区域内最独特.T
1 位点干扰强度最小, 植物群落重组现象相对较小, 基本保持原有植物群落的水平, 其独特性虽然小于T
0 位点, 但其依然保持较高的独特性.T
10 是研究区内高原鼠兔干扰强度最大的位点, 其有效洞口数密度高达1 633个·hm
-2 , 此时生境旱化严重, 适宜生物的物种相对减少, 基本以中生和旱生植物为主, 又形成了相对稳定的植物群落, 其显示出较强的独特性.位点T
5 对区域β多样性贡献最小, 主要原因是该位点处于中度干扰强度, 植物群落具有较高的物种丰富度(
Bagchi et al ., 2006 ), 这导致该位点物种组成与其他位点相比很普遍, 独特性较低, 因此其对β多样性的变化贡献相对较小.在高原鼠兔不同干扰强度下, 土壤动物丰富性存在明显变化, 其中中度干扰强度时会特别增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫(
叶国辉等, 2019 ), 这说明在高原鼠兔不同干扰强度下生态系统食物网的复杂性会有明显变化, 而土壤食物网的变化亦是小型啮齿草食动物干扰调节生态系统的重要内在原因, 从而使得不同干扰位点LCBD值存在差异.因此, 从较大的空间尺度而言, 高原鼠兔栖息地的独特性, 以及高原鼠兔干扰与其他土壤动物的关系, 成为特定干扰位点β多样性变化的驱动因素之一(
Bangert & Slobodchikoff, 2006 ).研究发现LCBD值与各位点物种丰富度呈显著负相关关系, 这与森林植物群落β多样性的研究结果(
Tan et al ., 2019 )一致, 说明LCBD值高的位点, 其物种组成更为独特, 但物种丰富度较低, 主要原因是物种丰富度高的位点与其他位点共享物种的机会更大, 导致在整个区域内的独特性降低(
Maloufi et al ., 2016 ).本研究与
Qiao等(2015) 在森林群落中对稀有种丰富度与LCBD的关系的研究相异, 这表明常见种和稀有种的比例可能决定着LCBD与丰富度的关系(
da Silva et al ., 2018 ).尽管如此, LCBD值的大小, 可以反映高原鼠兔干扰与物种丰富度降低或增加的关系.因此, 从生物多样性保护角度, 应该保护LCBD值较高的T
0 、T
1 、T
10 位点.而位点的LCBD值与有效洞口密度没有明显的关联性, 说明LCBD可能取决于位点, 即特定栖息地的物种组合. ...
Clipping by plateau pikas and impacts to plant community
1
2020
... 高原鼠兔(
Ochotona curzoniae )是青藏高原常见的、特有的小型草食动物, 主要栖息于青藏高原高寒草甸, 分布广, 数量多(
Guo et al ., 2012 ), 被认为是高寒草甸的“生态系统工程师” (
Smith et al ., 2019 ).高原鼠兔与高寒草甸在长期演化中形成了微妙的关系, 其中高原鼠兔刈割行为能够增加其有效洞口处的植物群落的物种多样性(
Zhang et al ., 2020 ), 适量的高原鼠兔干扰能够提高高寒草甸植物群落的稳定性, 增加禾本科植物的比例, 而高原鼠兔干扰程度过大时则会降低高寒草甸植物群落的丰富度和稳定性, 导致杂类草种类增加(
徐海鹏等, 2019 ), 说明高原鼠兔干扰往往会增加环境的异质性(
Liu et al ., 2013 ), 且为新物种的定居提供更多栖息地(
Bakker et al ., 2006 ).虽然大量研究证实, 高原鼠兔不同干扰强度必然会造成植物群落结构组分的差异, 从而影响高寒草甸的物种多样性(
Pang & Guo, 2018 ), 但高原鼠兔干扰引起的小尺度物种多样性变化, 是否能够引起大尺度范围内植物群落β多样性变化, 是确定高原鼠兔存在时如何保护高寒草甸植物多样性的重要依据.因此, 本研究以高原鼠兔干扰为例, 采用Whittaker指数和群落二元丰富度的方差分解法同步分析高原鼠兔干扰对青藏高原高寒草甸植物群落β多样性的影响, 以期说明高原鼠兔干扰对植物群落物种周转速率的影响, 揭示单个物种和物种组合(干扰位点)对β多样性的贡献, 从而确定高原鼠兔干扰是通过单个物种、还是位点(物种组合)、或两者结合而改变植物群落的β多样性, 也为通过高原鼠兔管理而维持青藏高原高寒草甸植物群落多样性提供科学依据. ...
高原鼠兔和高原鼢鼠在高寒草甸中的作用
2
2010
... 调查样区在刚察县哈尔盖镇的冷季放牧高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 主要伴生植物为莎草科的矮生嵩草(
Kobresia humilis )、禾本科的高原早熟禾(
Poa alpigena )和紫羊茅(
Festuca rubra )、菊科的美丽风毛菊(
Saussurea pulchra )和乳白香青(
Anaphalis lactea )、蔷薇科的莓叶委陵菜(
Potentilla fragarioides )等, 草层低矮、植株密聚.冷季放牧高寒草甸采用全年围封, 暖季(牧草生长季)不放牧, 冷季(牧草休眠期)打开围栏放牧, 因此从高寒草甸返青到植物枯黄期, 均无放牧动物干扰.野外调查时间为2019年7月底牧草生长旺盛时期, 该时期植物生物量最大, 高原鼠兔干扰也最为严重(
周雪荣等, 2010 ).首先, 选择地势相对平坦一致、草地类型一致的高寒草甸区, 确定是否有高原鼠兔干扰存在.高原鼠兔干扰的标准为高寒草甸内目视有高原鼠兔出没和有效洞口存在, 若存在高原鼠兔干扰, 则设置样地, 样地面积大小为35 m × 35 m, 接近高原鼠兔的巢域尺度(
Pang et al ., 2020 ).选定第1个样地后, 驱车继续前行距离超过5 km后, 选定第2样地.连续选定10个有高原鼠兔干扰的样地后, 在距离第10个样地不到5 km的范围内, 设置一个没有高原鼠兔干扰的样地, 其面积、地形地貌和植被类型均与所有干扰样地一致.高原鼠兔营家族式生活, 分布具有领域性, 因此高原鼠兔整体分布状态呈镶嵌分布, 因此, 有些高寒草甸虽然与干扰样地地貌、地形和植被类型一致, 调查时依然没有高原鼠兔干扰存在, 由于高原鼠兔种群扩散是一个逐渐的过程, 因此没有高原鼠兔干扰的样地仅是高原鼠兔潜在的适宜分布区, 因此设置非干扰样地(0干扰强度)是能够实现的.然后, 采用3天堵洞法测定每个干扰样地的有效洞口数.由于沿着公路布设样地, 而公路的走向并非直线, 这样客观上形成了一个样地群.以干扰样地内有效洞口密度测度该样地的干扰强度, 且将其视为一个位点, 其空间位置和干扰程度均存在差异.因未干扰样地没有高原鼠兔存在, 标记为T
0 , 然后根据每个干扰样地内的有效洞口密度, 由低到高标记为T
1 、T
2 、T
3 、T
4 、T
5 、T
6 、T
7 、T
8 、T
9 、T
10 , 共计11个位点(
图1 )(
Anderson et al ., 2011 ). ...
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
高原鼠兔和高原鼢鼠在高寒草甸中的作用
2
2010
... 调查样区在刚察县哈尔盖镇的冷季放牧高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 主要伴生植物为莎草科的矮生嵩草(
Kobresia humilis )、禾本科的高原早熟禾(
Poa alpigena )和紫羊茅(
Festuca rubra )、菊科的美丽风毛菊(
Saussurea pulchra )和乳白香青(
Anaphalis lactea )、蔷薇科的莓叶委陵菜(
Potentilla fragarioides )等, 草层低矮、植株密聚.冷季放牧高寒草甸采用全年围封, 暖季(牧草生长季)不放牧, 冷季(牧草休眠期)打开围栏放牧, 因此从高寒草甸返青到植物枯黄期, 均无放牧动物干扰.野外调查时间为2019年7月底牧草生长旺盛时期, 该时期植物生物量最大, 高原鼠兔干扰也最为严重(
周雪荣等, 2010 ).首先, 选择地势相对平坦一致、草地类型一致的高寒草甸区, 确定是否有高原鼠兔干扰存在.高原鼠兔干扰的标准为高寒草甸内目视有高原鼠兔出没和有效洞口存在, 若存在高原鼠兔干扰, 则设置样地, 样地面积大小为35 m × 35 m, 接近高原鼠兔的巢域尺度(
Pang et al ., 2020 ).选定第1个样地后, 驱车继续前行距离超过5 km后, 选定第2样地.连续选定10个有高原鼠兔干扰的样地后, 在距离第10个样地不到5 km的范围内, 设置一个没有高原鼠兔干扰的样地, 其面积、地形地貌和植被类型均与所有干扰样地一致.高原鼠兔营家族式生活, 分布具有领域性, 因此高原鼠兔整体分布状态呈镶嵌分布, 因此, 有些高寒草甸虽然与干扰样地地貌、地形和植被类型一致, 调查时依然没有高原鼠兔干扰存在, 由于高原鼠兔种群扩散是一个逐渐的过程, 因此没有高原鼠兔干扰的样地仅是高原鼠兔潜在的适宜分布区, 因此设置非干扰样地(0干扰强度)是能够实现的.然后, 采用3天堵洞法测定每个干扰样地的有效洞口数.由于沿着公路布设样地, 而公路的走向并非直线, 这样客观上形成了一个样地群.以干扰样地内有效洞口密度测度该样地的干扰强度, 且将其视为一个位点, 其空间位置和干扰程度均存在差异.因未干扰样地没有高原鼠兔存在, 标记为T
0 , 然后根据每个干扰样地内的有效洞口密度, 由低到高标记为T
1 、T
2 、T
3 、T
4 、T
5 、T
6 、T
7 、T
8 、T
9 、T
10 , 共计11个位点(
图1 )(
Anderson et al ., 2011 ). ...
... 高原鼠兔是青藏高原主要的小型掘洞类草食动物(
周雪荣等, 2010 ), 其不仅通过选择性采食直接影响部分植物生长发育(
刘伟等, 2008 ), 而且通过挖掘、排泄粪尿以及刈割行为产生过剩食物碎屑堆积等作用, 改变植物生境, 影响植物群落多样性(
Pang & Guo, 2018 ).研究结果表明, 高原鼠兔干扰会改变高寒草甸植物群落的β多样性指数, 说明小型草食动物干扰会改变植物群落的物种周转速率, 增加不同栖息地群落的异质性, 这趋同于欧洲地松鼠对温带草原群落β多样性指数的影响(
Lindtner et al ., 2020 ), 说明小型草食动物能够通过改变样方尺度的植物群落组分, 而引起更大尺度范围内的植物群落多样性变化.高原鼠兔中度干扰强度时物种更替速率最大, 主要是高原鼠兔不同干扰程度下不同植物对高原鼠兔干扰响应的整体体现, 当干扰程度从低干扰强度增加到中度干扰强度时, 改变了原始植物群落内的物种关系, 原来的优势植物虽然优势度有一定程度的降低, 但其依然处于相对优势的地位, 但原来处于劣势的植物, 则因植物群落内光照资源的重置和竞争压力的减弱, 而获得更好的生长机会, 更为重要的是高原鼠兔活动形成的裸斑, 增加了高寒草甸内部环境的异质性, 为部分机会主义物种和先锋物种创造了条件(
Pang & Guo, 2017 ), 从而为多物种共存提供了条件, 因此, β多样性指数会逐渐增加.然而, 当高原鼠兔干扰强度从中度水平继续增加时, 高原鼠兔产生的裸斑数量和面积增多, 环境干旱化程度增强(
Liu et al ., 2013 ), 有些湿生物种因不适应干旱生境而逐渐减少, 甚至退出群落, 群落内主要以旱生植物为主, 群落间共有种趋于稳定, 物种周转速率缓慢降低, 从而逐渐降低β多样性指数. ...
 ,*兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
,*兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020

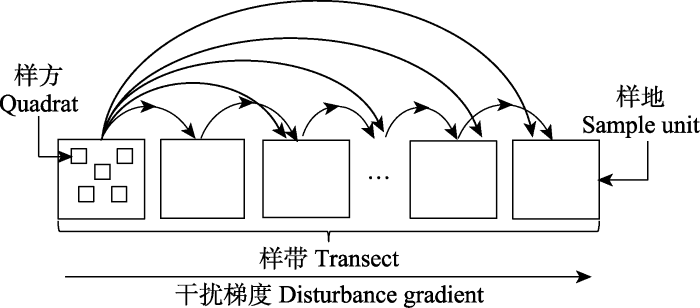 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT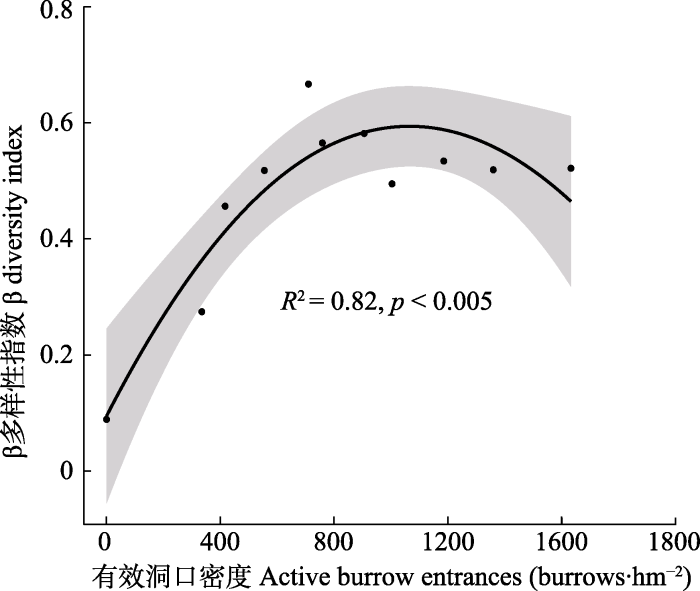 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT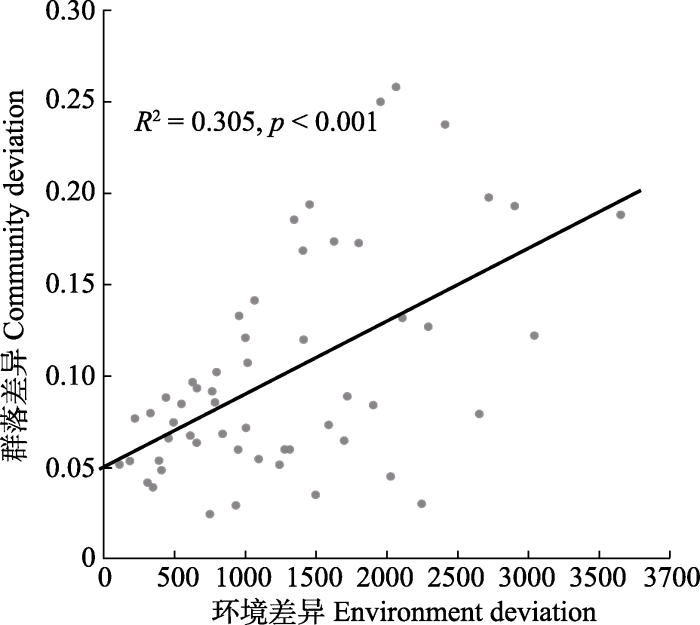 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT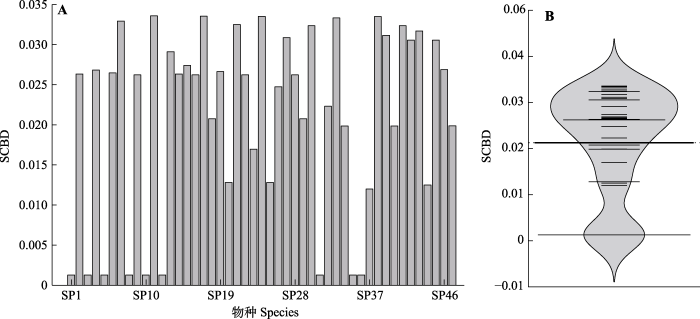 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT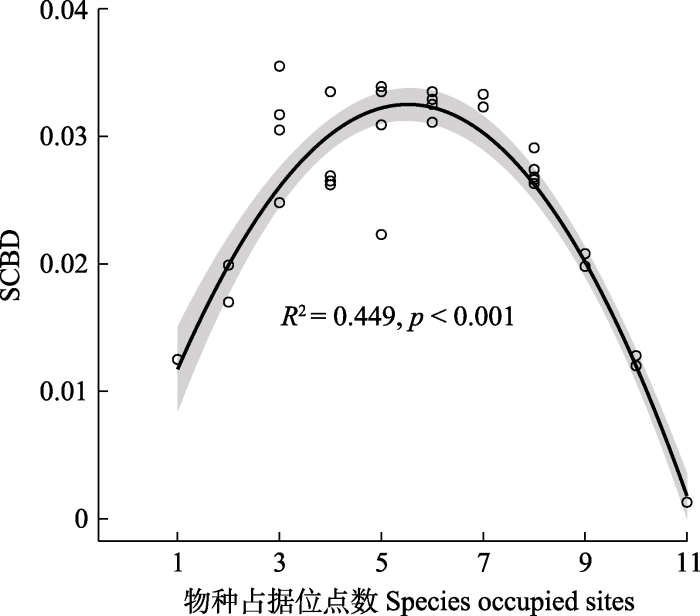 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT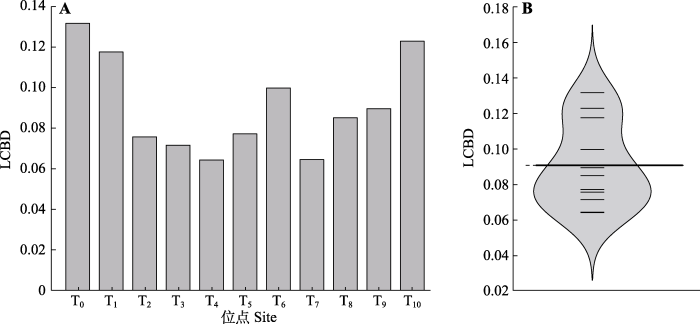 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT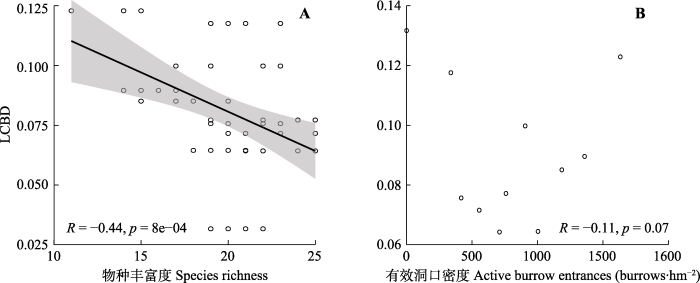 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}