Effects of nitrogen and phosphorus addition at early-spring and middle-summer on ecosystem carbon exchanges of a degraded community in Nei Mongol typical steppe
Ya-Xiang LÜ1,2, Zhi-Yan QI1,2, Wei LIU1, Jia-Mei SUN,1,*, Qing-Min PAN1,21State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China
National Natural Science Foundation of China(31870517)
摘要 过度放牧导致的养分“入不敷出”是我国天然草地大面积退化的主要原因之一, 而草地退化又显著影响了草原生态系统的固碳功能。能否通过补充土壤养分来恢复退化草地的固碳功能, 迄今相关研究较少。净生态系统碳交换(NEE)、生态系统呼吸(ER)和生态系统总初级生产力(GEP)是表征生态系统碳循环的重要指标。氮(N)和磷(P)是中国典型草原的主要限制性养分元素, 而草地退化进一步加剧了养分的限制。在退化草地上添加氮磷对碳循环的上述过程(NEE、ER和GEP)有何影响, 以及两种养分之间是否存在互作, 目前尚不清楚。为此, 该研究以内蒙古典型草原的退化草地为研究对象, 选择早春融雪期(4月)和夏季生长期(7月)两个时间节点, 设置不施肥(CK)、N添加(10.5 g·m-2·a-1, NH4NO3)、P添加(7 g·m-2·a-1, KH2PO4)和N、P共同添加((10.5 g N + 7 g P)·m-2·a-1) 4个养分处理, 探究早春和夏季氮磷添加对内蒙古典型草原退化群落碳交换的影响及其互作机制。结果表明: 1)在早春和夏季两个时期, 单独添加N或P对生态系统碳交换过程的影响均未达到显著水平, 而氮磷共同添加可显著提高NEE和GEP。2)早春(4月份)氮磷共同添加对NEE、ER和GEP的互作机制表现为正协同效应, 而夏季(7月份)氮磷共同添加对NEE、ER和GEP的互作机制表现为加性效应。为了恢复退化的典型草原的固碳功能, 氮磷共同添加比单一元素添加效果好, 且早春添加优于夏季添加。该研究对指导退化草地的恢复具有参考价值。 关键词:草地生态系统;草地施肥;草地恢复;碳循环;氮磷互作
Abstract Aims Overgrazing induced reduction in supply of soil nutrients is a major mechanism leading to extensive grassland degradation in China. The capacity of carbon (C) sequestration was demoted in the degraded grassland ecosystems. However, it remains unclear whether the capacity of carbon sequestration in a degraded grassland can be restored by the supplement of nutrients to the soil. Net ecosystem CO2 exchange (NEE), ecosystem respiration (ER) and gross ecosystem productivity (GEP) are important parameters describing the processes of ecosystem carbon cycle. Nitrogen (N) and phosphorus (P) are two limiting nutrients in typical steppe in China. To date, how these two nutrients alone or in combination affect the three parameters of carbon cycle (i.e., NEE, ER and GEP) in a degraded steppe community, especially their interactive effect, is poorly understood. Methods To address these scientific questions, we conducted a field experiment in a degraded typical steppe community. Four treatments of nutrient addition were implemented: no nutrient addition (CK, control), adding N alone (10.5 g·m-2, NH4NO3), adding P alone (7 g·m-2, KH2PO4), and adding two nutrients in combination. Two stages were selected for fertilization: early-spring (April 21) and middle-summer (July 15). Important findings Neither N nor P alone had significant effect on NEE, ER and GEP when nutrients applied at early-spring (April 21) or middle-summer (July 15), while their combination significantly increased the values ofNEE and GEP. 2) N and P exhibited strong synergistic effect on NEE, GEPand ER when applied in combination at early-spring (April 21), while a consistent additive effect between the two nutrients on the three carbon exchange parameters was observed when applied at middle-summer (July 15). Our findings have implications for the restoration of degraded grasslands. To restore the capacity of carbon sequestration of the degraded typical steppe ecosystem, supplying N and P in combination is better than a single nutrient alone, and appling these nutrients at spring is better than at summer. Keywords:grassland ecosystem;fertilization in grassland;grassland restoration;carbon cycle;interaction between nitrogen and phosphorus
PDF (1927KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 吕亚香, 戚智彦, 刘伟, 孙佳美, 潘庆民. 早春和夏季氮磷添加对内蒙古典型草原退化群落碳交换的影响. 植物生态学报, 2021, 45(4): 334-344. DOI: 10.17521/cjpe.2020.0277 LÜ Ya-Xiang, QI Zhi-Yan, LIU Wei, SUN Jia-Mei, PAN Qing-Min. Effects of nitrogen and phosphorus addition at early-spring and middle-summer on ecosystem carbon exchanges of a degraded community in Nei Mongol typical steppe. Chinese Journal of Plant Ecology, 2021, 45(4): 334-344. DOI: 10.17521/cjpe.2020.0277
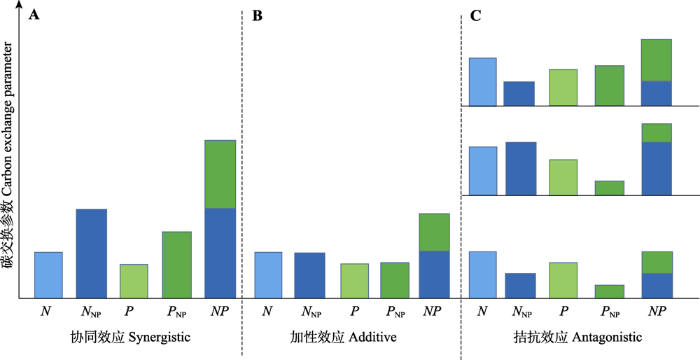
Fig. 1A conceptual framework illustrating interactive effects between nitrogen and phosphorus on parameters of carbon exchanges. N, effect of nitrogen on parameters of carbon exchanges under the scenario of only nitrogen addition; P, effect of phosphorus on parameters of carbon exchanges under the scenario of only phosphorus addition. NNP, effect of nitrogen on parameters of carbon exchanges under the scenario of nitrogen and phosphorus co-addition;NP, co-effect of nitrogen and phosphorus on parameters of carbon exchanges under the scenario of nitrogen and phosphorus co-addition; PNP, effect of phosphorus on parameters of carbon exchanges under the scenario of nitrogen and phosphorus co-addition.
新窗口打开|下载原图ZIP|生成PPT 图3不同时期氮磷添加对净生态系统碳交换(NEE, A)、生态系统呼吸(ER, B)和总生态系统生产力(GEP, C)的影响(平均值±标准误)。不同小写字母表示相同时期添加养分的各处理间差异达到显著水平(p< 0.05)。CK, 对照((0 g N + 0 g P)·m-2·a-1); N, 氮添加(10.5 g·m-2·a-1); P, 磷添加(7 g·m-2·a-1); NP, 氮磷共同添加((10.5 g N + 7 g P)·m-2·a-1)。
Fig. 3Effects of timing of nitrogen and phosphorus addition on net ecosystem carbon exchange (NEE, A), ecosystem respiration (ER, B) and gross ecosystem productivity (GEP, C)(mean ± SE). Different lowercase letters indicate significant difference between treatments of nutrient addition at the same date (p < 0.05). CK, control ((0 g N +0 g P)·m -2·a-1); N, nitrogen addition at a rate of 10.5 g·m-2·a-1; P, phosphorus addition at the rate of 7 g·m-2·a-1; NP, nitrogen in combination with phosphorus addition at the rates of 10.5 and 7 g·m-2·a-1, respectively.
新窗口打开|下载原图ZIP|生成PPT 图4氮(N)在无磷(P)添加和有磷添加情况下对净生态系统碳交换(NEE, A)、生态系统呼吸(ER, B)和总生态系统生产力(GEP, C)的作用以及磷在无氮添加和有氮添加情况下对NEE (D), ER (E)和GEP (F)的作用(平均值±标准误)。**,p < 0.01; ***, p < 0.001; ns, p> 0.05。
Fig. 4Nitrogen (N)-induced changes in net ecosystem carbon exchange (NEE, A), ecosystem respiration (ER,B) and gross ecosystem productivity (GEP, C) with and without phosphorus (P) fertilization, and P-induced changes in NEE (D), ER(E) and GEP (F) with and without N fertilization (mean ± SE). **, p < 0.01; ***, p < 0.001; ns, p> 0.05.
Fig. 5Net ecosystem carbon exchange (NEE, A, D), ecosystem respiration (ER, B, E) and gross ecosystem productivity (GEP, C, F) in relation to soil temperature and soil moisture.
ÅgrenGI, WetterstedtJÅM, BillbergerMFK (2012). Nutrient limitation on terrestrial plant growth—Modeling the interaction between nitrogen and phosphorus New Phytologist, 194, 953-960. DOI:10.1111/nph.2012.194.issue-4URL [本文引用: 1]
AllgeierJE, RosemondAD, LaymanCA (2011). The frequency and magnitude of non-additive responses to multiple nutrient enrichment Journal of Applied Ecology, 48, 96-101. DOI:10.1111/j.1365-2664.2010.01894.xURL [本文引用: 2]
BagchiS, RitchieME (2010). Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition Ecology Letters, 13, 959-968. [本文引用: 1]
BaiX, ChengJH, ZhengSX, ZhanSX, BaiYF ( 2014). Ecophysiological responses of Leymus chinensis to nitrogen and phosphorus additions in a typical steppe Chinese Journal of Plant Ecology, 38, 103-115. DOI:10.3724/SP.J.1258.2014.00010URL [本文引用: 2]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia Grasslands Global Change Biology, 16, 358-372. DOI:10.1111/(ISSN)1365-2486URL [本文引用: 4]
BaiYF, WuJG, XingQ, PanQM, HuangJH, YangDL, HanXG (2008). Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau Ecology, 89, 2140-2153. DOI:10.1890/07-0992.1URL [本文引用: 1]
BubierJL, MooreTR, BledzkiLA (2007). Effects of nutrient addition on vegetation and carbon cycling in an ombrotrophic bog Global Change Biology, 13, 1168-1186. DOI:10.1111/gcb.2007.13.issue-6URL [本文引用: 1]
ChenSP, LinGH, HuangJH, JeneretteGD (2009). Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe Global Change Biology, 15, 2450-2461. DOI:10.1111/gcb.2009.15.issue-10URL [本文引用: 1]
ChenZZ, ShengXW, YangZG, HuangDH ( 1985). The ecological effects of fertiltzation during rainy season on various types of steppe on the Xilin River Valley, Inner Mongolia//Inner Mongolia Grassland Ecosystem Research Station, Academia Sinica. Research on Grassland Ecosystem:No.1. Science Press, Beijing.225-232. [本文引用: 3]
DongCC, WangW, LiuHY, XuXT, ZengH (2019). Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: evidence from soil extracellular enzyme stoichiometry Ecological Indicators, 101, 453-464. DOI:10.1016/j.ecolind.2019.01.046URL [本文引用: 1]
ElserJJ, BrackenMES, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems Ecology Letters, 10, 1135-1142. DOI:10.1111/ele.2007.10.issue-12URL [本文引用: 2]
GuanSY, QiPQ, KangSA, ChangJB ( 1997). Effects on the steppe soil nutrient contents under different grazing intensities//Inner Mongolia Grassland Ecosystem Research Station, Academia Sinica. Research on Grassland Ecosystem: No.5. Science Press, Beijing. 17-22. [本文引用: 1]
HanWX, FangJY, GuoDL, ZhangY (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China New Phytologist, 168, 377-385. DOI:10.1111/nph.2005.168.issue-2URL [本文引用: 3]
HaoYB, ZhouCT, LiuWJ, LiLF, KangXM, JiangLL, CuiXY, WangYF, ZhouXQ, XuCY (2017). Aboveground net primary productivity and carbon balance remain stable under extreme precipitation events in a semiarid steppe ecosystem Agricultural and Forest Meteorology, 240- 241, 1-9. [本文引用: 1]
HarpoleWS, PottsDL, SudingKN (2007). Ecosystem responses to water and nitrogen amendment in a California grassland Global Change Biology, 13, 2341-2348. DOI:10.1111/gcb.2007.13.issue-11URL [本文引用: 1]
HasiMQ, ZhangXY, NiuGX, WangYL, HuangJH ( 2018). Effects of nitrogen addition on ecosystem CO2 exchange in a meadow steppe, Inner Mongolia Chinese Bulletin of Botany, 53, 27-41. [本文引用: 1]
JasoniRL, SmithSD, Arnone IIIJA (2005). Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2 Global Change Biology, 11, 749-756. DOI:10.1111/gcb.2005.11.issue-5URL [本文引用: 1]
JiangJ, WangYP, YangYH, YuMX, WangC, YanJH (2019). Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type Plant and Soil, 440, 523-537. DOI:10.1007/s11104-019-04119-5 [本文引用: 1] Aims Co-limitation of ecosystem productivity by nitrogen (N) and phosphorus (P) is gaining increasing recognition, but how co-limitation through N and P interactions differs among different terrestrial ecosystems remains unclear. Methods We performed a meta-analysis of 133 independent studies conducted in four natural terrestrial ecosystems to examine the interactive effects of N and P additions on ten plant growth-related variables. Results Adding N and P individually or in combination significantly increased aboveground biomass (AGB), and the interactions were uniformly synergistic for AGB, and additive for belowground biomass (BGB), but variable for other eight growth-related variables among four different ecosystems. The interaction was synergistic for leaf P and soil NO3-N only in tropical forests, and antagonistic for soil available P (AP) in tropical forests, leaf N in grasslands, root P in wetlands, and leaf P and soil NH4-N in tundra. The interaction for leaf N: P ratios was additive only in tropical forests, and synergistic in the other three ecosystems. Conclusions Our results highlighted the interactions of N and P additions can promote uptake of both nutrients by plants, and plants tend to maintain the optimal nutrient balance for growth and reproduction through regulating biomass production and tissue nutrient concentrations.
LiJD, LiuJX ( 1981). Preliminary report on application of nitrogen fertilizer in Leymus chinensisgrassland Chinese Grassland, 3, 58-59. [本文引用: 1]
LiSL, ChenYJ, GuanSY, KangSA ( 2002). Relationships between soil degradation and rangeland degradation Journal of Arid Land Resources and Environment, 16, 92-95. [本文引用: 1]
LiYH ( 1988). The divergence and convergence of Aneurolepidium chinense steppe and Stipa grandis steppe under the grazing influence in Xilin River valley, Inner Mongolia Acta Phytoecologica et Geobotanica Sinica, 12, 189-196. [本文引用: 1]
NiuSL, WuMY, HanY, XiaJY, ZhangZ, YangHJ, WanSQ (2010). Nitrogen effects on net ecosystem carbon exchange in a temperate steppe Global Change Biology, 16, 144-155. DOI:10.1111/(ISSN)1365-2486URL [本文引用: 1]
NiuSL, YangHJ, ZhangZ, WuMY, LuQ, LiLH, HanXG, WanSQ (2009). Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe Ecosystems, 12, 915-926. DOI:10.1007/s10021-009-9265-1URL [本文引用: 2]
OberbauerSF, TweedieCE, WelkerJM, FahnestockJT, HenryGHR, WebberPJ, HollisterRD, WalkerMD, KuchyA, ElmoreE, StarrG (2007). Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients Ecological Monographs, 77, 221-238. DOI:10.1890/06-0649URL [本文引用: 1]
PanQM, BaiYF, HanXG, YangJC ( 2004). Studies on the fate of labelled nitrogen applied to a Leymus Chinensis community of typical steppe in Inner Mongolia grassland Acta Phytoecologica Sinica, 28, 665-671. [本文引用: 1]
RaichJW, PotterCS (1995). Global patterns of carbon dioxide emissions from soils Global Biogeochemical Cycles, 9, 23-36. DOI:10.1029/94GB02723URL [本文引用: 1]
RongYP, HanJG, WangP, MaoPS ( 2001). The effects of grazing intensity on soil physics and chemical properties Grassland of China, 23, 41-47. [本文引用: 1]
SaarnioS, JärviöS, SaarinenT, VasanderH, SilvolaJ (2003). Minor changes in vegetation and carbon gas balance in a boreal mire under a raised CO2 or NH4NO3 supply Ecosystems, 6, 46-60. DOI:10.1007/s10021-002-0208-3URL [本文引用: 1]
SanyalSK, de DattaSK (1991). Chemistry of Phosphorus Transformations in Soil Springer,New York. [本文引用: 1]
ShenHH, ZhuYK, ZhaoX, GengXQ, GaoSQ, FangJY ( 2016). Analysis of current grassland resources in China Chinese Science Bulletin, 61, 139-154. [本文引用: 1]
TianDS, NiuSL, PanQM, RenTT, ChenSP, BaiYF, HanXG (2016). Nonlinear responses of ecosystem carbon fluxes and water-use efficiency to nitrogen addition in Inner Mongolia grassland Functional Ecology, 30, 490-499. DOI:10.1111/fec.2016.30.issue-3URL [本文引用: 1]
VoseJM, ElliottKJ, JohnsonDW, TingeyDT, JohnsonMG (1997). Soil respiration response to three years of elevated CO2 and N fertilization in ponderosa pine (Pinus ponderosa Doug. ex Laws.) Plant and Soil, 190, 19-28. DOI:10.1023/A:1004204129128URL [本文引用: 1]
WangB, LiJ, JiangWW, ZhaoL, GuS ( 2012). Impacts of the rangeland degradation on CO2 flux and the underlying mechanisms in the Three-River Source Region on the Qinghai-Tibetan Plateau China Environmental Science, 32, 1764-1771. [本文引用: 1]
WangJW, CaiYQ ( 1988). Study on genesis, types and characteristics of the soils of the Xilin River Basin//Inner Mongolia Grassland Ecosystem Research Station, Academia Sinica. Research on Grassland Ecosystem: No.3. Science Press, Beijing. 24-83. [本文引用: 2]
WangWY, WangQJ, WangG, JingZC ( 2007). Effects of land degradation and rehabilitation on vegetation carbon and nitrogen content of alpine meadow in China Journal of Plant Ecology(Chinese Version) , 31, 1073-1078. [本文引用: 1]
XiaJY, NiuSL, WanSQ (2009). Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe Global Change Biology, 15, 1544-1556. DOI:10.1111/gcb.2009.15.issue-6URL [本文引用: 1]
YanLM, ChenSP, HuangJH, LinGH (2011). Increasing water and nitrogen availability enhanced net ecosystem CO2 assimilation of a temperate semiarid steppe Plant and Soil, 349, 227-240. DOI:10.1007/s11104-011-0864-1URL [本文引用: 1]
YangQ, WangW, ZengH ( 2018). Effects of nitrogen addition on the plant diversity and biomass of degraded grasslands of Nei Mongol, China Chinese Journal of Plant Ecology, 42, 430-441. DOI:10.17521/cjpe.2017.0135URL [本文引用: 1]
YangY ( 2005). Relationship Between Soil Available Phosphorus and Above-ground Net Primary Productivity and Response of Soil Available Phosphorus to Nutrient Addition in Leymus chinensis Steppe in Inner Mongolia. Master degree dissertation, Institute of Botany, Chinese Academy of Sciences,Beijing. 27-31. [本文引用: 2]
ZhouGY, LuoQ, ChenYJ, HeM, ZhouLY, FrankD, HeYH, FuYL, ZhangBC, ZhouXH (2019). Effects of livestock grazing on grassland carbon storage and release override impacts associated with global climate change Global Change Biology, 25, 1119-1132. DOI:10.1111/gcb.2019.25.issue-3URL [本文引用: 1]
ZhouHK, ZhaoXQ, ZhouL, LiuW, LiYN, TangYH ( 2005). A study on correlations between vegetation degradation and soil degradation in the ‘Alpine Meadow’ of the Qinghai- Tibetan Plateau Acta Prataculturae Sinica, 14, 31-40. [本文引用: 1]
ZhouW, GangCC, ZhouL, ChenYZ, LiJL, JuWM, OdehI (2014). Dynamic of grassland vegetation degradation and its quantitative assessment in the northwest China Acta Oecologica, 55, 86-96. DOI:10.1016/j.actao.2013.12.006URL [本文引用: 1]
ZongN, ShiPL, JiangJ, MengFS, MaWL, XiongDP, SongMH, ZhangXZ ( 2013). Effects of fertilization and grazing exclosure on vegetation recovery in a degraded alpine meadow on the Tibetan Plateau Chinese Journal of Applied and Environmental Biology, 19, 905-913. DOI:10.3724/SP.J.1145.2013.00905URL [本文引用: 1]
ZongN, ShiPL, NiuB, JiangJ, SongMH, ZhangXZ, HeYT ( 2014). Effects of nitrogen and phosphorous fertilization on community structure and productivity of degraded alpine meadows in northern Tibet, China Chinese Journal of Applied Ecology, 25, 3458-3468. [本文引用: 1]
Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe 1 2009
... 实验采用静态同化箱和LI-850 CO2/H2O分析仪(LI-COR, Lincoln, USA)测定NEE和ER (Chen et al., 2009 ).在实验开始之前(4月10日), 在每个小区内随机安置了1个有水槽的同化箱底座(0.5 m × 0.5 m), 该底座入土5 cm, 其上沿与地面持平.每次测定时, 首先把同化箱与LI-850 CO2/H2O分析仪通过塑料管连接好, 保证气密性.然后把一个高透明的有机玻璃同化箱(0.5 m × 0.5 m × 0.5 m)安放在有水槽的底座上, 水槽注满水使同化箱处于密闭状态.同化箱内对角线装有2个小风扇, 使箱内气体混匀, 测试时选取晴朗无风的8:00-11:00, 如果计划测定日期的前一天有雨, 至少经历两个晴天, 排除水分干扰后再开始测定.测定分两个过程: NEE和ER.测量NEE前, 把同化箱举起通风待分析仪数据显示稳定后将同化箱放置底座上并水封, 然后开始测定, 1 s记录一次数据, 记录时间大约为1.5 min (90个数据), 为了避免初始数据因环境改变不稳定, 在计算时选取中间60个数据, 同时记录箱内初始温度和结束温度.NEE测定结束后, 把同化箱举起通风待分析仪数据显示稳定后将同化箱放置底座上并水封, 开始测定前用一块黑布完全罩住同化箱形成暗环境, 同样也是测量1.5 min取60个数据计算, 同样记录箱内初始温度和结束温度.本实验共进行了6次生态系统碳交换测定, 分别在8月13日、8月18日、8月19日、8月31日、9月7日和9月14日.全年逐日气温数据和逐日降水量数据由中国科学院内蒙古草原生态系统定位研究站提供. ...
Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: evidence from soil extracellular enzyme stoichiometry 1 2019
Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe 1 2009
... 大多数陆地生态系统的生产力都受到氮的限制(Elser et al., 2007 ; Harpole et al., 2011 ; Ågrenet al., 2012 ).对于退化的草地而言, 由于过度放牧或刈割将大量的氮带出系统, 导致氮的限制作用进一步加剧(Bai et al., 2010 ).因此, 在退化草地上施用氮肥, 可以缓解氮限制作用, 从而加速退化草地的恢复(陈佐忠等, 1985).由于氮循环和碳循环是密切耦联的两个过程, 前人就氮对草地生态系统碳交换的影响已经开展了一些研究.在内蒙古典型草原, 添加氮能显著提高草地的NEE和GEP (Xia et al., 2009 ; Niu et al., 2010 ; Yan et al., 2011 ), 但是当氮添加量达到10 g·m-2·a-1时, NEE和GEP的响应趋于饱和(Tian et al., 2016 ; 哈斯木其尔等, 2018).但是, 在其他系统的研究发现, 如果氮不是限制性资源, 氮添加对生态系统NEE可能没有显著影响(Vose et al., 1997 ; Saarnio et al., 2003 ), 甚至有负效应(Bubier et al., 2007 ; Harpole et al., 2007 ).值得一提的是, 以上研究大部分集中于多年围封样地为代表的成熟草地群落.由于退化草地的氮限制更为严重, 在退化草地上研究氮添加对生态系统碳交换的影响, 对于认识氮在草地生态系统碳循环中的作用, 特别是对退化草地生态系统碳平衡的影响至关重要. ...
Increasing water and nitrogen availability enhanced net ecosystem CO2 assimilation of a temperate semiarid steppe 1 2011
... 大多数陆地生态系统的生产力都受到氮的限制(Elser et al., 2007 ; Harpole et al., 2011 ; Ågrenet al., 2012 ).对于退化的草地而言, 由于过度放牧或刈割将大量的氮带出系统, 导致氮的限制作用进一步加剧(Bai et al., 2010 ).因此, 在退化草地上施用氮肥, 可以缓解氮限制作用, 从而加速退化草地的恢复(陈佐忠等, 1985).由于氮循环和碳循环是密切耦联的两个过程, 前人就氮对草地生态系统碳交换的影响已经开展了一些研究.在内蒙古典型草原, 添加氮能显著提高草地的NEE和GEP (Xia et al., 2009 ; Niu et al., 2010 ; Yan et al., 2011 ), 但是当氮添加量达到10 g·m-2·a-1时, NEE和GEP的响应趋于饱和(Tian et al., 2016 ; 哈斯木其尔等, 2018).但是, 在其他系统的研究发现, 如果氮不是限制性资源, 氮添加对生态系统NEE可能没有显著影响(Vose et al., 1997 ; Saarnio et al., 2003 ), 甚至有负效应(Bubier et al., 2007 ; Harpole et al., 2007 ).值得一提的是, 以上研究大部分集中于多年围封样地为代表的成熟草地群落.由于退化草地的氮限制更为严重, 在退化草地上研究氮添加对生态系统碳交换的影响, 对于认识氮在草地生态系统碳循环中的作用, 特别是对退化草地生态系统碳平衡的影响至关重要. ...
 ,1,*, 潘庆民1,2
,1,*, 潘庆民1,2

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT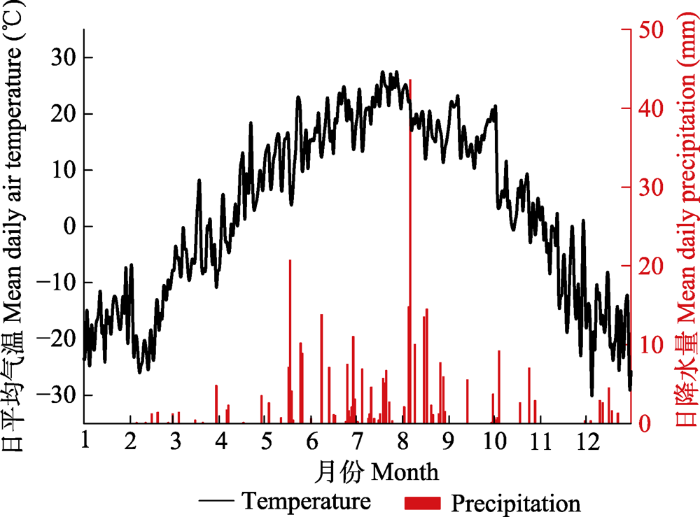 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT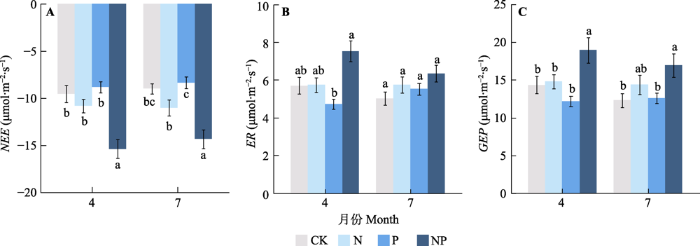 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT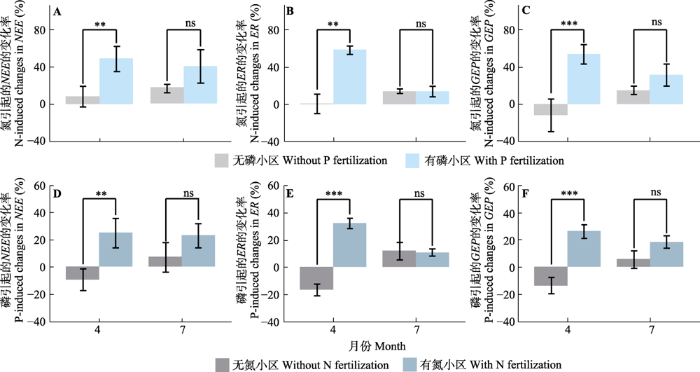 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT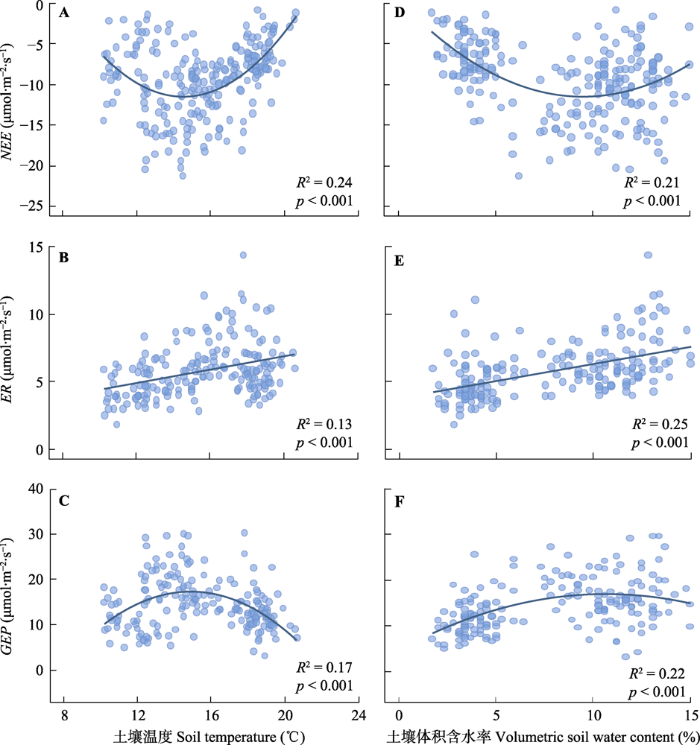 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}