 ,1,2,*
,1,2,*Latitudinal variability and driving factors of functional diversity in Pinus thunbergiicommunities across sea-islands in Eastern China
Jiao-Xing SHI1, Ming-Shan XU1, Xiao-Chen FANG1, Li-Ting ZHENG1, Yu ZHANG1, Di-Feng BAO1, An-Na YANG1, En-Rong YAN,1,2,*通讯作者: *(eryan@des.ecnu.edu.cn)
编委: 刘国方
责任编辑: 赵航
收稿日期:2020-07-8接受日期:2020-09-24网络出版日期:2021-02-20
| 基金资助: |
Corresponding authors: *(eryan@des.ecnu.edu.cn)
Received:2020-07-8Accepted:2020-09-24Online:2021-02-20
| Fund supported: |

摘要
黑松(Pinus thunbergii)群落在中国东海和黄海海域的海岛广泛分布, 研究其功能多样性的纬度变化特征及其影响因素, 有助于揭示是否在高度隔离的片断化景观中, 同一植物群落类型的生物多样性也具有显著的纬度地带性特征。该研究调查和测定了中国东部海域跨越13个纬度的27个海岛60个黑松群落的物种组成和植物功能性状, 分析了生物因素(黑松优势度和物种多样性)与非生物因素(年平均气温、总太阳辐射和干旱指数)与黑松群落功能多样性的关系, 并利用广义线性回归和方差分解揭示了各影响因素对黑松群落功能多样性的相对影响。主要结果: 随纬度升高, 群落的黑松优势度和物种丰富度, 以及枝干性状和枝叶性状总体的功能丰富度、功能分散度、Rao二次熵均显著降低, 但叶片功能多样性无显著变化趋势。黑松优势度、物种多样性和气候综合解释了枝叶性状总体功能丰富度、分散度和Rao二次熵变异的63%、47%和39%, 枝干性状功能丰富度、分散度和Rao二次熵变异的56%、67%和53%, 对叶片性状功能多样性的综合解释度较低(21%-30%)。物种丰富度和辛普森多样性显著增加叶功能多样性, 香农-维纳多样性显著降低叶功能丰富度。干旱度显著增加枝叶性状总体和枝干功能多样性, 年平均气温显著降低枝干功能分散度和Rao二次熵。该结果表明, 中国东部海岛黑松群落的功能多样性具有明显的纬度格局, 气候因素和物种丰富度是决定黑松群落功能多样性纬度变异的重要因素。
关键词:
Abstract
Aims The Pinus thunbergii communities is widespread across islands in both Yellow Sea and East China Sea. The objective of this study is to examine the latitudinal pattern of functional diversity and abiotic and biotic drivers of P. thunbergii communities. Our aims are to advance understanding of whether the zonal character of latitude oriented pattern of biodiversity still holds in the same community type across the highly isolated and fragmented landscape.
Methods We investigated community structure and measured plant functional traits across 60 P. thunbergii communities in 27 islands, spanning 13 degree in latitudes of Eastern China. Linear regression was used to analyze the relationships between functional diversity of the P. thunbergii community and each of the biotic factors (the dominance of P. thunbergii and species diversity) and abiotic factors (annual mean temperature, total solar radiation and aridity index). The relative importance of abiotic and biotic factors on the functional diversity of P. thunbergii communities was determined by using the generalized linear model and variance decomposition.
Important findings With the increasing latitude, the dominance of P. thunbergii,species richness, and functional richness, functional dispersion and Rao's quadratic entropy (RaoQ) of stem traits alone and stem and leaf traits in combination decreased significantly but those of leaf traits did not show clear trend, across P. thunbergii communities. The dominance of P. thunbergii, species richness and climatic factors jointly explained 63%, 47% and 39% of variation in each of functional richness, functional dispersion and RaoQ of the combination of leaf and stem traits, and 56%, 67% and 53% of variation in each of functional richness, functional dispersion and RaoQ of stem traits, but small variations in leaf traits (21%-30%). Species richness and Simpson diversity significantly increased but Shannon-Wiener diversity significantly decreased leaf functional richness. Aridity significantly increased functional diversity of stem traits and the combination of leaf and stem traits. Annual mean temperature significantly decreased functional dispersion and RaoQ of wood traits. These results indicate that there is a clear latitudinal pattern of functional diversity in P. thunbergii communities across islands. Climate and species richness play the key roles for shaping the latitudinal variations in functional diversity of P. thunbergii communities across islands in Eastern China.
Keywords:
PDF (1573KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
石娇星, 许洺山, 方晓晨, 郑丽婷, 张宇, 鲍迪峰, 杨安娜, 阎恩荣. 中国东部海岛黑松群落功能多样性的纬度变异及其影响因素. 植物生态学报, 2021, 45(2): 163-173. DOI: 10.17521/cjpe.2020.0227
SHI Jiao-Xing, XU Ming-Shan, FANG Xiao-Chen, ZHENG Li-Ting, ZHANG Yu, BAO Di-Feng, YANG An-Na, YAN En-Rong.

海岛在维持生物多样性方面扮演着重要角色(Courchamp et al., 2014; Pati?oet al., 2017)。我国是海岛大国, 拥有1万多个大小不同的岛屿。在东部海域, 海岛经历了长期的人类干扰和开发利用, 除少数无人小岛由于人类难以利用, 以及个别海岛由于寺庙保护而保留少量海岛特有植被外, 大部分海岛的天然森林和灌丛被严重破坏(妥彬等, 2019)。由于海岛环境条件一般比较恶劣, 且与大陆隔离, 天然植被种源难以自然传播, 因此, 长期以来, 我国东部海岛植被处于退化和半退化状态, 能够用于植被恢复的树种资源非常有限。
黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等, 2011), 因此被作为我国南北海岛植被恢复的优先推广树种。20世纪60年代开始, 通过人工种植和飞播等手段, 在沿海地区和海岛营造了大量的黑松林(许景伟等, 2003), 并成为了我国东部海域海岛的主要植被类型。经过长期自然更新, 黑松林下逐渐发育形成了地带性阔叶木本植物组成的灌木层, 从而有效扭转了海岛生态退化的不利局面, 在生物多样性维持、防风、固土、降盐碱和水源涵养等方面发挥着重要生态功能。然而, 自1996年以来, 松材线虫病在我国东部地区大规模暴发, 对黑松林构成了灭顶之灾, 导致绝大多数海岛黑松大树的死亡, 黑松在森林群落中的比例严重下降。与这个不利的方面相比, 黑松的大量消退却为林下被长期压制的地带性树种打开了生长空间, 经过近20年的自然演替, 黑松林的群落结构和物种组成已发生了显著改变。例如, 经过13年的自然恢复, 舟山群岛松材线虫灾害迹地的黑松林已演替为针阔混交林或阔叶林, 乔木层的物种多样性增加(王国明等, 2011)。在温带地区, 黑松群落的天然更新过程受到土壤、坡度、坡向等的影响(韩广轩等, 2010)。
气候和物种成分是影响大尺度植物群落多样性分布格局的重要因素。在我国的近岸海域, 海岛黑松林分布纬度自北向南跨越30°, 形成了典型的纬度地带梯度。同时, 这些海岛的海拔较低(除台湾岛外), 且经度变幅小, 一定程度上可以剔除海拔和经度变化对生物多样性的影响。因此, 研究我国东部海岛纬度梯度上黑松群落的物种和功能多样性的变化特征, 尤其是物种组成和气候等因素对功能多样性的影响, 有助于深入理解海岛植被结构和功能的维持机制。
大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al., 2019)。气候(温度、降水及光照)在纬度上的差异会导致植物功能群的变化(孟婷婷等, 2007), 通常, 常绿植物倾向于在低纬度分布, 而落叶植物更多出现在中高纬度(Givnish, 2002)。同时, 低纬度地区比高纬度地区拥有更高的物种多样性(MacArthur, 1965)。一般来讲, 物种多样性与功能多样性显著正关联(Whittaker et al., 2014; Kumordzi et al., 2015; 郑丽婷等, 2018), 但是, 当受环境胁迫或生物竞争等影响时, 二者的关系也可能为负相关或者不相关(de Bello et al., 2006; 李晓刚等, 2011)。基于此, 我们预测, 在中国东部海岛纬度梯度上, 随着年平均气温和降水量的减少, 黑松群落的物种多样性会显著下降; 同时, 黑松属于偏暖性树种, 其在群落中的优势度也会随纬度升高而下降。我们进一步预测(零假说), 在环境梯度上, 由于植物不同器官的性状分异性大, 某些性状更能体现其对环境变化的适应, 而有些性状对环境梯度的响应性差(Wright et al., 2001), 甚至, 如果枝干和叶片性状对环境的响应模式不一致(Kang et al., 2014), 那么黑松群落的功能多样性有可能不随纬度而变化。反之, 如果黑松群落的功能结构受气候和物种多样性的共同影响, 那么其功能多样性会随纬度梯度上物种多样性的减少而下降(备择假说)。
本研究以中国东海和黄海的27个海岛上的60个黑松群落为研究对象, 在探索群落物种和功能多样性纬度变化特征的基础上, 分析群落物种多样性和黑松优势度等生物因素, 以及干旱指数、年平均气温、总太阳辐射等非生物因素对群落功能丰富度、分散度和Rao二次熵的相对贡献, 以期揭示海岛纬度梯度上黑松群落功能多样性变化的驱动因素。本研究的开展有助于理解大尺度海岛的生物多样性维持机制, 也可为我国东部海岛的植被恢复提供理论支撑。
1 材料和方法
1.1 研究区概况
依据纬度梯度, 本研究选取我国黄海和东海海域从山东到福建的27个海岛(25.06°-37.98° N, 119.13°-122.79° E), 依次跨越暖温带、北亚热带、中亚热带和南亚热带气候区(图1)。随纬度上升, 年太阳辐射量和年平均风速增大, 年平均气温、年降水量和湿润度减小。年平均风速变化范围为3.4-4.8 m·s-1, 年平均气温变化范围为11.3-20.5 ℃, 年降水量变化范围为484-1 523 mm。土壤类型从北到南依次从棕壤、黄壤过渡到红壤。根据国家林业和草原科学数据共享服务平台(图1
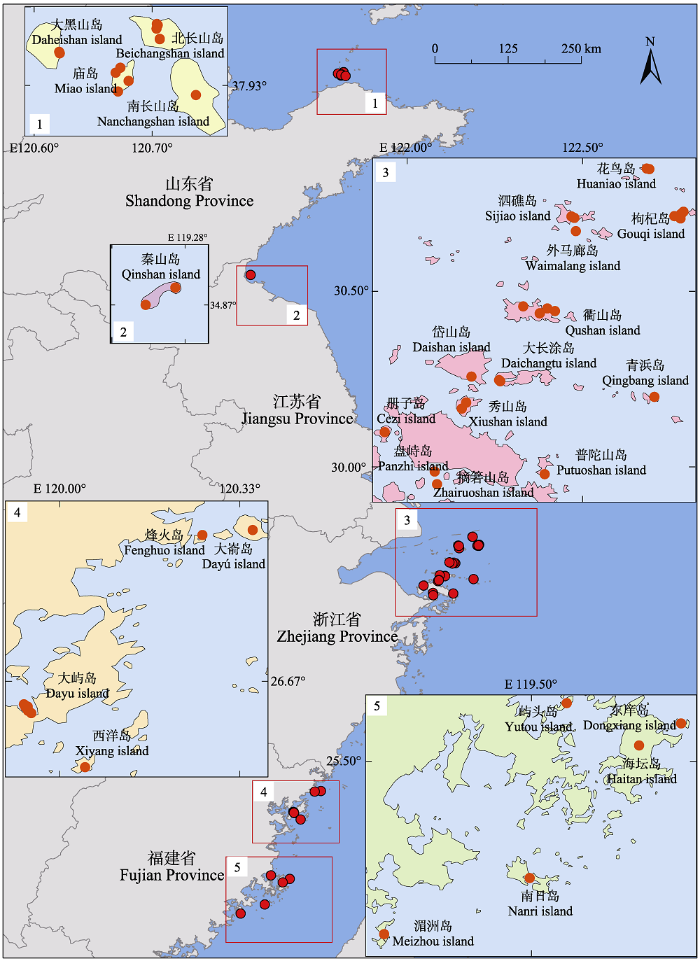 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1中国东部27个海岛与黑松群落研究样地(红/橙色圆点)的地理分布。
Fig. 1Geographic locations of studied plots of Pinus thunbergii community across 27 islands in Eastern China.
1.2 样地调查和气候因子获取
2016-2019年的每年7-8月, 在对各海岛植被踏查后, 选取典型黑松群落设置样地, 森林样方大小为20 m × 20 m, 灌丛样方大小为10 m × 10 m。样地建立后, 记录和测量所有高度大于0.5 m的木本植物名称、基径、胸径、高度、冠幅、枝下高、叶下高和郁闭度等群落学特征, 以及地形、地貌和土壤类型等。此外, 对于难以攀登的悬崖上的黑松群落, 采用法瑞学派的无样地调查法记录物种组成、物种的多盖度和聚集度, 以及群落分层情况(高度和盖度)等。本研究共调查了21个海岛的52个黑松群落样地, 另外, 从《福建海岛植被》(孔繁昇等, 1999)摘录了6个海岛8个黑松群落的样地资料用以合并分析, 详细信息见表1。本研究涉及的木本植物总计126种, 隶属51科93属。需要指出的是, 我国东部海域海岛主要集中分布在东南沿海, 而北部海域海岛数量偏少, 如果按照北部海域的海岛数量设置取样原则, 就需要牺牲南部海域的海岛样本数, 从而不能完全反映其整体特征。根据这种现实, 我们尽量将北部海域有黑松分布的海岛全纳入调查范围, 而在南方, 由于海岛数量太多, 尽量选取不同面积、不同隔离度的海岛来反映其整体状况。Table 1
表1
表1中国东部27个海岛的黑松群落样地概况
Table 1
| 区域 Region | 海岛 Island | 纬度/经度 Latitude/Longitude | 样地数 Number of plots | 物种 丰富度 Species richness | 年平均降水量 Mean annual precipitation (mm) | 干旱 指数 Aridity index | 年平均气温 Mean annual air temperature (℃) | 总太阳辐射 Total solar radiation (MJ·m-2·a-1) |
|---|---|---|---|---|---|---|---|---|
| 1 | 北长山岛、南长山岛、庙岛、大黑山岛 Beichangshan island, Nanchangshan island, Miao island, Daheishan island | 37.92°-37.99° N/ 120.61°-120.74° E | 12 | 5-10 | 480-505 | 1.78-1.88 | 11.3-11.8 | 43 566-43 944 |
| 2 | 秦山岛 Qinshan island | 34.87° N/119.28° E | 2 | 5-6 | 913 | 0.99 | 13.9 | 27 158 |
| 3 | 册子岛、大长涂岛、岱山岛、枸杞岛、花鸟岛、盘峙岛、普陀山岛、青浜岛、衢山岛、泗礁岛、外马廊岛、秀山岛、摘箬山岛 Cezi island, Dachangtu island, Daishan island, Gouqi island, Huaniao island, Panzhi island, Putuoshan island, Qingbang island, Qushan island, Sijiao island, Waimalang island, Xiushan island, Zhairuoshan island | 29.95°-30.85° N/ 121.93°-122.79° E | 28 | 8-21 | 981-1 275 | 0.86-1.12 | 15.8-16.8 | 32 469-33 335 |
| 4 | 大屿岛、大嵛岛*、烽火岛、西洋岛* Dayu island, Dayú island*, Fenghuo island, Xiyang island* | 26.51°-26.95° N/ 119.93°-120.36° E | 10 | 5-19 | 1 405-1 523 | 0.92-0.99 | 17-18.9 | 33 784-34 852 |
| 5 | 东庠岛*、海坛岛*、湄洲岛*、南日岛*、屿头岛 Dongxiang island*, Haitan island*, Meizhou island*, Nanri island*, Yutou island | 25.06°-25.65° N/ 119.13°-119.88° E | 8 | 7-20 | 1 116-1 258 | 1.09-1.25 | 19.6-20.5 | 18 036-35 862 |
新窗口打开|下载CSV
纬度梯度上水热变化最为显著, 与年降水量相比, 干旱指数能更好地表征环境的湿润和干旱胁迫程度, 因此本研究主要分析年平均气温、总太阳辐射和干旱指数对黑松群落生物多样性的影响。各样地的气候信息从全球气候图层数据库(
1.3 植物功能性状测定
与黑松群落学特征调查同步, 参照Pérez- Harguindeguy等(2013)的方法测量以下植物功能性状: 单叶面积、比叶面积、叶干物质含量、单位质量叶稠密度、比枝长、小枝含水率、小枝密度、干材密度以及胡伯尔值。在每个样地, 随机挑选各木本物种的健康植株3株(不足3株的在样地周围选取), 在没有明显遮阴的树冠剪取3-5个枝条, 摘取20片成熟叶片、当年生小枝和较粗枝条, 分别装进密封袋中带回驻地。在室内, 称量成熟叶鲜质量后, 用LI-3100C叶面积仪(LI-COR, Lincoln, USA)测量叶面积。之后, 将叶片风干, 尽快运回华东师范大学, 放置于75 ℃烘箱烘干48 h至恒质量, 称量叶干质量, 分别计算叶干物质含量(干质量除以叶片饱和鲜质量)和比叶面积(叶面积除以叶干质量)。在叶面积测定同时测定叶片厚度、叶柄长度等。对于当年生小枝, 先记录脱叶痕与当年生叶片的数量, 再将当年生叶片摘下, 为了减小误差, 枝长和直径用游标卡尺在不同位置测量3次, 然后计算枝横截面积和体积。之后, 称量其鲜质量, 装入信封, 放置于75 ℃烘箱中烘干至恒质量, 称量干质量。
根据小枝叶片数(包括叶痕)、枝干质量和枝体积, 计算单位质量叶稠密度、小枝含水率、比枝长、枝密度。另外, 将当年生叶片摘下, 测量总叶面积, 根据其与小枝横截面积比值计算胡伯尔值。
最后, 在野外取样过程中, 截取大枝条茎段, 在室内用游标卡尺在3个不同位置测量直径, 然后用美工刀削去树皮, 在称量鲜质量后, 用排水法测量枝条茎段体积, 之后, 放入105 ℃烘箱烘干48 h后称量其干质量, 计算干材密度(干质量除以体积)。
需要说明的是, 对于摘录自孔繁昇等(1999)的8个黑松群落, 本研究按照以下步骤对各群落物种性状进行赋值。首先, 按照就近原则, 利用本研究在福建烽火岛和大屿岛的植物性状测量值, 对8个群落中相应的物种赋值。其次, 若福建区域没有测量某物种的性状, 则利用本研究所测全部海岛中该物种的平均值代替。最后, 对于本研究所有海岛没有测量到的物种, 则利用全球植物性状数据库(
1.4 群落物种多样性和功能多样性指数计算
选用物种丰富度、香农-维纳指数和辛普森指数反映黑松群落的物种多样性情况, 具体计算方法参见张金屯(2011)。此外, 采用黑松在群落内的相对多度表示黑松的优势度。本研究采用多维性状计算各群落的功能丰富度、功能分散度(Laliberté & Legengdre,2010)和Rao二次熵(RaoQ)(Villégeret al., 2008)。考虑到枝叶性状对环境梯度的不同响应强度, 本研究在计算群落总体性状功能多样性的同时, 也分别单独计算基于叶片性状(单叶面积、比叶面积、干物质含量、单位质量叶稠密度)和枝干性状(比枝长、小枝含水率、小枝密度、干材密度以及胡伯尔值)的功能多样性。
功能丰富度(FRic)反映了群落内物种对多维生态空间的利用程度, 根据多维性状空间内生成的最小多边形的体积或面积计算, 如公式(1)所示:
式中, SFci为群落i内性状c所占据的功能性状空间的大小; Rc为所有群落中性状c占据的生态位空间。
功能分散度(FDis)反映了在多维空间中每个物种与重心的平均加权距离, 计算方法见公式(2):
式中, aj是物种j的相对多度, zj是物种j到重心g的加权距离。
Rao二次熵(RaoQ)反映的是群落内物种功能性状的趋异性, 计算方法见公式(3):
式中, Pi和Pj分别为群落中物种i和物种j的个体数占总个体数的百分比; dij为物种i和物种j的功能性状距离(dij在0-1之间);S为群落的总物种数。
1.5 数据处理和统计分析
利用R 4.0.2中的vegan、FD软件包计算群落的物种和功能多样性。为了揭示黑松群落物种多样性和功能多样性的纬度梯度格局, 同时考虑到数据分布的非正态和齐性等问题, 利用广义线性回归模型拟合了物种多样性、功能多样性与样地纬度的关系。为了确定生物和非生物因子对黑松群落功能多样性的影响, 采用广义线性回归和方差分解分析了黑松优势度、物种多样性和气候因子对群落功能多样性的相对影响。其中, 方差分解用相对效应量化的参数估计值与所有参数之和的百分比表示。在每个模型中, 方差分为黑松优势度、物种多样性和气候3个类别。所有分析在R 4.0.2中完成。2 结果
2.1 黑松群落物种和功能多样性的纬度变异趋势
随纬度增大, 海岛黑松群落香农-维纳多样性和辛普森多样性无显著变化趋势(p> 0.05)(图2A、2B), 物种丰富度和黑松优势度显著降低(p< 0.05) (图2C、2D)。图2
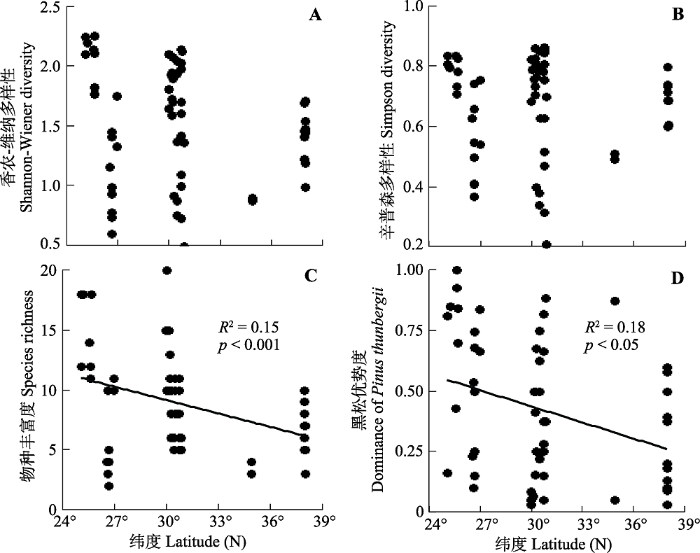 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2中国东部海岛黑松群落物种多样性沿纬度的变化。
Fig. 2Latitudinal variation in species diversity of Pinus thunbergii communities across sea-islands in Eastern China.
对于功能多样性, 枝叶性状总体和枝干的功能丰富度(图3A、3D)、离散度(图3B、3E)和Rao二次熵(图3C、3F)均随纬度增大显著下降(p< 0.05), 总体而言, 枝干功能多样性比总体性状功能多样性与纬度具有更高的拟合度(R2)。叶片功能多样性在纬度上无显著变化趋势(p> 0.05,图3G-3I)。
图3
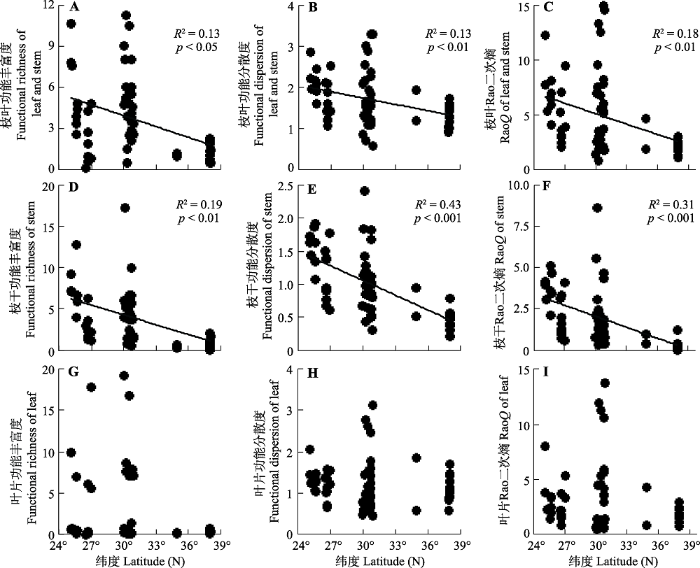 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3中国东部海岛黑松群落功能多样性沿纬度的变化。
Fig. 3Latitudinal variation in functional diversity of Pinus thunbergii communities across sea-islands in Eastern China.
2.2 生物和非生物因子对黑松群落功能多样性的相对影响
对于枝叶性状总体(图4A), 黑松优势度、物种多样性和气候综合解释了功能丰富度、分散度和RaoQ变异的63%、47%和39%。其中, 物种多样性(物种丰富度、辛普森和香农-维纳多样性)对功能丰富度、分散度和RaoQ的解释贡献率分别为53%、36%和31%。与之相比, 黑松优势度单独对功能多样性的解释度很低(<1%)。气候因子(总太阳辐射、年平均气温和干旱度)对功能丰富度、分散度和RaoQ的解释贡献率分别为8%、11%和7%。其中, 随干旱强度增加, 功能丰富度(5%)和RaoQ (2%)显著增大(p< 0.05)。图4
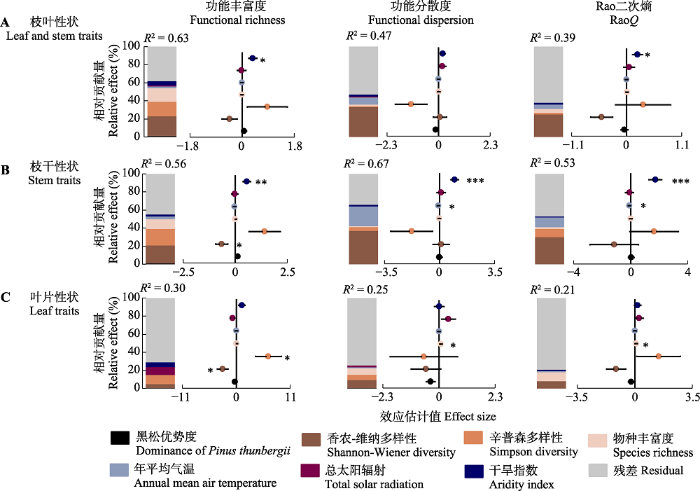 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4生物与非生物因子对中国东部海岛黑松群落功能多样性变异的相对影响。柱状图为各影响因子对功能多样性变异的相对贡献量, 其右侧坐标图指示各因子的影响效应量和方向(正或负)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 4Relative effect of abiotic and biotic factors on functional diversity variability of Pinus thunbergii communities across sea-islands in Eastern China. Histograms describe the relative contribution of different factors on functional diversity variability; coordinate graphs in right of histograms denote the effect size and direction (positive or negative) of different factors. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
对于枝干性状(图4B), 黑松优势度、物种多样性和气候综合解释了功能丰富度、分散度和RaoQ变异的56%、67%和53%。其中, 物种多样性对功能丰富度、分散度和RaoQ的解释度分别为32%、27%和22%; 气候因子对功能丰富度、分散度和RaoQ的解释贡献率分别为5%、24%和12%。干旱度显著增加功能多样性(p< 0.01), 年平均气温显著降低功能分散度(22%)和RaoQ (11%)(p< 0.05), 香农-维纳多样性减少功能丰富度(20%)(p< 0.05)。
对叶片性状而言(图4C), 黑松优势度、物种多样性和气候对功能多样性的综合解释度较低(21%-30%)。香农-维纳多样性显著降低功能丰富度(5%)(p< 0.05), 辛普森多样性显著增加功能丰富度(10%)(p< 0.05), 物种丰富度显著增加功能分散度(7%)和RaoQ (10%)(p< 0.05)。
3 讨论
对中国东部海岛黑松林这一特殊植物群落的研究发现, 物种丰富度、枝叶总体功能多样性、枝干功能多样性随纬度升高而显著下降, 这与全球大陆生态系统的变化格局基本一致(Clarke & Gaston, 2006)。与以往全球或生物群区水平上所揭示的生物多样性纬度地带性相比, 本研究所揭示的物种和功能多样性纬度梯度格局仅限于单一群落类型, 这在一定程度上说明了在纬度梯度上, 生物多样性的变化规律无论是在区域植被类型或复合群落水平, 还是在单一群落水平, 都是保持一致的。更重要的是, 本研究区属于海岛生态系统类型, 相对于大陆生态系统的连续性和广袤性, 海岛不仅与大陆保持隔离, 且面积不等和相互独立(刘翔宇等, 2019)。在这种特殊景观格局中, 同一植物群落类型的物种多样性和功能多样性也表现出了明显的纬度地带性特征。这进一步说明了生物多样性的纬度梯度地带性既不取决于生态系统或景观是否连续还是间断, 也不依赖于生态系统的分类等级高低(植被型或群落)。需要指出的是, 本研究中黑松群落的枝干功能多样性随纬度升高而显著下降, 但是叶片功能多样性并未随纬度升高而表现出显著的规律性。究其原因, 一方面是由于枝干性状比叶片性状更为保守, 而后者对生物竞争和环境过滤的响应更为敏感(Wright et al., 2001; Kang et al., 2014)。如果不同叶片性状对纬度梯度上生物和非生物环境变化的响应不一致, 那么不同方向的响应模式就会模糊多维性状功能多样性的纬度梯度规律(Butterfield & Suding, 2013)。另一方面, 本研究对象为以黑松为优势种的同一群落类型, 尽管表现出了一定的物种多样性和枝干功能多样性纬度梯度特征, 但黑松本身是针叶树种, 其叶性状的环境可塑性不高, 这也可能是导致其叶片功能多样性纬度地带性差的原因之一。以上两方面的原因也解释了, 在本研究中, 为什么随纬度升高, 枝叶性状总体比枝干性状单独的功能多样性变异更低, 这主要是枝叶性状总体功能多样性包含了叶性状。
在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie, 1991; Currie et al., 2004)。在温度(太阳辐射)较高的地区, 更多的能量供应可提高物种的繁衍能力, 降低物种灭绝率, 从而提高物种多样性(Schumm et al., 2019)。本研究显示, 黑松群落功能多样性的纬度梯度变化特征受到气候因子的显著影响, 年平均气温显著降低枝干功能多样性, 这意味着枝干功能多样性随环境的变化体现了植物群落物种在资源利用策略等方面的生态权衡, 以及对环境变化的响应(Garnier et al., 2004)。这表明在大尺度的温度等环境过滤作用下, 低纬度海岛的黑松群落物种具有更强的资源利用能力(de Bello et al., 2013)。本研究也发现, 干旱程度显著增加了黑松群落枝叶总体和枝干功能多样性。在纬度较高的海岛, 干旱程度高, 根据胁迫梯度假说, 植物多维性状的整合度高(Dwyer & Laughlin, 2017), 从而压缩功能或生态位空间(Stahl et al., 2014), 可维持的物种多样性也就较低(Marks et al., 2016)。
在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关。叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(图4)。这与其他研究结果一致, 即: 物种多样性的增大可提高群落功能多样性(Whittaker et al., 2014; Kumordzi et al., 2015)。群落功能多样性(功能空间)是以物种功能特征为基础的, 而群落内没有任何两个物种具有完全相同的功能特征(Díaz & Cabido,2001)。功能丰富度反映了群落内物种占据功能空间的大小, 功能分散度和RaoQ均体现了物种在功能性状空间分布的差异程度(Laliberté & Legendre,2010), 其值越高, 表明物种间的生态位互补程度越高(Díaz & Cabido,2001; Petchey & Gaston, 2002), 因而, 可以维持更高的物种多样性(Petchey & Gaston, 2006)。
海岛纬度梯度上黑松群落功能多样性的变化不是气候或物种多样性单独影响的结果, 而是两者的综合作用。其中, 气候因子、黑松优势度和物种多样性可以综合解释枝叶性状总体和枝干功能多样性变异的39%-67%, 但对叶片功能多样性的综合解释度较低。该结果反映了以下事实: 生物与非生物因素对海岛纬度梯度上黑松群落功能多样性变异的影响程度取决于植物性状类型。相对而言, 物种多样性对叶片功能多样性的影响更为显著, 而气候对枝干功能多样性影响更显著。从枝叶综合功能多样性看, 尽管物种多样性可以解释其纬度变异的31%-53%, 但其显著性不高, 这说明, 在区域大尺度, 气候因子(尤其是干旱)是影响群落功能多样性的关键因素。可以理解的是, 功能丰富度、功能分散度和RaoQ综合反映了环境过滤作用下群落共存物种间适应资源生态位的多样性(Laliberté & Legendre,2010; de Bello et al., 2013), 因此, 气候因子可以较好地解释其变异特征。需要提及的是, 气候、黑松优势度和物种多样性不能解释的功能多样性变异部分(即残差), 可能是不同黑松群落的人类干扰强度、群落层次结构和土壤因素等的差异造成的, 该部分需要后续进一步澄清。
值得注意的是, 本研究的回归分析显示, 物种香农-维纳多样性对叶片功能丰富度具有负向的影响, 但郑丽婷等(2018)发现功能丰富度与香农-维纳多样性正相关。造成不同结果的原因可能是由于香农-维纳多样性是对物种丰富度和均匀度两者的综合度量, 当一个群落既具有高的物种数又具有平均的个体数时, 在理想情况下, 则理应具有高的功能丰富度(Mason et al., 2005), 这也是本研究所反映的情形。相反, 当一个群落虽然具有高的物种数, 但物种的个体数并不均匀时, 例如本研究低纬度海岛的黑松群落, 其物种丰富度高, 黑松的优势度也高(图2), 则其并不能增加群落的功能丰富度。以上相反的分析结果提醒我们, 在研究群落功能多样性的影响机制时, 需要综合考虑物种多样性不同方面的叠加效应, 尤其需要将生物和非生物因素全部纳入分析框架, 才能比较客观地区分物种丰富度和均匀度两者的相对影响。
综上所述, 本研究揭示了中国东部海岛黑松群落功能多样性的纬度变异特征及其影响因素, 一定程度上确认了即使在隔离于大陆的海岛单一植被类型中, 物种和功能多样性仍然遵守传统的生物多样性纬度地带性格局。本研究有助于深入理解中国东部海岛植物多样性的生物地理格局和潜在的生态维持机制。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/jec.2012.101.issue-1URL [本文引用: 1]
DOI:10.1098/rspb.2006.3545URL [本文引用: 1]
DOI:10.1016/j.tree.2014.01.001URL [本文引用: 1]
DOI:10.1086/285144URL [本文引用: 1]
DOI:10.1111/ele.2004.7.issue-12URL [本文引用: 1]
DOI:10.1111/j.2006.0906-7590.04683.xURL [本文引用: 1]
DOI:10.1111/j.1600-0587.2012.07438.xURL [本文引用: 2]
DOI:10.1016/S0169-5347(01)02283-2URL [本文引用: 2]
DOI:10.1111/ele.12781PMID:28510261 [本文引用: 1]

Trade-offs maintain diversity and structure communities along environmental gradients. Theory indicates that if covariance among functional traits sets a limit on the number of viable trait combinations in a given environment, then communities with strong multidimensional trait constraints should exhibit low species diversity. We tested this prediction in winter annual plant assemblages along an aridity gradient using multilevel structural equation?modelling. Univariate and multivariate functional diversity measures were poorly explained by aridity, and were surprisingly poor predictors of community richness. By contrast, the covariance between maximum height and seed mass strengthened along the aridity gradient, and was strongly associated with richness declines. Community richness had a positive effect on local neighbourhood richness, indicating that climate effects on trait covariance indirectly influence diversity at local scales. We present clear empirical evidence that declines in species richness along gradients of environmental stress can be due to increasing constraints on multidimensional phenotypes.? 2017 John Wiley & Sons Ltd/CNRS.
DOI:10.1890/03-0799URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/jvs.12118URL [本文引用: 2]
[本文引用: 4]
[本文引用: 4]
DOI:10.1111/jec.2015.103.issue-6URL [本文引用: 2]
PMID:20380219 [本文引用: 3]
A new framework for measuring functional diversity (FD) from multiple traits has recently been proposed. This framework was mostly limited to quantitative traits without missing values and to situations in which there are more species than traits, although the authors had suggested a way to extend their framework to other trait types. The main purpose of this note is to further develop this suggestion. We describe a highly flexible distance-based framework to measure different facets of FD in multidimensional trait space from any distance or dissimilarity measure, any number of traits, and from different trait types (i.e., quantitative, semi-quantitative, and qualitative). This new approach allows for missing trait values and the weighting of individual traits. We also present a new multidimensional FD index, called functional dispersion (FDis), which is closely related to Rao's quadratic entropy. FDis is the multivariate analogue of the weighted mean absolute deviation (MAD), in which the weights are species relative abundances. For unweighted presence-absence data, FDis can be used for a formal statistical test of differences in FD. We provide the "FD" R language package to easily implement our distance-based FD framework.
DOI:10.3724/SP.J.1258.2011.01136URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2011.01136 [本文引用: 1]
通过在高寒矮嵩草(Kobresia humilis)草甸为期4年的刈割(留茬1 cm、3 cm及不刈割)、施肥(尿素7.5 g·m<sup>–2</sup>·a<sup>–1</sup> +磷酸二铵1.8 g·m<sup>–2</sup>·a<sup>–1</sup>、不施肥)和浇水(20.1 kg·m<sup>–2</sup>·a<sup>–1</sup>、不浇水)控制实验, 对该生态系统中功能多样性与物种多样性的变化及其与初级生产力关系进行了研究。结果表明: 刈割和施肥显著影响植物性状和生产力, 而浇水作用微弱。刈割对物种多样性无影响, 但对不同功能多样性指数的影响不同; 施肥降低了物种多样性和以所有性状(除植株高度外)量化的功能多样性; 浇水仅对物种丰富度有微弱影响。各功能多样性的平均值能更好地反映群落物种间的功能差异。功能多样性和物种多样性呈正相关或不相关, 且二者与初级生产力的关系可表现为正相关、负相关和不相关, 说明三者间的关系是性状依赖的, 并受生境资源状况和刈割扰动的影响。植物功能性状对群落初级生产力有更加直接的影响。在考虑生物多样性对生态系统功能的影响时, 应更加注重对植物功能属性变化的研究。
DOI:10.17520/biods.2018235URL [本文引用: 1]
DOI:10.17520/biods.2018235 [本文引用: 1]
beta多样性描述群落物种组成如何随环境梯度而变化。海岛具有边界清晰、面积和离岸距离不同以及环境变化剧烈等自然禀赋。目前, 我们对离岸距离、岛间距离和气候因素在海岛植物beta多样性变化格局中的相对贡献仍认识不足。本研究基于中国东部36个海岛的维管植物物种名录, 以Jaccard相异性指数度量beta多样性, 利用Mantel偏相关分析和beta多样性的变异分解, 探究了海岛不同生活型维管植物的beta多样性格局及其非生物影响因素。结果显示: 36个海岛共记录维管植物1,404种, 其中木本植物481种, 草本植物859种, 藤本植物64种。植物beta多样性随岛间距离和离岸距离差的增大而显著增加(P < 0.001); 海岛面积和气候要素对植物beta多样性无显著影响(P > 0.05)。岛间距离单独对beta多样性总变异的解释度为29.3%, 离岸距离独立解释了2.8%, 面积和气候共同解释了0.5%。木本植物与草本植物的beta多样性格局与总体一致, 距离因子对木本植物beta多样性的解释度高于草本植物(37.5% > 25.3%)。综上, 海岛植物beta多样性主要受岛间距离和离岸距离的影响, 反映了扩散限制是塑造中国东部海岛植物beta多样性格局的主要生态过程。
DOI:10.1111/brv.1965.40.issue-4URL [本文引用: 1]
DOI:10.1111/ele.2016.19.issue-7URL [本文引用: 1]
DOI:10.1111/oik.2005.111.issue-1URL [本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2007.0019 [本文引用: 1]
植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、 过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地 貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO<sub>2</sub>浓度升高 ) 对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国 的植物功能性状研究还需要更加明朗化和系统化。
DOI:10.1111/jbi.2017.44.issue-5URL [本文引用: 1]
DOI:10.1071/BT12225URL [本文引用: 1]
DOI:10.1046/j.1461-0248.2002.00339.xURL [本文引用: 1]
PMID:16706917 [本文引用: 1]
Functional diversity is a component of biodiversity that generally concerns the range of things that organisms do in communities and ecosystems. Here, we review how functional diversity can explain and predict the impact of organisms on ecosystems and thereby provide a mechanistic link between the two. Critical points in developing predictive measures of functional diversity are the choice of functional traits with which organisms are distinguished, how the diversity of that trait information is summarized into a measure of functional diversity, and that the measures of functional diversity are validated through quantitative analyses and experimental tests. There is a vast amount of trait information available for plant species and a substantial amount for animals. Choosing which traits to include in a particular measure of functional diversity will depend on the specific aims of a particular study. Quantitative methods for choosing traits and for assigning weighting to traits are being developed, but need much more work before we can be confident about trait choice. The number of ways of measuring functional diversity is growing rapidly. We divide them into four main groups. The first, the number of functional groups or types, has significant problems and researchers are more frequently using measures that do not require species to be grouped. Of these, some measure diversity by summarizing distances between species in trait space, some by estimating the size of the dendrogram required to describe the difference, and some include information about species' abundances. We show some new and important differences between these, as well as what they indicate about the responses of assemblages to loss of individuals. There is good experimental and analytical evidence that functional diversity can provide a link between organisms and ecosystems but greater validation of measures is required. We suggest that non-significant results have a range of alternate explanations that do not necessarily contradict positive effects of functional diversity. Finally, we suggest areas for development of techniques used to measure functional diversity, highlight some exciting questions that are being addressed using ideas about functional diversity, and suggest some directions for novel research.
DOI:10.1098/rspb.2019.0745URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/07-1206.1URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1218036111PMID:25225395 [本文引用: 2]
Analyses of species-diversity patterns of remote islands have been crucial to the development of biogeographic theory, yet little is known about corresponding patterns in functional traits on islands and how, for example, they may be affected by the introduction of exotic species. We collated trait data for spiders and beetles and used a functional diversity index (FRic) to test for nonrandomness in the contribution of endemic, other native (also combined as indigenous), and exotic species to functional-trait space across the nine islands of the Azores. In general, for both taxa and for each distributional category, functional diversity increases with species richness, which, in turn scales with island area. Null simulations support the hypothesis that each distributional group contributes to functional diversity in proportion to their species richness. Exotic spiders have added novel trait space to a greater degree than have exotic beetles, likely indicating greater impact of the reduction of immigration filters and/or differential historical losses of indigenous species. Analyses of species occurring in native-forest remnants provide limited indications of the operation of habitat filtering of exotics for three islands, but only for beetles. Although the general linear (not saturating) pattern of trait-space increase with richness of exotics suggests an ongoing process of functional enrichment and accommodation, further work is urgently needed to determine how estimates of extinction debt of indigenous species should be adjusted in the light of these findings.
DOI:10.1046/j.0269-8463.2001.00542.xURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1258.2011.00926URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2011.00926 [本文引用: 1]
沙质海岸空间梯度上环境差异较大, 黑松(Pinus thunbergii)在长期的适应过程中树冠结构变化也很大。为揭示黑松树冠结构与环境间的适应机制, 在山东省胶南市灵山湾国家森林公园距海岸线0–50、200–250和400–450 m梯度内各设置1个样带, 记为带I、带II、带III, 采用枝构型的理论和方法, 对黑松的分枝格局进行了研究, 并采用模拟自然风法测定了黑松枝条的抗风折能力。结果表明: 1)在海岸梯度上黑松分枝格局差异较大, 随着距离海岸越来越远, 黑松各级枝的分枝长度、总体分枝率均逐渐增大, 而枝径比和逐步分枝率逐渐减小, 各级分枝角度表现为带I >带III >带II。2)带I树冠背风面与迎风面相比, 分枝长度、分枝角度、分枝数量、枝条干枯率分别是迎风面的1.62、1.38、2.65和0.59倍, 随着距离海岸越来越远, 这种不对称性逐渐减弱, 至带III树冠基本对称。3)海风是影响带I分枝角度偏转、枝条干枯和冠型不对称现象的主要原因。4)带III枝条的抗风折能力高于带I, 且两个样带模拟风速与枝条所承受的拉力之间的关系均符合逻辑斯蒂方程, 相关系数R<sup>2</sup>均达0.97以上。该研究揭示了不同海岸梯度上黑松分枝格局的形成机制及其抗风折能力, 可为沿海黑松防护林的合理经营提供科学依据。
[本文引用: 1]
[本文引用: 1]
DOI:10.13287/j.1001-9332.201802.037PMID:29692046 [本文引用: 2]
Island vegetation plays an important role in biodiversity research across the world. The study of plant diversity in island is helpful for understanding the mechanism of plant diversity maintenance under land-sea interaction. Here, four typical plant communities (Quercus acutissima community, Robinia pseudoacacia community, Pinus thunbergii community and Vitex negundo community) in Miao Archipelago were selected to examine the species, functional and structural diversities and their responses to environmental factors at the community scale by using species diversity indices, functional diversity indices, as well as structural diversity indices. The results showed that the species richness and Rao index of P. thunbergii community was higher than that of Q. acutissima community and R. pseudoacacia community, but the structural diversity was lower. The species diversity and structural diversity of V. Negundo shrub were lower than that of forest community, but the functional diversity was higher than some forest communities. The relationship between the diversity of typical plant communities in island area illustrated a significant positive correlation between species richness with Rao index and tree height diversity, however the correlation with functional evenness was significantly negative. The structural diversity and functional evenness were determined by slope with negative and positive relationships, respectively. Functional heterogeneity, functional divergence and species diversity were affected largely by soil physical and chemical properties, displaying the positive relationship with soil bulk density and soil total carbon content, and a negative relationship with soil water content. In conclusion, diversity pattern of plant community in Miao Archipelago reflected not only the characteristics in mainland vegetation but also the special nature of the sea island.
PMID:29692046 [本文引用: 2]
Single-trait functional indices outperform multi-trait indices in linking environmental gradients and ecosystem services in a complex landscape
1
2013
... 需要指出的是, 本研究中黑松群落的枝干功能多样性随纬度升高而显著下降, 但是叶片功能多样性并未随纬度升高而表现出显著的规律性.究其原因, 一方面是由于枝干性状比叶片性状更为保守, 而后者对生物竞争和环境过滤的响应更为敏感(Wright et al.,
Climate, energy and diversity
1
2006
... 对中国东部海岛黑松林这一特殊植物群落的研究发现, 物种丰富度、枝叶总体功能多样性、枝干功能多样性随纬度升高而显著下降, 这与全球大陆生态系统的变化格局基本一致(Clarke & Gaston,
Climate change, sea-level rise, and conservation: keeping island biodiversity afloat
1
2014
... 海岛在维持生物多样性方面扮演着重要角色(Courchamp et al.,
Energy and large-scale patterns of animal- and plant-species richness
1
1991
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness
1
2004
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Variations in species and functional plant diversity along climatic and grazing gradients
1
2006
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
Hierarchical effects of environmental filters on the functional structure of plant communities: a case study in the French Alps
2
2013
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
... 海岛纬度梯度上黑松群落功能多样性的变化不是气候或物种多样性单独影响的结果, 而是两者的综合作用.其中, 气候因子、黑松优势度和物种多样性可以综合解释枝叶性状总体和枝干功能多样性变异的39%-67%, 但对叶片功能多样性的综合解释度较低.该结果反映了以下事实: 生物与非生物因素对海岛纬度梯度上黑松群落功能多样性变异的影响程度取决于植物性状类型.相对而言, 物种多样性对叶片功能多样性的影响更为显著, 而气候对枝干功能多样性影响更显著.从枝叶综合功能多样性看, 尽管物种多样性可以解释其纬度变异的31%-53%, 但其显著性不高, 这说明, 在区域大尺度, 气候因子(尤其是干旱)是影响群落功能多样性的关键因素.可以理解的是, 功能丰富度、功能分散度和RaoQ综合反映了环境过滤作用下群落共存物种间适应资源生态位的多样性(Laliberté & Legendre,
Vive la différence: plant functional diversity matters to ecosystem processes
2
2001
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
... ), 其值越高, 表明物种间的生态位互补程度越高(Díaz & Cabido,
Constraints on trait combinations explain climatic drivers of biodiversity: the importance of trait covariance in community assembly
1
2017
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Plant functional markers capture ecosystem properties during secondary succession
1
2004
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox
1
2002
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
山东半岛北部黑松海防林幼龄植株更新动态及其影响因素
1
2010
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
山东半岛北部黑松海防林幼龄植株更新动态及其影响因素
1
2010
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
Trait variability differs between leaf and wood tissues across ecological scales in subtropical forests
2
2014
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 需要指出的是, 本研究中黑松群落的枝干功能多样性随纬度升高而显著下降, 但是叶片功能多样性并未随纬度升高而表现出显著的规律性.究其原因, 一方面是由于枝干性状比叶片性状更为保守, 而后者对生物竞争和环境过滤的响应更为敏感(Wright et al.,
4
1999
... 2016-2019年的每年7-8月, 在对各海岛植被踏查后, 选取典型黑松群落设置样地, 森林样方大小为20 m × 20 m, 灌丛样方大小为10 m × 10 m.样地建立后, 记录和测量所有高度大于0.5 m的木本植物名称、基径、胸径、高度、冠幅、枝下高、叶下高和郁闭度等群落学特征, 以及地形、地貌和土壤类型等.此外, 对于难以攀登的悬崖上的黑松群落, 采用法瑞学派的无样地调查法记录物种组成、物种的多盖度和聚集度, 以及群落分层情况(高度和盖度)等.本研究共调查了21个海岛的52个黑松群落样地, 另外, 从《福建海岛植被》(孔繁昇等,
... *为孔繁昇等(
... * is the island investigated by Kong et al. (
... 需要说明的是, 对于摘录自孔繁昇等(
4
1999
... 2016-2019年的每年7-8月, 在对各海岛植被踏查后, 选取典型黑松群落设置样地, 森林样方大小为20 m × 20 m, 灌丛样方大小为10 m × 10 m.样地建立后, 记录和测量所有高度大于0.5 m的木本植物名称、基径、胸径、高度、冠幅、枝下高、叶下高和郁闭度等群落学特征, 以及地形、地貌和土壤类型等.此外, 对于难以攀登的悬崖上的黑松群落, 采用法瑞学派的无样地调查法记录物种组成、物种的多盖度和聚集度, 以及群落分层情况(高度和盖度)等.本研究共调查了21个海岛的52个黑松群落样地, 另外, 从《福建海岛植被》(孔繁昇等,
... *为孔繁昇等(
... * is the island investigated by Kong et al. (
... 需要说明的是, 对于摘录自孔繁昇等(
Linkage of plant trait space to successional age and species richness in boreal forest understorey vegetation
2
2015
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
A distance-based framework for measuring functional diversity from multiple traits
3
2010
... 本研究采用多维性状计算各群落的功能丰富度、功能分散度(Laliberté & Legengdre,
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
... 海岛纬度梯度上黑松群落功能多样性的变化不是气候或物种多样性单独影响的结果, 而是两者的综合作用.其中, 气候因子、黑松优势度和物种多样性可以综合解释枝叶性状总体和枝干功能多样性变异的39%-67%, 但对叶片功能多样性的综合解释度较低.该结果反映了以下事实: 生物与非生物因素对海岛纬度梯度上黑松群落功能多样性变异的影响程度取决于植物性状类型.相对而言, 物种多样性对叶片功能多样性的影响更为显著, 而气候对枝干功能多样性影响更显著.从枝叶综合功能多样性看, 尽管物种多样性可以解释其纬度变异的31%-53%, 但其显著性不高, 这说明, 在区域大尺度, 气候因子(尤其是干旱)是影响群落功能多样性的关键因素.可以理解的是, 功能丰富度、功能分散度和RaoQ综合反映了环境过滤作用下群落共存物种间适应资源生态位的多样性(Laliberté & Legendre,
刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响
1
2011
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响
1
2011
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
中国东部海岛维管植物的beta多样性及其驱动因素
1
2019
... 对中国东部海岛黑松林这一特殊植物群落的研究发现, 物种丰富度、枝叶总体功能多样性、枝干功能多样性随纬度升高而显著下降, 这与全球大陆生态系统的变化格局基本一致(Clarke & Gaston,
中国东部海岛维管植物的beta多样性及其驱动因素
1
2019
... 对中国东部海岛黑松林这一特殊植物群落的研究发现, 物种丰富度、枝叶总体功能多样性、枝干功能多样性随纬度升高而显著下降, 这与全球大陆生态系统的变化格局基本一致(Clarke & Gaston,
Patterns of species diversity
1
1965
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
Tree diversity, tree height and environmental harshness in eastern and western North America
1
2016
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Functional richness, functional evenness and functional divergence: the primary components of functional diversity
1
2005
... 值得注意的是, 本研究的回归分析显示, 物种香农-维纳多样性对叶片功能丰富度具有负向的影响, 但郑丽婷等(
植物功能性状与环境和生态系统功能
1
2007
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
植物功能性状与环境和生态系统功能
1
2007
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
A roadmap for island biology: 50 fundamental questions after 50 years of The Theory of Island Biogeography
1
2017
... 海岛在维持生物多样性方面扮演着重要角色(Courchamp et al.,
New handbook for standardised measurement of plant functional traits worldwide
1
2013
... 与黑松群落学特征调查同步, 参照Pérez- Harguindeguy等(
Functional diversity (FD), species richness and community composition
1
2002
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
Functional diversity: back to basics and looking forward
1
2006
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
Common latitudinal gradients in functional richness and functional evenness across marine and terrestrial systems
2
2019
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
Predicting species’ range limits from functional traits for the tree flora of North America
1
2014
... 在众多解释物种多样性大尺度格局的生态机制中, “能量假说”认为, 优越的水热条件适合更多物种的生存(Currie,
中国东部海岛森林和灌丛土壤碳氮磷养分库的纬度变化
1
2019
... 海岛在维持生物多样性方面扮演着重要角色(Courchamp et al.,
中国东部海岛森林和灌丛土壤碳氮磷养分库的纬度变化
1
2019
... 海岛在维持生物多样性方面扮演着重要角色(Courchamp et al.,
New multidimensional functional diversity indices for a multifaceted framework in functional ecology
1
2008
... 本研究采用多维性状计算各群落的功能丰富度、功能分散度(Laliberté & Legengdre,
浙江舟山岛松材线虫入侵后松林群落的自然演替和特征
1
2011
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
浙江舟山岛松材线虫入侵后松林群落的自然演替和特征
1
2011
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
Functional biogeography of oceanic islands and the scaling of functional diversity in the Azores
2
2014
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 在中国东部海岛纬度梯度上, 黑松群落的功能多样性变化特征除与大范围的气候差异有关, 也与群落本身的物种多样性变化有关.叶片功能多样性与物种多样性显著关联, 辛普森多样性和物种丰富度的增加, 导致了叶片功能丰富度、分散度和RaoQ的升高(
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats
2
2001
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 需要指出的是, 本研究中黑松群落的枝干功能多样性随纬度升高而显著下降, 但是叶片功能多样性并未随纬度升高而表现出显著的规律性.究其原因, 一方面是由于枝干性状比叶片性状更为保守, 而后者对生物竞争和环境过滤的响应更为敏感(Wright et al.,
沿海黑松防护林低产、低质、低效成因的调查报告
1
2003
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
沿海黑松防护林低产、低质、低效成因的调查报告
1
2003
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
沙质海岸黑松分枝格局特征及其抗风折能力分析
1
2011
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
沙质海岸黑松分枝格局特征及其抗风折能力分析
1
2011
... 黑松(Pinus thunbergii)具有优良的抗风、耐盐碱特性, 适合在土壤养分贫瘠的海岛生长, 对温度的适应范围广(张丹等,
1
2011
... 选用物种丰富度、香农-维纳指数和辛普森指数反映黑松群落的物种多样性情况, 具体计算方法参见张金屯(
1
2011
... 选用物种丰富度、香农-维纳指数和辛普森指数反映黑松群落的物种多样性情况, 具体计算方法参见张金屯(
庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应
2
2018
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 值得注意的是, 本研究的回归分析显示, 物种香农-维纳多样性对叶片功能丰富度具有负向的影响, 但郑丽婷等(
庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应
2
2018
... 大陆纬度梯度上植物功能多样性变化特征的研究表明, 在非生物因素的作用下, 随纬度上升, 植物群落功能丰富度降低, 功能均匀度增加(Schumm et al.,
... 值得注意的是, 本研究的回归分析显示, 物种香农-维纳多样性对叶片功能丰富度具有负向的影响, 但郑丽婷等(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}