黎松松
1 , 王宁欣
1 , 郑伟
, , 1 , 2 , * , 朱亚琼
1 , 王祥
1 , 马军
1 , 朱进忠
1 , 2 1 新疆农业大学草业与环境科学学院, 乌鲁木齐 8300522 新疆维吾尔自治区草地资源与生态重点实验室, 乌鲁木齐 830052Comparison of transgressive overyielding effect and plant diversity effects of annual and perennial legume-grass mixtures Song-Song LI
1 , Ning-Xin WANG
1 , Wei ZHENG
, , 1 , 2 , * , Ya-Qiong ZHU
1 , Xiang WANG
1 , Jun MA
1 , Jin-Zhong ZHU
1 , 2 1 College of Pratacultural and Environmental Science, Xinjiang Agricultural University, ürümqi 830052, China2 Xinjiang Key Laboratory of Grassland Resources and Ecology, ürümqi 830052, China 通讯作者: *郑伟: ORCID: 0000-0002-5627-9042(
zw065@126.com )
编委: 梁存柱
责任编辑: 赵航
收稿日期: 2020-02-24
接受日期: 2020-09-27
网络出版日期: 2021-01-20
基金资助: 新疆维吾尔自治区高校科研计划重点项目 (XJEDU2019Z18 )农业农村部“国家牧草现代产业技术体系”项目 (CARS-34 )国家自然科学基金 (31660692 )国家重点研发计划 (2017YFE0109200 )
Corresponding authors: *(
zw065@126.com )
Received: 2020-02-24
Accepted: 2020-09-27
Online: 2021-01-20
Fund supported: Key Project of University Research in Xinjiang Uygur Autonomous Region (XJEDU2019Z18 )“National Modern Forage and Grass Research System” Program of the Ministry of Agriculture and Rural Affairs (CARS-34 )National Natural Science Foundation of China (31660692 )National Key R&D Program of China (2017YFE0109200 )
摘要 为了测度一年生和多年生豆禾混播草地的超产效应与植物多样性效应的关系, 明确一年生和多年生混播草地的高产优势, 探索豆禾混播草地多样性效应和超产效应对其生态功能的响应机制, 于2013-2015年在新疆伊犁地区昭苏盆地开展了3年的牧草产量观测试验。该试验设置3种牧草混播种类和混播比例, 分别为一年生豆禾混播草地(2种牧草混播, AM2)和多年生豆禾混播草地(2种牧草混播, PM2; 4种牧草混播, PM4; 6种牧草混播, PM6), 豆禾混播比例分别为6:4、5:5和4:6。结果表明: 1) 2013、2014年和3年平均值AM2的超产幅度小于PM2和PM6, 2015年AM2的超产幅度大于PM2、PM4和PM6; 混播群落生产力与群落组分中生产力最高产物种单产以及各组分种平均单产的差值表现出相似的规律。2) 2013、2014年和3年平均值AM2的互补效应大于PM2、PM4和PM6, AM2的选择效应则远小于互补效应, PM2、PM4和PM6的互补效应则比较稳定。3)物种丰富度和物种均匀度与牧草产量(群落生产力)大部分情况下呈单峰的“饱和上升型”模式, 分别在4种牧草混播和豆禾混播比例为5:5时, 具有较高生产力。4)多年生豆禾混播草地的互补效应、选择效应和多样性净效应均随生长年限的延长而呈下降趋势, 也导致了超产幅度、超产效应及其稳定性的下降。由此可见, 在建植初期, 互补效应和选择效应共同主导了多年生豆禾混播草地的超产效应, 而随着生长年限的延长, 选择效应则成为主要影响因素; 一年生豆禾混播草地的超产效应则一直受互补效应的影响。
关键词: 物种丰富度 ;
物种均匀度 ;
超产效应 ;
互补效应 ;
选择效应 ;
多样性净效应 ;
生长年限 Abstract Aims Our objective was to explore mechanisms underlying the effects of transgressive overyielding and plant diversity effects associated with legume-grass mixtures in Zhaosu Basin, Yili Prefecture, Xinjiang Uygur Autonomous Region, China.
Methods Replicate plots (
n = 3) were sown in 2013 as legume and grass monocultures (8 in total, including
Vicia sativa, Avena sativa, Onobrychis viciaefolia, Bromus inermis, Trifolium pratense, Phleum pratense, Medicage sativa, Dactylis glomerata ), a two-species mixture (AM2, annual legume and grass; PM2, perennial legume and grass), a four-species mixture (PM4, 2 perennial legumes and 2 grasses), a six-species mixture (PM6, 3 perennial legumes and 3 grasses), and three mixed-ratio combinations (legume : grass = 6:4, 5:5 and 4:6). Total plot productivity was measured as forage yield (dry matter) each summer between 2013 to 2015, which were used to calculate transgressive overyielding effect (
OV ), transgressive overyielding effect 1 (
OV 1 ), and transgressive overyielding effect 2 (
OV 2 ). Plant diversity effects were determined by calculating the complementary effect (
CE ), selection effect (
SEF ), and net biodiversity effects (?
Y ).
Important findings The
OV amplitudes of AM2 in 2013, 2014 and 3 years average were less than those of PM2 and PM6. The
OV amplitude of AM2 in 2015 was greater than those of PM2, PM4 and PM6. The difference between the productivity of the mixed community and the yield of the most productive product species in the community components and the average yield of each species showed similar laws
. CE of AM2 in 2013, 2014, and 3 years average was greater than PM2, PM4 and PM6 in the respective years. While
SEF were much smaller than
CE in AM2, changes in
CE were relatively stable in PM2, PM4 and PM6. Fitted curves of species richness, species evenness, and forage yield (community productivity) predominantly showed a single maximum in PM4, yet productivity was highest in the 5:5 ratio plot.
CE, SEF, and ?
Y in perennial legume-grass mixtures decreased over time, which also led to declines in
OV amplitude,
OV 1 and
OV 2 , and their stability. Thus, in the initial study year,
CE and
SEF jointly dominated
OV, OV 1 and
OV 2 in perennial legume-grass mixtures. Over the next two growing years,
SEF became the main factor that influenced
OV, OV 1 and
OV 2 in perennial legume-grass mixtures.
OV, OV 1 and
OV 2 of annual legume-grass mixtures were all influenced by
CE from 2013 to 2015.
Keywords: species richness ;
species evenness ;
transgressive overyielding effect ;
complementarity effect ;
selection effect ;
net effect of biodiversity ;
growth years PDF (1497KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 黎松松, 王宁欣, 郑伟, 朱亚琼, 王祥, 马军, 朱进忠. 一年生和多年生豆禾混播草地超产与多样性效应的比较. 植物生态学报, 2021, 45(1): 23-37. DOI:
10.17521/cjpe.2020.0048 LI Song-Song, WANG Ning-Xin, ZHENG Wei, ZHU Ya-Qiong, WANG Xiang, MA Jun, ZHU Jin-Zhong.
Comparison of transgressive overyielding effect and plant diversity effects of annual and perennial legume-grass mixtures .
Chinese Journal of Plant Ecology , 2021, 45(1): 23-37. DOI:
10.17521/cjpe.2020.0048 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 )。与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 )。这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道。依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 )。超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 )。因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 )。
植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择。Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 )。Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势。而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱。在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 )。Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 )。由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义。
在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 )。多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围。但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 )。因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义。
本研究以新疆昭苏地区的一年生和多年生豆禾混作草地为研究对象, 在2013-2015年分别测度了一年生与多年生豆禾混作草地的牧草产量, 计算了二者的超产效应, 从群落层面(超产)和物种层面(超产效应)解析产生超产效应的原因, 并进一步对比分析选择效应、互补效应对超产效应的贡献。根据群落构建理论、植物多样性对生态功能作用的时间机制, 试图检验以下假设: (1)豆禾混作系统的超产和超产效应是互补效应和选择效应联合主导的; (2)多年生豆禾混作系统的互补效应随建植时间的延长而下降, 影响植物多样性-生态功能的正效应; (3) 3年中每年的超产效应均是一年生豆禾混作大于多年生豆禾混作系统。
1 材料和方法 1.1 试验地概况 试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 42.63°- 43.25° N), 属温带大陆性半干旱气候。年平均气温2.7 ℃, ≥10 ℃积温1 416.8 ℃, 无霜期85-100天, 年降水量512 mm, 降水集中于5-8月, 约占全年总降水量的68%, 年蒸发量1 261.6 mm, 是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm。试验小区为2013年开垦的多年生人工草地, 该地段2013年以前的植被类型为天然草地(山地草甸)。群落主要优势种有: 鸭茅(
Dactylis glomerata )、无芒雀麦(
Bromus inermis )、新疆披碱草(
Elymus sinkiangensis )、猫尾草(
Phleum pratense )、草原糙苏(
Phlomis pratensis )、草原老鹳草(
Geranium pratense )、大叶橐吾(
Ligularia macrophylla )等。盖度65%-100%, 草层高25-85 cm, 鲜草产量103 905 kg·hm
-2 。土壤为黑钙土, 土壤有机质含量为13.63%-14.89%, 全氮、全磷、全钾含量分别为9.53、2.16、12.20 g·kg
-1 , 碱解氮、有效磷、有效钾含量分别为459.20、144.43、473.58 mg·kg
-1 。
1.2 试验材料与设计 本研究的供试材料为燕麦(
Avena sativa )、救荒野豌豆(
Vicia sativa )、红车轴草(
Trifolium pratense )、驴食草(又名红豆草)(
Onobrychis viciifolia )、紫花苜蓿(
Medicage sativa )、无芒雀麦、鸭茅和猫尾草。一年生豆禾混播草地试验为以豆禾混播比例为主的随机区组设计, 豆禾比为6:4、5:5和4:6, 燕麦和救荒野豌豆单播作为对照(CK), 共5个处理, 重复3次, 共15个小区。多年生豆禾混播草地试验为豆禾混播比例和混播种类+搭配为主的随机区组设计, a因素为混播种类, 分别设2种牧草混播处理4个, 4种、6种牧草混播处理各1个; b因素为豆禾混播比例, 分别设6:4、5:5、4:6 (如出现豆科或禾本科多物种混播时, 该科各物种混播比例相同), 单播作为对照(CK), 共24个处理, 重复3次, 共63个小区。燕麦单播的播种量为180 kg·hm
-2 , 救荒野豌豆单播的播量为120 kg·hm
-2 ; 紫花苜蓿、红车轴草、猫尾草单播的播种量为15 kg·hm
-2 , 鸭茅、无芒雀麦单播的播种量为30 kg·hm
-2 , 驴食草单播的播量为60 kg·hm
-2 ; 混播比例按种子占单播重量的实际用价(种子纯净度与发芽率的乘积)来计算, 混播与单播密度相同。每小区3 m × 5 m, 条播, 行距15 cm, 播种深度2 cm, 播后覆土压实。前茬作物为春小麦, 整个生育期均不施肥, 不灌溉; 除杂草3次, 分别在返青后2周或出苗后2周、第1次除草20天后和刈割前1周。
1.3 取样时间和样品处理 一年生豆禾混播草地牧草地上生物量测定时间为每年的7月30日左右, 多年生豆禾混播草地牧草地上生物量测定时间为每年的7月15日左右。在每小区的对角线取5个1 m × 1 m的样方, 齐地面刈割, 65 ℃下恒温烘干至恒质量, 分种称量。以牧草地上生物量表征群落产量。
1.4 超产、超产效应和多样性效应的测度方法 超产(
OV )为混播群落生产力与该群落组分种单播生产力或该群落组分其他混播组合的生产力的差值。
(1) OV = B ploy - B mno 式中,
Bploy 为混播群落产量,
Bmno 为混播群落组各分种中的平均单产或该群落组分其他混播组合的产量,
OV > 0, 表明存在超产。
超产效应1 (
OV 1 )为混播群落生产力超过该群落组分种中生产力最高产物种的单产。
(2) OV 1 = (B ploy - B imno -max )/B imno -max 式中,
Bimno -max 为混播群落组分种中最高产物种的单产;
OV 1 > 0, 表明存在超产效应。
超产效应2 (
OV 2 )为混播群落生产力超过该群落内各组分种的平均单产。
(3) OV 2 = (B ploy - B mno -mean )/B mno -mean 式中,
Bmno -mean 为混播群落中各组分种的平均单产;
OV 2 > 0, 表明存在超产效应。
OV 2 解释了混播群落生产力与混播组分单播生产力之间的关系, 而
OV 1 强调了混播物种成分的差异与超产效应之间的联系(Siebenk?s
et al. ,
2016 )。
按照Loreau和Hector (
2001 )的方法, 将混播群落中生产力的增加量(多样性的净效应)分离出选择效应和互补效应。
?
Y =
Y O -
Y E =
N mean (?
RY ×
M ) +
(3) N cov (?RY, M) 式中, ?
Y 为多样性净效应,
Y O 为混播群落的实际产量,
Y E 为以单播产量为基础计算出的混播群落期望产量,
M 为混播群落中各物种单产的平均产量, ?
RY 为混播群落中各物种相对产量的变化量(各物种的实际相对产量与其期望相对产量之差),
N 为混播群落的物种数,
N mean (?
RY ×
M )为互补效应值(
CE ),
N cov (?
RY ,
M )为选择效应值(
SEF )。当?
Y = 0, 植物多样性对生产力无影响(零假设), 当?
Y > 0时, 表示群落高产受物种生态位互补作用影响, ?
Y < 0时, 表示群落中多物种竞争导致群落减产。
Table 1 表1 表1 豆禾混播草地的混播比例及各组分播量
Table 1
Mixed sowing ratios and sowing quantity of legume-grass mixtures 处理编号 混播比例 豆科牧草 Legume 禾本科牧草 Grass 物种 分播量 -2 ) 物种 分播量 -2 ) AM2-1 6:4 救荒野豌豆 Vicia sativa 72.00 燕麦 Avena sativa 60.00 AM2-2 5:5 救荒野豌豆 V. sativa 60.00 燕麦 A. sativa 75.00 AM2-3 4:6 救荒野豌豆 V. sativa 48.00 燕麦 A. sativa 90.00 PM2-A1 6:4 驴食草 Onobrychis viciaefolia 36.00 无芒雀麦 Bromus inermis 12.00 PM2-A2 5:5 驴食草 O. viciifolia 30.00 无芒雀麦 B. inermis 15.00 PM2-A3 4:6 驴食草 O. viciifolia 24.00 无芒雀麦 B. inermis 18.00 PM2-B1 6:4 驴食草 O. viciifolia 36.00 猫尾草 Phleum pratense 6.00 PM2-B2 5:5 驴食草 O. viciifolia 30.00 猫尾草P. pratense 7.50 PM2-B3 4:6 驴食草 O. viciifolia 24.00 猫尾草P. pratense 9.00 PM2-C1 6:4 红车轴草 Trifolium pratense 9.00 无芒雀麦 B. inermis 12.00 PM2-C2 5:5 红车轴草 T. pratense 7.50 无芒雀麦 B. inermis 15.00 PM2-C3 4:6 红车轴草 T. pratense 6.00 无芒雀麦 B. inermis 18.00 PM2-D1 6:4 红车轴草 T. pratense 9.00 猫尾草 P. pratense 6.00 PM2-D2 5:5 红车轴草 T. pratense 7.50 猫尾草P. pratense 7.50 PM2-D3 4:6 红车轴草 T. pratense 6.00 猫尾草P. pratense 9.00 PM4-1 6:4 驴食草 O. viciifolia 18.00 无芒雀麦B. inermis 6.00 红车轴草 T. pratense 4.50 猫尾草P. pratense 3.00 PM4-2 5:5 驴食草O. viciifolia 4.50 无芒雀麦B. inermis 7.50 红车轴草T. pratense 3.75 猫尾草P. pratense 3.75 PM4-3 4:6 驴食草O. viciifolia 12.00 无芒雀麦B. inermis 9.00 红车轴草T. pratense 3.00 猫尾草P. pratense 4.50 PM6-1 6:4 驴食草O. viciifolia 12.00 无芒雀麦B. inermis 4.00 红车轴草 T. pratense 3.00 猫尾草P. pretense 2.00 紫花苜蓿 Medicage sativa 3.00 鸭茅 Dactylis glomerata 2.00 PM6-2 5:5 驴食草O. viciifolia 10.00 无芒雀麦B. inermis 5.00 红车轴草 T. pratense 2.00 猫尾草P. pretense 2.50 紫花苜蓿M. sativa 2.00 鸭茅D. glomerata 2.50 PM6-3 4:6 驴食草O. viciifolia 8.00 无芒雀麦B. inermis 6.00 红车轴草 T. pratense 2.50 猫尾草P. pretense 3.00 紫花苜蓿 M. sativa 2.50 鸭茅D. glomerata 3.00
表中出现豆科或禾本科多物种混播时, 该科各物种混播比例相同。 When multiple species of Leguminosae or Gramineae are mixed in the table, the proportion of mixed sowing of species in this family is the same. 新窗口打开 |
下载CSV 1.5 数据处理 通过Microsoft Excel 2010软件对数据进行初步整理和常规计算, 再利用SPSS 19.0软件进行统计分析,即用一般线性模型分析对数据进行双因素方差分析, 统计完成后用Origin 8.0制图。采用q检验(SNK)法和最小差异显著(
LSD )法对产量、超产效应、多样性效应进行差异显著性检验。
2 结果和分析 2.1 一年生和多年生豆禾混播草地牧草产量、超产和超产效应的比较 由
表2 可知, 从不同年度来看, 2013、2014年AM2产量均显著高于多年生混播草地(
p < 0.05), PM2-C和PM2-D产量显著低于除PM2-B外其他多年生混播草地(
p < 0.05); 与单播相比, 2013年PM2-A、PM2-B、PM4和PM6增产较高, 达0.28-0.42 kg·m
-2 , 2014年PM2-A、PM2-B、PM4增产较高, 达0.17-0.28 kg·m
-2 ; 与PM2相比, 2013年PM4和PM6增产分别为0.42和0.32 kg·m
-2 , 2014年的则分别为0.29和0.23 kg·m
-2 ; 与PM4相比, 2013年PM6增产为0.32 kg·m
-2 , 2014年的则为0.22 kg·m
-2 。2015年AM2和PM2-D产量显著高于其他多年生混播草地(
p < 0.05); 与单播相比, AM2增产较高, 达0.16 kg·m
-2 ; 与PM2和PM4相比, 均未产生增产。从3年平均值来看, AM2产量显著高于多年生混播草地(
p < 0.05), PM2-C和PM2-D产量显著低于其他多年生混播草地(
p < 0.05); 与单播相比, PM2-A、PM2-B、PM6增产较高, 达0.14-0.19 kg·m
-2 ; 与PM2相比, PM4和PM6增产分别为0.22和0.20 kg·m
-2 , 与PM4相比, PM6增产不明显。各年份及3年平均值PM2-D豆禾产量比显著高于其他混播草地(
p < 0.05), 其他混播草地的豆禾产量比均小于40%。
Table 2 表2 表2 不同混播种类下牧草产量、超产和豆科牧草产量比的比较(平均值±标准误)
Table 2
Comparison of forage yield, overyielding and legume yield proportion under different mixed patterns (mean ± SE) 年份 处理编号 产量 Yield (kg·m-2 ) 超产 Overyielding (kg·m-2 ) 超产效应1 OV 1 ) 超产效应2 OV 2 ) 豆科产量比 混播处理 单播对照 与单播相比 与PM2相比 与PM4相比 2013 AM2 1.26 ± 0.11 a 1.22 ± 0.37 0.04 0.07 ± 0.14 b 0.08 ± 0.05 b 12.33 ± 1.52 c PM2-A 0.70 ± 0.06 b 0.39 ± 0.12 0.28 0.77 ± 0.13 a 0.92 ± 0.24 a 21.67 ± 5.40 b PM2-B 0.68 ± 0.12 b 0.40 ± 0.15 0.28 0.15 ± 0.09 b 0.62 ± 0.21 ab 27.03 ± 17.02 b PM2-C 0.28 ± 0.07 c 0.22 ± 0.06 0.06 -0.09 ± 0.04 c -0.31 ± 0.17 c 34.00 ± 5.53 b PM2-D 0.28 ± 0.05 c 0.24 ± 0.13 0.04 -0.13 ± 0.11 c 0.08 ± 0.03 b 81.67 ± 3.47 a PM4 0.73 ± 0.07 b 0.31 ± 0.06 0.42 0.42 0.86 ± 0.15 a 1.54 ± 0.35 a 29.33 ± 3.79 b PM6 0.63 ± 0.10 b 0.28 ± 0.10 0.35 0.32 0.32 0.62 ± 0.15 a 1.30 ± 0.24 a 32.33 ± 18.13 b 2014 AM2 0.75 ± 0.05 a 0.79 ± 0.33 -0.04 -0.15 ± 0.06 c -0.06 ± 0.06 cd 32.33 ± 1.53 b PM2-A 0.33 ± 0.01 bc 0.14 ± 0.02 0.19 0.55 ± 0.17 b 0.19 ± 0.06 b 10.00 ± 9.64 d PM2-B 0.26 ± 0.04 cd 0.09 ± 0.03 0.17 0.23 ± 0.13 b 0.07 ± 0.03 cd 21.33 ± 11.59 c PM2-C 0.19 ± 0.01 d 0.16 ± 0.11 0.03 -0.02 ± 0.01 c -0.15 ± 0.10 d 15.67 ± 5.13 cd PM2-D 0.15 ± 0.03 d 0.10 ± 0.07 0.05 0.30 ± 0.12 b 0.10 ± 0.02 b 52.33 ± 15.57 a PM4 0.41 ± 0.03 b 0.13 ± 0.06 0.28 0.18 1.01 ± 0.17 a 0.36 ± 0.03 a 13.67 ± 6.43 cd PM6 0.35 ± 0.04 bc 0.32 ± 0.21 0.03 0.12 0.22 -0.76 ± 0.02 d -0.07 ± 0.04 cd 12.00 ± 6.56 cd 2015 AM2 0.36 ± 0.03 a 0.21 ± 0.10 0.16 0.17 ± 0.14 a 0.80 ± 0.17 a 29.33 ± 8.33 c PM2-A 0.25 ± 0.04 c 0.27 ± 0.07 -0.02 0.01 ± 0.09 ab 0.64 ± 0.18 ab 4.33 ± 1.15 d PM2-B 0.29 ± 0.02 b 0.39 ± 0.16 -0.10 -0.04 ± 0.06 b 0.25 ± 0.11 b 10.67 ± 0.58 d PM2-C 0.33 ± 0.03 ab 0.37 ± 0.15 -0.04 -0.30 ± 0.12 d -0.08 ± 0.04 c 8.33 ± 2.31 d PM2-D 0.36 ± 0.01 a 0.31 ± 0.13 0.05 -0.06 ± 0.13 b -0.18 ± 0.11 c 80.67 ± 4.04 a PM4 0.25 ± 0.03 b 0.87 ± 0.29 -0.62 -0.08 -0.31 ± 0.06 d -0.07 ± 0.06 c 42.26 ± 4.62 b PM6 0.31 ± 0.01 b 0.27 ± 0.14 0.04 -0.02 -0.56 -0.21 ± 0.06 bc 0.29 ± 0.06 bc 21.67 ± 2.31 c 3年平均值 AM2 0.80 ± 0.25 a 0.74 ± 0.34 0.06 0.03 ± 0.07 b 0.27 ± 0.10 a 21.33 ± 2.08 b PM2-A 0.46 ± 0.10 b 0.27 ± 0.08 0.19 0.44 ± 0.10 a 0.58 ± 0.12 a 21.00 ± 10.44 b PM2-B 0.43 ± 0.07 b 0.29 ± 0.17 0.14 0.10 ± 0.23 ab 0.15 ± 0.20 ab 23.33 ± 10.41 b PM2-C 0.26 ± 0.03 c 0.25 ± 0.12 0.01 0.04 ± 0.10 b -0.20 ± 0.10 b 19.33 ± 1.53 b PM2-D 0.23 ± 0.05 c 0.22 ± 0.09 0.01 -0.01 ± 0.08 b -0.21 ± 0.24 b 74.67 ± 4.04 a PM4 0.48 ± 0.01 b 0.44 ± 0.12 0.04 0.22 0.52 ± 0.14 a 0.61 ± 0.18 a 29.67 ± 2.89 b PM6 0.46 ± 0.04 b 0.29 ± 0.13 0.17 0.12 0.02 -0.09 ± 0.12 b 0.51 ± 0.14 a 28.00 ± 11.53 b
相同年份同列不同小写字母表示差异显著(p < 0.05)。处理编号同 表1 。 Different lowercase letters within the same column of the same year indicate significant differences (p < 0.05). Treatment number see Table 1 . 新窗口打开 |
下载CSV 从混播种类来看, 2013年AM2的
OV 1 显著低于PM2-A、PM4和PM6 (
p < 0.05); 2014年PM4的
OV 1 显著高于其他混播种类(
p < 0.05), AM2和PM6的
OV 1 均为负值; 2015年AM2的
OV 1 显著高于PM4和PM6 (
p < 0.05); 从3年平均值来看, PM2-A、PM4的
OV 1 显著高于其他混播种类(
p < 0.05)。从混播比例来看, 各混播比例间
OV 1 在各年份及3年平均值间无显著差异(
p > 0.05); 随着建植年份的增加, 各混播比例
OV 1 呈下降趋势。从各混播组合来说, PM2和PM4各组合的
OV 1 显著高于其他混播(
p < 0.05)。从混播种类来看, 2013年AM2的
OV 2 显著低于PM2-A、PM4和PM6 (
p < 0.05); 2014年AM2和PM6的
OV 2 显著低于PM2-A、PM4 (
p < 0.05), 且均为负值, PM4的
OV 2 显著高于PM2-A (
p < 0.05); 2015年AM2的
OV 2 显著高于PM4和PM6 (
p < 0.05); 从3年平均值来看, 各混播种类处理间无显著差异(
p > 0.05)。从混播比例来看, 各混播比例间
OV2 在各年份及3年平均值间无显著差异(
p > 0.05); 2013年
OV 2 高于其他年份。从各混播组合来说, 各混播组合的
OV 2 无显著差异(
p > 0.05)。
2.2 一年生和多年生豆禾混播草地牧草产量多样性效应的比较 由
图1 可知, 从混播物种来看, 2013、2014年AM2的
CE 均显著高于其他混播种类(
p < 0.05); 2015年PM2-A的
CE 显著高于其他混播种类(
p < 0.05); 从3年平均值来看, AM2的
CE 显著高于其他混播种类(
p < 0.05)。从混播比例来看, 各混播比例间
CE 在各年份及3年平均值间无显著差异(
p > 0.05); 2013年
CE 高于其他年份。从各混播组合来看, AM2-3和AM2-2之间
CE 无显著差异(
p > 0.05), 但显著高于其他混播组合(
p < 0.05)。从混播物种来看, 2013年AM2的
SEF 显著低于PM2-A、PM4和PM6 (
p < 0.05); 2014年AM2的
SEF 也显著低于PM2-A、PM4和PM6 (
p < 0.05), PM2-A的
SEF 显著低于PM4和PM6 (
p < 0.05); 2015年AM2的
SEF 则显著高于其他混播种类(
p < 0.05), PM6的
SEF 显著高于PM2-A (
p < 0.05); 从3年平均值来看, AM2的
SEF 显著低于PM2-A、PM4和PM6 (
p < 0.05)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1不同混播物种、混播比例和处理间(3年平均值±标准误)互补效应、选择效应和多样性净效应的比较。不同小写字母表示各处理间差异显著(p < 0.05)。混播物种和处理编号同 表1。 Fig. 1Comparison of complementarities effect, selection effect and net effects of biodiversity under different mixed species, mixed proportion and treatments (three years average ± SE). Different lowercase letters within the same year indicate significant difference (p < 0.05). Mixed species and treatment number see Table 1. 从混播比例来看, 各混播比例间
SEF 在各年份及3年平均值间无显著差异(
p > 0.05); 2013年
SEF 低于其他年份。AM2各混播组合间
SEF 无显著差异(
p > 0.05), AM2-3显著高于PM2-A、PM4和PM6的各混播组合 (
p < 0.05)。2013年AM2的?
Y 均显著低于PM2-A、PM4和PM6 (
p < 0.05); 2014年AM2的?
Y 也显著低于PM2-A和PM4 (
p < 0.05), 2015年AM2的?
Y 则显著高于PM4 (
p < 0.05); 从3年平均值来看, AM2的?
Y 显著低于PM2-A和PM4 (
p < 0.05)。从混播比例来看, 各混播比例间?
Y 在各年份及3年平均值间无显著差异(
p > 0.05); 且呈逐年下降趋势。从各混播组合来看, AM2-1的?
Y 显著低于PM2-A、PM4的各组合及PM6 (
p < 0.05)。牧草产量、
OV 1 、
OV 2 、
CE 、
SEF 和?
Y 的双因素方差分析(
表3 )表明, 不同混播种类间牧草产量、
OV 1 、
CE 、
SEF 和?
Y 差异极显著(
p < 0.05); 不同混播比例间上述指标差异不显著(
p > 0.05), 混播种类与混播比例间的交互效应差异不显著(
p > 0.05)。
Table 3 表3 表3 混播物种与混播比例的双因素差异分析
Table 3
Two-way ANOVA analysis of mixed species and mixed proportion 参数 Parameter 变异来源 Source of variation 自由度 df 均方根 F p 产量 混播物种 Mixed species 6 0.852 12.756 0.000** 混播比例 Mixed proportion 2 0.005 0.080 0.923 年份 Year 2 18.983 96.640 0.000** 混播物种×混播比例 Mixed species × Mixed proportion 9 0.017 0.262 0.994 混播物种×年份 Mixed species × Year 9 2.746 13.978 0.000** 混播比例×年份 Mixed proportion ×Year 4 0.013 0.068 0.991 混播物种×混播比例×年份 Mixed species × Mixed proportion × Year 12 0.046 0.235 0.996 误差 Error 171 0.067 总计 Total 192 互补效应 混播物种 Mixed species 3 0.665 10.564 0.000** 混播比例 Mixed proportion 2 0.030 0.473 0.625 年份 Year 2 0.285 20.190 0.000** 混播物种×混播比例 Mixed species × Mixed proportion 6 0.070 1.106 0.365 混播物种×年份 Mixed species × Year 6 0.130 9.221 0.000** 混播比例×年份 Mixed proportion × Year 4 0.018 1.258 0.295 混播物种×混播比例×年份 Mixed species × Mixed proportion × Year 12 0.006 0.445 0.939 误差 Error 96 0.063 总计 Total 108 选择效应 混播物种 Mixed species 3 1.324 12.407 0.000** 混播比例 Mixed proportion 2 0.004 0.035 0.966 年份 Year 2 0.337 1.148 0.323 混播物种×混播比例 Mixed species × Mixed proportion 6 0.033 0.313 0.929 混播物种×年份 Mixed species × Year 6 5.210 17.742 0.000** 混播比例×年份 Mixed proportion × Year 4 0.134 0.455 0.768 混播物种×混播比例×年份 Mixed species × Mixed proportion × Year 12 0.137 0.468 0.927 误差 Error 96 0.107 总计 Total 108 多样性净效应 混播物种 Mixed species 3 0.143 4.168 0.008** 混播比例 Mixed proportion 2 0.022 0.630 0.535 年份 Year 2 1.405 15.086 0.000** 混播物种×混播比例 Mixed species × Mixed proportion 6 0.022 0.637 0.701 混播物种×年份 Mixed species × Year 6 2.450 26.300 0.000** 混播比例×年份 Mixed proportion × Year 4 0.007 0.077 0.989 混播物种×混播比例×年份 Mixed species × Mixed proportion × Year 12 0.058 0.625 0.815 误差 Error 96 0.034 总计 Total 108
**,p < 0.01. 新窗口打开 |
下载CSV 2.3 物种丰富度和均匀度对牧草产量的影响 由
图2 可知, 物种丰富度(混播种类)与牧草产量之间在2013和2014年呈二次曲线函数关系, 其拟合模型分别为
y = -0.0448
x 2 + 0.3618
x + 0.0347 (
R 2 = 0.386 8,
p < 0.01),
y = -0.0309
x 2 + 0.2544
x - 0.0947 (
R 2 = 0.560 4,
p < 0.01)。即牧草产量随着物种丰富度的增加先上升, 当物种数达到一定水平(4种)后开始呈现下降趋势。物种丰富度与牧草产量出现这种“上升-下降”变化的关系, 可能是因为随着混播物种的增加群体在空间结构上达到饱和或对于土壤养分的需求加剧所致。2015年牧草产量与物种丰富度的关系不明显, 可能是因为禾草处于竞争优势地位, 而豆科牧草处于竞争劣势地位(驴食草、紫花苜蓿和红车轴草对群体生物量的贡献率分别为5.68%、6.77%和8.03%), 无法对植物多样性-生态功能产生有效的正向影响。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2物种丰富度和均匀度与牧草产量的关系。AM2、PM2-A、PM4、PM6同表1。 Fig. 2Relationship between species richness, evenness and forage yield. AM2, PM2-A, PM4, PM6 see Table 1. 牧草产量3年平均值呈现二次函数的关系, 说明物种丰富度在一定程度上对混播草地的牧草产量具有正效应, 当超过阈值后会产生负效应。由
图2 和
表4 可知, AM2的牧草产量与物种均匀度(混播比例)之间在2013、2015年和3年平均值都呈二次曲线函数关系, 即牧草产量随着物种均匀度的增加先上升, 当物种均匀度达到一定水平(豆禾混播比例为5:5) 后开始呈现下降趋势; 2014年呈线性函数关系, 即牧草产量随着物种均匀度的增加呈线性下降趋势。PM2-A的牧草产量与物种均匀度之间在2013、2014、2015年和3年平均值都呈二次曲线函数关系; PM4的牧草产量与物种均匀度之间在2013、2014年和3年平均值都呈二次曲线函数关系, 2015年呈三项式函数关系; PM6的牧草产量与物种均匀度在2013、2015年和3年平均值都呈二次曲线函数关系, 2014年呈线性增加趋势。
Table 4 表4 表4 物种均匀度与牧草产量的函数拟合模型
Table 4
Function fitting models between species evenness and forage yield 年份 Year 处理编号 Treatment number 拟合曲线 Fitting curve R2 p 2013 AM2 y = -1.3682x 2 + 1.0218x + 1.1065 0.282 5 >0.05 PM2-A y = -1.0161x 2 + 1.4789x + 0.1569 0.688 1 <0.01 PM4 y = -1.6284x 2 + 1.8951x + 0.1763 0.667 3 <0.01 PM6 y = -1.3567x 2 + 1.624x + 0.1476 0.601 7 <0.01 2014 AM2 y = -0.0333x + 0.7827 0.002 7 >0.05 PM2-A y = -0.6501x 2 + 0.5594x + 0.2012 0.416 8 <0.05 PM4 y = -1.2421x 2 + 1.2671x + 0.0981 0.777 1 <0.01 PM6 y = 0.3897x + 0.1251 0.769 9 <0.01 2015 AM2 y = -0.6322x 2 + 0.8796x + 0.0887 0.782 0 <0.01 PM2-A y = -0.2212x 2 + 0.1058x + 0.3687 0.307 4 <0.05 PM4 y = -1.125x 3 + 1.7721x 2 - 0.7338x + 0.3097 0.603 1 <0.01 PM6 y = -0.1765x 2 + 0.0925x + 0.3122 0.535 4 <0.01 3年平均值 AM2 y = -0.7384x 2 + 0.7472x + 0.6114 0.042 2 >0.05 PM2-A y = -0.5024x 2 + 0.6473x + 0.2434 0.164 4 <0.05 PM4 y = -0.9286x 2 + 1.0004x + 0.1930 0.258 7 <0.05 PM6 y = -0.5301x 2 + 0.7211x + 0.1922 0.237 5 <0.05
处理编号同表1 。 Treatment number see Table 1 . 新窗口打开 |
下载CSV 2.4 多样性效应与增产效应、超产效应和建植时间的关系 由
图3 可知, 将
OV 1 与
CE 、
SEF 和?
Y 进行拟合, 发现
OV 1 与
CE 呈负相关的发展趋势(
p > 0.05), 而与
SEF 和?
Y 均呈显著正相关关系(
p < 0.05)。由
图4 可知, PM2-A的
CE 、
SEF 和?
Y 均呈现2013年显著高于2014年, 2014年显著高于2015年; PM4和PM6的
CE 表现为2013和2014年差异不显著但显著高于2015年; 而
SEF 为2014年显著高于2015年, 2015年显著高于2013年; ?
Y 呈现2014年显著高于2013和2015年; 从年际间的变化趋势来看PM2-A、PM4和PM6的
CE 、
SEF 和?
Y 均随建植时间的延长呈下降趋势。由
图5 可知, 随建植时间的延长, PM2-A处理
OV 1 和
OV 2 的变异系数均呈增加趋势, PM4处理
OV 1 的变异系数也呈增加趋势, 但
OV 2 的变异系数呈下降趋势; PM6处理
OV 1 和
OV 2 的变异系数也均呈增加趋势。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3超产效应1与互补效应、选择效应和多样性效应的关系。 Fig. 3Relationship between transgressive overyielding effect 1 and complementary effect, selection effect and net effect of biodiversity. 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4互补效应、选择效应和多样性效应的年际间变化(平均值±标准误)。不同小写字母、大写字母和a′、b′和c′等符号分别表示PM2-A、PM4和PM6年际间的差异显著(p < 0.05)。PM2-A、PM4、PM6同 表1。 Fig. 4Change of the complementary effect, selection effect and net effect of biodiversity with establishment years (mean ± SE). Different lowercase letters, uppercase letters, and a′, b′ and c′ symbols respectively indicate significant differences of PM2-A, PM4 and PM6 treatments from 2013 to 2015 (p < 0.05). PM2-A、PM4、PM6 see Table 1. 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5PM2-A、PM4和PM6处理超产效应(OV)稳定性分析。PM2-A、PM4、PM6同表1。 Fig. 5Stability analysis of transgressive overyieding effect (OV) for PM2-A, PM4 and PM6. CV, coefficient of variation. PM2-A, PM4, PM6 see Table 1. 3 讨论 3.1 生长时间对超产效应和多样性效应的影响 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 )。对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面。现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 )。如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化。本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 )。相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小。因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应。本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 )。因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 )。这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关。
与此同时, 生长时间还影响了超产效应与多样性效应的关系。本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者。该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 )。但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应。故而在建植后期, 互补效应的作用则更为关键。朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合。本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 )。一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致。
3.2 生长时间对物种丰富度和均匀度与生产力间关系的影响 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 )。物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 )。本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致。这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 )。但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 )。由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 )。
物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 )。物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 )。对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 )。本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 )。这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 )。随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 )。陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加。本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 )。因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力。这可能与本试验设计的豆禾混播比例较高有关。
4 结论 在新疆昭苏地区建植的一年生和多年生豆禾混播草地均可产生超产效应, 有利于当地草牧业的健康发展。(1)在建植初期, 互补效应和选择效应共同主导了多年生豆禾混播草地的超产效应, 而随着生长年限的延长, 互补效应则成为主要影响因素; 一年生豆禾混播草地的超产效应则一直受互补效应的影响。(2)多年生豆禾混播草地的互补效应、选择效应和多样性净效应均随生长年限的延长而呈下降趋势, 也导致了超产效应的下降。(3)一年生豆禾混播草地的超产效应在建植的第1、2年以及3年平均值均可低于多年生豆禾混播草地, 只有在建植第3年才高于多年生豆禾混播草地。由此可见, 合适的混播种类、种类搭配和混播比例可以使多年生豆禾混播草地的生产优势大于一年生豆禾混播草地。
[1] Atwater DZ Callaway RM 2015 ). Testing the mechanisms of diversity-dependent overyielding in a grass speciesEcology 3332 -3342 . PMID:
26909438 [本文引用: 2] Plant diversity enhances many ecosystem processes, including productivity, but these effects have been studied almost exclusively at the taxonomic scale of species. We explore the effect of intraspecific diversity on the productivity of a widespread and dominant grassland species using accessions collected from populations throughout its range. We found that increasing population/ecotype diversity of Pseudoroegneria spicata increased productivity to a similar degree as that reported for species diversity. However, we did not find evidence that overyielding was related to either resource depletion or to pathogenic soil fungi, two causes of overyielding in species-diverse communities. Instead, larger accessions overyielded at low diversity at the expense of smaller accessions, and small accessions overyielded through complementarity at all levels of diversity. Furthermore, overyielding was stronger for accessions from mesic environments, suggesting that local adaptation might predictably influence how plants respond to increases in diversity. This suggests that mass-based competition or other cryptic accession-specific processes had complex but important effects on overyielding. Our results indicate that the effects of diversity within a species can be substantial but that overyielding by intraspecifically diverse populations may not be through the same processes thought to cause overyielding in species diverse communities.
[2] Beckage B Gross LJ 2006 ). Overyielding and species diversity: What should we expect?New Phytologist 140 -148 . PMID:
16945096 [本文引用: 1] Recent empirical studies have found evidence of increased biomass production ('overyielding') in species mixtures relative to monoculture, but the interpretation of these results remains controversial, in part, because of the lack of a theoretical expectation. Here, we examined the expected frequency and stability of overyielding species mixtures using Lotka-Volterra models of species dynamics in two- and four-species systems in conjunction with community, population, and specific rate of biomass production (SRP) definitions of overyielding. Overyielding plant mixtures represented > 55% of potential species assemblages under community definitions and approximately 100% of species were either overyielding or underyielding under the population definition. Our species simulations approached their equilibria in 1-2 yr, supporting the relevancy of an equilibrial analysis. The range of parameter space that we explored produced realistic values of plot biomass, supporting their biological relevance. We show that overyielding is expected to be common under community definitions and population definitions. Overyielding, under community or population definitions, does not imply an actual increase in the specific rate of biomass production. In addition, assemblages of overyielding and underyielding species under all three definitions can be stable over time with underyielding species persisting in the presence of overyielding species.
[3] Cardinale BJ Wright JP Cadotte MW Carroll IT Hector A Srivastava DS Loreau M Weis JJ 2007 ). Impacts of plant diversity on biomass production increase through time because of species complementarityProceedings of the National Academy of Sciences of the United States of America 18123 -18128 . PMID:
17991772 [本文引用: 2] Accelerating rates of species extinction have prompted a growing number of researchers to manipulate the richness of various groups of organisms and examine how this aspect of diversity impacts ecological processes that control the functioning of ecosystems. We summarize the results of 44 experiments that have manipulated the richness of plants to examine how plant diversity affects the production of biomass. We show that mixtures of species produce an average of 1.7 times more biomass than species monocultures and are more productive than the average monoculture in 79% of all experiments. However, in only 12% of all experiments do diverse polycultures achieve greater biomass than their single most productive species. Previously, a positive net effect of diversity that is no greater than the most productive species has been interpreted as evidence for selection effects, which occur when diversity maximizes the chance that highly productive species will be included in and ultimately dominate the biomass of polycultures. Contrary to this, we show that although productive species do indeed contribute to diversity effects, these contributions are equaled or exceeded by species complementarity, where biomass is augmented by biological processes that involve multiple species. Importantly, both the net effect of diversity and the probability of polycultures being more productive than their most productive species increases through time, because the magnitude of complementarity increases as experiments are run longer. Our results suggest that experiments to date have, if anything, underestimated the impacts of species extinction on the productivity of ecosystems.
[4] Chang SH Hou FJ Yu YW Nan ZB 2004 ). Vegetation and soil characteristic of three forage legume pastures on the Loess PlateauActa Ecologica Sinica 932 -937 . [本文引用: 1] [ 常生华 , 侯扶江 , 于应文 , 南志标 (2004 ). 黄土丘陵沟壑区三种豆科人工草地的植被与土壤特征生态学报 , 24,932 -937 .] [本文引用: 1] [5] Chen C Zhu ZH Li YN Yao TH Pan SY Wei XH Kong BB Du JL 2016 ). Effects of interspecific trait dissimilarity and species evenness on the relationship between species diversity and functional diversity in an alpine meadowActa Ecologica Sinica 661 -674 . [本文引用: 2] [ 陈超 , 朱志红 , 李英年 , 姚天华 , 潘石玉 , 卫欣华 , 孔彬彬 , 杜家丽 (2016 ). 高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响生态学报 , 36,661 -674 .] [本文引用: 2] [6] Chen N Zhang YJ Zhu JT Li JX Liu YJ Zu JX Cong N Huang K Wang L 2018 ). Nonlinear responses of productivity and diversity of alpine meadow communities to degradationChinese Journal of Plant Ecology 50 -65 . DOI:
10.17521/cjpe.2017.0252 URL PMID:
a4431765-e95a-47c7-ba86-e990a189bbdb [本文引用: 1] [ 陈宁 , 张扬建 , 朱军涛 , 李军祥 , 刘瑶杰 , 俎佳星 , 丛楠 , 黄珂 , 王荔 (2018 ). 高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究植物生态学报 , 42,50 -65 .] DOI:
10.17521/cjpe.2017.0252 PMID:
a4431765-e95a-47c7-ba86-e990a189bbdb [本文引用: 1] 为了阐释青藏高原高寒草甸退化的关键生态过程, 该研究依托藏北高原草地生态系统研究站(那曲站), 设置不同退化梯度实验, 即对照、轻度退化、中度退化、重度退化和极度退化5个梯度, 探究群落生产力和物种多样性对不同退化强度的响应机制。结果表明: 1)随着退化程度不断加剧, 地上生物量呈现线性或非线性增加趋势, 在重度退化处理下, 地上生物量显著高于对照32.3%, 其中高山嵩草(Kobresia pygmaea)地上生物量呈非线性下降趋势, 而矮火绒草(Leontopodium nanum)地上生物量呈非线性增加趋势; 2)与地上生物量的响应模式相反, 随着退化程度加剧, 地下生物量与总生物量均呈现非线性降低趋势; 3)高寒草甸退化过程中, 物种辛普森指数、丰富度指数、香农多样性指数和均匀度指数均呈现非线性上升趋势。结构等式方程结果表明, 土壤碳含量和体积含水量与地下生物量均呈现显著的正相关关系。土壤碳含量、体积含水量和砾石质量比对地上生物量无显著影响, 土壤碳、氮含量与物种多样性指数呈现显著的负相关关系。研究认为地上生产力的变化不能很好地指示草地的退化程度, 建议今后研究应以可食性牧草和毒杂草等植物功能群的变化来衡量草地退化。
[7] Chen YX Liu J Chen XP Zhang CC Huang W Tang YQ Xu KW 2013 ). Dry matter accumulation, yield and nitrogen use efficiency of crops rotation and intercropping systems in SichuanJournal of China Agricultural University 6 ),68 -79 . [本文引用: 2] [ 陈远学 , 刘静 , 陈新平 , 张朝春 , 黄蔚 , 唐义琴 , 徐开未 (2013 ). 四川轮套作体系的干物质积累、产量及氮素利用效率研究中国农业大学学报 , 18(6 ),68 -79 .] [本文引用: 2] [8] Cox S Peel MD Creech JE Waldron BL Eun JS Zobell DR Miller LR Snyder DL 2017 ). Forage production of grass-legume binary mixtures on Intermountain Western USA irrigated pasturesCrop Science 1742 -1753 . DOI:
10.2135/cropsci2016.04.0235 URL [本文引用: 1] [9] Dai W Zhang R Du ZB Wang F 2009 ). Soil fertility and species identity control community productivity in an experimental plant community in an area of subalpine meadowChinese Journal of Plant Ecology 45 -52 . PMID:
2F18E407-FF71-4C30-A737-379FF8009B84 [本文引用: 1] [ 代巍 , 张荣 , 独占彪 , 王璠 (2009 ). 土壤肥力和物种属性决定亚高寒草甸实验群落的生产力植物生态学报 , 33,45 -52 .] DOI:
10.3773/j.issn.1005-264x.2009.01.005 PMID:
2F18E407-FF71-4C30-A737-379FF8009B84 [本文引用: 1] 生物多样性与生态系统功能之间的关系及其形成的内在机制还存在很多争议。为了揭示植物群落生产力形成的生态学机制, 采用盆栽方法探讨了物种多样性、物种属性以及施肥水平与植物群落生产力之间的关系。研究结果显示: 在不施肥和每盆施5.0 g磷酸二铵的条件下, 随着物种多样性的增加, 地上生物量增加不显著; 在每盆施10.0 g磷酸二铵的条件下, 随着物种多样性的增加, 地上生物量显著增加。相对于中华羊茅(Festuca sinensis)而言, 垂穗披碱草(Elymus nutans)和垂穗鹅观草(Roegneria nutans)对群落生产力的贡献较大, 但在不同施肥水平和播种密度下, 其影响不完全相同。这表明物种多样性对群落生产力的影响随着土壤肥力的变化而变化; 并且植物群落生产力受组成群落的物种属性影响较大, 而物种属性又与特定时间和特定生境下资源的利用方式相联系。在高肥力水平下, 物种多样性之所以对群落生产力具有正效应, 可能是因为高肥力水平增加了可利用的生态位空间, 最终仍体现在物种组合上。因此, 植物群落的生产力与物种多样性之间没有必然的联系, 而与土壤肥力和土壤肥力决定的物种属性有关。
[10] de Mazancourt C Isbell F Larocque A Berendse F de Luca E Grace JB Haegeman B Polley HW Roscher C Schmid B Tilman D van Ruijven J Weigelt A Wilsey BJ Loreau M 2013 ). Predicting ecosystem stability from community composition and biodiversityEcology Letters 617 -625 . DOI:
10.1111/ele.12088 PMID:
23438189 [本文引用: 1] As biodiversity is declining at an unprecedented rate, an important current scientific challenge is to understand and predict the consequences of biodiversity loss. Here, we develop a theory that predicts the temporal variability of community biomass from the properties of individual component species in monoculture. Our theory shows that biodiversity stabilises ecosystems through three main mechanisms: (1) asynchrony in species' responses to environmental fluctuations, (2) reduced demographic stochasticity due to overyielding in species mixtures and (3) reduced observation error (including spatial and sampling variability). Parameterised with empirical data from four long-term grassland biodiversity experiments, our prediction explained 22-75% of the observed variability, and captured much of the effect of species richness. Richness stabilised communities mainly by increasing community biomass and reducing the strength of demographic stochasticity. Our approach calls for a re-evaluation of the mechanisms explaining the effects of biodiversity on ecosystem stability.? 2013 Blackwell Publishing Ltd/CNRS.
[11] Finn JA Kirwan L Connonlly J Sebastià MT Helgadottir A Baadshaug OH Belanger G Black A Brophy C Collins RP Cop J Dalmannsdóttir S Delgodo I Elgersma A Fothergill M et al 2013 ). Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: a 3-year continental scale field experimentJournal of Applied Ecology 365 -375 . DOI:
10.1111/1365-2664.12041 URL [本文引用: 5] [12] Frankow-Lindberg BE Halling M H?glind M Forkman J 2009 ). Yield and stability of yield of single- and multi-clover grass-clover swards in two contrasting temperate environmentsGrass and Forage Science 236 -245 . DOI:
10.1111/gfs.2009.64.issue-3 URL [本文引用: 1] [13] Grime JP 1998 ). Benefits of plant diversity to ecosystems: immediate, filter and founder effectsJournal of Ecology 902 -910 . DOI:
10.1046/j.1365-2745.1998.00306.x URL [本文引用: 1] [14] Hector A Hautier Y Saner P Wacker L Bagchi R Joshi J Scherer-Lorenzen M Spehn EM Bazeley-White E Weilenmann M Caldeira MC Dimitrakopoulos PG Finn JA Huss-Danell K Jumpponen A et al 2010 ). General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyieldingEcology 2213 -2220 . PMID:
20836442 [本文引用: 4] Insurance effects of biodiversity can stabilize the functioning of multispecies ecosystems against environmental variability when differential species' responses lead to asynchronous population dynamics. When responses are not perfectly positively correlated, declines in some populations are compensated by increases in others, smoothing variability in ecosystem productivity. This variance reduction effect of biodiversity is analogous to the risk-spreading benefits of diverse investment portfolios in financial markets. We use data from the BIODEPTH network of grassland biodiversity experiments to perform a general test for stabilizing effects of plant diversity on the temporal variability of individual species, functional groups, and aggregate communities. We tested three potential mechanisms: reduction of temporal variability through population asynchrony; enhancement of long-term average performance through positive selection effects; and increases in the temporal mean due to overyielding. Our results support a stabilizing effect of diversity on the temporal variability of grassland aboveground annual net primary production through two mechanisms. Two-species communities with greater population asynchrony were more stable in their average production over time due to compensatory fluctuations. Overyielding also stabilized productivity by increasing levels of average biomass production relative to temporal variability. However, there was no evidence for a performance-enhancing effect on the temporal mean through positive selection effects. In combination with previous work, our results suggest that stabilizing effects of diversity on community productivity through population asynchrony and overyielding appear to be general in grassland ecosystems.
[15] Hector A Schmid B Beierkuhnlein C Caldeira MC Diemer M Dimitrakopoulos PG Finn JA Freitas H Giller PS Good J Harris R H?gberg P Huss-Danell K Joshi J Jumpponen A et al 1999 ). Plant diversity and productivity experiments in European grasslandsScience 1123 -1127 . DOI:
10.1126/science.286.5442.1123 URL [本文引用: 3] [16] Hendriks M Mommer L de Caluwe H Smit-Tiekstra AE van der Putten WH de Kroon H 2013 ). Independent variations of plant and soil mixtures reveal soil feedback effects on plant community overyieldingJournal of Ecology 287 -297 . DOI:
10.1111/jec.2013.101.issue-2 URL [本文引用: 1] [17] Hooper DU Chapin III FS Ewel JJ Hector A Inchausti P Lavorel S Lawton JH Lodge DM Loreau M Naeem S Schmid B Set?l? H Symstad AJ Vandermeer J Wardle DA 2005 ). Effects of biodiversity on ecosystem functioning: a consensus of current knowledgeEcological Monographs 3 -35 . DOI:
10.1890/04-0922 URL [本文引用: 3] [18] Hu ZZ Dong SK Long RJ 2001 ). Evaluation of ecological efficiency of mixed pastures with perennial grasses in alpine region of ChinaGrassland Science Outlook of 21 Century—Proceedings of International Prataculture (Grassland) Congress 482 -487 . [本文引用: 1] [ 胡自治 , 董世魁 , 龙瑞军 (2001 ). 高寒地区多年生禾草混播草地生产——生态效益评价21世纪草业科学展望——国际草业(草地)学术大会论文集 .482 -487 .] [本文引用: 1] [19] Huston MA Aarssen LW Austin MP Cade BS Fridley JD Garnier E Grime JP Hodgson J Lauenroth WK Thompson K Vandermeer JH Wardle DA 2000 ). No consistent effect of plant diversity on productivityScience 10.1126/science.289.5483.1255a . DOI:
10.1126/science.289.5483.1255a [本文引用: 1] [20] Isbell FI Polley HW Wilsey BJ 2009 ). Biodiversity, productivity and the temporal stability of productivity: patterns and processesEcology Letters 443 -451 . DOI:
10.1111/ele.2009.12.issue-5 URL [本文引用: 1] [21] Jiang XL Li WQ Yue J Hao M Zhang WG Liu YJ Li F 2010 ). Effects of available soil space on the diversity- productivity relationship of speciesJournal of Lanzhou University Natural Sciences ), 46(6 ),62 -67 . [本文引用: 4] [ 江小雷 , 李伟绮 , 岳静 , 郝敏 , 张卫国 , 刘允佳 , 李飞 (2010 ). 土壤有效空间对物种多样性-生产力关系的影响兰州大学学报(自然科学版) , 46(6 ),62 -67 .] [本文引用: 4] [22] Jiang XL Zhang WG Duan ZH 2005 ). Effects of complementarity on diversity-productivity relationshipActa Phytoecologica Sinica 523 -529 . PMID:
16213ACD-1657-40DB-A770-148AF8BE78BF [本文引用: 3] [ 江小雷 , 张卫国 , 段争虎 (2005 ). 资源互补效应对多样性-生产力关系的影响植物生态学报 , 29,523 -529 .] DOI:
10.17521/cjpe.2005.0070 PMID:
16213ACD-1657-40DB-A770-148AF8BE78BF [本文引用: 3] 许多有关物种多样性-生态系统功能关系的观察、理论和实验研究都表明, 在局域尺度范围内, 植物种多样性对生态系统生产力存在正效应。 然而, 对于促成这种关系的潜在生态学机制却缺乏足够的了解。 该实验利用9种一年生栽培牧草, 采用各物种单播及混播的方法, 构建不同多样性梯度的实验群落, 对物种多样性与生态系统生产力的关系及资源互补效应对系统生产力的影响进行了研究。 结果表明, 在一年生植物群落内,植物种多样性在一定程度内对系统生产力存在正效应, 物种多样性与生产力呈二次函数关系, 关系式为y = -98.449x2 + 1 039.2 x - 42.407, (R2 = 0.423 1)。 各物种在资源利用、生长速度和竞争能力等功能特征方面存在较大差异, 最高产物种和最低产物种间产量相差5.8倍。 在同一多样性梯度内, 不同物种组合的群落间生产力和互补效应也存在较大差异, 说明物种的成分对生态系统生产力也有重要影响。 同时,在混播群落中程度不同地存在着资源的互补性利用, 说明物种多样性对系统生产力有增强作用, 但相关分析表明, 互补效应和物种多样性间不存在显著相关关系。互补效应的4种计算方法所反映的资源互补程度有所不同, 每种方法各有利弊, 在对系统的多样性效应作用机制进行评价时, 应根据具体情况, 同时采用几种方法, 以利于对资源互补效应做出恰当的估测。
[23] Jiao NY Ning TY Zhao C Wang Y Shi ZQ Hou LT Fu GZ Jiang XD Li ZJ 2006 ). Characters of photosynthesis in intercropping system of maize and peanutActa Agronomica Sinica 917 -923 . [本文引用: 2] [ 焦念元 , 宁堂原 , 赵春 , 王芸 , 史忠强 , 侯连涛 , 付国占 , 江晓东 , 李增嘉 (2006 ). 玉米花生间作复合体系光合特性的研究作物学报 , 32,917 -923 .] [本文引用: 2] [24] Küchenmeister F Küchenmeister K Wrage N Kayser M Isselstein J 2012 ). Yield and yield stability in mixtures of productive grassland species: Does species number or functional group composition matter?Grassland Science 94 -100 . DOI:
10.1111/grs.2012.58.issue-2 URL [本文引用: 3] [25] Li A Zhang M Du GZ 2012 ). Impacts of species composition, richness, sowing density, and soil nutrients on the complementary effect of plant communitiesChinese Journal of Ecology 2443 -2448 . [本文引用: 4] [ 李昂 , 张鸣 , 杜国祯 (2012 ). 物种组成、丰富度、播种密度和土壤养分对群落补偿效应的影响生态学杂志 , 31,2443 -2448 .] [本文引用: 4] [26] Li CJ Li YY Yu CB Sun JH Christie P An M Zhang FS Li L 2011 ). Crop nitrogen use and soil mineral nitrogen accumulation under different crop combinations and patterns of strip intercropping in northwest ChinaPlant and Soil 221 -231 . DOI:
10.1007/s11104-010-0686-6 URL [本文引用: 2] [27] Li L Li SM Sun JH Zhou LL Bao XG Zhang HG Zhang FS 2007 ). Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus- deficient soilsProceedings of the National Academy of Sciences of the United States of America 11192 -11196 . [本文引用: 3] [28] Li L Tilman D Lambers H Zhang FS 2014 ). Plant diversity and overyielding: insights from belowground facilitation of intercropping in agricultureNew Phytologist 63 -69 . DOI:
10.1111/nph.2014.203.issue-1 URL [本文引用: 4] [29] Li L Zhang F Li X Christie P Sun J Yang S Tang C 2003 ). Interspecific facilitation of nutrient uptake by intercropped maize and faba beanNutrient Cycling in Agroecosystems 61 -71 . DOI:
10.1023/A:1021885032241 URL [本文引用: 1] [30] Lithourgidis AS Vasilakoglou IB Dhima KV Dordas CA Yiakoulaki MD 2006 ). Forage yield and quality of common vetch mixtures with oat and triticale in two seeding ratiosField Crops Research 106 -113 . DOI:
10.1016/j.fcr.2006.03.008 URL [本文引用: 1] [31] Liu WH Zhang YJ Shi SL Sun J Wei XX 2017 ). Effect of variety, fertilization and mixture sowing on oat biomass allocation in the alpine cultivated pastureJournal of Plant Nutrition and Fertilizer 398 -407 . [本文引用: 1] [ 刘文辉 , 张英俊 , 师尚礼 , 孙建 , 魏小星 (2017 ). 高寒地区燕麦(Avena sativa L.)人工草地生物量分配对施肥和混播措施的响应植物营养与肥料学报 , 23,398 -407 .] [本文引用: 1] [32] Loreau M 1998 ). Separating sampling and other effects in biodiversity experimentsOikos 600 -602 . DOI:
10.2307/3546381 URL [本文引用: 1] [33] Loreau M Hector A 2001 ). Partitioning selection and complementarity in biodiversity experimentsNature 72 -76 . PMID:
11452308 [本文引用: 4] The impact of biodiversity loss on the functioning of ecosystems and their ability to provide ecological services has become a central issue in ecology. Several experiments have provided evidence that reduced species diversity may impair ecosystem processes such as plant biomass production. The interpretation of these experiments, however, has been controversial because two types of mechanism may operate in combination. In the 'selection effect', dominance by species with particular traits affects ecosystem processes. In the 'complementarity effect', resource partitioning or positive interactions lead to increased total resource use. Here we present a new approach to separate the two effects on the basis of an additive partitioning analogous to the Price equation in evolutionary genetics. Applying this method to data from the pan-European BIODEPTH experiment reveals that the selection effect is zero on average and varies from negative to positive in different localities, depending on whether species with lower- or higher-than-average biomass dominate communities. In contrast, the complementarity effect is positive overall, supporting the hypothesis that plant diversity influences primary production in European grasslands through niche differentiation or facilitation.
[34] Ma CH Han JG Li HX Mao PS Rong YP 1999 ). The dynamical studies on biomass, quality and interspecific competition in annual forage crop swardsActa Agrestia Sinica 1 ),62 -71 . [本文引用: 1] [ 马春晖 , 韩建国 , 李鸿祥 , 毛培胜 , 戎郁萍 (1999 ). 一年生混播草地生物量和品质以及种间竞争的动态研究草地学报 , 7(1 ),62 -71 .] [本文引用: 1] [35] Mulder C Jumpponen A H?gberg P Huss-Danell K 2002 ). How plant diversity and legumes affect nitrogen dynamics in experimental grassland communitiesOecologia 412 -421 . DOI:
10.1007/s00442-002-1043-0 PMID:
28466208 [本文引用: 2] Positive relationships between species richness and ecosystem processes such as productivity or nitrogen cycling can be the result of a number of mechanisms. We examined how species richness, biomass, and legume presence, diversity, and abundance explained nitrogen dynamics in experimental grassland plots in northern Sweden. Nitrogen concentrations and δN values were measured in plants grown in 28 mixtures (58 plots) including 1, 2, 4, 8 or 12 local grassland species over four years. Values for δN declined over time for all three functional groups (grasses, legumes, and non-leguminous forbs), suggesting greater reliance on N fixed by legumes over time by all species. Above ground percent nitrogen (%N) also declined over time but root %N and total N did not. Path analysis of above ground data suggested that two main factors affected %N and the size of the N pool. First, higher plant diversity (species richness) increased total N through increased biomass in the plot. Although in the first two years of the experiment this was the result of a greater probability of inclusion of at least one legume, in the last two years diversity had a significant effect on biomass beyond this effect. Second, percent legumes planted in the plots had a strong effect on above ground %N and δN, but a much smaller effect on above ground biomass. In contrast, greater plant diversity affected N in roots both by increasing biomass and by decreasing %N (after controlling for effects mediated by root biomass and legume biomass). Increased legume biomass resulted in higher %N and lower δN for both non-legume forbs and grasses in the first year, but only for grasses in the third year. We conclude that a sampling effect (greater probability of including a legume) contributed towards greater biomass and total N in high-diversity communities early on in the experiment, but that over time this effect weakened and other positive effects of diversity became more important.
[36] Nyfeler D Huguenin-Elie O Suter M Frossard E Connolly J Lüscher A 2009 ). Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyieldingJournal of Applied Ecology 683 -691 . DOI:
10.1111/jpe.2009.46.issue-3 URL [本文引用: 5] [37] Pacala S Tilman D 2001 ). The transition from sampling to complementarityThe Functional Consequences of Biodiversity: Empirical Progress and Theoretical Extensions [本文引用: 2] [38] Russelle MP 1992 ). Nitrogen cycling in pasture and rangeJournal of Production Agriculture 13 -23 . DOI:
10.2134/jpa1992.0013 URL [本文引用: 1] [39] Schuldt A Bruelheide H H?rdtle W Assmann T Li Y Ma K von Oheimb G Zhang J 2015 ). Early positive effects of tree species richness on herbivory in a large-scale forest biodiversity experiment influence tree growthJournal of Ecology 563 -571 . DOI:
10.1111/jec.2015.103.issue-3 URL [本文引用: 1] [40] Siebenk?s A Schumacher J Roscher C 2016 ). Resource availability alters biodiversity effects in experimental grass-forb mixturesPLOS ONE e0158110 . DOI: 10.1371/journal.pone.0158110 . DOI:
10.1371/journal.pone.0158110 URL [本文引用: 1] [41] Song R Mu Y Wang YL Wu CS Guo JX 2002 ). Effects of intercropping of maize and soybean on the morphological character of rootsJournal of Northeast Normal University Natural Science Edition ), 34(3 ),83 -86 . [本文引用: 1] [ 宋日 , 牟瑛 , 王玉兰 , 吴春胜 , 郭继勋 (2002 ). 玉米、大豆间作对两种作物根系形态特征的影响东北师大学报(自然科学版) , 34(3 ),83 -86 .] [本文引用: 1] [42] Thibaut LM Connolly SR 2013 ). Understanding diversity stability relationships: towards a unified model of portfolio effectsEcology Letters 140 -150 . DOI:
10.1111/ele.12019 PMID:
23095077 [本文引用: 1] A major ecosystem effect of biodiversity is to stabilise assemblages that perform particular functions. However, diversity-stability relationships (DSRs) are analysed using a variety of different population and community properties, most of which are adopted from theory that makes several restrictive assumptions that are unlikely to be reflected in nature. Here, we construct a simple synthesis and generalisation of previous theory for the DSR. We show that community stability is a product of two quantities: the synchrony of population fluctuations, and an average species-level population stability that is weighted by relative abundance. Weighted average population stability can be decomposed to consider effects of the mean-variance scaling of abundance, changes in mean abundance with diversity and differences in species' mean abundance in monoculture. Our framework makes explicit how unevenness in the abundances of species in real communities influences the DSR, which occurs both through effects on community synchrony, and effects on weighted average population variability. This theory provides a more robust framework for analysing the results of empirical studies of the DSR, and facilitates the integration of findings from real and model communities.? 2012 Blackwell Publishing Ltd/CNRS.
[43] Tilman D Downing JA 1994 ). Biodiversity and stability in grasslandsNature 363 -365 . DOI:
10.1038/367363a0 URL [本文引用: 1] [44] Tilman D Reich PB Knops J Wedin D Mielke T Lehman C 2001 ). Diversity and productivity in a long-term grassland experimentScience 843 -845 . DOI:
10.1126/science.1060391 URL [本文引用: 3] [45] Wang HD Zhang LL Zhu ZH 2013 ). Effects of clipping and fertilizing on the relationships between species diversity and ecosystem functioning and mechanisms of community stability in alpine meadowChinese Journal of Plant Ecology 279 -295 . DOI:
10.3724/SP.J.1258.2013.00028 URL PMID:
48B5B4B7-C6C4-422D-91B5-E81985690251 [本文引用: 2] [ 王海东 , 张璐璐 , 朱志红 (2013 ). 刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制植物生态学报 , 37,279 -295 .] DOI:
10.3724/SP.J.1258.2013.00028 PMID:
48B5B4B7-C6C4-422D-91B5-E81985690251 [本文引用: 2] 植物群落中不同“功能身份”物种的多样性与特定生态系统功能之间具有何种关系及其作用机制尚不明确。通过在高寒矮嵩草(Kobresia humilis)草甸为期5年的刈割(不刈割、留茬3 cm、留茬1 cm)、施肥(施肥、不施肥)和浇水(浇水、不浇水)控制实验, 研究了刈割与土壤资源获得性梯度上不同“功能身份”物种(群落中所有物种、响应物种、作用物种和共有物种)的多样性变化与群落地上净初级生产力和稳定性的关系以及稳定性机制。研究结果显示: 群落中响应物种、作用物种和共有物种数分别占全部物种数的36.6%、18.3%和64.8%, 物种多样性对生态系统功能具有不同的效应, 净初级生产力主要受响应物种和作用物种的多样性变化影响, 而稳定性则主要由共有物种的多样性变化决定; 群落稳定性的维持主要依赖于共有物种的多样性增加, 其作用机制是投资组合效应, 而超产效应和异步性效应对稳定性并无作用; 刈割和施肥对物种多样性、稳定性和净初级生产力具有相反的影响, 前者能增加物种多样性和稳定性, 并降低净初级生产力, 而后者的作用正相反。这与群落中全部物种的多样性变化受刈割影响较大, 而作用物种的多样性变化受资源获得性影响较大有关。上述结果表明高寒草甸生态系统地上净初级生产力主要由少数影响生产力的作用物种的多样性决定, 而稳定性则由大量共有物种的多样性所掌控。投资组合效应是物种多样性导致稳定性的机制。由于群落中不同物种的多样性效应具有分异性, 对于特定的生态系统功能而言, 物种的“功能身份”可能比物种多样性本身更重要, 不加区别地笼统定义物种多样性与生态系统功能的关系可能欠妥。
[46] Wang P Wang TH Zhou DW 2007 ). The productivity of grass-legume mixture in Songnen area of ChinaSciencepaper Online 2 ),121 -128 . [本文引用: 1] [ 王平 , 王天慧 , 周道玮 (2007 ). 松嫩地区禾-豆混播草地生产力研究中国科技论文在线 , 2(2 ),121 -128 .] [本文引用: 1] [47] Wang P Zhou DW Jiang SC 2010 ). Research on biological nitrogen fixation of grass-legume mixtures in a semi-arid area of ChinaActa Prataculturae Sinica 6 ),276 -280 . [本文引用: 1] [ 王平 , 周道玮 , 姜世成 (2010 ). 半干旱地区禾-豆混播草地生物固氮作用研究草业学报 , 19(6 ),276 -280 .] [本文引用: 1] [48] Wang P Zhou DW Zhang BT 2009 ). Coexistence and inter- specific competition in grass-legume mixtureActa Ecologica Sinica 2560 -2567 . [本文引用: 1] [ 王平 , 周道玮 , 张宝田 (2009 ). 禾-豆混播草地种间竞争与共存生态学报 , 29,2560 -2567 .] [本文引用: 1] [49] Wang YK Ding XF Wang XP Wu M Gao SB Yang X Zhao NX Gao YB 2019 ). Genotypic diversity of a dominant species Leymus chinensis inhibited ecological function of species diversity in the Inner Mongolia SteppeActa Ecologica Sinica 1507 -1516 . [本文引用: 2] [ 王宇坤 , 丁新峰 , 王小平 , 吴曼 , 高韶勃 , 杨雪 , 赵念席 , 高玉葆 (2019 ). 内蒙古典型草原建群种羊草基因型多样性抑制群落物种多样性的生态功能生态学报 , 39,1507 -1516 .] [本文引用: 2] [50] Wilsey BJ Potvin C 2000 ). Biodiversity and ecosystem functioning: importance of species evenness in an old fieldEcology 887 -892 . DOI:
10.1890/0012-9658(2000)081[0887:BAEFIO]2.0.CO;2 URL [本文引用: 1] [51] Xiao YB Li L Zhang FS 2005 ). The interspecific nitrogen facilitation and the subsequent nitrogen transfer between the intercropped wheat and faba beanScientia Agricultura Sinica 965 -973 . [本文引用: 1] [ 肖焱波 , 李隆 , 张福锁 (2005 ). 小麦/蚕豆间作体系中的种间相互作用及氮转移研究中国农业科学 , 38,965 -973 .] [本文引用: 1] [52] Xie KY Zhao Y Li XL He F Wan LQ Wang D Han DM 2013 ). Relationships between grasses and legumes in mixed grassland: a reviewActa Prataculturae Sinica 3 ),284 -296 . [本文引用: 1] [ 谢开云 , 赵云 , 李向林 , 何峰 , 万里强 , 王丹 , 韩冬梅 (2013 ). 豆-禾混播草地种间关系研究进展草业学报 , 22(3 ),284 -296 .] [本文引用: 1] [53] Yang XC Hu YG Qian X Ren CZ Lin YC Guo LC Wang CL Zeng ZH 2012 ). Effects of nitrogen application level on system productivity, nitrogen absorption and accumulation in mung bean ‖ oat intercropping systemJournal of China Agricultural University 4 ),46 -53 . [本文引用: 1] [ 杨学超 , 胡跃高 , 钱欣 , 任长忠 , 林叶春 , 郭来春 , 王春龙 , 曾昭海 (2012 ). 施氮量对绿豆‖燕麦间作系统生产力及氮吸收累积的影响中国农业大学学报 , 17(4 ),46 -53 .] [本文引用: 1] [54] Zhang JH Huang YM 2016 ). Biodiversity and stability mechanisms: understanding and future researchActa Ecologica Sinica 3859 -3870 . [本文引用: 2] [ 张景慧 , 黄永梅 (2016 ). 生物多样性与稳定性机制研究进展生态学报 , 36,3859 -3870 .] [本文引用: 2] [55] Zhang QG Zhang DY 2006 ). Resource availability and biodiversity effects on the productivity, temporal variability and resistance of experimental algal communitiesOikos 385 -396 . DOI:
10.1111/j.2006.0030-1299.14723.x URL [本文引用: 1] [56] Zhao C Chai Q Qiao YY Wang JK 2016 ). Effect of cereal- legume spacing in intercropping system on alleviating “N inhibition” in pea plantsChinese Journal of Eco- Agriculture 1169 -1176 . [本文引用: 2] [ 赵财 , 柴强 , 乔寅英 , 王建康 (2016 ). 禾豆间距对间作豌豆“氮阻遏”减缓效应的影响中国生态农业学报 , 24,1169 -1176 .] [本文引用: 2] [57] Zhao CX He WQ Hu YG Dong HM She XL 2006 ). Effect of intercropping or mixture and harvest time on forage yield and quality of oat and pea under low soil nitrogen environmentAgricultural Research in the Arid Areas 4 ),5 -9 . [本文引用: 2] [ 赵彩霞 , 何文清 , 胡跃高 , 董慧明 , 佘小玲 (2006 ). 低氮环境下燕麦与箭筈豌豆间混作与刈割时间对饲草产量及质量特性的影响干旱地区农业研究 , 23(4 ),5 -9 .] [本文引用: 2] [58] Zheng W Jianaerguli , Tang GR Zhu JZ 2015 ). Effects of nitrogen addition on competitive ability and productive performance of legume-grass mixtureActa Agrestia Sinica 422 -425 . [本文引用: 2] [ 郑伟 , 加娜尔古丽 , 唐高溶 , 朱进忠 (2015 ). 氮素添加对豆禾牧草种间竞争力与生产性能的影响草地学报 , 23,422 -425 .] [本文引用: 2] [59] Zheng W Zhu JZ Jianaerguli (2012 ). A comprehensive evaluation of the productive performance of legume-grass mixture under different mixed sowing patternsActa Prataculturae Sinica 6 ),242 -251 . [本文引用: 1] [ 郑伟 , 朱进忠 , 加娜尔古丽 (2012 ). 不同混播方式豆禾混播草地生产性能的综合评价草业学报 , 21(6 ),242 -251 .] [本文引用: 1] [60] Zhu SX Chang J Ge Y Wang H Wang FY Zhang JM Su CH 2014 ). Effects of plant diversity on productivity and diversity effect in a full-scale constructed wetlandEcology and Environmental Sciences 35 -42 . [本文引用: 4] [ 朱四喜 , 常杰 , 葛滢 , 王海 , 王凤友 , 张建民 , 苏春花 (2014 ). 全尺度人工湿地中植物多样性对生产力与多样性效应的影响生态环境学报 , 23,35 -42 .] [本文引用: 4] [61] Zhu YQ Zheng W Wang X Guan ZX 2018 ). Effects plant spacing pattern on root morphological and architectural characteristics of legume-grass mixturesActa Prataculturae Sinica 1 ),73 -85 . [本文引用: 1] [ 朱亚琼 , 郑伟 , 王祥 , 关正翾 (2018 ). 混播方式对豆禾混播草地植物根系构型特征的影响草业学报 , 27(1 ),73 -85 .] [本文引用: 1] Testing the mechanisms of diversity-dependent overyielding in a grass species
2
2015
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Overyielding and species diversity: What should we expect?
1
2006
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
Impacts of plant diversity on biomass production increase through time because of species complementarity
2
2007
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
黄土丘陵沟壑区三种豆科人工草地的植被与土壤特征
1
2004
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
黄土丘陵沟壑区三种豆科人工草地的植被与土壤特征
1
2004
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响
2
2016
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
... ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响
2
2016
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
... ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究
1
2018
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究
1
2018
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
四川轮套作体系的干物质积累、产量及氮素利用效率研究
2
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
四川轮套作体系的干物质积累、产量及氮素利用效率研究
2
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
Forage production of grass-legume binary mixtures on Intermountain Western USA irrigated pastures
1
2017
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
土壤肥力和物种属性决定亚高寒草甸实验群落的生产力
1
2009
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
土壤肥力和物种属性决定亚高寒草甸实验群落的生产力
1
2009
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
Predicting ecosystem stability from community composition and biodiversity
1
2013
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: a 3-year continental scale field experiment
5
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2013 ). ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
Yield and stability of yield of single- and multi-clover grass-clover swards in two contrasting temperate environments
1
2009
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
Benefits of plant diversity to ecosystems: immediate, filter and founder effects
1
1998
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding
4
2010
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ,
2010 ; 朱四喜等,
2014 ). ...
Plant diversity and productivity experiments in European grasslands
3
1999
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
Independent variations of plant and soil mixtures reveal soil feedback effects on plant community overyielding
1
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge
3
2005
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
高寒地区多年生禾草混播草地生产——生态效益评价
1
2001
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
高寒地区多年生禾草混播草地生产——生态效益评价
1
2001
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
No consistent effect of plant diversity on productivity
1
2000
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Biodiversity, productivity and the temporal stability of productivity: patterns and processes
1
2009
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
土壤有效空间对物种多样性-生产力关系的影响
4
2010
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
土壤有效空间对物种多样性-生产力关系的影响
4
2010
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
资源互补效应对多样性-生产力关系的影响
3
2005
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
资源互补效应对多样性-生产力关系的影响
3
2005
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
玉米花生间作复合体系光合特性的研究
2
2006
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
玉米花生间作复合体系光合特性的研究
2
2006
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
Yield and yield stability in mixtures of productive grassland species: Does species number or functional group composition matter?
3
2012
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
物种组成、丰富度、播种密度和土壤养分对群落补偿效应的影响
4
2012
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
物种组成、丰富度、播种密度和土壤养分对群落补偿效应的影响
4
2012
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
Crop nitrogen use and soil mineral nitrogen accumulation under different crop combinations and patterns of strip intercropping in northwest China
2
2011
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus- deficient soils
3
2007
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
Plant diversity and overyielding: insights from belowground facilitation of intercropping in agriculture
4
2014
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
Interspecific facilitation of nutrient uptake by intercropped maize and faba bean
1
2003
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Forage yield and quality of common vetch mixtures with oat and triticale in two seeding ratios
1
2006
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
高寒地区燕麦(
Avena sativa L.)人工草地生物量分配对施肥和混播措施的响应
1
2017
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
高寒地区燕麦(
Avena sativa L.)人工草地生物量分配对施肥和混播措施的响应
1
2017
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
Separating sampling and other effects in biodiversity experiments
1
1998
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Partitioning selection and complementarity in biodiversity experiments
4
2001
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 按照Loreau和Hector (
2001 )的方法, 将混播群落中生产力的增加量(多样性的净效应)分离出选择效应和互补效应. ...
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
一年生混播草地生物量和品质以及种间竞争的动态研究
1
1999
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
一年生混播草地生物量和品质以及种间竞争的动态研究
1
1999
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
How plant diversity and legumes affect nitrogen dynamics in experimental grassland communities
2
2002
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyielding
5
2009
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
The transition from sampling to complementarity
2
2001
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
Nitrogen cycling in pasture and range
1
1992
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
Early positive effects of tree species richness on herbivory in a large-scale forest biodiversity experiment influence tree growth
1
2015
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
Resource availability alters biodiversity effects in experimental grass-forb mixtures
1
2016
... 式中,
Bmno -mean 为混播群落中各组分种的平均单产;
OV 2 > 0, 表明存在超产效应.
OV 2 解释了混播群落生产力与混播组分单播生产力之间的关系, 而
OV 1 强调了混播物种成分的差异与超产效应之间的联系(Siebenk?s
et al. ,
2016 ). ...
玉米、大豆间作对两种作物根系形态特征的影响
1
2002
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
玉米、大豆间作对两种作物根系形态特征的影响
1
2002
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Understanding diversity stability relationships: towards a unified model of portfolio effects
1
2013
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
Biodiversity and stability in grasslands
1
1994
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
Diversity and productivity in a long-term grassland experiment
3
2001
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
... ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制
2
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制
2
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
松嫩地区禾-豆混播草地生产力研究
1
2007
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
松嫩地区禾-豆混播草地生产力研究
1
2007
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
半干旱地区禾-豆混播草地生物固氮作用研究
1
2010
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
半干旱地区禾-豆混播草地生物固氮作用研究
1
2010
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
禾-豆混播草地种间竞争与共存
1
2009
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
禾-豆混播草地种间竞争与共存
1
2009
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
内蒙古典型草原建群种羊草基因型多样性抑制群落物种多样性的生态功能
2
2019
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
内蒙古典型草原建群种羊草基因型多样性抑制群落物种多样性的生态功能
2
2019
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
Biodiversity and ecosystem functioning: importance of species evenness in an old field
1
2000
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
小麦/蚕豆间作体系中的种间相互作用及氮转移研究
1
2005
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
小麦/蚕豆间作体系中的种间相互作用及氮转移研究
1
2005
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
豆-禾混播草地种间关系研究进展
1
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
豆-禾混播草地种间关系研究进展
1
2013
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
施氮量对绿豆‖燕麦间作系统生产力及氮吸收累积的影响
1
2012
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
施氮量对绿豆‖燕麦间作系统生产力及氮吸收累积的影响
1
2012
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
生物多样性与稳定性机制研究进展
2
2016
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
生物多样性与稳定性机制研究进展
2
2016
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
Resource availability and biodiversity effects on the productivity, temporal variability and resistance of experimental algal communities
1
2006
... 植物多样性-生产力关系的选择效应、互补效应或二者的共同作用是否会受到生态系统建植时间的影响? 一年生与多年生间混作系统植物多样性-生产力关系的本质是否是一致的? 这不仅影响了我们理解植物多样性-生态功能关系的内涵与实质, 还影响了农业耕作实践中人们的选择.Zhang和Zhang (
2006 )经过文献统计与分析, 发现物种丰富度-生产力正相关和正选择效应主要出现在一年的试验中, 无论其建植的是一年生植物(江小雷等,
2005 ,
2010 ), 还是多年生植物(Nyfeler
et al. ,
2009 ; 李昂等,
2012 ); 长期的试验结果往往发现多年生草地中物种丰富度-生产力无显著关系(王海东等,
2013 ; 陈宁等,
2018 ).Tilman等(
2001 )发现, 随着多年生植物生长时间延长, 草地群落发生超产现象的比例、强度以及正多样性效应均呈增加趋势.而Mulder等(
2002 )则发现, 豆科植物在试验初期起到了正选择效应的作用, 物种丰富度-生产力呈正相关关系; 随着试验时间的延长, 豆科植物的正选择效应逐渐减弱.在多年生草地中, 群落稳定性是一个时间函数(张景慧和黄永梅,
2016 ), 不仅受到物种丰富度和均匀度的影响(Isbell
et al. ,
2009 ), 超产现象的存在还对多年生草地的功能稳定性起到了一定作用(Hector
et al. ,
2010 ; de Mazancourt
et al. ,
2013 ).Biodiversity- Ecosystem Functioning (BEF)项目通过多年的研究也发现, 森林中植物多样性与生态系统功能的关系也受到群落建成时间的影响(Schuldt
et al. ,
2015 ).由此可见, 通过多年的长期试验, 同时对比一年生间混作和多年生间混作系统的选择效应、互补效应或二者的共同作用, 对于正确理解建植时间对植物多样性-生产力关系的影响, 具有十分重要的科学意义. ...
禾豆间距对间作豌豆“氮阻遏”减缓效应的影响
2
2016
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
禾豆间距对间作豌豆“氮阻遏”减缓效应的影响
2
2016
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
... 物种多样性包括物种丰富度或指定区域内的物种数量, 还包括物种均匀度或群落中物种间生物量分布(Wilsey & Potvin,
2000 ).物种均匀度不仅是功能多样性的重要组分(陈超等,
2016 ), 还是影响种群间变异和群落变异的重要因素(Thibaut & Connolly,
2013 ).对于豆禾混作系统来说, 物种的混播比例, 特别是豆科牧草的混播比例可以作为物种均匀度表征物种多样性或功能多样性(Nyfeler
et al. ,
2009 ; Küchenmeister
et al. ,
2012 ).本试验中, 物种均匀度与生产力的关系也呈单峰“饱和上升型”模式(2013、2014年和3年平均值), 即当豆科牧草混播比例为5:5时群落生产力达到峰值(
图2 ;
表4 ).这种情景下, 增加豆科牧草比例, 期望通过此措施增加群体养分(主要是土壤氮养分)供应达到增加群落生产力的目的无法实现(赵财等,
2016 ).随着生长年限的延长, 物种均匀度与生产力的关系也发生了变化, 但大部分混播组合呈单峰“饱和上升型”模式(
图2 ;
表4 ).陈超等(
2016 )指出, 当物种均匀度的增加程度大于多性状种间差异的减少程度(物种丰富度减少)时, 功能多样性还可增加, 进而导致生产力增加.本试验中物种丰富度影响了牧草产量(生产力)、超产效应和多样性效应的变化, 而物种均匀度和物种均匀度与物种丰富度的互作效应均没有影响上述指标的变化(
表3 ).因而, 本试验中均匀度的增加程度并没有影响到功能多样性, 从而增加群落生产力.这可能与本试验设计的豆禾混播比例较高有关. ...
低氮环境下燕麦与箭筈豌豆间混作与刈割时间对饲草产量及质量特性的影响
2
2006
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
低氮环境下燕麦与箭筈豌豆间混作与刈割时间对饲草产量及质量特性的影响
2
2006
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
... 超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据(Hector
et al. ,
1999 ; Tilman
et al. ,
2001 ; Beckage & Gross,
2006 ; Li
et al. ,
2014 ).对于农业生产来说, 超产效应1 (
OV 1 , 混播/混作的产量高于组分中产量最高的单播/单作)的发现是具有重要实践意义的, 其可用于指导提高土地产出效率(赵彩霞等,
2006 ), 还可应用在提高土壤养分的利用效率(Li
et al. ,
2014 )和土地空间的利用效率(焦念元等,
2006 )等方面.现有的半人工或人工草地以及农作物间混作试验发现的超产效应, 并由此推断植物多样性与群落或生态系统生产力正相关的研究中, 很多是以建植当年或以一年生植物/作物的表现作为证据(Li
et al. ,
2007 ; 江小雷等,
2010 ; 王宇坤等,
2019 ).如果以多年生植物建植的混播/混作群落, 则发现随着生长时间的延长, 超产效应和多样性效应均会发生变化.本试验中, 2013年所有混播组合均产生了超产效应, 2014年所有的多年生牧草混播组合也均产生了超产效应, 2015年则只有PM2-D和PM6产生了超产效应, 且超产效应呈下降趋势(
表2 ); 2015年不同混播种类和混播比例的
OV 1 均小于2013和2014年, 2014年不同混播种类和混播比例的
OV 2 则小于2013和2015年(
表1 ).相关研究也证实, 生长多年后, 有些物种会几乎消失在混播群落中, 有些物种则产量占比(相对产量)大幅减少(Küchenmeister
et al. ,
2012 ), 那么建植早期的由强优势物种主导的选择效应和
OV 1 就会减弱; 有些物种建植前期表现较好, 有些物种则在整个观测期内表现稳定(Frankow-Lindberg
et al. ,
2009 ),
OV 2 和选择效应则变化较小.因此, 混播物种属性(选择效应)及混播后物种间相互作用类型、强度(互补效应)的变化影响了超产效应.本试验中, 2013年不同混播种类和比例的互补效应大于2014和2015年, 选择效应也是大于2014和2015年(除AM2外), 多样性净效应则与互补效应表现出相似的规律(
图1 ,
图4 ).因而, 本试验的互补效应、选择效应和多样性净效应均表现出随生长时间的延长而下降的趋势, 进而导致了超产效应的下降和超产效应稳定性的下降(
图5 ).这与Tilman等(
2001 )和李昂等(
2012 )的研究结果不一致, 这可能与上述群落受到了土壤肥力的影响(施肥)有关. ...
氮素添加对豆禾牧草种间竞争力与生产性能的影响
2
2015
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
... ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
氮素添加对豆禾牧草种间竞争力与生产性能的影响
2
2015
... 在生产实践中, 一年生间混作应用范围更广: 在生长季节较短的地区, 可用于改善生态系统氮营养平衡(赵彩霞等,
2006 ; 刘文辉等,
2017 ), 促进草地动物蛋白质的形成(Russelle,
1992 ), 满足家畜冷季对饲草的需求(马春晖等,
1999 ; Lithourgidis
et al. ,
2006 ); 在生长季节较长, 光热资源较好的地区, 可与农作物轮作倒茬、复播, 对建立稳产、高产、高效和可持续的农业种植体系具有重要作用(杨学超等,
2012 ).多年生间混作建植的初期(第1年)生长速度较慢, 难以在初期获得较高的经济产量, 其高产年份往往在建植的中期(第2-4年)(郑伟等,
2015 ), 相对来说占地时间长(常生华等,
2004 ), 往往难与常规作物进行轮作倒茬、复播(陈远学等,
2013 ), 需要与常规作物“争地”, 限制了其应用范围.但如果多年生间混作系统具有较好的生态功能稳定性, 其经济产量/品质稳定性较好, 则适合长期利用, 与一年生间混作系统相比则具有成本优势, 节省了种子、春季播种和秋季翻耕等费用, 多年综合收益较高(胡自治等,
2001 ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
... ; 郑伟等,
2015 ).因此, 通过多年的长期试验, 同时对比一年生和多年生间混作系统的超产效应, 对于在生产实践中选择适宜的间混作系统类型, 具有十分重要的实践意义. ...
不同混播方式豆禾混播草地生产性能的综合评价
1
2012
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
不同混播方式豆禾混播草地生产性能的综合评价
1
2012
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
全尺度人工湿地中植物多样性对生产力与多样性效应的影响
4
2014
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ; 朱四喜等,
2014 ). ...
全尺度人工湿地中植物多样性对生产力与多样性效应的影响
4
2014
... 与此同时, 生长时间还影响了超产效应与多样性效应的关系.本试验中, 超产效应与互补效应呈正相关关系, 与选择效应和多样性净效应则呈负相关关系(
图3 ), 且选择效应值小于互补效应(
图1 ), 说明互补效应是影响超产的主要贡献者.该结果得到了许多实验的证实(Loreau & Hector,
2001 ; 代巍等,
2009 ; 李昂等,
2012 ).但
OV 1 与互补效应呈负相关关系, 与选择效应和多样性净效应则呈正相关关系(
图3 ), 且随着生长年限的延长, 超产效应与多样性效应的变化趋势相同(
图1 ,
图4 ), 这表明在多年生豆禾混播系统建植前期, 选择效应对生态系统功能的响应优先于互补效应, 但互补效应的稳定性优于选择效应.故而在建植后期, 互补效应的作用则更为关键.朱四喜等(
2014 )则发现在物种多样性与生态功能的关系中选择效应比互补效应更普遍, 与本试验的结果基本吻合.本试验中的一年生豆禾混播草地的超产效应和多样性净效应年际间变化呈“V”形变化, 互补效应和选择效应则向趋异方向变化(
图1 ;
表1 ).一年生豆禾混播草地的超产效应主要受互补效应的影响, 与多年生豆禾混播草地基本一致. ...
... 提高表现效应(超产效应)是物种丰富度调控群落功能(生产力和稳定性)的内在机制之一, 而超产效应的发生机制与选择效应和互补效应有着密切的关系(Hector
et al. ,
2010 ; 张景慧和黄永梅,
2016 ).物种丰富度对群落生产功能的研究尚未明确解析出普遍性结论, 但是在草地生态系统和农作物系统上, 物种丰富度对群落生产力存在正效应得到了多数****的证实(Hector
et al. ,
1999 ; 江小雷等,
2005 ; Finn
et al. ,
2013 ; Li
et al. ,
2014 ).本研究也进一步证实多年生混播草地物种丰富度在一定程度上对群落生产性能具有促进作用, 其变化规律呈单峰格局(2013、2014年和3年平均值), 较高物种丰富度具有较高生产力(
图2 ), 这与江小雷等(
2005 ,
2010 )、朱四喜等(
2014 )的研究结果一致.这种物种丰富度与生产力关系的“饱和上升型”模式, 则说明人工草地生产力在较低物种丰富度时就达到饱和(Hooper
et al. ,
2005 ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ; 朱四喜等,
2014 ).但年际间物种丰富度和群落生产力之间的关系是有波动的, 2015年则呈双峰格局(
图2 ).由于混播物种间的竞争作用和物种本身属性(如生长速度、生长寿命、对不良环境的耐受性能等)会随着生长年限的延长而产生变化, 导致物种属性及其相互作用成为控制生长后期物种丰富度-生产力关系的主导因素(Pacala & Tilman,
2001 ; Hector
et al. ,
2010 ; 朱四喜等,
2014 ). ...
... ; 朱四喜等,
2014 ). ...
混播方式对豆禾混播草地植物根系构型特征的影响
1
2018
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
混播方式对豆禾混播草地植物根系构型特征的影响
1
2018
... 在豆科与禾本科植物间混作系统中, 间混作的豆禾植物在形态(焦念元等,
2006 ; 王平等,
2009 )、生长发育节律(Cox
et al. ,
2017 )、土壤养分资源利用(Li
et al. ,
2007 ,
2014 ; 王平等,
2010 )、根系分布层位(宋日等,
2002 ; 朱亚琼等,
2018 )上都存在着相互补充而非彼此竞争的可能(谢开云等,
2013 ), 因而, 豆禾间作系统通过改善其土壤养分(特别是氮)供给(肖焱波等,
2005 ; Li
et al. ,
2011 ; 赵财等,
2016 ), 提高了系统的产量或质量(王平等,
2007 ; 郑伟等,
2012 ; 陈远学等,
2013 ).与此同时, 增加半人工草地或栽培草地物种多样性可以改善资源利用效率(空间生态位的差异), 增加种间正相互作用(Hendriks
et al. ,
2013 )和选择效应(选择了高产物种)的可能(Hooper
et al. ,
2005 ; Finn
et al. ,
2013 ), 从而使物种/种群丰富度更高的系统生产力更高(Cardinale
et al. ,
2007 ; Atwater & Callaway,
2015 ).这种比单播/单作系统生产力更高的现象称作超产效应(Loreau,
1998 ; Nyfeler
et al. ,
2009 ; Hector
et al. ,
2010 ), 在土壤肥力一般的半人工草地(Cardinale
et al. ,
2007 )、施肥的栽培草地(Nyfeler
et al. ,
2009 ; 江小雷等,
2010 ; Finn
et al. ,
2013 )和豆科与禾本科作物间混作的农作系统(Li
et al. ,
2003 ,
2007 ,
2011 )均有报道.依据物种生态位添加分离方法, 将混播/混作群落生产力的增加量(多样性净效应)分解为群落互补效应(
CE )和选择效应(
SEF ), 其中选择效应是指具有特殊性质的物种在群落占优势会影响生态系统过程, 而互补效应是指不同的物种具有特定的生态位, 实现了资源的合理分配, 导致资源利用总量增加(Loreau & Hector,
2001 ).超产效应的产生可能是选择了高产物种/种群的结果(Grime,
1998 ; Huston
et al. ,
2000 ; Atwater & Callaway,
2015 ), 物种/种群属性(选择效应)控制着生态系统功能(Hooper
et al. ,
2005 ; Küchenmeister
et al. ,
2012 ; 王海东等,
2013 ); 也可能是间混作物种间生态位互补作用的结果(Tilman & Downing,
1994 ; Hector
et al. ,
1999 ; Nyfeler
et al. ,
2009 ), 互补效应决定着生态系统生产力(李昂等,
2012 ; Finn
et al. ,
2013 ; 王宇坤等,
2019 ); 或是二者共同起作用的结果(Loreau & Hector,
2001 ; Pacala & Tilman,
2001 ; Mulder
et al. ,
2002 ).因此, 在农业生态系统中依据植物多样性-生产力关系的选择效应、互补效应或二者的共同作用, 使其产量、资源利用效率得到提升, 已成为促进农业可持续发展的重要措施(Finn
et al. ,
2013 ). ...
 ,
, ,1,2,*, 朱亚琼1, 王祥1, 马军1, 朱进忠1,2
,1,2,*, 朱亚琼1, 王祥1, 马军1, 朱进忠1,2

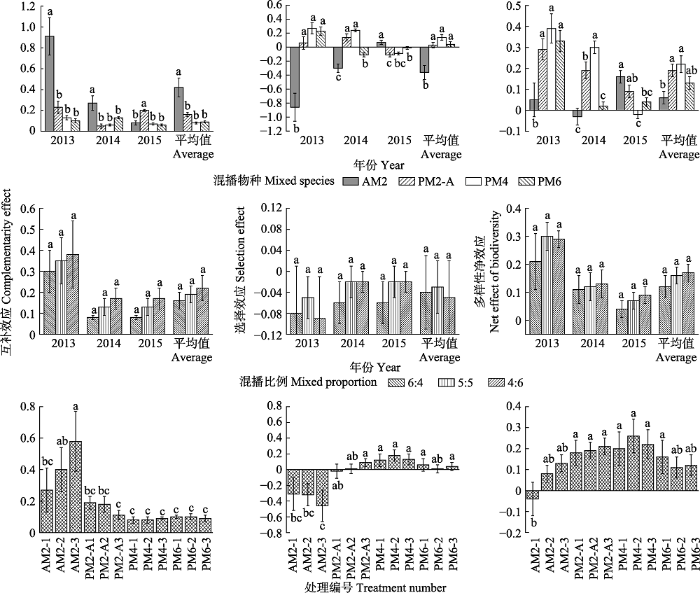 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT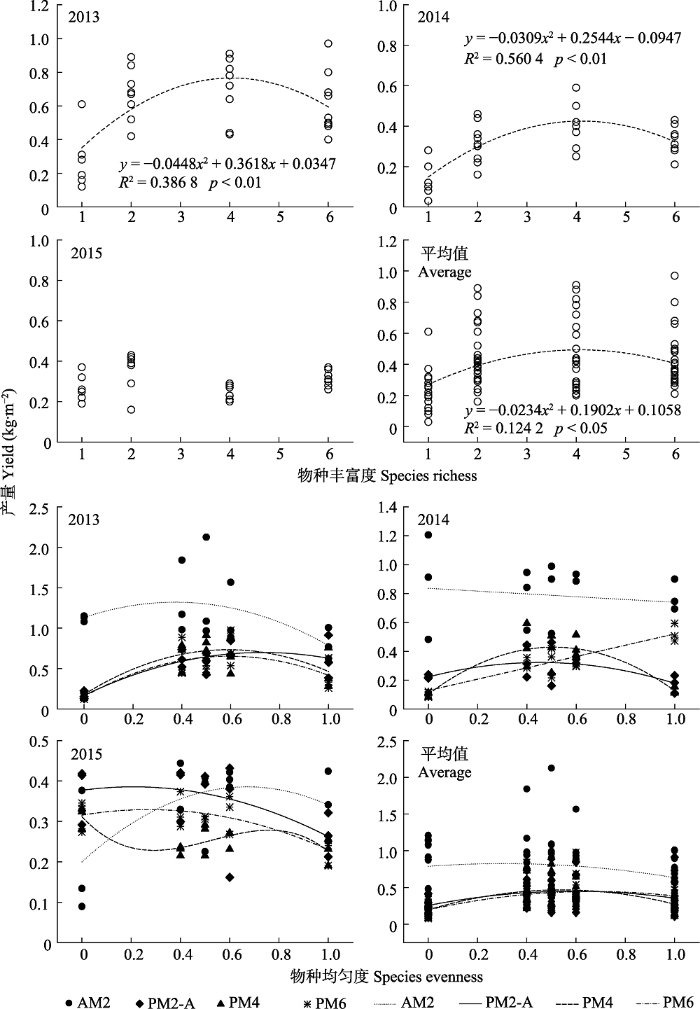 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT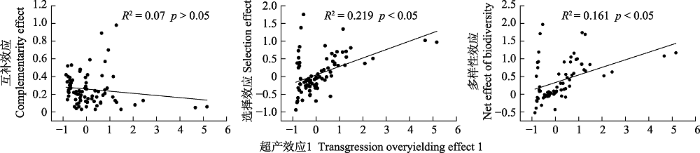 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT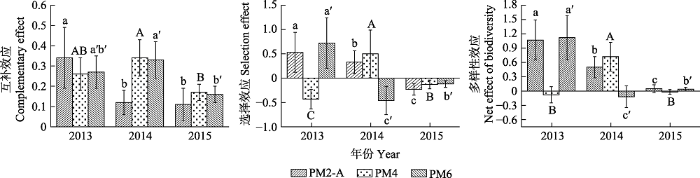 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT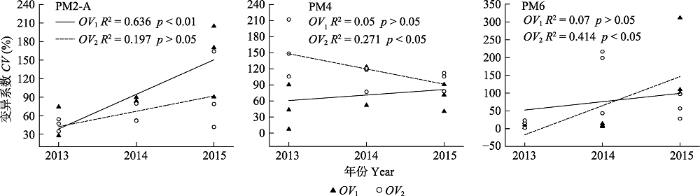 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}