Key characteristics for facilitating Leucaena leucocephala to successfully invade pioneer communities of tropical rain forests
Jin-Huan LUO1,*, Zhao-Yuan TAN2,3,*, Bin CHEN4, Guang-Wu CHEN4, Kai JIANG2,3, Qi-Fang HEI2,3, Hui ZHANG,2,3,**1Sanya Academy of Forestry, Sanya, Hainan 572000, China 2College of Forestry/Wuzhishan National Forest Ecosystem Monitoring Research Station, Hainan University, Haikou 570228, China 3Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants (Hainan University), Ministry of Education, College of Forestry, Hainan University, Haikou 570228, China 4Sanya Forestry Bureau, Sanya, Hainan 572000, China
Abstract Aims Due to fast-growing and high drought stress tolerance, Leucaena leucocephala has been widely used for afforestation in degraded tropical forests worldwide, but it is also a global invasive exotic species. Studies have shown that fast-growing can help L. leucocephala successfully invade subtropical forests. In this study, we aimed to investigate whether fast-growing and high drought stress tolerance can help L. leucocephala invade tropical rain forests. Methods The pioneer community of tropical rain forest which had been invaded by L. leucocephala in the Baopoling Mountain, Sanya, China was the research object. Through the t-test, we compared the differences in key functional traits that were related to growth rate (photosynthesis rate, stomatal conductance and transpiration rate) and drought stress tolerance (leaf turgor loss point) in both wet and dry seasons between L. leucocephala and eight dominant native species of pioneer community of tropical rain forest. And the principal component analysis (PCA) was used to investigate whether these functional traits can best discriminate between Leucaena leucocephala and the eight dominant native species. Important findings Leucaena leucocephala could be invariably growing fast (photosynthesis rate, stomatal conductance and transpiration rate much higher than native species) from wet to dry seasons and had higher drought stress tolerance (leaf turgor loss point much lower than native species) in the dry season. The results of PCA showed that these functional traits could significantly discriminate between L. leucocephala and the eight dominant native species. Therefore, invariable fast-growing from wet to dry season and high drought stress tolerance in the dry season make L. leucocephala successfully invade pioneer communities of tropical rain forests. In the future, these functional traits can be used to select many native species to perform biological control of L. leucocephala in other tropical forests. Keywords:drought stress tolerance;fast-growing;functional trait;invasion;pioneer community of tropical rain forest
PDF (14836KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 罗金环, 谭照远, 陈斌, 陈广武, 姜凯, 何奇芳, 张辉. 银合欢成功入侵热带雨林先锋群落的关键因素. 植物生态学报, 2020, 44(12): 1215-1223. DOI: 10.17521/cjpe.2020.0137 LUO Jin-Huan, TAN Zhao-Yuan, CHEN Bin, CHEN Guang-Wu, JIANG Kai, HEI Qi-Fang, ZHANG Hui. Key characteristics for facilitating Leucaena leucocephala to successfully invade pioneer communities of tropical rain forests. Chinese Journal of Plant Ecology, 2020, 44(12): 1215-1223. DOI: 10.17521/cjpe.2020.0137
新窗口打开|下载原图ZIP|生成PPT 图3银合欢和三亚抱坡岭8个本地优势种在干湿季时的4种功能性状之间的差异(平均值+标准误差)。***, p < 0.05; NS, p > 0.05。
Fig. 3Differences in four functional traits between dry and wet season for Leucaena leucocephala and the eight dominant native species in the Baopoling Mountain, Sanya, China (mean + SE). ***, p < 0.05; NS, p > 0.05.
A, 银合欢; B, 土蜜树; C, 美叶菜豆树; D, 赤才; E, 石斑木; F, 翻白叶树; G, 瓜馥木; H, 九节; I, 葨芝。***, p < 0.05; NS, p > 0.05。 Fig. 4Differences in four functional traits between Leucaena leucocephala and each of the eight dominant native plant species in the Baopoling Mountain, Sanya, China (mean + SE).
A, Leucaena leucocephala; B, Bridelia tomentosa; C, Radermachera frondosa; D, Lepisanthes rubiginosa; E, Rhaphiolepis indica; F, Pterospermum heterophyllum; G, Fissistigma oldhamii; H, Psychotria asiatica; I, Maclura cochinchinensis. ***, p < 0.05; NS, p > 0.05
A, 干季。B, 湿季。 Fig. 5Principal component analysis of the four functional traits between Leucaena leucocephala and the eight doimant native plant species in the Baopoling Mountain, Sanya, China.
A, Dry season. B, Wet season.
Table 1 表1 表1银合欢和三亚抱坡岭8个本地优势种基于4种功能性状的主成分分析前两轴 Table 1The first two axes of a principal component analysis for Leucaena leucocephala and the eight dominant native species in the Baopoling Mountain, Sanya, China, based on four plant functional traits
功能性状 Functional trait
轴1 PC1
轴2 PC2
蒸腾速率 Transpiration rate
0.51
-0.16
最大光合速率 Maximum photosynthesis rate
0.77
-0.15
气孔导度 Stomatal conductance
0.58
-0.19
叶片膨压丧失点 Leaf turgor loss point
0.11
0.53
加粗字体表示PC1 > 0.5或PC2 > 0.5。 Bold data means PC1 > 0.5 or PC2 > 0.5.
BarrosV, MeloA, SantosM, NogueiraL, FrosiG, SantosMG (2020). Different resource-use strategies of invasive and native woody species from a seasonally dry tropical forest under drought stress and recovery Plant Physiology and Biochemistry, 147, 181-190. URLPMID:31865164 [本文引用: 1]
BartlettMK, ScoffoniC, SackL (2012). The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis Ecology Letters, 15, 393-405. URLPMID:22435987 [本文引用: 1]
BartlettMK, ZhangY, YangJ, KreidlerN, SunSW, LinL, HuYH, CaoKF, SackL (2015). Drought tolerance as a driver of tropical forest assembly: resolving spatial signatures for multiple processes Ecology, 97,503-514. DOI:10.1890/15-0468.1URLPMID:27145624 [本文引用: 1] 1 cm dbh) in a 20-ha seasonal tropical rainforest plot in Xishuangbanna, China, we compared the ability of drought-tolerance traits, other physiological traits, and commonly measured functional traits to predict the spatial patterns expected from the assembly processes of habitat associations, niche-overlap-based competition, and hierarchical competition. We distinguished the neighborhood-scale (0-20 m) patterns expected from competition from larger-scale habitat associations with a wavelet method. Species' drought tolerance and habitat variables related to soil water supply were strong drivers of habitat associations, and drought tolerance showed a significant spatial signal for influencing competition. Overall, the traits most strongly associated with habitat, as quantified using multivariate models, were leaf density, leaf turgor loss point (pi(tlp); also known as the leaf wilting point), and stem hydraulic conductivity (r2 range for the best fit models = 0.27-0.36). At neighborhood scales, species spatial associations were positively correlated with similarity in pi(tlp), consistent with predictions for hierarchical competition. Although the correlation between pi(tlp) and interspecific spatial associations was weak (r2
BlackmanCJ, BrodribbTJ, JordanGJ (2010). Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms New Phytologist, 188, 1113-1123. [本文引用: 1]
BucciSJ, Carbonell-SillettaLM, GarréA, CavallaroA, EfronST, AriasNS, GoldsteinG, ScholzFG (2019). Functional relationships between hydraulic traits and the timing of diurnal depression of photosynthesis Plant, Cell & Environment, 42,1603-1614. URLPMID:30613989 [本文引用: 1]
ChiouCR, ChenYJ, WangHH, GrantWE (2016). Predicted range expansion of the invasive plant Leucaena leucocephala in the Hengchun Peninsula, Taiwan Biological Invasions, 18, 381-394. [本文引用: 1]
dosSantos VAHF, FerreiraMJ, RodriguesJVFC, GarciaMN, CeronJVB, NelsonBW, SaleskaSR (2018). Causes of reduced leaf-level photosynthesis during strong El Ni?o drought in a Central Amazon forest Global Change Biology, 24, 4266-4279. URLPMID:29723915 [本文引用: 1]
FunkJL, ClelandEE, SudingKN, ZavaletaES (2008). Restoration through reassembly: plant traits and invasion resistance Trends in Ecology & Evolution, 23, 695-703. URLPMID:18951652 [本文引用: 4]
GibsonAH, DreyfusBL, DommerguesYR (1982). Nitrogen Fixation by Legumes in the Tropics Microbiology of Tropical Soils and Plant Productivity, Springer, Dordrecht, the Netherlands. [本文引用: 1]
GoelVL, BehlHM (2002). Selection of Leucaena species for afforestation and amelioration of sodic soils Land Degradation & Development, 13, 387-393. [本文引用: 1]
HuaL, ChenY, ZhangH, FuP, FanZ (2017). Stronger cooling effects of transpiration and morphology of the plants from a hot dry habitat than from a hot wet habitat Functional Ecology, 31, 2202-2211. [本文引用: 1]
HulmePE, Py?ekP, Jaro?íkV, PerglJ, SchaffnerU, VilaM (2013). Bias and error in understanding plant invasion impacts Trends in Ecology & Evolution, 28, 212-218. URLPMID:23153723 [本文引用: 1]
IshiharaKL, HondaMD, BageelA, BorthakurD (2018). Leucaena leucocephala: a leguminous tree suitable for eroded habitats of Hawaiian islands Ravine Lands: Greening for Livelihood and Environmental Security, 413-431. [本文引用: 1]
JiZH, FangHD, YangYX, PanZX, YueXW, LiJZ (2010). Mechanism research of gully controlling with Leucaena leucocephala in the Yuanmou Arid Hot Valley Journal of Soil and Water Conservation, 24, 19-22, 26. [本文引用: 1]
KirschbaumMU (2011). Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies Plant Physiology, 155, 117-124. URLPMID:21088226 [本文引用: 2]
KüsterEC, KühnI, BruelheideH, KlotzS (2008). Trait interactions help explain plant invasion success in the German flora Journal of Ecology, 96, 860-868. [本文引用: 1]
LaughlinDC (2014). Applying trait-based models to achieve functional targets for theory-driven ecological restoration Ecology Letters, 17, 771-784. DOI:10.1111/ele.12288URLPMID:24766299 [本文引用: 1] Manipulating community assemblages to achieve functional targets is a key component of restoring degraded ecosystems. The response-and-effect trait framework provides a conceptual foundation for translating restoration goals into functional trait targets, but a quantitative framework has been lacking for translating trait targets into assemblages of species that practitioners can actually manipulate. This study describes new trait-based models that can be used to generate ranges of species abundances to test theories about which traits, which trait values and which species assemblages are most effective for achieving functional outcomes. These models are generalisable, flexible tools that can be widely applied across many terrestrial ecosystems. Examples illustrate how the framework generates assemblages of indigenous species to (1) achieve desired community responses by applying the theories of environmental filtering, limiting similarity and competitive hierarchies, or (2) achieve desired effects on ecosystem functions by applying the theories of mass ratios and niche complementarity. Experimental applications of this framework will advance our understanding of how to set functional trait targets to achieve the desired restoration goals. A trait-based framework provides restoration ecology with a robust scaffold on which to apply fundamental ecological theory to maintain resilient and functioning ecosystems in a rapidly changing world.
LawsonT, Vialet-ChabrandS (2019). Speedy stomata, photosynthesis and plant water use efficiency New Phytologist, 221, 93-98. [本文引用: 1]
LiHS, CaiHJ, LiJM, DengHY, ZhongBL (2015). A preliminary study on the alien invasive plant species in Huangshanlu Forest Park, Guangzhou Journal of Guangdong University of Education, 35(5), 73-77. [本文引用: 1]
LiuF, GaoC, ChenM, LiK (2018). Above- and below-ground biomass relationships of Leucaena leucocephala(Lam.) de Wit in different plant stands PLOS ONE, 13, e0207059. DOI: 10.1371/journal.pone.0207059. [本文引用: 2]
LuoHX, DaiSP, LiMF, XieZH (2018). Analysis on climate change characteristics of Hainan Island from 1959 to 2015 Jiangsu Agricultural Sciences, 46, 261-268. [本文引用: 2]
MacArthurR, LevinsR (1967). The limiting similarity, convergence, and divergence of coexisting species The American Naturalist, 101, 377-385. [本文引用: 2]
MaheraliH, SherrardME, CliffordMH, LattaRG (2008). Leaf hydraulic conductance and photosynthesis are genetically correlated in an annual grass New Phytologist, 180, 240-247. [本文引用: 1]
McAuslandL, Vialet-ChabrandS, DaveyP, BakerNR, BrendelO, LawsonT (2016). Effects of kinetics of light-induced stomatal responses on photosynthesis and water-use efficiency New Phytologist, 211, 1209-1220. [本文引用: 1]
MitchellPJ, VeneklaasEJ, LambersH, BurgessSS (2008). Leaf water relations during summer water deficit: differential responses in turgor maintenance and variation in leaf structure among different plant communities in south- western Australia Plant, Cell & Environment, 31, 1791-1802. URLPMID:18761698 [本文引用: 1]
MoranVC, HoffmannJH, ZimmermannHG (2005). Biological control of invasive alien plants in South Africa: necessity, circumspection, and success Frontiers in Ecology and the Environment, 3, 71-77. [本文引用: 1]
OstertagR, WarmanL, CordellS, VitousekPM (2015). Using plant functional traits to restore Hawaiian rainforest Journal of Applied Ecology, 52, 805-809. [本文引用: 1]
PengSH, WangHH, KuoYL (2019). Methods for preventing the invasion of Leucaena leucocephala in coastal forests of the Hengchun Peninsula, Taiwan Taiwan Journal of Forest Science, 34, 99-112. [本文引用: 3]
PengY (2008). Study on Diversity and Invasion in Plantation of Alien Tree Species—Take Eucalyptus spp. Acacia dealbata, Leucaena leucocephala for Example Master degree dissertation, Southwest Forestry University, Kunming. [本文引用: 1]
RichardsonDM, RejmánekM (2011). Trees and shrubs as invasive alien species—A global review Diversity and Distributions, 17, 788-809. [本文引用: 1]
RowlandL, daCosta ACL, OliveiraAAR, AlmeidaSS, MeirP (2018). Shock and stabilisation following long-term drought in tropical forest from 15 years of litterfall dynamics Journal of Ecology, 106, 1673-1682. [本文引用: 1]
SackL, CowanP, JaikumarN, HolbrookN (2003). The “hydrology” of leaves: co-ordination of structure and function in temperate woody species Plant, Cell & Environment, 26, 1343-1356. [本文引用: 1]
SchulteP, HinckleyT (1985). A comparison of pressure-volume curve data analysis techniques Journal of Experimental Botany, 36, 1590-1602. [本文引用: 1]
SeastedtTR (2015). Biological control of invasive plant species: a reassessment for the Anthropocene New Phytologist, 205, 490-502. [本文引用: 1]
vanKleunen M, WeberE, FischerM (2010). A meta-analysis of trait differences between invasive and non-invasive plant species Ecology Letters, 13, 235-245. URLPMID:20002494 [本文引用: 1]
WangC, ZhangH, LiuH, JianS, YanJ, LiuN (2020). Application of a trait-based species screening framework for vegetation restoration in a tropical coral island of China Functional Ecology, 267, 25-34. [本文引用: 2]
WangW, ZhangXM, ShaLH Cheng LS (2007). Roster of alien: invasive perilous species of animals and plants in Hainan Island (one) Chinese Journal of Tropical Agriculture, 27, 58-64. [本文引用: 3]
WolfeBT, vanBloem SJ (2012). Subtropical dry forest regeneration in grass-invaded areas of Puerto Rico: understanding why Leucaena leucocephala dominates and native species fail Forest Ecology and Management, 267, 253-261. [本文引用: 5]
ZhangH, ChenHYH, LianJ, ChandranRJ, LiRH, LiuH, YeW, BerningerF, YeQ (2018a). Using functional trait diversity patterns to disentangle the scale-dependent ecological processes in a subtropical forest Functional Ecology, 32, 1379-1389. [本文引用: 1]
ZhangH, ChandranRJ, ZhuS, LiuH, XuQ, QiW, LiuK, ChenHYH, YeQ (2018b). Shifts in functional trait-species abundance relationships over secondary subalpine meadow succession in the Qinghai-Tibetan Plateau Oecologia, 188, 547-557. URLPMID:30043232 [本文引用: 3]
ZhouLP, HeYJ, MaHC, ZhuCF, GaoZ, LiW, LiFX (2010). Effects of fertilization treatment on seedlings drought tolerance of Leucaena leucephala under different water conditions Jiangxi Science, 28, 311-319, 358. [本文引用: 1]
ZhuSD, SongJJ, LiRH, YeQ (2013). Plant hydraulics and photosynthesis of 34 woody species from different successional stages of subtropical forests Plant, Cell & Environment, 36, 879-891. URLPMID:23057774 [本文引用: 2]
ZouHP, ZhangJH, ChenXM, LiuSJ, LiWG (2015). Spatiotemporal change characteristics of agricultural climate resources in Hainan Island Chinese Journal of Agrometeorology, 36, 417-427. [本文引用: 1]
Speedy stomata, photosynthesis and plant water use efficiency 1 2019
... 光合速率会影响植物的能量平衡, 植物的光合速率越高, 其生长速度就越快(Kirschbaum, 2011; Zhang et al., 2018b).研究发现, 无论是在湿季还是干季, 银合欢的光合速率都比其他8个本地优势种高5-10倍.而且植物光合速率较高也会导致植物拥有较高的气孔性状值(McAusland et al., 2016; Lawson & Vialet-Chabrand, 2019)和蒸腾速率(Maherali et al., 2008; dos Santos et al., 2018; Bucci et al., 2019).结果显示无论在湿季还是干季, 银合欢的气孔导度和蒸腾速率也均比其他8个本地优势种高(5-10倍).这些结果表明, 银合欢比本地优势种拥有更适应当地环境的叶片性状, 可以最大限度地提高光合速率和整体生长速度.众所周知, 水是限制植物最大光合速率的关键因素(Guan et al., 2015).由于此热带雨林先锋群落干季的降水量仅为湿季的1/10 (罗红霞等, 2018), 所以植物的光合速率会受到抑制.然而, 研究结果发现银合欢的生长速率在干季和湿季均较高, 差异不大, 而8个本地优势种的生长速率在干季则会受到抑制.因此, 研究结果进一步表明, 较高的生长速率是一个关键因素, 使银合欢优于其他8个本地优势种, 从而更好地生长在这片热带雨林先锋群落中.已有研究在亚热带的干旱森林中观察到, 快速的生长可以成功地帮助银合欢竞争过本地优势种, 从而成功入侵亚热带森林(Wolfe & van Bloem, 2012).我们的结果也表明高生长速率能帮助银合欢成功入侵热带雨林先锋群落. ...
广州南沙黄山鲁森林公园外来入侵植物初步研究 1 2015
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
广州南沙黄山鲁森林公园外来入侵植物初步研究 1 2015
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
Above- and below-ground biomass relationships of Leucaena leucocephala(Lam.) de Wit in different plant stands 2 2018
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... ; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
Leaf hydraulic conductance and photosynthesis are genetically correlated in an annual grass 1 2008
... 光合速率会影响植物的能量平衡, 植物的光合速率越高, 其生长速度就越快(Kirschbaum, 2011; Zhang et al., 2018b).研究发现, 无论是在湿季还是干季, 银合欢的光合速率都比其他8个本地优势种高5-10倍.而且植物光合速率较高也会导致植物拥有较高的气孔性状值(McAusland et al., 2016; Lawson & Vialet-Chabrand, 2019)和蒸腾速率(Maherali et al., 2008; dos Santos et al., 2018; Bucci et al., 2019).结果显示无论在湿季还是干季, 银合欢的气孔导度和蒸腾速率也均比其他8个本地优势种高(5-10倍).这些结果表明, 银合欢比本地优势种拥有更适应当地环境的叶片性状, 可以最大限度地提高光合速率和整体生长速度.众所周知, 水是限制植物最大光合速率的关键因素(Guan et al., 2015).由于此热带雨林先锋群落干季的降水量仅为湿季的1/10 (罗红霞等, 2018), 所以植物的光合速率会受到抑制.然而, 研究结果发现银合欢的生长速率在干季和湿季均较高, 差异不大, 而8个本地优势种的生长速率在干季则会受到抑制.因此, 研究结果进一步表明, 较高的生长速率是一个关键因素, 使银合欢优于其他8个本地优势种, 从而更好地生长在这片热带雨林先锋群落中.已有研究在亚热带的干旱森林中观察到, 快速的生长可以成功地帮助银合欢竞争过本地优势种, 从而成功入侵亚热带森林(Wolfe & van Bloem, 2012).我们的结果也表明高生长速率能帮助银合欢成功入侵热带雨林先锋群落. ...
Effects of kinetics of light-induced stomatal responses on photosynthesis and water-use efficiency 1 2016
... 光合速率会影响植物的能量平衡, 植物的光合速率越高, 其生长速度就越快(Kirschbaum, 2011; Zhang et al., 2018b).研究发现, 无论是在湿季还是干季, 银合欢的光合速率都比其他8个本地优势种高5-10倍.而且植物光合速率较高也会导致植物拥有较高的气孔性状值(McAusland et al., 2016; Lawson & Vialet-Chabrand, 2019)和蒸腾速率(Maherali et al., 2008; dos Santos et al., 2018; Bucci et al., 2019).结果显示无论在湿季还是干季, 银合欢的气孔导度和蒸腾速率也均比其他8个本地优势种高(5-10倍).这些结果表明, 银合欢比本地优势种拥有更适应当地环境的叶片性状, 可以最大限度地提高光合速率和整体生长速度.众所周知, 水是限制植物最大光合速率的关键因素(Guan et al., 2015).由于此热带雨林先锋群落干季的降水量仅为湿季的1/10 (罗红霞等, 2018), 所以植物的光合速率会受到抑制.然而, 研究结果发现银合欢的生长速率在干季和湿季均较高, 差异不大, 而8个本地优势种的生长速率在干季则会受到抑制.因此, 研究结果进一步表明, 较高的生长速率是一个关键因素, 使银合欢优于其他8个本地优势种, 从而更好地生长在这片热带雨林先锋群落中.已有研究在亚热带的干旱森林中观察到, 快速的生长可以成功地帮助银合欢竞争过本地优势种, 从而成功入侵亚热带森林(Wolfe & van Bloem, 2012).我们的结果也表明高生长速率能帮助银合欢成功入侵热带雨林先锋群落. ...
Leaf water relations during summer water deficit: differential responses in turgor maintenance and variation in leaf structure among different plant communities in south- western Australia 1 2008
... 银合欢干季的叶片膨压丧失点明显低于8个本地优势种(1/2), 这说明在该热带雨林先锋群落中银合欢的耐旱性比本地优势种的更强.研究发现, 在面临非常有限的水分供应时, 只有叶片膨压丧失点更低的植物才会拥有更强的耐旱性, 从而维持更高的气孔导度、光合速率和蒸腾速率(Sack et al., 2003; Mitchell et al., 2008; Blackman et al., 2010).这可能解释了干季时期银合欢的光合速率、气孔导度和蒸腾速率仍比8个本地优势种要高得多的原因.因此银合欢能优于本地优势种的关键可能是其在干季具有更强的耐旱性, 可以在水分供应不足时仍然保持较高的生长速率. ...
Biological control of invasive alien plants in South Africa: necessity, circumspection, and success 1 2005
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
Using plant functional traits to restore Hawaiian rainforest 1 2015
... 利用功能性状找出外来入侵植物优于本地植物的关键特征, 可以揭示为什么外来入侵植物能成功入侵本地生态系统(Küster et al., 2008; van Kleunen et al., 2010; Hulme et al., 2013; Ostertag et al., 2015).一般而言, 入侵植物想要入侵成功, 其功能性状不仅要具有比本地优势物种更好地适应本地生长环境的能力, 而且要与本地优势物种完全不相似(Funk et al., 2008).功能性状如叶片膨压丧失点和光合速率分别是耐旱性和快速生长的良好指标(Kirschbaum, 2011; Zhang et al., 2018a), 因此研究银合欢生长速率和耐旱性的功能性状能帮助我们更好地了解银合欢适应本地生长环境的能力及其与本地优势物种的功能性状不相似性.相似性限制假说表明, 如果本地植物在功能上与外来植物相似, 会和外来植物竞争资源, 从而具有消除外来植物的巨大潜力(MacArthur & Levins, 1967).因此, 通过功能性状找到的外来入侵植物优于本地物种的关键特征, 也可以筛选出一些与外来入侵植物功能性状相似的本地植物, 用来防治外来入侵植物(Funk et al., 2008). ...
Methods for preventing the invasion of Leucaena leucocephala in coastal forests of the Hengchun Peninsula, Taiwan 3 2019
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... ; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... ; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
外来树种人工林下植物多样性及其入侵性研究——以桉树、银荆、银合欢为例 1 2008
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
外来树种人工林下植物多样性及其入侵性研究——以桉树、银荆、银合欢为例 1 2008
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
Trees and shrubs as invasive alien species—A global review 1 2011
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
Shock and stabilisation following long-term drought in tropical forest from 15 years of litterfall dynamics 1 2018
... 银合欢自身的固氮能力可以让其具有很高的生长速率进而比本地优势物种能更好地适应本地生长环境(Gibson et al., 1982; Wolfe & van Bloem, 2012).但是热带季风气候区通常具有明显的干湿季(Rowland et al., 2018), 从而会限制银合欢的生长.我们假设银合欢的高生长速率和强耐旱性不仅能比本地优势物种在干湿季都更好地适应本地环境, 而且其功能性状与本地优势物种完全不相似.为验证这一假设, 本研究以位于中国三亚抱坡岭被银合欢成功入侵的热带雨林先锋群落为研究对象, 测定了银合欢和本地优势种干季和湿季期间与生长速率(光合速率、气孔导度和蒸腾速率)和耐旱性(叶片膨压丧失点)高度相关的关键功能性状.利用这些功能性状我们可以探究: 1)银合欢的高生长速率是否比本地优势种在干季与湿季更加适应本地环境? 2)银合欢的高生长速率和强耐旱性的功能性状是否与本地优势物种完全不相似? 目前银合欢是海南危害严重的外来入侵植物之一(王伟等, 2007).如果这些功能性状确实能使银合欢优于本地优势种最终成功入侵热带雨林先锋群落, 我们也可利用这些性状和我们开发出的筛选本地物种的软件(Wang et al., 2020), 从三亚和整个海南岛的植物物种库中筛选出多种合适的本地物种对整个海南岛成功入侵热带森林的银合欢进行生物防治. ...
The “hydrology” of leaves: co-ordination of structure and function in temperate woody species 1 2003
... 银合欢干季的叶片膨压丧失点明显低于8个本地优势种(1/2), 这说明在该热带雨林先锋群落中银合欢的耐旱性比本地优势种的更强.研究发现, 在面临非常有限的水分供应时, 只有叶片膨压丧失点更低的植物才会拥有更强的耐旱性, 从而维持更高的气孔导度、光合速率和蒸腾速率(Sack et al., 2003; Mitchell et al., 2008; Blackman et al., 2010).这可能解释了干季时期银合欢的光合速率、气孔导度和蒸腾速率仍比8个本地优势种要高得多的原因.因此银合欢能优于本地优势种的关键可能是其在干季具有更强的耐旱性, 可以在水分供应不足时仍然保持较高的生长速率. ...
A comparison of pressure-volume curve data analysis techniques 1 1985
Subtropical dry forest regeneration in grass-invaded areas of Puerto Rico: understanding why Leucaena leucocephala dominates and native species fail 5 2012
... 银合欢(Leucaena leucocephala)具有很强的耐旱性, 即使面对干旱胁迫也能保持较高的生长速率(周利平等, 2010; Wolfe & van Bloem, 2012; Chiou et al., 2016; Barros et al., 2020).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... ).因此被广泛单一种植, 用于修复全球高度退化的热带和亚热带森林(Wolfe & van Bloem, 2012; Liu et al., 2018; Peng et al., 2019).但它目前也是全球性外来入侵植物(Richardson & Rejmánek, 2011), 威胁着全球热带和亚热带森林的生物多样性(Goel & Behl, 2002; Ishihara et al., 2018; Peng et al., 2019).在中国, 银合欢也被广泛用于造林, 修复退化的森林生态系统, 已有研究发现银合欢能成功入侵热带亚热带森林(王伟等, 2007; 彭芸, 2008; 纪中华等, 2010; 李海生等, 2015; Liu et al., 2018; Peng et al., 2019), 因此急需探究银合欢的生物防治方法.有研究表明, 想要对外来入侵植物进行生物防治, 关键是找到帮助外来入侵植物成功入侵本地生态系统的主要特征(Moran et al., 2005; Funk et al., 2008; Seastedt, 2015).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... ).因此, 迫切需要量化银合欢的关键特性(生长速率高、耐旱性强)是否真的能够帮助其成功入侵到本地生态系统中.已有研究表明, 较高的生长速率能够帮助银合欢成功入侵亚热带森林(Wolfe & van Bloem, 2012), 但是目前还不清楚高生长速率和强耐旱性是否能帮助银合欢入侵热带森林生态系统. ...
... 银合欢自身的固氮能力可以让其具有很高的生长速率进而比本地优势物种能更好地适应本地生长环境(Gibson et al., 1982; Wolfe & van Bloem, 2012).但是热带季风气候区通常具有明显的干湿季(Rowland et al., 2018), 从而会限制银合欢的生长.我们假设银合欢的高生长速率和强耐旱性不仅能比本地优势物种在干湿季都更好地适应本地环境, 而且其功能性状与本地优势物种完全不相似.为验证这一假设, 本研究以位于中国三亚抱坡岭被银合欢成功入侵的热带雨林先锋群落为研究对象, 测定了银合欢和本地优势种干季和湿季期间与生长速率(光合速率、气孔导度和蒸腾速率)和耐旱性(叶片膨压丧失点)高度相关的关键功能性状.利用这些功能性状我们可以探究: 1)银合欢的高生长速率是否比本地优势种在干季与湿季更加适应本地环境? 2)银合欢的高生长速率和强耐旱性的功能性状是否与本地优势物种完全不相似? 目前银合欢是海南危害严重的外来入侵植物之一(王伟等, 2007).如果这些功能性状确实能使银合欢优于本地优势种最终成功入侵热带雨林先锋群落, 我们也可利用这些性状和我们开发出的筛选本地物种的软件(Wang et al., 2020), 从三亚和整个海南岛的植物物种库中筛选出多种合适的本地物种对整个海南岛成功入侵热带森林的银合欢进行生物防治. ...
... 光合速率会影响植物的能量平衡, 植物的光合速率越高, 其生长速度就越快(Kirschbaum, 2011; Zhang et al., 2018b).研究发现, 无论是在湿季还是干季, 银合欢的光合速率都比其他8个本地优势种高5-10倍.而且植物光合速率较高也会导致植物拥有较高的气孔性状值(McAusland et al., 2016; Lawson & Vialet-Chabrand, 2019)和蒸腾速率(Maherali et al., 2008; dos Santos et al., 2018; Bucci et al., 2019).结果显示无论在湿季还是干季, 银合欢的气孔导度和蒸腾速率也均比其他8个本地优势种高(5-10倍).这些结果表明, 银合欢比本地优势种拥有更适应当地环境的叶片性状, 可以最大限度地提高光合速率和整体生长速度.众所周知, 水是限制植物最大光合速率的关键因素(Guan et al., 2015).由于此热带雨林先锋群落干季的降水量仅为湿季的1/10 (罗红霞等, 2018), 所以植物的光合速率会受到抑制.然而, 研究结果发现银合欢的生长速率在干季和湿季均较高, 差异不大, 而8个本地优势种的生长速率在干季则会受到抑制.因此, 研究结果进一步表明, 较高的生长速率是一个关键因素, 使银合欢优于其他8个本地优势种, 从而更好地生长在这片热带雨林先锋群落中.已有研究在亚热带的干旱森林中观察到, 快速的生长可以成功地帮助银合欢竞争过本地优势种, 从而成功入侵亚热带森林(Wolfe & van Bloem, 2012).我们的结果也表明高生长速率能帮助银合欢成功入侵热带雨林先锋群落. ...
 ,2,3,**
,2,3,**
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT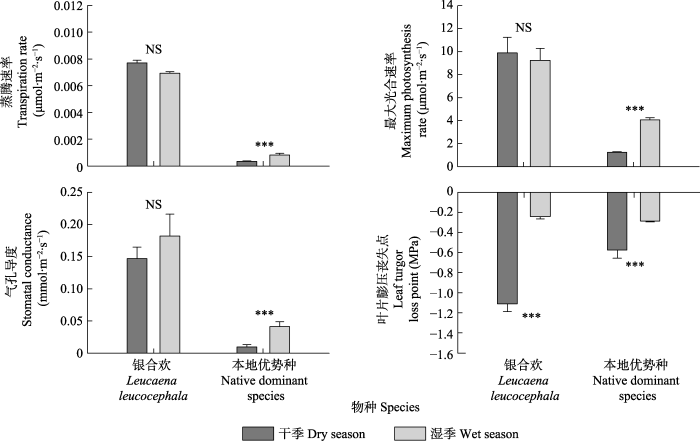 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT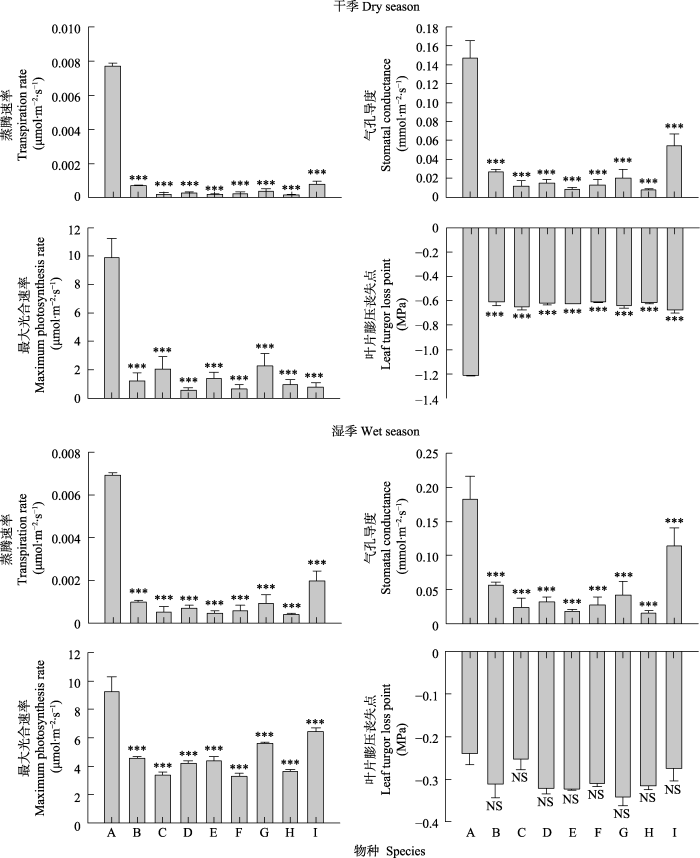 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT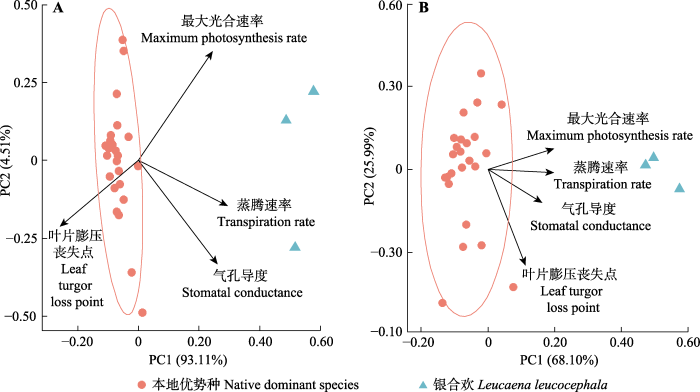 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}