王艳红
1 , 李帅锋
1 , 郎学东
1 , 黄小波
1 , 刘万德
1 , 徐崇华
2 , 苏建荣
, 1 , * 1 中国林业科学研究院资源昆虫研究所, 国家林业和草原局云南普洱森林生态系统国家定位观测研究站, 昆明 6502242 云南省太阳河自然保护区管理局, 云南普洱 665000Effects of topographic heterogeneity on species diversity in a monsoon evergreen broad- leaved forest in Pu?er, Yunnan, China Yan-Hong WANG
1 , Shuai-Feng LI
1 , Xue-Dong LANG
1 , Xiao-Bo HUANG
1 , Wan-De LIU
1 , Chong-Hua XU
2 , Jian-Rong SU
, 1 , * 1 Research Institute of Resource Insects, Chinese Academy of Forestry, Pu’er Forest Ecosystem Research Station, National Forestry and Grassland Administration of China, Kunming 6502242 Taiyanghe Provincinal Nature Reserve of Yunnan, Pu’er, Yunnan 665000, China通讯作者: *E-mail:
jianrongsu@vip.sina.com 编委: 曹敏
责任编辑: 赵航
收稿日期: 2020-05-12
接受日期: 2020-08-21
网络出版日期: 2020-10-20
基金资助: 云南省科技创新人才计划 (2018HC013 )云南省基础研究计划 (2019FB058 )云南省基础研究计划 (202001AS070005 )
Corresponding authors: *E-mail:
jianrongsu@vip.sina.com Received: 2020-05-12
Accepted: 2020-08-21
Online: 2020-10-20
Fund supported: Yunnan Science and Technology Innovation Talent Program (2018HC013 )Applied Basic Research Foundation of Yunnan Province (2019FB058 )Applied Basic Research Foundation of Yunnan Province (202001AS070005 )
摘要 地形异质性通过调控树木生长所需的养分、水分和光照等而成为亚热带森林结构与物种组成的重要驱动因子。但是, 地形异质性对季风常绿阔叶林物种多样性及其分布影响的研究还相对较少。该文基于云南普洱30 hm
2 森林动态监测样地(大样地) 750个20 m × 20 m的样方调查数据, 以海拔、坡度、凹凸度和坡向4个地形因子为变量, 采用C均值模糊聚类分析大样地的地形类型, 进而分析不同地形条件下的群落物种组成及群落物种多样性; 采用Torus转换检验法, 探讨物种与地形关联性, 为季风常绿阔叶林生物多样性保护提供科学依据。研究结果表明, 大样地可分为山脊、陡坡、缓坡、高谷和沟谷等5类地形, 地形面积分别是8.00、6.04、7.68、2.76和5.52 hm
2 。大样地中胸径(
DBH ) ≥ 1 cm的木本植物个体153β418株, 分属79科179属271种。5类地形中, 物种丰富度、不同径级的植株密度和比例明显不同, 多样性及优势物种多度分布具有较大差异。种-面积曲线表明, 同等面积条件下, 随着取样面积增加, 山脊的物种丰富度始终最小, 高谷次之, 沟谷的物种丰富度始终最大。种-个体数累积曲线表明, 随着个体数增加, 山脊物种丰富度的累积速率最小, 种丰富度增加缓慢, 高谷次之。在被检验的123个物种中, 与地形相关的物种有83个, 高达67.5%的物种与至少一类地形存在显著相关关系。山脊和缓坡中与地形具有显著负相关关系的物种数超过显著正相关的物种数; 而与陡坡、高谷和沟谷显著正相关的物种数高于显著负相关的物种数。普洱大样地地形异质性对物种多样性维持的贡献率为7.8%。
关键词: 地形异质性 ;
季风常绿阔叶林 ;
物种组成 ;
群落结构 ;
重要值 ;
物种多样性 ;
物种地形关联 Abstract Aims In subtropical forests, topographic heterogeneity drives ecosystem structure and species composition by regulating the available nutrients, water and light needed for tree growth. A gap in current research is the role of topographic heterogeneity in plant species diversity and distribution in monsoon evergreen broad-leaved forests. Our study aims to fill this gap, providing scientific grounds for the conservation of biodiversity through an exploration of floral structure and ecosystem development in monsoon evergreen broad-leaved forests.
Methods Within a 30 hm
2 dynamics plot, we established 750 20 m × 20 m sample plots, using fuzzy C-mean clustering to classify different topographic types based on elevations, convexities, slopes, and aspects of each sample plots. We compared community composition and community species diversity between different topographic types, using Torus-translation tests to examine the relationship between floral species and topography.
Important findings We identified five distinct topography types: ridge (8.00 hm
2 ), steep slope (6.04 hm
2 ), less-steep slope (7.68 hm
2 ), high plateau (2.76 hm
2 ), and valley (5.52 hm
2 ). We counted a total of 153 418 woody plant individuals with a diameter at breast height (
DBH ) ≥ 1 cm, belonging to 271 species, 179 genera and 79 families. Among the topographic types, we noted differences in species richness, plant density, proportion of different diameters, species diversity and the abundance of dominant species. When comparing species-area relationships between the five topographic types, we noticed that the ridge had the lowest number of species, while the valley had the highest. The cumulative species-individual relationships within the topographic types revealed that the ridge possesses the lowest cumulative rate. Of the 123 examined species, 67.5% were significantly related to at least one type of topography. Among species significantly related to topography, those species with significant negative correlation were more abundant in the ridge and steep slope topographic types. The opposite was true in steep slope, high plateau and valley. Based on our data, we determined that topographic heterogeneity contributed 7.8% to the maintenance of species diversity in our study area.
Keywords: topographic heterogeneity ;
monsoon broad-leaved evergreen forest ;
species composition ;
community structure ;
importance value ;
species diversity ;
topographic association of woody plants PDF (1353KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 王艳红, 李帅锋, 郎学东, 黄小波, 刘万德, 徐崇华, 苏建荣. 地形异质性对云南普洱季风常绿阔叶林物种多样性的影响. 植物生态学报, 2020, 44(10): 1015-1027. DOI:
10.17521/cjpe.2020.0148 WANG Yan-Hong, LI Shuai-Feng, LANG Xue-Dong, HUANG Xiao-Bo, LIU Wan-De, XU Chong-Hua, SU Jian-Rong.
Effects of topographic heterogeneity on species diversity in a monsoon evergreen broad- leaved forest in Pu?er, Yunnan, China .
Chinese Journal of Plant Ecology , 2020, 44(10): 1015-1027. DOI:
10.17521/cjpe.2020.0148 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 )。在众多的生境因子中, 地形越来越受到重视。在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 )。局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 )。在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 )。地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 )。海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 )。
评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 )。森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 )。因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 )。国际上关于这方面的研究主要集中在热带森林动态监测样地内。近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究。但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏。
本文基于普洱30 hm
2 森林动态监测样地(大样地)的第一次调查数据, 分析不同地形条件下的群落物种组成及群落物种多样性差异, 评估大样地中物种与地形的关联性及地形对物种共存的影响, 量化地形对群落物种组成及多样性的影响, 旨在回答: (1)地形异质性对季风常绿阔叶林木本植物群落物种组成的影响如何? (2)地形异质性对群落物种多样性的维持的贡献多大? 通过研究, 可为揭示普洱季风常绿阔叶林木本植物物种分布与共存规律积累数据资料, 为制定和优化保护措施提供科学依据。
1 材料和方法 1.1 研究区概况 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区(
图1 )。保护区地理坐标22.50o-22.63o N, 101.12o-101.25o E。地处横断山脉的无量山南延末端, 滇南热带与南亚热带的过渡位置(
李帅锋等, 2020 ), 保护对象主要为典型、完整连片的南亚热带季风常绿阔叶林和珍稀、特有动植物种类。该区气候受印度洋西南季风暖湿气流影响, 亚热带季风气候特征明显, 日温差大, 年温差小, 年平均气温高; 雨量充沛集中, 夏秋季多雨, 冬春季天气多晴朗, 全年干湿季分明。年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
苏建荣等, 2015 ;
李帅锋等, 2020 )。植被以季风常绿阔叶林为主, 人为干扰较小。森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1云南普洱30 hm2 森林动态样地地理位置示意图。 Fig. 1Location of the 30 hm2 forest dynamics plot in Pu’er, Yunnan. 1.2 森林动态监测样地的调查 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m。样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究。
1.3 地形异质性分析 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 )。其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 )。凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 )。基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型。
1.4 群落数量特征分析 统计大样地内胸径(
DBH ) ≥ 1 cm的乔木、藤本与灌木物种, 建立物种数据库, 包括科、属、种, 物种中文名与拉丁名参考《中国植物志》电子版(
http://www.iplant.cn/frps )(
中国科学院中国植物志编辑委员会, 2019 )的物种名称。分析不同地形群落数量特征、物种多样性及物种-面积及物种-个体多度曲线差异。
根据
Bellingham和Tanner (2000) 以及
苏建荣等(2015) 的划分方法, 将物种萌生类型划分为4种类型: 根萌生, 萌生位置在地下; 干基萌生, 萌生位置在地面上高度小于等于0.5 m; 干萌生, 萌生位置在地面上高度大于0.5 m, 主干受损并从残留树干上萌生; 枝萌生, 萌生位置在地面上高度大于0.5 m, 小枝受损, 从小枝与大树枝之间萌生。
通过物种重要值确定其优势物种, 重要值(
IV )的计算公式(
叶万辉等, 2008 )为:
(1) IV = (RD + RF + RP )/3 式中,
RD 为相对密度,
RF 为相对频度,
RP 为相对显著度。
RD = (某种植物的个体数/全部植物的个体数) × 100,
RF = (该种的频度/所有种的频度总和) × 100,
RP = (样方中该种胸高断面积和/样方中所有个体胸高断面积的总和) × 100。
按1-5、5-10、10-20、20-30、30-40、40-50和≥50 cm的划分标准, 将样地植株划分为7个径级。
1.5 群落物种多样性分析 Shannon-Wiener指数(
H ):
(2) $H=-\sum\limits_{i=1}^{S}{({{P}_{i}}{{\operatorname{lnP}}_{i}})}$ 式中,
S 为样地物种总数,
Pi 是样地种
i 的所有个体占总株数的比例。
参照
Hubbell和Foster (1986) 的定义, 将每hm
2 个体数不多于1株的物种定义为稀有种, 1-10株的物种定义为偶见种。
1.6 物种-面积及物种-个体多度曲线分析 从各类地形样方(20 m × 20 m)中, 依次随机抽取1、2……、
n 个样方, 直至全部抽完, 分别统计物种数量随样方数量(面积)增长的变化过程, 据此绘制物种-面积曲线; 同时统计被抽取样方中的个体数和物种数, 据此绘制物种-多度曲线。所有抽样、统计和曲线绘制均在R 3.5.1中完成。
1.7 物种与生境关联性分析 采用Torus转换检验法检验物种与生境之间的关联性, 分析生境偏好对物种多样性维持的贡献。Torus转换检验法是目前检验物种与生境关联最常用的方法之一, 能够较大程度上排除生境和物种分布的空间自相关从而使检验更加敏感(
Harms et al ., 2001 )。该方法首先计算目标种在每种生境中的实际相对多度(观测值); 其次, 让每个样方中目标种的个体数固定, 按照行或列叠加样方, 计算叠加后目标种在每种生境中的相对多度(理论值); 最后, 通过比较观测值与理论值的分位数来确定物种与生境的关联程度。如果观测值小于2.5%分位数, 则认为物种与生境负相关, 目标物种不喜好或者排斥该生境; 实际观测值大于97.5%分位数, 则认为该物种与生境正相关, 目标物种偏好这种生境; 观测值位于理论值的95%置信区间中, 则认为该物种对该生境没有什么特别排斥或者偏好。地形对物种分布与共存的贡献, 即地形异质性贡献率, 采用与地形负相关的物种平均数占被检测物种总数的比例表示(
Harms et al ., 2001 )。
2 结果和分析 2.1 地形特征 本研究中, 在海拔、坡度、坡向和凹凸度的基础上, 将大样地划分为山脊、陡坡、缓坡、高谷和沟谷5类地形(
图2 ), 地形特征见
表1 , 其中山脊面积最大, 海拔较高, 凹凸度较大, 坡向为南; 高谷面积最小, 平均海拔最高, 坡度较大, 坡向为东; 沟谷海拔最低, 凹凸度最小, 坡向为西北; 陡坡和缓坡属于高海拔向低海拔的过渡带, 坡向分别为北和南。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2云南普洱30 hm2 大样地等高线与地形类型。 Fig. 2Contour map with 5 m intervals and topography types within the 30 hm2 dynamics plot in Pu’er, Yunnan. Table 1 表1 表1 云南普洱30 hm
2 大样地5类地形特征及样地数(平均值±标准误差)
Table 1
Characteristics and numbers of plots of different topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan (mean ± SE ) 地形 海拔 坡度 凹凸度 坡向 样地数 山脊 Ridge 1 547.0 ± 0.96a 19.6 ± 0.33a 0.67 ± 0.14a 164.41 ± 1.56a 200 陡坡 Steep slope 1 525.7 ± 1.62b 19.4 ± 0.50a -0.17 ± 0.18b 324.74 ± 2.19b 151 缓坡 Less-steep slope 1 504.5 ± 1.18c 18.4 ± 0.34b -0.42 ± 0.12b 185.70 ± 1.27c 192 高谷 High plateau 1 548.7 ± 3.10a 19.5 ± 0.56a 0.41 ± 0.21a 52.53 ± 4.52d 69 沟谷 Valley 1 498.3 ± 1.89d 18.8 ± 0.52a -0.91 ± 0.20bc 294.68 ± 2.23e 138
同一列中不同小写字母表示差异显著(p < 0.05)。 Values within the same column followed by different lowercase letters are significantly different (p < 0.05). 新窗口打开 |
下载CSV 2.2 不同地形群落的数量特征 (1)多度与密度
普洱大样地中,
DBH ≥ 1 cm的木本植物个体数为153 418株(其中包括萌生株16 128株), 整体植株密度为5 114株·hm
-2 , 植株密度表现为山脊>缓坡>大样地>高谷>陡坡>沟谷, 分别为5 608、5 120、5 114、5 016、4 918和4 654株·hm
-2 (
表2 )。271个物种(乔木: 140种, 藤本: 30种, 灌木: 101种)中有148个物种植株数量不到30株·hm
-2 , 其中有40种的植株数量仅有1株, 有66个物种的植株数量在1-10株·hm
-2 之间。5类地形中缓坡物种丰富度最高, 为217; 沟谷物种丰富度为200, 位于其次; 陡坡、山脊和高谷物种丰富度分别为198、145和143。高谷每hm
2 物种数最高, 山脊最低, 沟谷>陡坡>缓坡。5类地形中, 萌生株数量差异明显, 山脊萌生株数量最多, 其次是缓坡、陡坡和沟谷, 最小为高谷(
表2 )。
Table 2 表2 表2 云南普洱30 hm
2 大样地5类地形群落的基本特征
Table 2
Basic characteristics of the communities in the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan 参数 面积2 ) 个体数 Number of individuals 植株密度 Plant density (individual·hm-2 ) 根萌生 干基萌生 干萌生 枝萌生 大样地 Big sample plot 30.00 153 418 5 114 14 691 675 721 - 山脊 Ridge 8.00 44 861 5 608 5 222 98 142 - 陡坡 Steep slope 6.04 29 703 4 918 2 687 118 113 1 缓坡 Less-steep slope 7.68 39 320 5 120 3 295 203 207 2 高谷 High plateau 2.76 13 843 5 016 1 682 62 80 - 沟谷 Valley 5.52 25 691 4 654 1 805 195 179 38
新窗口打开 |
下载CSV (2)重要值
5类地形中重要值最大的物种均为短刺锥(
Castanopsis echidnocarpa ), 且重要值要高于其他物种(
表3 ), 重要值排在其次的为泥柯(
Lithocarpus fenestratus )和西南木荷(
Schima wallichii )。这3个物种的个体数多、生存能力强, 重要值较大, 为群落的优势物种。
Table 3 表3 表3 云南普洱30 hm
2 大样地5类地形重要值前10的物种
Table 3
Top ten dominant species in the five topographic types based on importance values within the 30 hm2 dynamics plot in Pu’er, Yunnan 地形 物种 相对频度 相对多度 相对胸高断面积 重要值 山脊 短刺锥 Castanopsis echidnocarpa 3.7 28.5 34.0 22.0 泥柯 Lithocarpus fenestratus 3.7 14.5 6.5 8.2 截果柯 Lithocarpus truncatus 3.7 5.3 7.1 5.3 西南木荷 Schima wallichii 3.6 2.6 9.7 5.3 茶梨 Anneslea fragrans 3.5 4.4 5.4 4.4 隐距越桔 Vaccinium exaristatum 3.5 5.4 3.6 4.2 红梗润楠 Machilus rufipes 3.6 4.6 2.5 3.6 红叶木姜子 Litsea rubescens 3.6 4.3 2.5 3.5 红皮水锦树 Wendlandia tinctoria subsp. intermedia 3.3 3.0 2.8 3.0 深绿山龙眼 Helicia nilagirica 3.5 3.3 2.1 3.0 陡坡 短刺锥 Castanopsis echidnocarpa 2.8 16.4 22.8 14.0 泥柯 Lithocarpus fenestratus 2.6 9.8 4.2 5.5 西南木荷 Schima wallichii 2.8 3.4 9.8 5.3 红梗润楠 Machilus rufipes 2.8 5.5 4.4 4.2 枹丝锥 Castanopsis calathiformis 1.9 5.6 5.0 4.2 截果柯 Lithocarpus truncatus 2.6 4.1 5.4 4.0 红叶木姜子 Litsea rubescens 2.7 4.5 2.6 3.2 西桦 Betula alnoides 0.9 0.5 7.4 3.0 茶梨 Anneslea fragrans 2.5 3.4 2.9 2.9 隐距越桔 Vaccinium exaristatum 2.2 2.8 1.8 2.2 缓坡 短刺锥 Castanopsis echidnocarpa 2.8 21.8 24.7 16.5 西南木荷 Schima wallichii 2.8 3.2 14.3 6.8 泥柯 Lithocarpus fenestratus 2.8 10.1 6.3 6.4 枹丝锥 Castanopsis calathiformis 2.5 7.2 4.4 4.7 红梗润楠 Machilus rufipes 2.9 6.0 4.8 4.6 截果柯 Lithocarpus truncatus 2.7 4.2 6.3 4.4 茶梨 Anneslea fragrans 2.6 4.0 3.7 3.5 红叶木姜子 Litsea rubescens 2.7 3.7 3.8 3.4 隐距越桔 Vaccinium exaristatum 2.0 2.4 1.7 2.0 艾胶算盘子 Glochidion lanceolarium 2.7 2.4 0.6 1.9 高谷 短刺锥 Castanopsis echidnocarpa 3.1 16.7 24.0 14.6 泥柯 Lithocarpus fenestratus 3.0 17.2 5.7 8.6 西南木荷 Schima wallichii 3.2 3.2 13.5 6.7 红梗润楠 Machilus rufipes 3.2 5.7 5.0 4.7 截果柯 Lithocarpus truncatus 2.8 3.9 5.7 4.1 红叶木姜子 Litsea rubescens 3.1 5.4 2.9 3.8 西桦 Betula alnoides 1.4 0.8 7.7 3.3 隐距越桔 Vaccinium exaristatum 2.7 4.0 2.4 3.0 茶梨 Anneslea fragrans 3.1 2.8 2.7 2.9 枹丝锥 Castanopsis calathiformis 2.0 2.6 2.7 2.5 沟谷 短刺锥 Castanopsis echidnocarpa 2.5 12.9 15.6 10.4 西南木荷 Schima wallichii 2.4 3.4 10.9 5.6 泥柯 Lithocarpus fenestratus 2.3 9.5 4.2 5.4 红梗润楠 Machilus rufipes 2.6 5.6 6.2 4.8 截果柯 Lithocarpus truncatus 2.4 3.7 6.3 4.1 枹丝锥 Castanopsis calathiformis 1.9 5.6 3.1 3.5 云南瘿椒树 Tapiscia yunnanensis 1.8 1.4 4.7 2.6 茶梨 Anneslea fragrans 2.2 3.2 2.2 2.6 红叶木姜子 Litsea rubescens 2.2 3.2 2.2 2.5 齿叶黄杞 Engelhardtia serrata 1.7 2.4 3.2 2.4
新窗口打开 |
下载CSV 大样地整个群落的优势种相对稳定, 但同一物种在不同地形中的重要值、相对多度各不相同, 每类地形群落物种组成有一定的差异(
表3 )。5类地形中重要值最大的前10个物种有7种相同(
图3 ), 在山脊、沟谷和缓坡分别有2、2、1个物种是在其他地形中没有出现的物种, 说明5种地形群落结构存在一定差异。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3云南普洱30 hm2 大样地5类地形中优势物种重叠情况。数字代表物种数量。 Fig. 3Overlap of dominant species in five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan. Values represent number of species. (3)径级结构
表4 表明, 平均胸径为高谷>缓坡>沟谷>山脊>陡坡, 其中高谷、缓坡和沟谷的平均胸径大于大样地平均水平;
DBH < 20 cm时, 山脊1-5和5-10 cm径级的植株密度都最大, 缓坡次之, 其次是高谷、陡坡和沟谷; 总体上,
DBH < 30 cm时山脊植株密度大于沟谷;
DBH ≥ 30 cm时, 沟谷植株密度大于山脊。
Table 4 表4 表4 云南普洱30 hm
2 大样地5类地形不同径级植株密度(株?hm
-2 )及各径级植株数占总植株数比例(%)
Table 4
Plant density (individual?hm-2 ) and percentage (%) of trees with different DBH values in five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan 地形 植株密度(比例) Plant density (Percentage) 平均胸径 Average DBH (cm) 1-5 cm 5-10 cm 10-20 cm 20-30 cm 30-40 cm 40-50 cm ≥50 cm 大样地 Big sample plot 6.34 3 213 (62.83) 1 041 (20.35) 589 (11.51) 184 (3.60) 57 (1.12) 20 (0.40) 10 (0.19) 山脊 Ridge 6.33 ± 0.05a 3 376 (60.20) 1 226 (21.86) 723 (12.89) 210 (3.74) 53 (0.95) 15 (0.27) 5 (0.09) 陡坡 Steep slope 6.28 ± 0.07a 3 162 (64.31) 950 (19.32) 532 (10.83) 178 (3.61) 63 (1.27) 24 (0.48) 9 (0.18) 缓坡 Less-steep slope 6.38 ± 0.07a 3 254 (63.56) 1 012 (19.76) 584 (11.41) 180 (3.51) 54 (1.06) 22 (0.42) 14 (0.28) 高谷 High plateau 6.42 ± 0.15a 3 171 (63.22) 983 (19.61) 576 (11.49) 190 (3.78) 65 (1.30) 21 (0.41) 10 (0.19) 沟谷 Valley 6.35 ± 0.09a 2 998 (64.40) 941 (20.21) 466 (10.02) 157 (3.37) 57 (1.21) 22 (0.49) 13 (0.30)
同一列中不同小写字母表示差异显著(p < 0.05)。 Values within the same column followed by different lowercase letters are significantly different (p < 0.05). DBH, diameter at breast height. 新窗口打开 |
下载CSV 每一类地形中
DBH 为1-5 cm的个体数量最多, 该径级个体数均占各类地形中个体数的60%以上, 但在沟谷中所占比例大于其他4类地形;
DBH 为5-20 cm的个体数占总个体数的比例均表现为山脊大于其他4类地形;
DBH 为20-30、30-40、40-50和≥50 cm的个体数在各类地形中所占比例高低顺序分别为: 高谷>山脊>陡坡>缓坡>沟谷、高谷>陡坡>沟谷>缓坡>山脊、沟谷>陡坡>缓坡>高谷>山脊和沟谷>缓坡>高谷>陡坡>山脊。
2.3 不同地形群落的物种多样性 5类地形中, 科数最多的是缓坡, 其次是陡坡; 各类地形中拥有的属数为陡坡>缓坡>沟谷>山脊>高谷; 物种丰富度指数的大小顺序为缓坡>沟谷>陡坡>山脊>高谷; Shannon-Wiener多样性指数呈沟谷>陡坡>缓坡>高谷>山脊的变化趋势, 且陡坡与缓坡间并没有显著差异(
表5 )。大样地稀有种的比例为54.6%; 偶见种为24.4%, 5类地形中的稀有种比例低于大样地水平。除缓坡外, 其他地形中的偶见种数量均高于大样地水平。缓坡的稀有种和偶见种都最多, 稀有种为缓坡>陡坡>沟谷>山脊>高谷, 偶见种为缓坡>高谷>沟谷>陡坡>山脊(
表5 )。
Table 5 表5 表5 云南普洱30hm
2 大样地5类地形群落物种多样性特征
Table 5
Species diversity characteristics within the communities in the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan 参数 科数 No. 属数 丰富度 香农威纳指数 稀有种 稀有物种比例 偶见种 偶见种比例 大样地 Big sample plot 79 178 271 - 148 54.6 66 24.4 山脊 Ridge 48 104 145 2.47 ± 0.02d 63 43.4 44 30.3 陡坡 Steep slope 67 148 198 2.87 ± 0.03b 78 39.4 56 28.3 缓坡 Less-steep slope 69 147 217 2.81 ± 0.03b 101 46.5 67 30.9 高谷High plateau 51 103 143 2.69 ± 0.04c 29 20.3 65 45.5 沟谷 Valley 64 143 200 3.05 ± 0.03a 74 37.0 62 31.0
同一列中不同小写字母表示差异显著(p < 0.05)。 Values within the same column followed by different lowercase letters are significantly different (p < 0.05). 新窗口打开 |
下载CSV 2.4 物种-面积及物种-个体多度曲线 种-面积曲线(
图4 )表明, 随着取样面积增加, 同等面积条件下, 山脊的物种丰富度始终最小, 高谷次之, 沟谷的物种丰富度始终最大, 缓坡、沟谷和陡坡3类地形物种丰富度基本处于缓坡类型估计值的置信区间内; 当取样面积在0-1.48 hm
2 时, 各地形中丰富度随着面积的增加而急剧增加; 面积超过1.48 hm
2 后曲线斜率趋于稳定, 物种丰富度随取样面积增加缓慢, 当取样面积达到4.32 hm
2 时缓坡物种丰富度超过陡坡。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4云南普洱30 hm2 大样地5类地形种-面积曲线。虚线表示估计值在±1.96 × SE 的变化范围。 Fig. 4Cumulative species-area relationships of the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan. The dotted lines indicated the expected values ±1.96 × SE . 种-个体数累积曲线(
图5 )表明, 随着个体数增加, 山脊物种丰富度的累积速率最小, 种丰富度增加缓慢, 高谷次之, 沟谷最大, 当取样株数达到20 000株时, 缓坡的物种丰富度超过陡坡。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5云南普洱30 hm2 大样地5类地形种-个体数累积曲线。虚线表示估计值在±1.96 × SE 的变化范围。 Fig. 5Cumulative species-individual relationships of the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan. The dotted lines indicated the expected values ±1.96 × SE . 2.5 地形异质性与物种分布的相关性 对5类地形中出现的123个木本植物物种进行了物种与地形关联分析(
图6 )。Torus转换检验结果表明, 有83个物种具有96个显著相关关系(
p < 0.05), 其中显著正相关48个, 显著负相关48个, 换言之, 83个物种至少与一类地形存在显著相关关系(
p < 0.05)。检验结果也表明, 与5类地形显著相关的物种数明显不同, 在正相关的检验结果中, 沟谷>高谷>山脊>陡坡>缓坡; 在负相关的检验结果中, 山脊>缓坡>高谷>沟谷>陡坡。在山脊和缓坡中, 呈显著正相关关系的物种数显著低于呈显著负相关关系的物种数, 而在陡坡、高谷和沟谷中呈显著正相关关系的物种数显著高于呈显著负相关关系的物种数。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6云南普洱30 hm2 大样地与5类地形显著相关的物种数。 Fig. 6Number of species significantly correlated with each of the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan. 5类地形中重要值前10的物种共有15种, 其与地形的相关性见
表6 。茶梨(
Anneslea fragrans )、红叶木姜子(
Litsea rubescens )、截果柯(
Lithocarpus truncatus )、西桦(
Betula alnoides )、西南木荷与5类地形中任何一类均不显著相关(
p > 0.05); 艾胶算盘子(
Glochidion lanceolarium )、红梗润楠(
Machilus rufipes )、云南瘿椒树(
Tapiscia yunnanensis )与一类地形呈显著负相关关系; 短刺锥、红皮水锦树(
Wendlandia tinctoria subsp
. intermedia )、泥柯、深绿山龙眼(
Helicia nilagirica )、隐距越桔(
Vaccinium exaristatum )至少与一类地形呈显著正相关关系(
p < 0.05); 枹丝锥(
Castanopsis calathiformis )、齿叶黄杞(
Engelhardtia serrata )至少与两类地形呈显著正或负相关关系(
p < 0.05)。
Table 6 表6 表6 云南普洱30 hm
2 大样地15种物种与5类地形的相关性
Table 6
Fifteen species associated with the five topographic types within the 30 hm2 dynamics plot in Pu’er, Yunnan 物种 山脊 陡坡 缓坡 高谷 沟谷 艾胶算盘子 Glochidion lanceolarium 0 0 0 0 - 枹丝锥 Castanopsis calathiformis - 0 0 + 0 茶梨 Anneslea fragrans 0 0 0 0 0 齿叶黄杞 Engelhardtia serrata - 0 0 0 + 短刺锥 Castanopsis echidnocarpa + 0 0 0 0 红梗润楠 Machilus rufipes - 0 0 0 0 红皮水锦树 Wendlandia tinctoria subsp. intermedia + 0 0 0 0 红叶木姜子 Litsea rubescens 0 0 0 0 0 泥柯 Lithocarpus fenestratus 0 0 + 0 0 截果柯 Lithocarpus truncatus 0 0 0 0 0 深绿山龙眼 Helicia nilagirica + 0 0 0 0 西桦 Betula alnoides 0 0 0 0 0 西南木荷 Schima wallichii 0 0 0 0 0 隐距越桔 Vaccinium exaristatum + 0 0 0 0 云南瘿椒树 Tapiscia yunnanensis 0 0 - 0 0
+、-和0分别表示物种在0.05水平上与地形显著正相关、显著负相关和不显著相关。 +, -, and 0 indicates significantly positive, negative association and no significant association at level of 0.05, respectively. 新窗口打开 |
下载CSV 与某类地形正相关的物种并不完全与其他地形负相关, 大部分物种对其他地型相关性表现为中性。普洱大样地中与山脊、缓坡、陡坡、高谷和沟谷呈负相关关系的物种数分别为42、3、0、2和1。用与地形负相关的平均物种数占被检测的物种数的比例来表示地形异质性对物种多样性的贡献程度, 则大样地的地形异质性对物种多样性维持的贡献率为7.8%。
3 讨论 3.1 地形异质性对物种组成与多样性的影响 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致。本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地。
研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%)。这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 )。普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 )。山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 )。高谷物种丰富度最低, 但每hm
2 物种数最大。该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会。陡坡物种丰富度明显高于高谷和山脊。陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新。生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用。不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 )。本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响。
3.2 地形异质性对群落更新的影响 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 )。小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用。短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主。萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 )。山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多。有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多。萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 )。
沟谷植株密度最小, 但
DBH ≥ 40 cm径级的植株数占总植株数的比例在5类地形中最高。虽然沟谷中水、磷、氮等资源的含量比山脊更丰富(
Clark et al ., 1998 ), 但是, 由于沟谷地势低洼, 大量降水容易形成溪流和沟壑, 由于水分胁迫, 减少了部分不耐水湿条件植物的生长空间(
Punchi-Manage et al ., 2012 ); 土壤水分湿度过大, 透气性差, 抑制了植物生长(
郭屹立等, 2016 ); 同时, 沟谷大部分处于西北坡, 光照条件不足, 植物群落接受的光合有效辐射可能低于其他地形。由于沟谷大径级植株(
DBH ≥ 40 cm)的遮挡, 导致群落中下层光照资源更为缺乏, 阴生小径级物种,
DBH 1-5 cm的个体数比例高, 因而植株平均胸径在5类地形中偏小。
3.3 物种与生境关联 被检验的123个物种中, 与地形相关的物种有83个, 即高达67.5%的物种与至少一类地形存在显著相关关系。在5类地形中, 呈显著相关关系的物种数各不相同(
图6 ), 物种对地形生境的选择具有差异性, 说明物种经过长时间的环境适应, 群落生态位分化明显, 不同物种更多地生存于对各自有利的地形中。
Harms等(2001) 认为如果某物种与某类生境负相关, 代表该物种不能在该生境中长期生存, 正相关或中性则代表物种能在该生境中长期生存。本研究中, 陡坡、高谷和沟谷中呈显著正相关关系的物种数比呈显著负相关关系的物种数多(
图6 ), 说明植物偏好陡坡、高谷和沟谷, 尤其是沟谷, 其地形条件具有海拔低和凹凸度较小的特性, 同时具有丰富的养分和水分条件; 陡坡分布位置特殊, 为喜湿植物提供了良好的生存条件, 高谷光照充足。因此, 以上3种地形有利于物种的分布; 山脊海拔较高, 坡度大, 坡度会造成土壤中水分和矿质元素的流失, 山脊和缓坡坡向为南, 光照充足, 同时水分蒸发较快, 受此影响, 这两类生境中物种很少表现为正相关关系。此外, 地形中的海拔、坡度、坡向、凹凸度可能包含了多种环境因子的综合影响(
Chuyong et al ., 2011 ), 在地形复杂的环境中, 光照、温度、土壤养分、水分可能存在高度的异质性, 从而形成多样的小生境(
Irl et al ., 2015 ), 在生物多样性维持中起到积极作用。
4 结论 本研究基于普洱大样地野外调查数据, 分析不同地形条件下的群落物种组成及群落物种多样性, 评估物种与地形的关联性。发现不同地形条件下物种组成及多样性具有较大差异, 5类地形中, 多样性呈沟谷>陡坡>缓坡>高谷>山脊的变化趋势, 物种丰富度缓坡>沟谷>陡坡>山脊>高谷, 植株密度表现为山脊>缓坡>大样地>高谷>陡坡>沟谷; 不同地形中, 优势物种多度分布、群落物种组成及结构具有较大差异; 不同径级的植株密度和比例在不同地形中明显不同。在被检验的123个物种中, 与地形相关的物种有83个, 即高达67.5%的物种与至少一类地形存在显著相关关系。各类地形中, 呈显著相关的物种数明显不同, 在正相关的检验结果中, 沟谷>高谷>山脊>陡坡>缓坡; 在负相关的检验结果中, 山脊>缓坡>高谷>沟谷>陡坡; 在山脊和缓坡中, 呈显著正相关关系的物种数显著低于呈显著负相关关系的物种数, 而在陡坡、高谷和沟谷中呈显著正相关关系的物种数显著高于呈显著负相关关系的物种数。普洱大样地地形异质性对物种多样性维持的贡献率为7.8%, 物种对地形生境的选择具有差异性。
[1] Anderson MJ Crist TO Chase JM Vellend M Inouye BD Freestone AL Sanders NJ Cornell HV Comita LS Davies KF Harrison SP Kraft NJB Stegen JC Swenson NG 2011 ). Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologistEcology Letters, 19 -28 . [本文引用: 1] [2] Bell G Lechowicz MJ Waterway MJ 2000 ). Environmental heterogeneity and species diversity of forest sedgesJournal of Ecology, 67 -87 . [本文引用: 1] [3] Bellingham PJ Tanner EVJ 2000 ). The influence of topography on tree growth, mortality, and recruitment in a tropical montane forestBiotropica, 378 -384 . [本文引用: 1] [4] Bezdek JC 1981 ). Pattern Recognition with Fuzzy Objective Function Algorithms. [本文引用: 1] [5] Bu WS 2013 ). The Relationships Between Biodiversity and Ecosystem Functioning in Natural Tropical Forests of Hainan Island, China. PhD dissertation, Chinese Academy of Forestry, Beijing . 89 -95 . [本文引用: 1] [ 卜文圣 (2013 ). 海南岛热带天然林生物多样性与生态系统功能关系的研究博士学位论文, 中国林业科学研究院, 北京 . 89 -95 .] [本文引用: 1] [6] Chase JM 2003 ). Community assembly: When should history matter?Oecologia, 489 -498 . [本文引用: 1] [7] Chen L Mi XC Comita LS Zhang LW Ren HB Ma KP 2010 ). Community-level consequences of density dependence and habitat association in a subtropical broad-leaved forestEcology Letters, 695 -704 . [本文引用: 1] [8] Chen M Fang H Cao M 2008 ). Sprouting characteristics of sprouted woody plants in the mid-mountain humid evergreen broad-leaved forest on Ailao Mountain, Yunnan ProvinceGuihaia, 627 -632 . [本文引用: 2] [ 陈沐 , 房辉 , 曹敏 (2008 ). 云南哀牢山中山湿性常绿阔叶林树种萌生特征研究广西植物 , 28, 627 -632 .] [本文引用: 2] [9] Chuyong GB Kenfack D Harms KE Thomas DW Condit R Comita LS 2011 ). Habitat specificity and diversity of tree species in an African wet tropical forestPlant Ecology, 1363 -1374 . [本文引用: 1] [10] Clark DB Clark DA Read JM 1998 ). Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forestJournal of Ecology, 101 -112 . [本文引用: 2] [11] Condit R 1998 ). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Springer-Verlag, Berlin . [本文引用: 1] [12] Forrester D Bauhus J Connell M 2003 ). Competition in thinned Silvertop ash ( Eucalyptus sieberi L. Johnson) stands from early coppice growthForest Ecology and Management, 459 -475 . [本文引用: 1] [13] Gong GQ Chen JM Mi XC Chen SW Fang T 2007 ). Habitat associations of wood species in the Gutianshan subtropical broad-leaved evergreen forestScience of Soil and Water Conservation, 3 ), 79 -83 . [本文引用: 3] [ 宫贵权 , 程积民 , 米湘成 , 陈声文 , 方腾 (2007 ). 古田山常绿阔叶林木本植物与生境的相关性中国水土保持科学 , 5(3 ), 79 -83 .] [本文引用: 3] [14] Guo YL Wang B Xiang WS Ding T Lu SH Huang FZ Wen SJ Li DX He YL Li XK 2016 ). Responses of spatial pattern of woody plants’ basal area to topographic factors in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern ChinaBiodiversity Science, 30 -39 . [本文引用: 1] [ 郭屹立 , 王斌 , 向悟生 , 丁涛 , 陆树华 , 黄甫昭 , 文淑均 , 李冬兴 , 何运林 , 李先琨 (2016 ). 喀斯特季节性雨林木本植物胸高断面积分布格局及其对地形因子的响应生物多样性 , 24, 30 -39 .] [本文引用: 1] [15] Guo YL Wang B Xiang WS Ding T Lu SH Wen SJ Huang FZ Li DX Li XK 2015 ). Sprouting characteristics of tree species in 15 hm2 plot of northern tropical karst seasonal rain forest in Nonggang, Guangxi, southern ChinaChinese Journal of Ecology, 955 -961 . [本文引用: 1] [ 郭屹立 , 王斌 , 向悟生 , 丁涛 , 陆树华 , 文淑均 , 黄甫昭 , 李冬兴 , 李先琨 (2015 ). 弄岗北热带喀斯特季节性雨林15 hm2 样地木本植物萌生特征 生态学杂志 , 34, 955 -961 .] [本文引用: 1] [16] Hao MH Zhang ZH Zhao SS Zhao XH Beiketuerhan Y Zhang CY 2017 ). Habitat associations of tree growth in a coniferous and broad-leaved mixed forest in Jiaohe, Jilin ProvinceActa Ecologica Sinica, 3437 -3444 . [本文引用: 1] [ 郝珉辉 , 张忠辉 , 赵珊珊 , 赵秀海 , 叶尔江·拜克吐尔汉 , 张春雨 (2017 ). 吉林蛟河针阔混交林树木生长与生境的关联性生态学报 , 37, 3437 -3444 .] [本文引用: 1] [17] Harms KE Condit R Hubbell SP Foster RB 2001 ). Habitat associations of trees and shrubs in a 50 hm2 neotropical forest plot Journal of Ecology, 947 -959 . [本文引用: 5] [18] Hennenberg KJ Bruelheide H 2003 ). Ecological investigations on the northern distribution range of Hippocrepis comosa L. in GermanyPlant Ecology, 167 -188 . [本文引用: 1] [19] Hofer G Wagner HH Herzog F Edwards PJ 2008 ). Effects of topographic variability on the scaling of plant species richness in gradient dominated landscapesEcography, 131 -139 . [本文引用: 1] [20] Hubbell S Foster RB 1986 ). Commonness and rarity in a neotropical forest: implications for tropical tree conservation //Soule ME. Conservation Biology: Science of Scarcity and Diversity. Sinauer Press, Sunderland, UK. [本文引用: 1] [21] Irl SDH Harter DEV Steinbauer MJ Puyol GD Fernández- Palacios JM Jentsch A Beierkuhnlein C 2015 ). Climate vs. topography-spatial patterns of plant species diversity and endemism on a high-elevation islandJournal of Ecology, 1621 -1633 . [本文引用: 1] [22] Jiang P Ye J Wu G 2005 ). Woody species composition and biomass of main tree species in a 25 hm2 plot of broad-leaved and Korean pine mixed forests of Changbai Mountain, northeast China Journal of Beijing Forestry University, Supp.2 ), 112 -115 . [本文引用: 1] [ 姜萍 , 叶吉 , 吴钢 (2005 ). 长白山阔叶红松林大样地木本植物组成及主要树种的生物量北京林业大学学报 , 27(增刊2 ), 112 -115 .] [本文引用: 1] [23] Lai JS Mi XC Ren HB Ma KP 2010 ). Numerical classification of associations in subtropical evergreen broad- leaved forest based on multivariate regression trees—A case study 24 hm2 Gutianshan forest plot in China Chinese Journal of Plant Ecology, 761 -769 . [本文引用: 1] [ 赖江山 , 米湘成 , 任海保 , 马克平 (2010 ). 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例植物生态学报 , 34, 761 -769 .] [本文引用: 1] [24] Lan GY Hu YH Cao M Zhu H 2011 ). Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in ChinaForest Ecology and Management, 1507 -1513 . [本文引用: 2] [25] Lan GY Hu YH Cao M Zhu H Wang H Zhou SS Deng XB Cui JY Huang JG Liu LY Xu HL Song JP He YC 2008 ). Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patternsJournal of Plant Ecology (Chinese Version )287 -298 . [本文引用: 1] [ 兰国玉 , 胡跃华 , 曹敏 , 朱华 , 王洪 , 周仕顺 , 邓晓保 , 崔景云 , 黄建国 , 刘林云 , 许海龙 , 宋军平 , 何有才 (2008 ). 西双版纳热带森林动态监测样地: 树种组成与空间分布格局植物生态学报 , 32, 287 -298 .] [本文引用: 1] [26] Li SF Lang XD Huang XB Wang YH Liu WD Xu CH Su JR 2020 ). Association classification of a 30 hm2 dynamics plot in the monsoon broad-leaved evergreen forest in Pu’er, Yunnan Province, ChinaChinese Journal of Plant Ecology, 248 -259 . [本文引用: 3] [ 李帅锋 , 郎学东 , 黄小波 , 王艳红 , 刘万德 , 徐崇华 , 苏建荣 (2020 ). 云南普洱30公顷季风常绿阔叶林动态监测样地群丛数量分类植物生态学报 , 44, 248 -259 .] [本文引用: 3] [27] Li SF Liu WD Su JR Zhang ZJ Liu QY 2012 ). Woody seedling regeneration in secondary succession of monsoon broad-leaved evergreen forest in Pu’er, Yunnan, southwest ChinaActa Ecologia Sinica, 5653 -5662 . [本文引用: 1] [ 李帅锋 , 刘万德 , 苏建荣 , 张志钧 , 刘庆云 (2012 ). 普洱季风常绿阔叶林次生演替中木本植物幼苗更新特征生态学报 , 32, 5653 -5662 .] [本文引用: 1] [28] Li X Nie Y Song X Zhang R Wang G 2011 ). Patterns of species diversity and functional diversity along the south to north-facing slope gradient in a sub-alpine meadowCommunity Ecology, 179 -187 . [本文引用: 1] [29] Liu JJ Tan YH Slik JWF 2014 ). Topography related habitat associations of tree species traits, composition and diversity in a Chinese tropical forestForest Ecology and Management, 75 -81 . [本文引用: 1] [30] Liu WP Cao HL Liu W Lian JY Wu LF 2011 ). Study on diversity of monsoon evergreen broad leaved forest in diferent kinds of habitat in DinghushanJournal of Anhui Agricultural Science, 16159 -16163 . [本文引用: 2] [ 刘文平 , 曹洪麟 , 刘卫 , 练琚愉 , 吴林芳 (2011 ). 鼎湖山季风常绿阔叶林不同生境物种多样性研究安徽农业科学 , 39, 16159 -16163 .] [本文引用: 2] [31] Lundholm JT 2009 ). Plant species diversity and environmental heterogeneity: spatial scale and competing hypothesesJournal of Vegetation Science, 377 -391 . [本文引用: 1] [32] Ma KP 2017 ). Forest dynamics plot is a crosscutting research platform for biodiversity scienceBiodiversity Science, 227 -228 . [本文引用: 1] [ 马克平 (2017 ). 森林动态大样地是生物多样性科学综合研究平台生物多样性 , 25, 227 -228 .] [本文引用: 1] [33] Pigott CD 1975 ). Experimental studies on the influence of climate on the geographical distribution of plantsWeather, 82 -90 . [本文引用: 1] [34] Punchi-Manage R Getzin S Wiegand T Kanagaraj R Savitri GCV Nimal GIAU Wiegand K Huth A 2012 ). Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forestJournal of Ecology, 149 -160 . [本文引用: 1] [35] Ru HL Zhang HD Jiao F Xue CY Guo ML 2015 ). Impact of micro-landform on grassland plant community structure and function in the hilly Loess Plateau region, ChinaChinese Journal of Applied Ecology, 25 -32 . [本文引用: 1] [ 汝海丽 , 张海东 , 焦峰 , 薛超玉 , 郭美丽 (2015 ). 黄土丘陵区微地形对草地植物群落结构组成和功能特征的影响应用生态学报 , 27, 25 -32 .] [本文引用: 1] [36] Sharma CM Suyal S Gairola S Ghildiyal SK 2009 ). Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal HimalayaJournal of American Science, 119 -128 . [本文引用: 1] [37] Shen GC Yu MJ Hu XS Mi XC Ren HB Sun IF Ma KP 2009 ). Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneityEcology, 3033 -3041 . [本文引用: 1] [38] Shen ZH Zhang XS Jin YX 2000 ). Spatial pattern analysis and topographical interpretation of species diversity in the forests of Dalaoling in the region of the Three GorgesActa Botanica Sinica, 620 -627 . [本文引用: 1] [ 沈泽昊 , 张新时 , 金义兴 (2000 ). 三峡大老岭森林物种多样性的空间格局分析及其地形解释植物学报 , 42, 620 -627 .] [本文引用: 1] [39] Song YC Yan ER Song K 2015 ). Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, ChinaBiodiversity Science, 139 -148 . [本文引用: 1] [ 宋永昌 , 阎恩荣 , 宋坤 (2015 ). 中国常绿阔叶林8大动态监测样地植被的综合比较生物多样性 , 23, 139 -148 .] [本文引用: 1] [40] Su JR Liu WD Li SF Lang XD 2015 ). Restoration Ecology of Monsoon Broad-leaved Evergreen Forest in West China. 3 -4 . [本文引用: 2] [ 苏建荣 , 刘万德 , 李帅锋 , 郎学东 (2015 ). 西部季风常绿阔叶林恢复生态学 . 科学出版社, 北京. 3 -4 .] [本文引用: 2] [41] Su JR Liu WD Zhang ZJ Li SF 2012 ). Sprouting characteristic in restoration ecosystems of monsoon evergreen broadleaved forest in south-central of Yunnan ProvinceActa Ecologica Sinica, 805 -814 . [本文引用: 1] [ 苏建荣 , 刘万德 , 张志钧 , 李帅锋 (2012 ). 云南中南部季风常绿阔叶林恢复生态系统萌生特征生态学报 , 32, 805 -814 .] [本文引用: 1] [42] Su SJ 2015 ). Spatial Heterogeneity and Vegetation Maintaining Mechanism of Pinus taiwanensis Forest in Daiyun Mountain High Altitude Area PhD dissertation, Fujian Agriculture and Forestry University, Fuzhou , 49 -65 . [本文引用: 1] [ 苏松锦 (2015 ). 戴云山高海拔黄山松林的空间异质性与植被维持机制博士学位论文, 福建农林大学, 福州 . 49 -65 .] [本文引用: 1] [43] Sun XW Yang QS Liu HM Wang XH 2018 ). Classification of plant associations based on a 20 hm2 dynamics plot of evergreen broad-leaved forest in Mt. Tiantong, Zhejiang, ChinaChinese Journal of Plant Ecology, 550 -561 . [本文引用: 1] [ 孙小伟 , 杨庆松 , 刘何铭 , 王希华 (2018 ). 基于浙江天童20 hm2 常绿阔叶林动态监测样地的群丛划分 植物生态学报 , 42, 550 -561 .] [本文引用: 1] [44] The Editorial Committee of Flora Reipublicae Popularis Sinicae, Chinese Academy of Sciences (2019 ). Flora Reipublicae Popularis Sinicae URL [本文引用: 1] [ 中国科学院中国植物志编辑委员会 (2019 ). 中国植物志 . http://www.iplant.cn/frps.] URL [本文引用: 1] [45] Tilman D Pacala S 1993 ). The Maintenance of Species Richness in Plant Communities [本文引用: 1] [46] Wang JM Xu H Li YD Lin MX Zhou Z Luo TS Chen DX 2018 ). Effects of topographic heterogeneity on community structure and diversity of woody plants in Jianfengling tropical montane rainforestSciental Silvae Sinicae, 1 ), 1 -11 . [本文引用: 2] [ 王家鸣 , 许涵 , 李意德 , 林明献 , 周璋 , 骆土寿 , 陈德祥 (2018 ). 地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响林业科学 , 54(1 ), 1 -11 .] [本文引用: 2] [47] Wang W Luo ZR Zhou RF Xu DM Ai JG Ding BY 2011 ). Habitat associations of woody plant species in Baishanzu subtropical broad-leaved evergreen forestBiodiversity Science, 134 -142 . [本文引用: 2] [ 王伟 , 骆争荣 , 周荣飞 , 许大明 , 哀建国 , 丁炳扬 (2011 ). 百山祖常绿阔叶林木本植物的生境相关性分析生物多样性 , 19, 134 -142 .] [本文引用: 2] [48] Wen HD Lin LX Yang J Hu YH Cao M Liu YH Lu ZY Xie YN 2018 ). Species composition and community structure of a 20 hm2 plot of mid-mountain moist evergreen broad-leaved forest on the Mts. Ailaoshan, Yunnan Province, China Chinese Journal of Plant Ecology, 419 -429 . [本文引用: 4] [ 温韩东 , 林露湘 , 杨洁 , 胡跃华 , 曹敏 , 刘玉洪 , 鲁志云 , 谢有能 (2018 ). 云南哀牢山中山湿性常绿阔叶林 20 hm2 动态样地的物种组成与群落结构 植物生态学报 , 42, 419 -429 .] [本文引用: 4] [49] Yang QS Ma ZP Xie YB Zhang ZG Wang ZH Liu HM Li P Zhang N Wang DL Yang HB Fang XF Yan ER Wang XH 2011 ). Community structure and species composition of an evergreen broadleaved forest in Tiantong’s 20 hm2 dynamic plot, Zhejiang Province, eastern China Biodiversity Science, 215 -223 . [本文引用: 2] [ 杨庆松 , 马遵平 , 谢玉彬 , 张志国 , 王樟华 , 刘何铭 , 李萍 , 张娜 , 王达力 , 杨海波 , 方晓峰 , 阎恩荣 , 王希华 (2011 ). 浙江天童20 hm2 常绿阔叶林动态监测样地的群落特征 生物多样性 , 19, 215 -223 .] [本文引用: 2] [50] Yang YC Da LJ You WH 2005 ). Vegetation structure in relation to micro-landform in Tiantong National Forest Park, Zhejiang, ChinaActa Ecologica Sinica, 38 -48 . [本文引用: 1] [ 杨永川 , 达良俊 , 由文辉 (2005 ). 浙江天童国家森林公园微地形与植被结构的关系生态学报 , 25, 38 -48 .] [本文引用: 1] [51] Ye WH Cao HL Huang ZL Lian JY Wang ZG Li L Wei SG Wang ZM 2008 ). Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, ChinaJournal of Plant Ecology (Chinese Version )274 -286 . [本文引用: 4] [ 叶万辉 , 曹洪麟 , 黄忠良 , 练琚愉 , 王志高 , 李林 , 魏识广 , 王章明 (2008 ). 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究植物生态学报 , 32, 274 -286 .] [本文引用: 4] [52] Yuan TX Zhang HP Ou ZY Tan YB 2014 ). Effects of topography on the diversity and distribution pattern of ground plants in karst montane forests in southwest Guangxi, ChinaChinese Journal of Applied Ecology, 2803 -2810 . [本文引用: 1] [ 袁铁象 , 张合平 , 欧芷阳 , 谭一波 (2014 ). 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响应用生态学报 , 25, 2803 -2810 .] [本文引用: 1] [53] Zhang YD Liu SR Zhao CM 2005 ). Spatial pattern of sub- alpine forest restoration in west SichuanChinese Journal of Applied Ecology, 1706 -1710 . [本文引用: 1] [ 张远东 , 刘世荣 , 赵常明 (2005 ). 川西亚高山森林恢复的空间格局分析应用生态学报 , 16, 1706 -1710 .] [本文引用: 1] [54] Zhu H 2019 ). Floristic divergence of the evergreen broad-leaved forests in Yunnan, southwestern ChinaPhytotaxa, 1 -20 . [本文引用: 1] [55] Zhu Y Zhao GF Zhang LW Shen GC Mi XC Ren HB Yu MJ Chen JH Chen SW Fang T Ma KP 2008 ). Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, east ChinaJournal of Plant Ecology (Chinese Version )262 -273 . [本文引用: 1] [ 祝燕 , 赵谷风 , 张俪文 , 沈国春 , 米湘成 , 任海保 , 于明坚 , 陈建华 , 陈声文 , 方腾 , 马克平 (2008 ). 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构植物生态学报 , 32, 262 -273 .] [本文引用: 1] Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist
1
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
Environmental heterogeneity and species diversity of forest sedges
1
2000
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
The influence of topography on tree growth, mortality, and recruitment in a tropical montane forest
1
2000
... 根据
Bellingham和Tanner (2000) 以及
苏建荣等(2015) 的划分方法, 将物种萌生类型划分为4种类型: 根萌生, 萌生位置在地下; 干基萌生, 萌生位置在地面上高度小于等于0.5 m; 干萌生, 萌生位置在地面上高度大于0.5 m, 主干受损并从残留树干上萌生; 枝萌生, 萌生位置在地面上高度大于0.5 m, 小枝受损, 从小枝与大树枝之间萌生. ...
1
1981
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
海南岛热带天然林生物多样性与生态系统功能关系的研究
1
2013
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
海南岛热带天然林生物多样性与生态系统功能关系的研究
1
2013
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
Community assembly: When should history matter?
1
2003
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Community-level consequences of density dependence and habitat association in a subtropical broad-leaved forest
1
2010
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
云南哀牢山中山湿性常绿阔叶林树种萌生特征研究
2
2008
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
... ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
云南哀牢山中山湿性常绿阔叶林树种萌生特征研究
2
2008
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
... ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
Habitat specificity and diversity of tree species in an African wet tropical forest
1
2011
... 被检验的123个物种中, 与地形相关的物种有83个, 即高达67.5%的物种与至少一类地形存在显著相关关系.在5类地形中, 呈显著相关关系的物种数各不相同(
图6 ), 物种对地形生境的选择具有差异性, 说明物种经过长时间的环境适应, 群落生态位分化明显, 不同物种更多地生存于对各自有利的地形中.
Harms等(2001) 认为如果某物种与某类生境负相关, 代表该物种不能在该生境中长期生存, 正相关或中性则代表物种能在该生境中长期生存.本研究中, 陡坡、高谷和沟谷中呈显著正相关关系的物种数比呈显著负相关关系的物种数多(
图6 ), 说明植物偏好陡坡、高谷和沟谷, 尤其是沟谷, 其地形条件具有海拔低和凹凸度较小的特性, 同时具有丰富的养分和水分条件; 陡坡分布位置特殊, 为喜湿植物提供了良好的生存条件, 高谷光照充足.因此, 以上3种地形有利于物种的分布; 山脊海拔较高, 坡度大, 坡度会造成土壤中水分和矿质元素的流失, 山脊和缓坡坡向为南, 光照充足, 同时水分蒸发较快, 受此影响, 这两类生境中物种很少表现为正相关关系.此外, 地形中的海拔、坡度、坡向、凹凸度可能包含了多种环境因子的综合影响(
Chuyong et al ., 2011 ), 在地形复杂的环境中, 光照、温度、土壤养分、水分可能存在高度的异质性, 从而形成多样的小生境(
Irl et al ., 2015 ), 在生物多样性维持中起到积极作用. ...
Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest
2
1998
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
... 沟谷植株密度最小, 但
DBH ≥ 40 cm径级的植株数占总植株数的比例在5类地形中最高.虽然沟谷中水、磷、氮等资源的含量比山脊更丰富(
Clark et al ., 1998 ), 但是, 由于沟谷地势低洼, 大量降水容易形成溪流和沟壑, 由于水分胁迫, 减少了部分不耐水湿条件植物的生长空间(
Punchi-Manage et al ., 2012 ); 土壤水分湿度过大, 透气性差, 抑制了植物生长(
郭屹立等, 2016 ); 同时, 沟谷大部分处于西北坡, 光照条件不足, 植物群落接受的光合有效辐射可能低于其他地形.由于沟谷大径级植株(
DBH ≥ 40 cm)的遮挡, 导致群落中下层光照资源更为缺乏, 阴生小径级物种,
DBH 1-5 cm的个体数比例高, 因而植株平均胸径在5类地形中偏小. ...
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. 1
1998
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
Competition in thinned Silvertop ash (
Eucalyptus sieberi L. Johnson) stands from early coppice growth
1
2003
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
古田山常绿阔叶林木本植物与生境的相关性
3
2007
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
古田山常绿阔叶林木本植物与生境的相关性
3
2007
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
喀斯特季节性雨林木本植物胸高断面积分布格局及其对地形因子的响应
1
2016
... 沟谷植株密度最小, 但
DBH ≥ 40 cm径级的植株数占总植株数的比例在5类地形中最高.虽然沟谷中水、磷、氮等资源的含量比山脊更丰富(
Clark et al ., 1998 ), 但是, 由于沟谷地势低洼, 大量降水容易形成溪流和沟壑, 由于水分胁迫, 减少了部分不耐水湿条件植物的生长空间(
Punchi-Manage et al ., 2012 ); 土壤水分湿度过大, 透气性差, 抑制了植物生长(
郭屹立等, 2016 ); 同时, 沟谷大部分处于西北坡, 光照条件不足, 植物群落接受的光合有效辐射可能低于其他地形.由于沟谷大径级植株(
DBH ≥ 40 cm)的遮挡, 导致群落中下层光照资源更为缺乏, 阴生小径级物种,
DBH 1-5 cm的个体数比例高, 因而植株平均胸径在5类地形中偏小. ...
喀斯特季节性雨林木本植物胸高断面积分布格局及其对地形因子的响应
1
2016
... 沟谷植株密度最小, 但
DBH ≥ 40 cm径级的植株数占总植株数的比例在5类地形中最高.虽然沟谷中水、磷、氮等资源的含量比山脊更丰富(
Clark et al ., 1998 ), 但是, 由于沟谷地势低洼, 大量降水容易形成溪流和沟壑, 由于水分胁迫, 减少了部分不耐水湿条件植物的生长空间(
Punchi-Manage et al ., 2012 ); 土壤水分湿度过大, 透气性差, 抑制了植物生长(
郭屹立等, 2016 ); 同时, 沟谷大部分处于西北坡, 光照条件不足, 植物群落接受的光合有效辐射可能低于其他地形.由于沟谷大径级植株(
DBH ≥ 40 cm)的遮挡, 导致群落中下层光照资源更为缺乏, 阴生小径级物种,
DBH 1-5 cm的个体数比例高, 因而植株平均胸径在5类地形中偏小. ...
弄岗北热带喀斯特季节性雨林15 hm
2 样地木本植物萌生特征
1
2015
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
弄岗北热带喀斯特季节性雨林15 hm
2 样地木本植物萌生特征
1
2015
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
吉林蛟河针阔混交林树木生长与生境的关联性
1
2017
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
吉林蛟河针阔混交林树木生长与生境的关联性
1
2017
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
Habitat associations of trees and shrubs in a 50 hm
2 neotropical forest plot
5
2001
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
... ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
... 采用Torus转换检验法检验物种与生境之间的关联性, 分析生境偏好对物种多样性维持的贡献.Torus转换检验法是目前检验物种与生境关联最常用的方法之一, 能够较大程度上排除生境和物种分布的空间自相关从而使检验更加敏感(
Harms et al ., 2001 ).该方法首先计算目标种在每种生境中的实际相对多度(观测值); 其次, 让每个样方中目标种的个体数固定, 按照行或列叠加样方, 计算叠加后目标种在每种生境中的相对多度(理论值); 最后, 通过比较观测值与理论值的分位数来确定物种与生境的关联程度.如果观测值小于2.5%分位数, 则认为物种与生境负相关, 目标物种不喜好或者排斥该生境; 实际观测值大于97.5%分位数, 则认为该物种与生境正相关, 目标物种偏好这种生境; 观测值位于理论值的95%置信区间中, 则认为该物种对该生境没有什么特别排斥或者偏好.地形对物种分布与共存的贡献, 即地形异质性贡献率, 采用与地形负相关的物种平均数占被检测物种总数的比例表示(
Harms et al ., 2001 ). ...
... ).该方法首先计算目标种在每种生境中的实际相对多度(观测值); 其次, 让每个样方中目标种的个体数固定, 按照行或列叠加样方, 计算叠加后目标种在每种生境中的相对多度(理论值); 最后, 通过比较观测值与理论值的分位数来确定物种与生境的关联程度.如果观测值小于2.5%分位数, 则认为物种与生境负相关, 目标物种不喜好或者排斥该生境; 实际观测值大于97.5%分位数, 则认为该物种与生境正相关, 目标物种偏好这种生境; 观测值位于理论值的95%置信区间中, 则认为该物种对该生境没有什么特别排斥或者偏好.地形对物种分布与共存的贡献, 即地形异质性贡献率, 采用与地形负相关的物种平均数占被检测物种总数的比例表示(
Harms et al ., 2001 ). ...
... 被检验的123个物种中, 与地形相关的物种有83个, 即高达67.5%的物种与至少一类地形存在显著相关关系.在5类地形中, 呈显著相关关系的物种数各不相同(
图6 ), 物种对地形生境的选择具有差异性, 说明物种经过长时间的环境适应, 群落生态位分化明显, 不同物种更多地生存于对各自有利的地形中.
Harms等(2001) 认为如果某物种与某类生境负相关, 代表该物种不能在该生境中长期生存, 正相关或中性则代表物种能在该生境中长期生存.本研究中, 陡坡、高谷和沟谷中呈显著正相关关系的物种数比呈显著负相关关系的物种数多(
图6 ), 说明植物偏好陡坡、高谷和沟谷, 尤其是沟谷, 其地形条件具有海拔低和凹凸度较小的特性, 同时具有丰富的养分和水分条件; 陡坡分布位置特殊, 为喜湿植物提供了良好的生存条件, 高谷光照充足.因此, 以上3种地形有利于物种的分布; 山脊海拔较高, 坡度大, 坡度会造成土壤中水分和矿质元素的流失, 山脊和缓坡坡向为南, 光照充足, 同时水分蒸发较快, 受此影响, 这两类生境中物种很少表现为正相关关系.此外, 地形中的海拔、坡度、坡向、凹凸度可能包含了多种环境因子的综合影响(
Chuyong et al ., 2011 ), 在地形复杂的环境中, 光照、温度、土壤养分、水分可能存在高度的异质性, 从而形成多样的小生境(
Irl et al ., 2015 ), 在生物多样性维持中起到积极作用. ...
Ecological investigations on the northern distribution range of
Hippocrepis comosa L. in Germany
1
2003
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Effects of topographic variability on the scaling of plant species richness in gradient dominated landscapes
1
2008
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
1
1986
... 参照
Hubbell和Foster (1986) 的定义, 将每hm
2 个体数不多于1株的物种定义为稀有种, 1-10株的物种定义为偶见种. ...
Climate vs. topography-spatial patterns of plant species diversity and endemism on a high-elevation island
1
2015
... 被检验的123个物种中, 与地形相关的物种有83个, 即高达67.5%的物种与至少一类地形存在显著相关关系.在5类地形中, 呈显著相关关系的物种数各不相同(
图6 ), 物种对地形生境的选择具有差异性, 说明物种经过长时间的环境适应, 群落生态位分化明显, 不同物种更多地生存于对各自有利的地形中.
Harms等(2001) 认为如果某物种与某类生境负相关, 代表该物种不能在该生境中长期生存, 正相关或中性则代表物种能在该生境中长期生存.本研究中, 陡坡、高谷和沟谷中呈显著正相关关系的物种数比呈显著负相关关系的物种数多(
图6 ), 说明植物偏好陡坡、高谷和沟谷, 尤其是沟谷, 其地形条件具有海拔低和凹凸度较小的特性, 同时具有丰富的养分和水分条件; 陡坡分布位置特殊, 为喜湿植物提供了良好的生存条件, 高谷光照充足.因此, 以上3种地形有利于物种的分布; 山脊海拔较高, 坡度大, 坡度会造成土壤中水分和矿质元素的流失, 山脊和缓坡坡向为南, 光照充足, 同时水分蒸发较快, 受此影响, 这两类生境中物种很少表现为正相关关系.此外, 地形中的海拔、坡度、坡向、凹凸度可能包含了多种环境因子的综合影响(
Chuyong et al ., 2011 ), 在地形复杂的环境中, 光照、温度、土壤养分、水分可能存在高度的异质性, 从而形成多样的小生境(
Irl et al ., 2015 ), 在生物多样性维持中起到积极作用. ...
长白山阔叶红松林大样地木本植物组成及主要树种的生物量
1
2005
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
长白山阔叶红松林大样地木本植物组成及主要树种的生物量
1
2005
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China
2
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 地形异质性通过海拔、坡度、凹凸度和坡向4个地形因子进行度量(
郝珉辉等, 2017 ).其中, 海拔为每个20 m × 20 m样方4个角的海拔平均值; 坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的平均值(
Harms et al ., 2001 ).凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值, 处于大样地边缘的样地凹凸度为样地中心的海拔减去4个顶点海拔的平均值(
Lan et al ., 2011 ), 凹凸度为正值则说明目标样方比周围样方高; 为负值则说明目标样方比周围样方低; 为零则说明目标样方与周围样方高度相等(
Harms et al ., 2001 ).基于普洱森林动态监测样地750个样方的平均海拔、坡度、坡向和凹凸度进行C均值模糊聚类分析(
Bezdek, 1981 ;
宫贵权等, 2007 ), 划分大样地不同的地形类型. ...
西双版纳热带森林动态监测样地: 树种组成与空间分布格局
1
2008
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
西双版纳热带森林动态监测样地: 树种组成与空间分布格局
1
2008
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
云南普洱30公顷季风常绿阔叶林动态监测样地群丛数量分类
3
2020
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区(
图1 ).保护区地理坐标22.50o-22.63o N, 101.12o-101.25o E.地处横断山脉的无量山南延末端, 滇南热带与南亚热带的过渡位置(
李帅锋等, 2020 ), 保护对象主要为典型、完整连片的南亚热带季风常绿阔叶林和珍稀、特有动植物种类.该区气候受印度洋西南季风暖湿气流影响, 亚热带季风气候特征明显, 日温差大, 年温差小, 年平均气温高; 雨量充沛集中, 夏秋季多雨, 冬春季天气多晴朗, 全年干湿季分明.年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
苏建荣等, 2015 ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
云南普洱30公顷季风常绿阔叶林动态监测样地群丛数量分类
3
2020
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区(
图1 ).保护区地理坐标22.50o-22.63o N, 101.12o-101.25o E.地处横断山脉的无量山南延末端, 滇南热带与南亚热带的过渡位置(
李帅锋等, 2020 ), 保护对象主要为典型、完整连片的南亚热带季风常绿阔叶林和珍稀、特有动植物种类.该区气候受印度洋西南季风暖湿气流影响, 亚热带季风气候特征明显, 日温差大, 年温差小, 年平均气温高; 雨量充沛集中, 夏秋季多雨, 冬春季天气多晴朗, 全年干湿季分明.年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
苏建荣等, 2015 ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
普洱季风常绿阔叶林次生演替中木本植物幼苗更新特征
1
2012
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
普洱季风常绿阔叶林次生演替中木本植物幼苗更新特征
1
2012
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
Patterns of species diversity and functional diversity along the south to north-facing slope gradient in a sub-alpine meadow
1
2011
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
Topography related habitat associations of tree species traits, composition and diversity in a Chinese tropical forest
1
2014
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
鼎湖山季风常绿阔叶林不同生境物种多样性研究
2
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
鼎湖山季风常绿阔叶林不同生境物种多样性研究
2
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
Plant species diversity and environmental heterogeneity: spatial scale and competing hypotheses
1
2009
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
森林动态大样地是生物多样性科学综合研究平台
1
2017
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
森林动态大样地是生物多样性科学综合研究平台
1
2017
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
Experimental studies on the influence of climate on the geographical distribution of plants
1
1975
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forest
1
2012
... 沟谷植株密度最小, 但
DBH ≥ 40 cm径级的植株数占总植株数的比例在5类地形中最高.虽然沟谷中水、磷、氮等资源的含量比山脊更丰富(
Clark et al ., 1998 ), 但是, 由于沟谷地势低洼, 大量降水容易形成溪流和沟壑, 由于水分胁迫, 减少了部分不耐水湿条件植物的生长空间(
Punchi-Manage et al ., 2012 ); 土壤水分湿度过大, 透气性差, 抑制了植物生长(
郭屹立等, 2016 ); 同时, 沟谷大部分处于西北坡, 光照条件不足, 植物群落接受的光合有效辐射可能低于其他地形.由于沟谷大径级植株(
DBH ≥ 40 cm)的遮挡, 导致群落中下层光照资源更为缺乏, 阴生小径级物种,
DBH 1-5 cm的个体数比例高, 因而植株平均胸径在5类地形中偏小. ...
黄土丘陵区微地形对草地植物群落结构组成和功能特征的影响
1
2015
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
黄土丘陵区微地形对草地植物群落结构组成和功能特征的影响
1
2015
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal Himalaya
1
2009
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity
1
2009
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
三峡大老岭森林物种多样性的空间格局分析及其地形解释
1
2000
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
三峡大老岭森林物种多样性的空间格局分析及其地形解释
1
2000
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
中国常绿阔叶林8大动态监测样地植被的综合比较
1
2015
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
中国常绿阔叶林8大动态监测样地植被的综合比较
1
2015
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
2
2015
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区(
图1 ).保护区地理坐标22.50o-22.63o N, 101.12o-101.25o E.地处横断山脉的无量山南延末端, 滇南热带与南亚热带的过渡位置(
李帅锋等, 2020 ), 保护对象主要为典型、完整连片的南亚热带季风常绿阔叶林和珍稀、特有动植物种类.该区气候受印度洋西南季风暖湿气流影响, 亚热带季风气候特征明显, 日温差大, 年温差小, 年平均气温高; 雨量充沛集中, 夏秋季多雨, 冬春季天气多晴朗, 全年干湿季分明.年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
苏建荣等, 2015 ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... 根据
Bellingham和Tanner (2000) 以及
苏建荣等(2015) 的划分方法, 将物种萌生类型划分为4种类型: 根萌生, 萌生位置在地下; 干基萌生, 萌生位置在地面上高度小于等于0.5 m; 干萌生, 萌生位置在地面上高度大于0.5 m, 主干受损并从残留树干上萌生; 枝萌生, 萌生位置在地面上高度大于0.5 m, 小枝受损, 从小枝与大树枝之间萌生. ...
2
2015
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区(
图1 ).保护区地理坐标22.50o-22.63o N, 101.12o-101.25o E.地处横断山脉的无量山南延末端, 滇南热带与南亚热带的过渡位置(
李帅锋等, 2020 ), 保护对象主要为典型、完整连片的南亚热带季风常绿阔叶林和珍稀、特有动植物种类.该区气候受印度洋西南季风暖湿气流影响, 亚热带季风气候特征明显, 日温差大, 年温差小, 年平均气温高; 雨量充沛集中, 夏秋季多雨, 冬春季天气多晴朗, 全年干湿季分明.年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
苏建荣等, 2015 ;
李帅锋等, 2020 ).植被以季风常绿阔叶林为主, 人为干扰较小.森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤. ...
... 根据
Bellingham和Tanner (2000) 以及
苏建荣等(2015) 的划分方法, 将物种萌生类型划分为4种类型: 根萌生, 萌生位置在地下; 干基萌生, 萌生位置在地面上高度小于等于0.5 m; 干萌生, 萌生位置在地面上高度大于0.5 m, 主干受损并从残留树干上萌生; 枝萌生, 萌生位置在地面上高度大于0.5 m, 小枝受损, 从小枝与大树枝之间萌生. ...
云南中南部季风常绿阔叶林恢复生态系统萌生特征
1
2012
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
云南中南部季风常绿阔叶林恢复生态系统萌生特征
1
2012
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
戴云山高海拔黄山松林的空间异质性与植被维持机制
1
2015
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
戴云山高海拔黄山松林的空间异质性与植被维持机制
1
2015
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
基于浙江天童20 hm
2 常绿阔叶林动态监测样地的群丛划分
1
2018
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
基于浙江天童20 hm
2 常绿阔叶林动态监测样地的群丛划分
1
2018
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
1
2019
... 统计大样地内胸径(
DBH ) ≥ 1 cm的乔木、藤本与灌木物种, 建立物种数据库, 包括科、属、种, 物种中文名与拉丁名参考《中国植物志》电子版(
http://www.iplant.cn/frps )(
中国科学院中国植物志编辑委员会, 2019 )的物种名称.分析不同地形群落数量特征、物种多样性及物种-面积及物种-个体多度曲线差异. ...
1
2019
... 统计大样地内胸径(
DBH ) ≥ 1 cm的乔木、藤本与灌木物种, 建立物种数据库, 包括科、属、种, 物种中文名与拉丁名参考《中国植物志》电子版(
http://www.iplant.cn/frps )(
中国科学院中国植物志编辑委员会, 2019 )的物种名称.分析不同地形群落数量特征、物种多样性及物种-面积及物种-个体多度曲线差异. ...
1
1993
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响
2
2018
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
... ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响
2
2018
... 研究发现地形异质性对普洱大样地的物种丰富度具有显著影响, 这与
王家鸣等(2018) 对海南尖峰岭热带山地雨林的研究结论一致, 同时具有较高比例的稀有种(54.6%), 高于鼎湖山(52.3%)、哀牢山(47.1%)、古田山(37.1%)和天童山(36.2%).这与物种多样性格局主要受生境条件限制有极大的关系(
Clark et al ., 1998 ).普洱大样地中缓坡的物种丰富度最大, 稀有种和偶见种都最多, 可能的原因是该地形在大样地中分布面积较大, 仅次于山脊, 为物种生存提供更大的面积; 其次, 该地形的环境异质性强, 拥有丰富的资源, 可为更多的物种提供定居的生态位(
卜文圣, 2013 ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
... ;
王家鸣等, 2018 ).山脊面积最大, 但物种丰富度较低, 植株密度高, 说明生境限制性强, 山脊不利于水分和养分的积累从而限制了物种的定居与更新(
Li et al ., 2011 ).高谷物种丰富度最低, 但每hm
2 物种数最大.该地形大部分样方为东坡向, 阳光比较容易穿透到森林底层, 充足光照为更多物种的定居、存活和更新提供了机会.陡坡物种丰富度明显高于高谷和山脊.陡坡主要分布在箐沟两侧, 空气湿度相对较大, 而常绿木本植物多为喜湿植物, 因而成为常绿木本植物繁衍的庇护所, 有利于物种的定居与更新.生境选择的存在使得大量物种能够共存(
宫贵权等, 2007 ), 尤其是生境对稀有种、偶见种具有选择作用.不同地形可利用资源的差异导致了不同地形对不同物种的生境限制不同, 从而可以满足不同物种特异性的生态位需求(
刘文平等, 2011 ).本研究中不同地形条件下物种组成及多样性存在较大差异, 表明多样性空间格局受到局部地形因子的影响. ...
百山祖常绿阔叶林木本植物的生境相关性分析
2
2011
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
百山祖常绿阔叶林木本植物的生境相关性分析
2
2011
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
云南哀牢山中山湿性常绿阔叶林 20 hm
2 动态样地的物种组成与群落结构
4
2018
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
云南哀牢山中山湿性常绿阔叶林 20 hm
2 动态样地的物种组成与群落结构
4
2018
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 普洱30 hm
2 森林动态监测样地东西长600 m, 南北宽500 m, 样地总体上东北高、西南低, 地形复杂, 最高海拔1 586.7 m, 最低海拔1 467.4 m, 海拔高差119.3 m.样地按CTFS调查技术规范(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等, 2018 )建设, 样地设置与调查方法参考
李帅锋等(2020) 的研究. ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
... 普洱大样地小径级植株密度大, 与哀牢山样地相比, 平均胸径小于哀牢山样地(9.19 cm)(
温韩东等, 2018 ).小径级个体占总个体数的比例较高, 表明物种有足够数量的幼树和幼苗, 尤其是优势物种, 包括短刺锥、泥柯和截果柯, 对森林群落的更新具有显著的积极作用.短刺锥、泥柯和截果柯种子产量高, 且多分布在母树周围, 有更大的几率在森林环境中定居并形成幼苗库进入乔木层(
李帅锋等, 2012 ); 同时, 壳斗科物种在季风常绿阔叶林中萌生更新较多, 当存在较多的人为或自然干扰时, 它们的更新策略以萌生更新为主.萌生使物种在长时间大范围的干扰后植物种群能够继续维持, 对物种周转率、群落结构和物种组成变化及生物多样性的维持具有重要的影响(
陈沐等, 2008 ;
苏建荣等, 2012 ).山脊和缓坡植株密度较大的一个原因就是根萌生、干基萌生及干萌生个体较多.有研究表明萌生在山顶周围分布较多(
郭屹立等, 2015 ), 原因是随着光照增加, 植物体内可移动的碳水化合物含量升高, 植物萌生能力较强, 萌生数量较多.萌生存在于沟谷中, 可能的原因是外部生存威胁及内部竞争导致(
陈沐等, 2008 ). ...
浙江天童20 hm
2 常绿阔叶林动态监测样地的群落特征
2
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
浙江天童20 hm
2 常绿阔叶林动态监测样地的群落特征
2
2011
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
浙江天童国家森林公园微地形与植被结构的关系
1
2005
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
浙江天童国家森林公园微地形与植被结构的关系
1
2005
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
4
2008
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 通过物种重要值确定其优势物种, 重要值(
IV )的计算公式(
叶万辉等, 2008 )为: ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
4
2008
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
... 评估地形因子对物种组成的影响和群落结构的相对贡献是物种共存机制研究的重要内容(
Anderson et al ., 2011 ;
Liu et al ., 2014 ).森林动态监测大样地是研究群落结构、功能及动态、群落与环境因子的关系以及生物多样性维持机制等的重要平台(
兰国玉等, 2008 ;
宋永昌等, 2015 ;
马克平, 2017 ); 与传统采用随机离散小样方取样相比, 大样地更加注重地理实体空间信息的表达(
苏松锦, 2015 ), 研究的准确性更高(
姜萍等, 2005 ).因此, 不同****基于大样地对地形或生境对物种分布与共存的相对贡献进行了评估(
Chen et al ., 2010 ;
Lan et al ., 2011 ).国际上关于这方面的研究主要集中在热带森林动态监测样地内.近年, 我国关于亚热带森林地形或生境对物种分布影响的研究逐渐增加, 围绕浙江天童山(
杨庆松等, 2011 )、古田山(
宫贵权等, 2007 )、百山祖(
王伟等, 2011 ), 云南哀牢山(
温韩东等, 2018 )及广东鼎湖山(
刘文平等, 2011 )等5个固定监测样地开展了相关研究.但是, 关于南亚热带季风常绿阔叶林物种多样性维持机制的研究还不够充分(
叶万辉等, 2008 ), 对群落中植物与环境之间相互关系的研究还相对缺乏. ...
... 通过物种重要值确定其优势物种, 重要值(
IV )的计算公式(
叶万辉等, 2008 )为: ...
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响
1
2014
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响
1
2014
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
川西亚高山森林恢复的空间格局分析
1
2005
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
川西亚高山森林恢复的空间格局分析
1
2005
... 异质的环境因子是大量物种得以稳定共存的重要因素(
Tilman & Pacala, 1993 ), 环境异质性是自然植物群落物种多样性的重要维持机制(
Chase, 2003 ;
叶万辉等, 2008 ;
Shen et al ., 2009 ;
王伟等, 2011 ).在众多的生境因子中, 地形越来越受到重视.在局域尺度上, 地形是重要的非地带性因素和植物群落生境异质性的重要来源, 也是影响植被格局的重要因子(
杨永川等, 2005 ).局部地形通过土壤养分、水分等资源的空间再分配对立地条件产生影响(
Forrester et al ., 2003 ;
汝海丽等, 2015 ).在气候相对一致的情况下, 地形是群落物种多样性的决定因子(
Bell et al ., 2000 ), 植物多样性的维持很大程度上依赖于地形异质性(
Hofer et al ., 2008 ;
Lundholm, 2009 ).地形对物种组成及多样性的影响, 又可分为海拔、坡度、坡向和凹凸度等地形因子对物种组成和多样性的影响(
沈泽昊等, 2000 ;
张远东等, 2005 ).海拔是影响水热条件的主导因子(
袁铁象等, 2014 ), 物种丰富度在区域尺度上一般随海拔的升高而下降(
Sharma et al ., 2009 ), 凹凸度在土壤水分、养分分配方面具有重要作用, 与坡向、坡度一起间接导致了物种组成和多样性的差异(
Pigott, 1975 ;
Hennenberg & Bruelheide, 2003 ). ...
Floristic divergence of the evergreen broad-leaved forests in Yunnan, southwestern China
1
2019
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
1
2008
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
1
2008
... 普洱大样地植被类型为季风常绿阔叶林, 与哀牢山(
温韩东等, 2018 )、天童山(
杨庆松等, 2011 )、古田山(
祝燕等, 2008 )和鼎湖山(
叶万辉等, 2008 )植被类型一致.本研究表明普洱大样地每hm
2 物种数为9种, 高于哀牢山(5种·hm
-2 )、天童山(8种·hm
-2 )、古田山(7种·hm
-2 ) 3个森林动态监测大样地, 其原因可能是普洱大样地面积更大, 包含的物种可能更多, 地形对物种的影响更大, 同时普洱大样地位于热带边缘向亚热带过渡的区域, 群落内包含较多的热带区系组成部分(
Zhu, 2019 ), 群落交错带效应明显; 普洱大样地与鼎湖山(11种·hm
-2 )大样地相比, 纬度相差不大, 而海拔较高, 因而物种丰富度低于鼎湖山大样地. ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT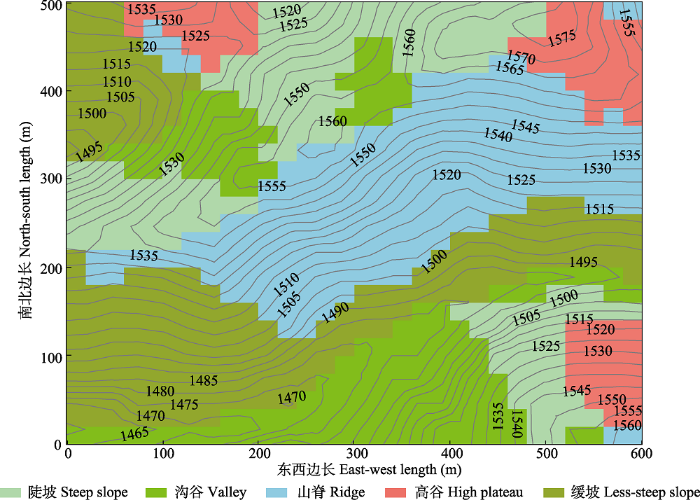 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT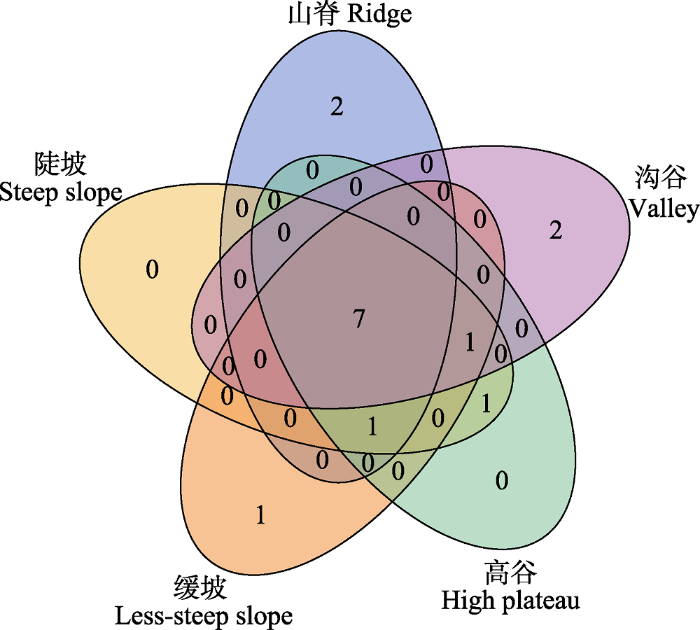 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT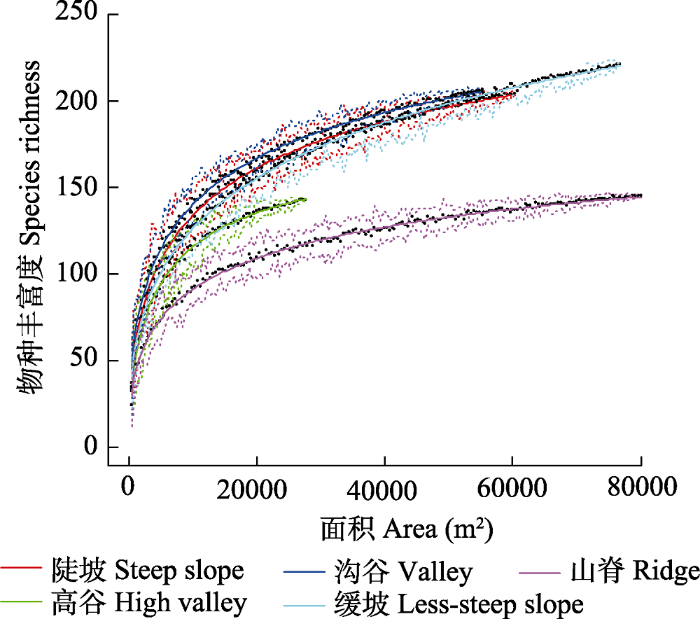 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT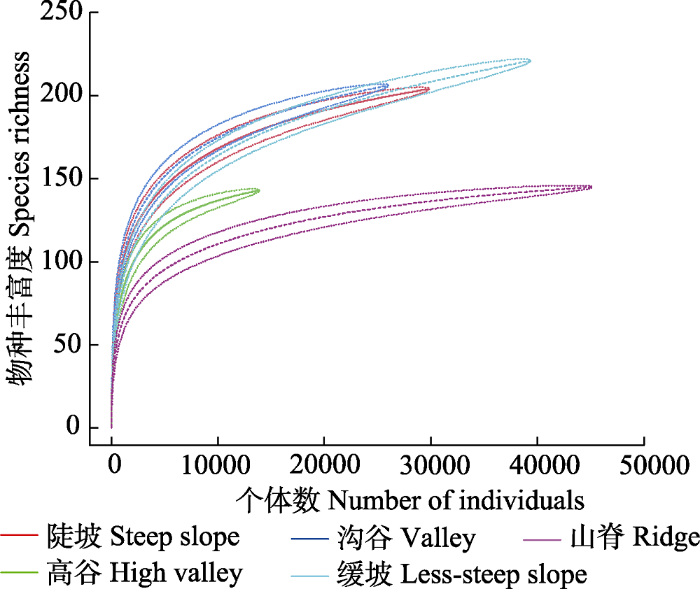 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT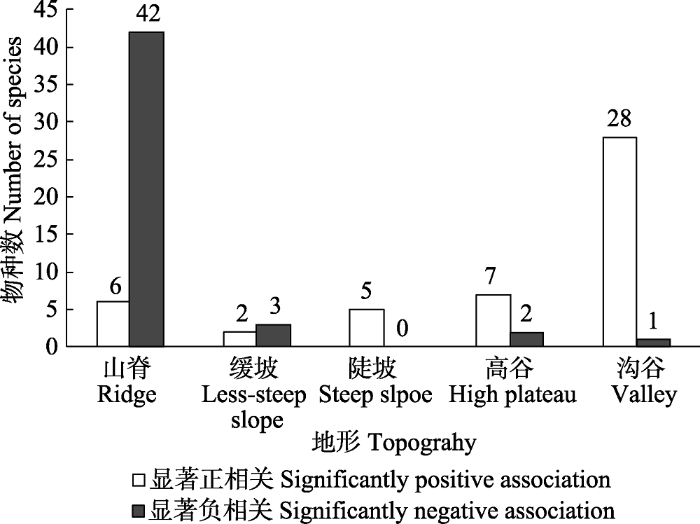 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}