 ,1,2,*
,1,2,*Basic characteristics of Stipa sareptana var. krylovii communities in China
Shuai-Zhi LU1,2, Xian-Guo QIAO1,2, Li-Qing ZHAO3, Zi WANG1,2, Chen-Guang GAO4, Jing WANG1,2, Ke GUO,1,2,*通讯作者: *guoke@ibcas.ac.cn
编委: 王德利
责任编辑: 李敏
收稿日期:2020-03-17网络出版日期:2020-10-20
| 基金资助: |
Corresponding authors: *guoke@ibcas.ac.cn
Received:2020-03-17Online:2020-10-20
| Fund supported: |

摘要
西北针茅(Stipa sareptana var. krylovii)群系是亚洲中部地区特有的典型草原之一, 也是生态适应性最广的草原类型, 向东可以在呼伦贝尔高原与草甸草原重叠分布, 向西可以在乌兰察布高原、天山等地区与荒漠草原形成复合分布, 向南可分布至黄土高原与暖温带草原镶嵌分布, 还可在青藏高原的东缘与高寒草原混生。该研究调查了中国西北针茅群系的主要植被类型, 通过对117个样地的调查数据分析, 量化描述了该群系的基本群落特征。结果表明, 中国西北针茅群系共有种子植物336种, 分属于36科131属, 物种数大于15的科有禾本科、菊科、豆科、蔷薇科、藜科和百合科; 物种存在度等级划分中, I级(0-20%)植物占比91.67%, 多为群落中的偶见种或稀有种, 最为常见的植物除西北针茅外, 也有糙隐子草(Cleistogenes squarrosa)、阿尔泰狗娃花(Heteropappus altaicus)、冰草(Agropyron cristatum)、
关键词:
Abstract
Stipa sareptana var. krylovii alliance is one of the typical steppes endemic to Central Asia. The eastern border of its distribution is adjacent to the meadow steppes on the Hulunbeir Plateau, the western border extends to the desert steppes on the Ulanqabu Plateau and Tianshan Mountains, and the southern border lies among the warm temperate steppes on the Loess Plateau. The distribution of this alliance also spreads into the alpine steppes on the eastern margin of the Qinghai-Xizang Plateau. The broad its distribution suggests that S. sareptana var. krylovii alliance has wide ecological adaptability. In this study, we described and analyzed the eco-geographical distribution, community characteristics and classification of S. sareptana var. krylovii alliance based on data from 117 plots. The results showed that there are 336 seed plants belonging to 36 families and 131 genera in these plots. The families with more than 15 species are Gramineae, Compositae, Leguminosae, Rosaceae, Chenopodiaceae and Liliaceae. Among all species in these plots, 91.67% are rare species with occurrence frequency smaller than 20%. Besides S. sareptana var. krylovii, the most common species in this alliance include Cleistogenes squarrosa, Heteropappus altaicus, Agropyron cristatum, Koeleria cristata, Potentilla bifurca, Salsola collina, Allium tenuissimum and Leymus chinensis. In addition, hemicryptophytes are the most common life form within the surveyed plots, accounting for 66.37% of all species. Xerophytes are the most common water ecological type, accounting for 61.19% of all species. East Palearctic element was the major floristic geographical element, accounting for 33.33% of all species. Based on the China Vegetation Classification System, S. sareptana var. krylovii alliance could be classified into 6 association groups and further into 45 associations.
Keywords:
PDF (1003KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
陆帅志, 乔鲜果, 赵利清, 王孜, 高趁光, 王静, 郭柯. 中国西北针茅草原的基本群落特征. 植物生态学报, 2020, 44(10): 1087-1094. DOI: 10.17521/cjpe.2020.0072
LU Shuai-Zhi, QIAO Xian-Guo, ZHAO Li-Qing, WANG Zi, GAO Chen-Guang, WANG Jing, GUO Ke.
西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区。西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(中国植被编辑委员会, 1980; 中国科学院内蒙古宁夏综合考察队, 1985; 周兴民等, 1987; 黄大燊, 1997; 马子清, 2001)。
建群种西北针茅, 是多年生旱生丛生禾草(内蒙古植物志编辑委员会, 1998)。中国的内蒙古、河北、山西、宁夏、甘肃、青海、新疆和西藏都有分布, 生长在海拔440-4 510 m的山前洪积扇、平坦地或河谷阶地上(中国科学院中国植物志编辑委员会, 2004)。在形态特征上, 西北针茅与新疆针茅(S. sareptana)和针茅(S. capillata)极其接近, 因此一些植物学家将其作为新疆针茅的变种或亚种(郭本兆和孙永华, 1982; Wu et al., 2006), 但是更多的植物学家将其作为独立的种(Lomonosova, 1990; Gudkova, 2012; Nobis & Gudkova, 2016)。三者在营养枝叶舌长度、叶片内表面的被毛情况、外稃被毛情况, 以及适应生境的差异较为明显(Gudkova et al., 2017; 表1)。西北针茅与针茅的区别: 西北针茅外稃顶端有一圈毛环, 针茅没有; 西北针茅与新疆针茅的区别: 西北针茅营养枝叶片远轴面光滑或微粗糙, 新疆针茅有长达0.25 mm的毛而比较粗糙; 针茅与新疆针茅的区别: 新疆针茅的外稃、基盘和颖长都短于针茅。
Table 1
表1
表1西北针茅、针茅和新疆针茅形态特征区别(Gudkova et al., 2017)
Table 1
| 形态特征 Charaters | 针茅 S. capillata | 西北针茅 S. krylovii | 新疆针茅 S. sareptana |
|---|---|---|---|
| 茎秆高度 Length of culm (cm) | 30-130 | 20-60 | 30-80 |
| 叶片宽度 The leaf blade width (mm) | 0.5-1.3 | 0.3-0.6 | 0.4-0.7 |
| 营养枝叶片远轴面 Abaxial surface of vegetative leaves | 光滑或粗糙 Glabrous or scabrous | 光滑, 极少粗糙 Glabrous or rarely somewhat scabrous | 粗糙, 表面有0.25 mm长的微刺 Scabrous due to 0.25 mm long spinules |
| 营养枝叶片近轴面 Adaxial surface of vegetative leaves | 具0.2-0.5 mm的毛 With hair 0.2-0.5 mm long | 具0.05-0.1 mm的毛 With hair 0.05-0.1 mm long | 具0.1 mm的毛, 边缘毛更长 With hair 0.1 mm long, admixture of longer hairs near the margins |
| 营养枝叶舌长度 Ligules of vegetative leaves (mm) | (0.6) 1-1.5 (2.5) | 0.1-0.3 | 0.2-1.0 |
| 芒长 Length of the awn (cm) | 10-18 | (8) 12-16 (20) | 10-15 |
| 芒柱长 Length of column (cm) | 3-5 (7.5) | (1.8) 2.5-3.5 (4) | 2.5-5 |
| 外稃长 Length of anthecium (mm) | 9-14 | (8.5) 9-12 | 9-11 |
| 基盘长 Length of callus (mm) | 3.0-4.5 | (2.2) 2.4-3.5 (4.5) | 2-3 |
| 外稃顶端的毛环 Coronula of hairs at the top of lemma | 缺失 Absent | 存在 Present | 存在或稀疏 Present rarely reduced |
新窗口打开|下载CSV
1 研究方法
1.1 调查方法
野外调查于2010-2017年生长季在西北针茅群系的分布区域展开, 包括呼伦贝尔高原、锡林郭勒高原、乌兰察布高原、黄土高原、河西走廊等地理单元, 以及祁连山、天山等荒漠区山地。每个样地选取3-5个具有代表性的1 m × 1 m的样方调查。样方的基本描述, 包括样方编号、调查日期、经纬度、海拔高度、地形、干扰情况、斑块类型、样方平均高度、平均盖度等。其次, 对于样方内每一个物种, 都测量高度, 目视估计分盖度, 数算株/丛数, 最后齐地面收获各物种的地上生物量, 带回实验室在65 ℃的烘箱内烘干后称质量。对调查过程中不能确定的物种, 采集标本并带回室内进行鉴定。
1.2 数据处理及分析
西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(中国科学院中国植物志编辑委员会, 2004)为依据来确定, 物种生活型、水分生态类型和区系地理成分的确定参考相关文献(中国植被编辑委员会, 1980; 吴玉虎, 2008; 赵一之, 2012)。样地内物种重要值(IV)的计算是基于3-5个样方的平均盖度、平均高度和平均生物量, 计算单个物种的盖度、高度和生物量占样地内所有物种的百分比, 即相对盖度、相对高度和相对生物量, 采用公式“IV = (相对盖度+相对高度+相对生物量)/3”, 在生物量缺失的样方中, 采用公式“IV = (相对盖度+相对高度)/2”。最后以建群种和次优势层/次优势层片优势物种命名群丛组和群丛(陈灵芝等, 2014)。为表达群落的气候特征, 下载Worldclim (
2 结果和分析
2.1 西北针茅群系的分布与环境特征
西北针茅群系在中国主要分布在呼伦贝尔高原中西部、锡林郭勒高原和黄土高原东北部等典型草原分布区。另外, 在草甸草原区的阳坡, 如河北西北部、山西北部等; 荒漠草原区的丘陵阴坡, 如乌兰察布高原、阴山等; 以及荒漠区山地, 如祁连山、天山、阿拉套山、塔尔巴哈台山等, 也有分布。在黄土高原与青藏高原接壤的地区, 如青海湖周边地区, 也有较多分布。根据117个样地(见附录)统计, 西北针茅群系主要发育在典型的栗钙土上, 也可出现在棕钙土或淡黑钙土上。分布区的平均海拔为1 653.23 m, 最高海拔达3 467.00 m, 分布在青海省贵南县拉脊山; 最低海拔是549 m, 分布在内蒙古呼伦贝尔的新巴尔虎右旗。平均年降水量为274.56 mm, 最小值为64.00 mm, 最大值是525.00 mm; 平均最暖季降水量172.80 mm, 最高298.00 mm, 最低44.00 mm; 平均最冷季降水量10.36 mm, 最低2.00 mm, 最高65.00 mm。年平均气温为0.43 ℃, 最冷年平均气温-6.22 ℃, 最高年平均气温6.93 ℃; 最暖季平均气温16.03 ℃, 最低6.05 ℃, 最高20.98 ℃; 最冷季平均气温-16.44 ℃, 最高-5.58 ℃, 最低-30.20 ℃。
2.2 西北针茅群系的群落特征
2.2.1 群落数量及结构特征西北针茅群系的群落平均高度为34.07 cm, 最低可达10.17 cm, 最高可达65.67 cm; 群落平均盖度38.39%, 最小7.40%, 最大98.23%; 群落平均生物量111.61 g·m-2, 最低22.76 g·m-2, 最高353.13 g·m-2; 丰富度是13.62种·m-2, 最少3种·m-2, 最大27种·m-2。
2.2.2 物种组成
西北针茅群系在117个调查样地中共记录到种子植物336种, 另有未鉴定到种的植物12种, 包括黄耆属(Astragalus)、薹草属(Carex)、棘豆属(Oxytropis)、绢蒿属(Seriphidium)、鸢尾属(Iris)、葱属(Allium)、柴胡属(Bupleurum)、蒿属(Artemisia)和伞形科等的植物。含10个种以上的科有禾本科(53种)、菊科(51种)、豆科(42种)、蔷薇科(21种)、藜科(19种)、百合科(16种)、莎草科(13种)、石竹科(12种)、唇形科(12种)和十字花科(10种)。含5个种以上的属有委陵菜属(Potentilla, 15种)、针茅属(Stipa, 14种)、蒿属(14种)、黄耆属(13种)、葱属(12种)、棘豆属(12种)、薹草属(10种)、鸢尾属(8种)、鸦葱属(Scorzonera, 7种)、柴胡属(6种)、点地梅属(Androsace, 6种)、锦鸡儿属(Caragana, 5种)、石头花属(Gypsophila, 5种)、早熟禾属(Poa, 5种)、藜属(Chenopodium, 5种)、青兰属(Dracocephalum, 5种)和芨芨草属(Achnatherum, 5种)。
存在度是某个种在有多个样地记录的群落中出现于样地记录的百分数(宋永昌, 2001)。按照其出现次数可以将其划分为5级: 0 < I ≤ 20%; 20% < II ≤ 40%; 40% < III ≤ 60%; 60% < IV ≤ 80%; 80% < V ≤ 100% (图1)。V级植物只有西北针茅一种, 存在度100%。IV级物种有糙隐子草(Cleistogenes squarrosa)和阿尔泰狗娃花(Heteropappus altaicus), 存在度分别是64.11%和62.39%, 为群落的伴生种或亚优势种。III级植物有6种, 冰草(Agropyron cristatum)、
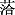
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1西北针茅群系物种的存在度等级分布。0 < I ≤ 20%; 20% < II ≤ 40%; 40% < III ≤ 60%; 60% < IV ≤ 80%; 80% < V ≤ 100%。
Fig. 1The frequency distribution of species with different presence class in Stipa sareptana var. krylovii alliance. 0 < I ≤ 20%; 20% < II ≤ 40%; 40% < III ≤ 60%; 60% < IV ≤ 80%; 80% < V ≤ 100%.
2.2.3 生活型及水分生态类型
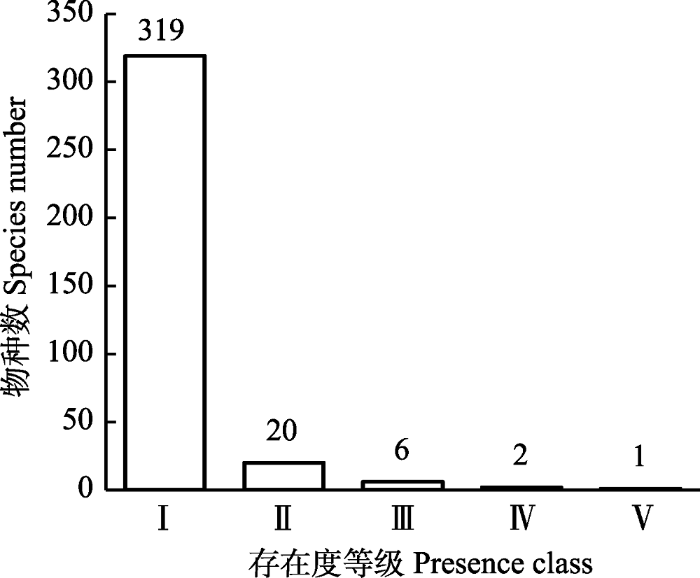
西北针茅群系中的物种按照《中国植被》的生活型系统可划分为6类(图2), 一年生草本、多年生杂类草、多年生根茎禾草、多年生丛生禾草、半灌木、灌木。半灌木和灌木一共24种, 比较常见的有狭叶锦鸡儿(Caragana stenophylla)、木地肤(Kochia prostrata)、燥原荠(Ptilotricum canescens)、百里香(Thymus mongolicus)、小叶锦鸡儿(Caraganamicrophylla)等。多年生丛生禾草主要是针茅属的植物, 多年生杂类草在委陵菜属、黄耆属、鸢尾属、柴胡属、葱属等较多, 多年生根茎禾草以赖草属植物居多。一年生植物在群落中占据17.86%, 以藜科和禾本科一年生草本较多, 常见的有猪毛菜、刺藜(Chenopodium aristatum)、尖头叶藜(Chenopodium acuminatum)、猪毛蒿(Artemisia scoparia)、藜(Chenopodium album)、鹤虱(Lappula myosotis)、画眉草(Eragrostis pilosa)等。
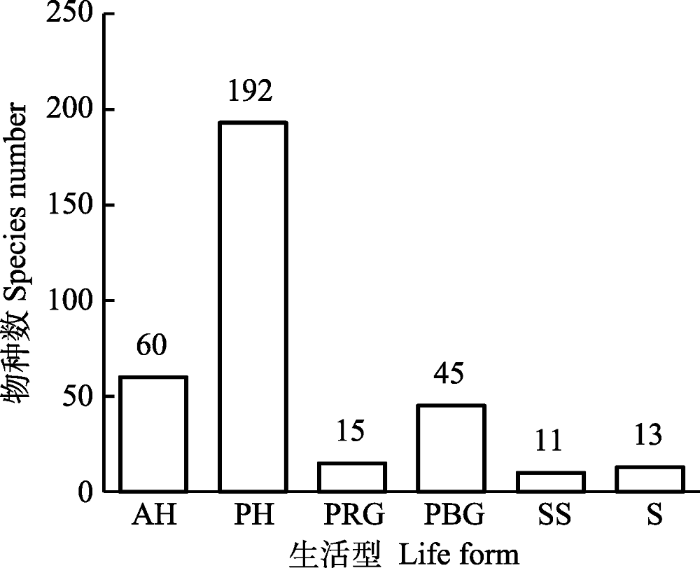
按照物种水分生态类型可划分为6类(图3)。湿生植物只有1种: 毛薹草(Carex lasiocarpa), 常生长于水分充足的沼泽等生境, 也可偶见于草原区域。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2西北针茅群系物种的生活型谱。AH, 一年生草本植物; PBG, 多年生丛生禾草; PH, 多年生杂类草; PRG, 多年生根茎禾草; S, 灌木植物; SS, 半灌木植物。
Fig. 2Frequency distribution of species with different life forms in Stipa sareptana var. krylovii alliance. AH, annual herbs; PBG, perennial bunch grasses; PH, perennial herbs; PRG, perennial rhizome grasses; S, shrubs; SS, semi-shrubs.
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3西北针茅群系物种的水分生态类型谱。HP, 湿生植物; MP, 中生植物; MX, 中旱生植物; SX, 超旱生植物; TX, 旱生植物; XM, 旱中生植物。
Fig. 3Frequency distribution of species belonging to different water ecological types in Stipa sareptana var. krylovii alliance. HP, hygrophyte; MP, mesophytes; MX, meso-xerophytes; SX, super-xerophytes; TX, typical xerophytes; XM, xero-mesophytes.
中生植物一共有129种, 占38.50%, 可继续划分为典型中生和旱中生植物。旱生植物比例最大, 有205种, 占61.19%, 典型旱生种类多于中旱生植物, 荒漠区的超旱生植物也较多地出现在西北针茅群系, 其中以碱韭(Allium polyrhizum)、荒漠镰芒针茅(Stipa caucasica subsp. desertorum)、白皮锦鸡儿(Caragana leucophloea)、矮锦鸡儿(Caragana pygmaea)、双穗麻黄(Ephedra distachya)、胶黄耆状棘豆(Oxytropis tragacanthoides)、灌木亚菊(Ajania fruticulosa)等较常见。
2.2.4 物种区系地理成分
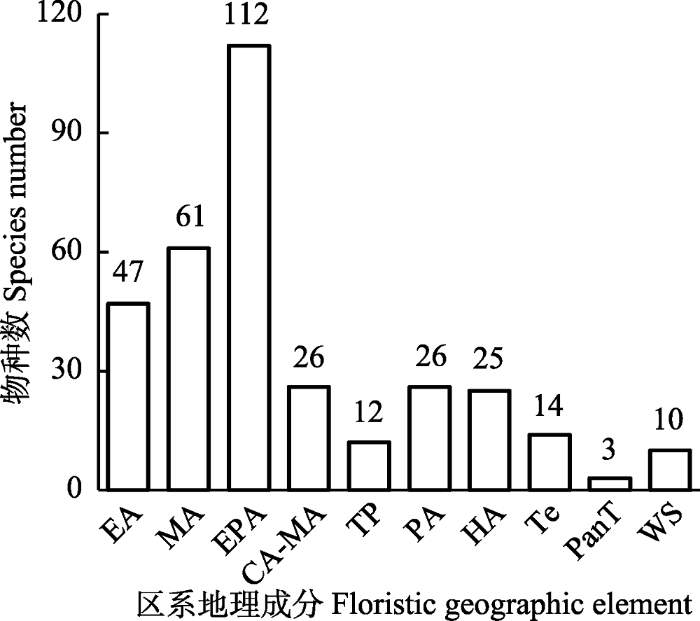
西北针茅群系的物种按照区系地理成分可以划分为10类(图4)。东古北极成分最多, 占总物种数的33.33%。亚洲中部成分其次, 占18.15%, 东亚成分、古北极和泛北极成分在群落中也较为常见。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4西北针茅群系物种区系地理成分组成。CA-MA, 中亚-亚洲中部成分; EA, 东亚分布种; EPA, 东古北极成分; HA, 泛北极成分; MA, 亚洲中部成分; PA, 古北极成分; PanT, 泛温带成分; Te, 古地中海北极成分; TP, 青藏高原分布种; WS, 世界分布种。
Fig. 4Frequency distribution of species belonging to different floristic geographic elements in Stipa sareptana var. krylovii alliance. CA-MA, central Asia-middle Asia element; EA, east Asia element; EPA, east Palearctic element; HA, Holarctic element; MA, middle Asia element; PA, Palearctic element; PanT, pan temperate element; Te, Tethys element; TP, Qinghai-Xizang Plateau element; WS, world spread element.
2.3 西北针茅群系分类
依照陈灵芝等(2014)中的植被分类原则和系统、植物群落命名原则对西北针茅群系进行分类和命名。根据生活型层片及物种优势度将117个西北针茅群系样地划分为6个群丛组类型, 45个群丛类型(表2)。Table 2
Table 2The classification of Stipa sareptana var. krylovii steppe
| 群丛组类型 Association group | 群丛类型 Association | 样地编号 Plot number |
|---|---|---|
| 西北针茅、丛生禾草或莎草 S. sareptana var. krylovii, bunchgrasses or bunch sedges | 西北针茅+冰草 S. sareptana var. krylovii + Agropyron cristatum | P1, P26, P27, P28, P79, P101, P110, P115 |
| 西北针茅+糙隐子草 S. sareptana var. krylovii + Cleistogenes squarrosa | P17, P25, P44, P46, P52, P57, P58, P60, P62, P67, P93, P95, P106, P113 | |
| 西北针茅+短花针茅 S. sareptana var. krylovii + S. breviflora | P64, P82, P84 | |
| 西北针茅+ | P16 | |
| 西北针茅+瑞士羊茅 S. sareptana var. krylovii + Festuca valesiaca | P13 | |
| 西北针茅+新疆针茅S. sareptana var. krylovii + S. sareptana | P19 | |
| 西北针茅+羊茅 S. sareptana var. krylovii + Festuca sp. | P31 | |
| 西北针茅+沙芦草 S. sareptana var. krylovii + Agropyron mongolicum | P34 | |
| 西北针茅+华灰早熟禾 S. sareptana var. krylovii + Poa sinoglauca | P85 | |
| 西北针茅+硬质早熟禾 S. sareptana var. krylovii + Poa sphondylodes | P89 | |
| 西北针茅+紫花针茅 S. sareptana var. krylovii + S. purpurea | P91 | |
| 西北针茅+大针茅 S. sareptana var. krylovii + S. grandis | P109 | |
| 西北针茅+嵩草 S. sareptana var. krylovii + Kobresia sp. | P86 | |
| 西北针茅、根茎禾草或莎草 S. sareptana var. krylovii rhizomatous grasses or sedges | 西北针茅+羊草 S. sareptana var. krylovii + Leymus chinensis | P2, P45, P47, P48, P50, P53, P96, P105, P107, P114 |
| 西北针茅+干生薹草 S. sareptana var. krylovii + Carex aridula | P3, P8, P80, P83, P87, P88 | |
| 西北针茅+赖草 S. sareptana var. krylovii + Leymus secalinus | P12, P30, P42, P92, | |
| 西北针茅+寸草 S. sareptana var. krylovii + Carex duriuscula | P98, P102 | |
| 西北针茅+窄叶薹草 S. sareptana var. krylovii + Carex montis-everestii | P9 | |
| 西北针茅、杂类草 S. sareptana var. krylovii, forbs | 西北针茅+柔毛蒿 S. sareptana var. krylovii + Artemisia pubescens | P49, P65, P73, P74, P75, P90 |
| 西北针茅+星毛委陵菜 S. sareptana var. krylovii + Potentilla acaulis | P18, P24, P35, P38, P41, P112 | |
| 西北针茅+银灰旋花 S. sareptana var. krylovii+ Convolvulus ammannii | P11, P68, P72 | |
| 西北针茅+碱韭 S. sareptana var. krylovii + Allium polyrhizum | P61, P66, P100, P103 | |
| 西北针茅+棘豆 S. sareptana var. krylovii + Oxytropis sp. | P32, P33 | |
| 西北针茅+砂韭 S. sareptana var. krylovii + Allium bidentatum | P55, P56 | |
| 西北针茅+绢蒿 S. sareptana var. krylovii + Seriphidium sp. | P116, P117 | |
| 西北针茅+蓝花棘豆 S. sareptana var. krylovii + Oxytropis caerulea | P10 | |
| 西北针茅+香叶蒿 S. sareptana var. krylovii + Artemisia rutifolia | P14 | |
| 西北针茅+达乌里芯芭 S. sareptana var. krylovii+ Cymbaria dahurica | P51 | |
| 西北针茅+麻花头 S. sareptana var. krylovii + Serratula centauroides | P54 | |
| 西北针茅+北柴胡 S. sareptana var. krylovii + Bupleurum chinense | P59 | |
| 西北针茅+草麻黄 S. sareptana var. krylovii + Ephedra sinica | P63 | |
| 西北针茅+北芸香 S. sareptana var. krylovii + Haplophyllum dauricum | P71 | |
| 西北针茅+甘青青兰 S. sareptana var. krylovii + Dracocephalum tanguticum | P76 | |
| 西北针茅+西山委陵菜 S. sareptana var. krylovii + Potentilla sischanensis | P77 | |
| 西北针茅+青海苜蓿 S. sareptana var. krylovii + Medicago archiducis-nicolai | P78 | |
| 西北针茅+小果黄耆 S. sareptana var. krylovii + Astragalus tataricus | P81 | |
| 西北针茅+草木犀状黄耆 S. sareptana var. krylovii + Astragalus melilotoides | P99 | |
| 西北针茅、一年生草本 S. sareptana var. krylovii, annual grasses | 西北针茅+猪毛菜 S. sareptana var. krylovii + Salsola collina | P69, P70, P108 |
| 西北针茅+猪毛蒿 S. sareptana var. krylovii + Artemisia scoparia | P29 | |
| 西北针茅+尖头叶藜 S. sareptana var. krylovii + Chenopodium acuminatum | P94 | |
| 西北针茅、半灌木 S. sareptana var. krylovii, dwarf shrubs | 西北针茅-白莲蒿 S. sareptana var. krylovii - Artemisia sacrorum | P4 |
| 西北针茅-百里香 S. sareptana var. krylovii - Thymus mongolicus | P5 | |
| 西北针茅-灌木亚菊 S. sareptana var. krylovii - Ajania fruticulosa | P6 | |
| 西北针茅 S. sareptana var. krylovii | 西北针茅 S. sareptana var. krylovii | P21, P36, P37, P39, P97, P111 |
新窗口打开|下载CSV
西北针茅、多年生丛生禾草或莎草群丛组: 该群丛组共记录到35个样地, 14个群丛类型。西北针茅+糙隐子草群丛最为常见。糙隐子草是中亚-亚洲中部地区草原的稳定伴生种和次优势种, 因此该群丛在中国的分布跨度也比较大, 从内蒙古的新巴尔虎左旗、新巴尔虎右旗, 经锡林郭勒的苏尼特左旗、西乌珠穆沁旗到新疆的吉木乃县、裕民县都有比较广泛的分布。在不同的区域和生境中, 西北针茅可与针茅属的不同植物形成不同的群丛, 成为西北针茅群系与其他针茅群系之间的过渡类型。例如, 西北针茅+短花针茅群丛在青海省共和县和内蒙古苏尼特左旗记录到3个样地, 分布在栗钙土或棕钙土上, 是偏干旱的类型; 西北针茅+大针茅群丛在锡林浩特有分布; 西北针茅+紫花针茅群丛在甘肃省的祁连山山地有分布, 偏高寒性质, 是向紫花针茅草原逐渐过渡的类型; 西北针茅+新疆针茅群丛在新疆和布克赛尔县有分布。
西北针茅+冰草群丛主要分布在偏沙化的基质上, 群落中的细沙植物也比较多, 比如砂韭、蒙古韭、沙生针茅、石生针茅(Stipa tianschanica var. klemenzii)、沙生薹草等; 另外, 群落中的退化指示植物的优势度较高, 比如冷蒿、
西北针茅、多年生根茎禾草或莎草群丛组: 西北针茅+羊草群丛是比较典型的西北针茅草原类型, 分布区域偏东, 主要分布在呼伦贝尔地区, 在锡林郭勒和张北也有小面积的分布, 群落中生境较为湿润, 丰富度也略高。另外, 干生薹草(Carex aridula)、赖草、窄叶薹草(Carex montis-everestii)和寸草也可成为群落的亚优势种形成相应的群丛类型。
西北针茅、多年生杂类草群丛组: 西北针茅+星毛委陵菜群丛在内蒙古乌兰察布、包头、锡林郭勒, 新疆托里县、和布克赛尔有记载。星毛委陵菜是群落退化的标志性植物, 这个群丛所在的草场均是处于中度或重度放牧压力之下, 且土壤已经明显的沙化而成为砂壤土或粉砂土。西北针茅+碱韭群丛和西北针茅+砂韭群丛分布在西北针茅草原区东部的草原风沙土或砂性的栗钙土上。另外, 蓝花棘豆(Oxytropis caerulea)、西山委陵菜、青海苜蓿(Medicago archiducis-nicolai)、草木樨状黄耆(Astragalus melilotoides)、麻花头等中生杂类草和柔毛蒿、银灰旋花、香叶蒿(Artemisia rutifolia)、达乌里芯芭、草麻黄、甘青青兰(Dracocephalum tanguticum)、绢蒿、北柴胡(Bupleurum chinense)、北芸香等旱生杂类草也可成为群落的次优势种形成相应的群丛类型。
西北针茅、一年生草本群丛组: 一年生草本多是常见的农田杂草, 猪毛菜是草原中最常见的一年生草本植物, 这类群丛是受降雨或人为干扰影响较大的类型。
西北针茅、半灌木群丛组: 西北针茅-冷蒿群丛是草原过度利用下的退化变型, 分布较广, 在内蒙古乌兰察布、呼伦贝尔, 新疆吉木乃县、托里县, 以及甘肃白银和张掖地区都有分布。西北针茅-百里香群丛分布在山西右玉县风蚀性的土壤环境中, 其他还包括西北针茅-灌木亚菊群丛和西北针茅-白莲蒿群丛。
西北针茅群丛组: 该群丛组/群丛共记录到6个样地。群落中无明显的次优势种, 形成较纯的西北针茅草原景观, 群落盖度最高可达95%。在新疆托里县, 内蒙古乌兰察布、锡林浩特和呼和浩特有分布。
3 结论和讨论
1)西北针茅群系在中国的呼伦贝尔高原、锡林郭勒高原、乌兰察布高原、青藏高原东缘以及祁连山、天山等荒漠区的山地有分布, 发育在典型栗钙土上。群落中物种以菊科、禾本科、豆科和蔷薇科居多, 针茅属、委陵菜属、蒿属、葱属、棘豆属和黄耆属在草原中作用显著。群落中的旱生植物多于中生植物, 多年生杂类草、东古北极成分和亚洲中部成分居多。冰草、糙隐子草、羊草、冷蒿、柔毛蒿、星毛委陵菜等是群落中常见的亚优势种。2)西北针茅群系的广幅适应性, 使得其东界和西界可以分别作为草甸草原与典型草原、典型草原与荒漠草原的过渡区。与蒙古高原地区同样是以光芒组植物建群的大针茅群系、狼针草(Stipa baicalensis)群系和长芒草群系相比, 西北针茅群系的分布区偏西或北, 降水量较少, 群落盖度、生物量和丰富度都偏低(乔鲜果, 2019)。群落中的湿生和中生植物较少, 旱生植物尤其是典型旱生植物和强旱生植物明显较多。麻花头、硬质早熟禾、兴安胡枝子(Lespedeza daurica)、草木樨状黄耆等植物的优势度明显较低; 耐牧压植物如寸草、银灰旋花、星毛委陵菜、冷蒿等优势度明显增加, 羊草、
3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(乔鲜果, 2019)。但是, 西北针茅群系是最重要的放牧场和打草场, 因不合理利用引起的土地覆盖从草地转变为未利用地的比例相对更大(李绍良等, 2002; 曹鑫等, 2006; Eckert et al., 2015)。而且在西北针茅群系的调查中发现, 中度和重度放牧引起的退化现象非常普遍, 群落中冷蒿、星毛委陵菜、冰草、糙隐子草等退化指示植物优势度较高。因此, 西北针茅群系在草场恢复、放牧管理上应该及时采取措施以免进一步的退化。
致谢
感谢中国科学院植物研究所潘庆民研究员提供的部分样地数据, 感谢中国科学院地理科学与资源研究所赵海卫和内蒙古大学旭日、要振宇在野外调查中的帮助。附录 中国西北针茅草原样方数据
Supplement Plot data of Stipa sareptana var. krylovii communities in Chinahttps://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0072-D1.xlsx

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.//
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
基于遥感的草原退化人为因素影响趋势分析
1
2006
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
基于遥感的草原退化人为因素影响趋势分析
1
2006
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
2
2014
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
... 依照
2
2014
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
... 依照
Trend analysis of MODIS NDVI time series for detecting land degradation and regeneration in Mongolia
1
2015
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
WorldClim 2: new 1 km spatial resolution climate surfaces for global land areas
1
2017
... 为表达群落的气候特征, 下载Worldclim (
Annotated list of the Stipa L. species (Poaceae) from South Sibiria
1
2012
... 建群种西北针茅, 是多年生旱生丛生禾草(
Stipa krylovii Roshev. (Poaceae), a new record for the Flora of Nepal.
3
2017
... 建群种西北针茅, 是多年生旱生丛生禾草(
... 西北针茅、针茅和新疆针茅形态特征区别(
... Main differences in the characteristics of Stipa sareptana var. krylovii, S. capillata and S. sareptana (
中国针茅属分类、分布和生态的初步研究
1
1982
... 建群种西北针茅, 是多年生旱生丛生禾草(
中国针茅属分类、分布和生态的初步研究
1
1982
... 建群种西北针茅, 是多年生旱生丛生禾草(
1
1997
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
1
1997
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
土壤退化与草地退化关系的研究
1
2002
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
土壤退化与草地退化关系的研究
1
2002
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
Stipa L
1
1990
... 建群种西北针茅, 是多年生旱生丛生禾草(
1
2001
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
1
2001
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
1
1985
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
1
1985
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
Taxonomic notes on feather grasses (Poaceae: Stipa) from eastern Kazakhstan with typification of seven names and one new combination
1
2016
... 建群种西北针茅, 是多年生旱生丛生禾草(
中国针茅草原群落生态学研究
2
2019
... 2)西北针茅群系的广幅适应性, 使得其东界和西界可以分别作为草甸草原与典型草原、典型草原与荒漠草原的过渡区.与蒙古高原地区同样是以光芒组植物建群的大针茅群系、狼针草(Stipa baicalensis)群系和长芒草群系相比, 西北针茅群系的分布区偏西或北, 降水量较少, 群落盖度、生物量和丰富度都偏低(
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
中国针茅草原群落生态学研究
2
2019
... 2)西北针茅群系的广幅适应性, 使得其东界和西界可以分别作为草甸草原与典型草原、典型草原与荒漠草原的过渡区.与蒙古高原地区同样是以光芒组植物建群的大针茅群系、狼针草(Stipa baicalensis)群系和长芒草群系相比, 西北针茅群系的分布区偏西或北, 降水量较少, 群落盖度、生物量和丰富度都偏低(
... 3)西北针茅群系的分布区更干旱一些, 农业发展潜力不如狼针草群系、大针茅群系和长芒草群系, 因此西北针茅群系在过去30年中的分布面积被农田和人工林所占比例更小(
1
2001
... 存在度是某个种在有多个样地记录的群落中出现于样地记录的百分数(
1
2001
... 存在度是某个种在有多个样地记录的群落中出现于样地记录的百分数(
2
2004
... 建群种西北针茅, 是多年生旱生丛生禾草(
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
2
2004
... 建群种西北针茅, 是多年生旱生丛生禾草(
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
1998
... 建群种西北针茅, 是多年生旱生丛生禾草(
1
1998
... 建群种西北针茅, 是多年生旱生丛生禾草(
2
1980
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
2
1980
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
2008
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
2008
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
2006
... 建群种西北针茅, 是多年生旱生丛生禾草(
1
2012
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
2012
... 西北针茅群系中出现的所有物种的名称及科属以《中国植物志》(
1
1987
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(
1
1987
... 西北针茅(也叫克氏针茅, Stipa sareptana var. krylovii)群系属于典型草原, 分布中心在中温带半干旱地区的蒙古高原(中国、蒙古), 在中国主要分布在呼伦贝尔高原西部、锡林郭勒高原和黄土高原地区.西北针茅群系是生态适应性最广的一个草原类型, 向东与大针茅群系分布区重叠, 甚至渗透入草甸草原区; 向西分布至荒漠草原区和荒漠区的山地, 与短花针茅群系等荒漠草原复合分布; 向南分布至黄土高原, 与长芒草群系等暖温性草原交替分布(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}