 ,1, 陈全1, 张宇杰2, 汪正祥1,3, 戴璨
,1, 陈全1, 张宇杰2, 汪正祥1,3, 戴璨 ,1,3,*
,1,3,*Effects of mating distance and number of pollen donors on sexual reproduction of Sagittaria trifolia
Pan-Pan ZHOU,1, Quan CHEN1, Yu-Jie ZHANG2, Zheng-Xiang WANG1,3, Can DAI,1,3,*通讯作者: * (daican@hubu.edu.cn). ORCID:周盼盼: 0000-0002-8326-3062
编委: 廖万金
责任编辑: 李敏
收稿日期:2020-06-18修回日期:2020-07-20网络出版日期:2020-09-20
| 基金资助: |
Received:2020-06-18Revised:2020-07-20Online:2020-09-20
| Fund supported: |

摘要
植物的有性繁殖是生活史和进化的核心, 母本及环境对繁殖的影响固然重要, 父本的贡献亦不容忽视。父本来源与多样性对坐果结籽和后代质量的影响明显, 但由于不同物种或种群的繁殖特点和进化历程, 往往也会呈现其独特性。该研究旨在探究挺水植物野慈姑(Sagittaria trifolia)是否存在自交或远交衰退, 以及父本数量对其繁殖和后代表现的影响。通过人工控制授粉, 设置自交、近距离异交(<50 km)、远距离异交(>200 km) 3种交配距离以及单、双两类父本数量, 共计5种授粉处理, 观测野慈姑坐果概率、单果种子数量、种子面积、萌发率(2018和2019年)、幼苗芽长(2018和2019年)共7项指标。结果显示: 不同距离的交配对野慈姑的坐果概率、单果种子数量、种子面积、萌发率、幼苗芽长没有显著影响, 野慈姑未出现明显的自交衰退或远交衰退现象。父本数量的增加对野慈姑的结实数量(坐果概率、单果种子数量、种子面积)无影响, 但结实质量方面, 双父本处理的种子萌发率略高于单父本处理。综上所述, 交配距离与父本数量对野慈姑的繁殖表现影响较小, 这可能与其频繁的自交历史、远距离的基因扩散以及高度相似的水生环境有关; 同时, 该研究基于20余个自然种群的基因型, 研究结果也表明了野慈姑物种水平有性繁殖的优异和稳定性。
关键词:
Abstract
Aims Sexual reproduction is the core of plant life history and evolution. Besides the influence of maternal genotype and environment, the contribution of paternal parts on reproduction should not be ignored. In particular, the distance and diversity of pollen donor have significant impacts on fruit setting and offspring quality, which often show distinctive patterns due to specific reproductive characteristics and evolutionary processes of different species or populations. Our objective was to study the effects of mating distance and number of pollen donors on reproductive performance of Sagittaria trifolia.
Methods Three mating distances including selfing, outcrossing of short distance (<50 km), and outcrossing of long distance (>200 km) and two types of pollen donors (single- and double-donor) were investigated in S. trifolia using hand-pollination. Besides selfing, the remaining factors formed a factorial design, resulting in a total of five pollination treatments. We analyzed the differences in fruiting probability, seed number per fruit, seed size, germination rate of F1 seed (year 2018 and 2019) and their seedling length (year 2018 and 2019).
Important findings Different mating distances had no significant effects on fruiting probability, seed number per fruit, seed size, germination rate or seedling length of S. trifolia, implying that there was no inbreeding or outbreeding depression in this species. The increase from single to double pollen donors had no effect on seed production of S. trifolia, but seed germination rate in double-donor treatment was slightly higher than that of single- donor. All together, mating distance and number of donors had little influence on reproductive performance of S. trifolia, which might be related to frequent selfing history, long-distance gene dispersal and high similarity in aquatic habitats of this species. Meanwhile, this study was based on mutual mating designs between genotypes from over 20 natural populations. Such results also imply high and stable reproductive output of S. trifolia at species level.
Keywords:
PDF (1167KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
周盼盼, 陈全, 张宇杰, 汪正祥, 戴璨. 交配距离与父本数量对野慈姑有性繁殖的影响. 植物生态学报, 2020, 44(9): 895-904. DOI: 10.17521/cjpe.2020.0197
ZHOU Pan-Pan, CHEN Quan, ZHANG Yu-Jie, WANG Zheng-Xiang, DAI Can.

利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(Minnaar et al., 2019)。在花粉转移的过程中, 供体与受体在空间或遗传上的远近程度往往会影响植物的繁殖表现及后代的质量(Rhode & Emmett Duffy, 2004)。其中, 关注较多的有自交衰退和远交衰退现象。近亲间的交配使等位基因的纯合性提升从而导致后代的适合度降低的现象称为自交衰退(Trame et al., 1995)。从遗传学的角度解释主要有两点: 有害隐性基因的表达(Charlesworth & Charlesworth, 1987)和失去了某些生存发育能力相关位点的杂合优势(heterozygote advantage at viability loci)(Charlesworth & Charlesworth, 1987; Ziehe & Roberds, 1989; Agren & Schemske, 1993)。相对而言, 远交衰退是指遗传上相隔较远的同一物种个体之间交配后, 后代适合度下降的现象(Waser & Price, 1994)。造成这种现象的机制可能包括杂合子劣势(heterozygote disadvantage)、共适应基因复合物的解离(breaking up of coadapted gene complexes)、等位基因间的上位效应(epistatic interactions between alleles)以及局部适应的破坏(disruption of local adaptations)等等(Price & Waser, 1979; Edmands, 2002, 2007; Hufford & Mazer, 2003; Frankham et al., 2011)。个体间的遗传距离与地理距离往往存在正相关关系(Schiemann et al., 2000; Alberto et al., 2010), 因此许多研究者通过设置不同的交配距离来观测物种的自交或远交衰退情况(Waser & Price, 1994; Trame et al., 1995; Volis et al., 2011)。
交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式。(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离。如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(Fischer & Matthies, 1997)。在花柱草属(Stylidium)植物S. hispidum里, 相对于短距离(3-10 km)的异交, 种群内的交配出现了自交衰退, 而长距离(111-124 km)的异交则表现出了远交衰退, 表明其有最适的中间异交距离(Hufford et al., 2012)。(2)存在自交衰退, 但远交衰退现象不明显, 甚至显示出杂合优势。例如, Polylepis australis的自然授粉、自交和30 km异交相比, 仅自交的种子萌发率显著降低(Seltmann et al., 2009)。Silene vulgaris自花受精产生的自交衰退现象非常普遍, 但是不同种群间的交配却显现出杂合子优势(Bailey & Mccauley, 2006)。(3)存在远交衰退, 但无近交衰退。如, 钝稃野大麦(Hordeum spontaneum)的F1代异交种子与自花授粉种子相比, 活力明显降低(Volis et al., 2011)。(4)交配距离对繁殖和子代的影响有限。例如, 路边青属(Geum)植物G. urbanum即使在很小的种群中也未出现自交衰退现象, 除了远距离(20 km)异交的萌发率稍低外, 没有发现远交对子代适合度的负面影响(Vandepitte et al., 2010)。以上每种表现在自然界中都有可能存在, 这也许体现了物种/种群间不同的繁衍历史和进化历程。频繁自花传粉的植物物种不会累积大量的有害突变(张大勇和姜新华, 2001; Vandepitte et al., 2010), 因为有害等位基因在纯合状态下会被清除, 从而缓解了以自花传粉为主的植物物种的自交衰退(Lande & Schemske, 1985; Johnston & Schoen, 1996; Byers & Waller, 1999; Crnokrak & Barrett, 2002)。然而, 也可能存在轻度有害的等位基因由于选择压力不大而无法被清除的现象(Charlesworth et al., 1990; Glémin, 2003), 因此可能会在自交亲和的种群中累积并固定下来。这种情况下, 与种群内的交配相比, 种群间的异交就有可能提高子代的适合度。可若是种群间的地理或时间隔离程度大, 导致基因流动降低, 植物适应了本地的遗传背景, 那么这些遗传分化大的种群间的交配会因为基因组的不相容性或歧化适应(disrupting adaptation)的表达而导致繁殖水平和远交后代的适合度下降(Hufford & Mazer, 2003; Edmands, 2007)。
除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果。多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(Karron et al., 2006; Koffi et al., 2013; 周庆源, 2014)。这也许是因为: (1)多父本混合授粉, 使母株获得较高兼容性花粉的机会增加, 从而提高果实中受精的胚珠数量, 减少果实或种子的败育(Paschke et al., 2002; 周庆源, 2014)。(2)与单父本的全同胞相比, 多父本的半同胞对母本资源的竞争更加激烈(Kress, 1981), 且激烈的竞争会产生更有活力的后代(Marshall et al., 2007)。但父本数量的增加并不是对所有的物种都有促进作用, 也有研究发现父本的多样性对繁殖表现没有显著影响(Sork & Schemske, 1992; Holland et al., 2009; Pélabon et al., 2015)。这可能是因为柱头上有过量的花粉应对花粉竞争和母本选择过程, 此时不管父本数量如何, 均能够保证充分结实(Bertin, 1986)。也有研究指出, 当纳入实验的多个父本之间竞争能力及其与母本个体的兼容性没有明显差异时, 繁殖水平与后代表现也不会优于单个父本的交配(Sork & Schemske, 1992; Pélabon et al., 2015, 2016)。
野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(陈家宽, 1989)。Chen等(2008)对全国范围内42个野慈姑种群的研究结果表明, 野慈姑区域间或种群间的遗传分化较低, 暗示存在长距离的基因散布和频繁的基因交流。这可能是因为野慈姑的种子非常小且漂浮于水面, 加上水系的连通流动或城镇的各类导水设施作用, 导致其散布距离长。同时, 由于水生植物物理环境的相似, 不同种群之间出现歧化选择(disruptive selection)或局部适应性(local adaptation)的可能性不大, 所以地理距离应该对基因组之间的兼容性影响较小。另外, 自然条件下野慈姑的自交率在20%左右(Dai et al., 2018a), 这种低自交频率的植物可能存在一定的遗传负荷(张大勇和姜新华, 2001); 前期也有实验发现一野慈姑自然种群的自交种子萌发率偏低(覃道凤等, 2015)。因此我们推测, 野慈姑的远交衰退现象不会太明显, 而存在自交衰退现象。本研究以多个野外种群采集的植株为材料, 设置不同的交配距离(自交、小于50 km近距离异交和大于200 km的远距离异交), 探究野慈姑的自交衰退或远交衰退程度。同时, 还试图探讨野慈姑的繁殖水平是否会随着授粉父本数量(单父本处理与双父本处理)的提升而相应提高。
1 材料和方法
1.1 实验材料与研究地点
野慈姑是多年生挺水植物, 既可以通过地下球茎进行营养繁殖, 亦可以通过蜜蜂、食蚜蝇等昆虫授粉完成有性生殖, 且自交亲和(Dai et al., 2018a, 2018b)。野慈姑雌雄同株异花, 总状花序, 每轮3朵花, 花序基部为雌花, 顶部为雄花, 由基至顶依次开放, 雌花一般在1-2天内开放完毕, 雄花持续开放4-10天, 雌雄花单花期均为1天(5:30-15:00)。在湖北省地区, 野慈姑花期为6-10月。果实在授粉后20天左右成熟, 成熟的果实易散落, 种子为褐色, 可水培。实验于2018年8月在中国科学院武汉植物园(114.42° E, 30.54° N)内野慈姑的同质园进行(罗文杰等, 2018)。同质园的植株来自于野慈姑自然种群2013年采集的球茎后代(李婷等, 2015), 各自然种群和同质园的地理位置如图1所示。
图1
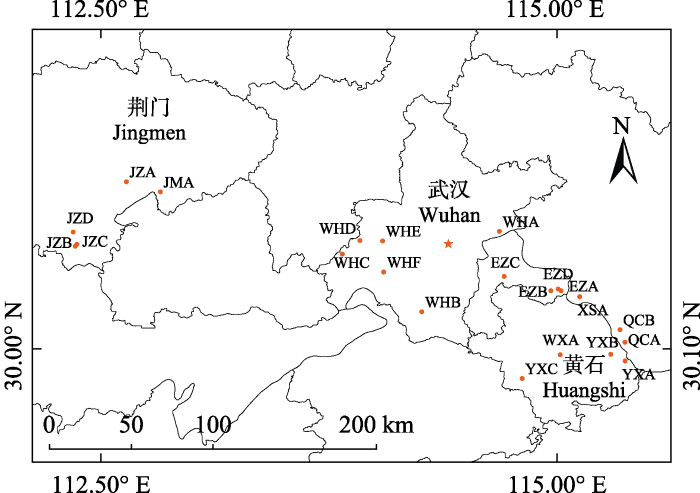 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1野慈姑种群来源。实验地为中国科学院武汉植物园。EZA-EZD, 鄂州A-D; JMA, 荆门A; JZA-JZD, 荆州A-D; QCA-QCB, 蕲春A-B。WHA-WHF, 武汉A-F; WXA, 武穴A; XSA, 浠水A; YXA-YXC, 阳新A-C; 各自然种群的详细描述见李婷等(2015)。
Fig. 1Source populations of Sagittaria trifolia. The experimental site is Wuhan Botanical Garden, Chinese Academy of Sciences. EZA-EZD, A-D of Ezhou; JMA, A of Jingmen; JZA-JZD, A-D of Jingzhou; QCA-QCB, A-B of Qichun. WHA-WHF, A-F of Wuhan; WXA, A of Wuxue; XSA, A of Xishui; YXA-YXC, A-C of Yangxin; See Li et al. (2015) for specific information of each natural population.
1.2 实验设计
1.2.1 授粉处理实验主要通过人工操控授粉的方法, 控制野慈姑雌花交配过程中花粉的来源以及父本的多样性, 以探究野慈姑是否存在自交衰退或远交衰退现象, 以及父本多样性对繁殖及后代表现产生的影响。
为探究野慈姑是否存在自交或远交衰退现象, 本实验设置自交、近距离异交和远距离异交3个水平, 拟以近距离异交的子代性状为参照, 将自交后代、远距离异交后代与其进行对比。在本实验中, 综合考虑了水系连通性和传粉昆虫的飞行能力分别影响的种子与花粉散布, 把种群之间直线距离小于50 km的交配定义为近距离异交, 大于200 km的交配定义为远距离异交。具体方法为以每一个种群为中心, 列出它与其他种群之间的距离, 把植株来源小于50 km的作为可选近距离父本, 大于200 km的作为可选远距离父本(图1)。人工授粉时, 雌雄花须满足同一天开放, 自交授粉的花粉供体来源于同一基因型的植株, 近距离异交的花粉供体优先选择来源于同一种群不同基因型的其他植株(野外最常见的交配格局)(Dai et al., 2018a), 远距离异交的花粉供体优先选择距离最远的种群。
为探究父本多样性对繁殖及后代的影响, 本实验设置单父本与双父本两个水平。人工授粉中每朵雌花均被两朵雄花授粉, 其中单父本处理的两朵雄花来源于同一个植株, 而双父本处理的两朵雄花分别来自基因型不同的两个植株。父本数量与交配距离采用了阶乘设计, 但由于自交只可能是单父本, 故共计5种授粉处理: 自交、单父本近距离异交、双父本近距离异交、单父本远距离异交以及双父本远距离异交(下文分别简称为自交、单近、双近、单远和双远)。
人工授粉以每朵雌花为重复单元, 采用雄花花药与雌花心皮直接接触的方法, 每朵雌花用两朵雄花授粉, 直至球形心皮上的花粉均匀饱和。为了控制不同基因型的母本效应, 5个处理都尽量选择在同一个花序上进行, 每种处理对应的轮次位置随机, 以避免位置效应(Dai et al., 2018a)。有少数花序内的雌花数太少, 故处理之间的样本量略有差异。在实验过程中, 为避免传粉昆虫的影响, 对即将开放以及完成人工授粉的花序进行套袋处理。在人工授粉后的第二天摘除套袋, 以免影响后续果实的发育。授粉操作于2018年8月盛花期进行, 每天8:00-12:00进行人工授粉, 最终样本量为自交51, 单近53, 双近52, 单远50, 双远54。
1.2.2 繁殖水平
不同授粉处理下野慈姑的繁殖水平用授粉雌花的坐果概率、单果种子数量和种子面积(该植物扁平状种子的平铺面积, 种子大小的指标)为评价指标。在授粉操作10天后, 用透气透光的网袋对已发育的果实进行套袋, 防止果实成熟后散落。从授粉后20天开始, 对完全成熟的果实(果实从紧实变为疏松、种子呈准散落状态)进行采集, 单个存放于6 cm × 10 cm的牛皮纸袋中, 并统计坐果情况(坐果成功记为1, 失败记为0), 每日检查, 直至所有果实采集完成。果实袋带回实验室后立即对种子指标进行统计。具体操作为: 将每个果实的所有种子人工平铺于一张A3白纸上, 尽量保证种子之间没有重合, 用固定位置和焦距的单反照相机(尼康D7000)对其拍照, 然后用ImageJ软件对照片进行颗粒计数与大小估测, 得出单果种子数量与种子的平均面积。为了转化种子的面积单位, 边长为1 cm的黑色正方形标块被放置在白纸上和种子一起拍摄, 通过求得该正方形的平均像素面积, 将种子面积单位全部转化为mm2。
1.2.3 萌发实验
于2018和2019年分别进行两次种子萌发实验。2018年8-9月, 采集回来的果实统计完种子数量后, 立即将种子放入水中进行萌发(模拟自然状态)。每个果实随机挑取100粒种子, 不足100粒的则全部选取, 剩余的种子放回原袋进行烘干(50 ℃, 12 h), 以备后续使用。将每个果实待萌发的种子放入装满自来水的塑料杯中, 在MGC-350HP-2人工气候箱(上海一恒科学仪器有限公司, 上海)中进行培养。本实验设定的人工气候箱的环境温度为30 ℃, 相对湿度20%, 光照时间每天18 h, 光照强度选择仪器最大值20 000 lx。培养10天后, 种子萌发基本趋于顶峰(大于10天的培养新增萌发较少, 且杯内易滋生藻类等低等植物), 记录发芽个数, 并测量最长的幼苗芽长。由于果实数量较多, 所有种子陆续被随机分配到了3个温箱进行培养。
2019年10月对剩余的种子进行了第二次萌发实验。由于这批种子进行了烘干处理, 因此在这次萌发之前对种子进行了两个月的浸泡(覃道凤等, 2015), 其余的萌发操作和条件均与第一次萌发实验相同。
1.3 统计分析
本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年)。除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布。单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据。全部统计用R软件(v.3.6.1, Team, 2016)分析, 使用package包括car (Fox & Weisberg, 2019), lme4 (Bates et al., 2015), lmerTest (Kuznetsova et al., 2017), lsmeans (Lenth, 2016), psych (Revelle, 2018)。为了探究野慈姑是否存在自交衰退或远交衰退, 将3种单父本的处理(自交、单近、单远)组成子数据集对7个变量进行方差分析。对坐果概率, 采用因变量二项分布的广义线性模型分析交配距离对坐果的影响, 处理类型作为固定因素, 实验日期作为随机因素。对其他6个变量, 均采用混合线性模型分析交配距离对各繁殖指标的影响, 处理类型作为固定因素, 实验日期作为随机因素, 其中2018年的萌发率与幼苗芽长加入了温箱作为随机因素。
为了探究双父本情况下近距离与远距离交配的区别, 将双近与双远两种处理组成子数据集分析, 模型同上。
最后, 为了整体研究父本数量对繁殖的影响, 将单近、单远、双近、双远4种处理集合, 以“父本数量”作为固定因素, 模型内其他要素同上, 对每个变量进行了分析。
以上3个方面的比较虽涉及重复使用数据, 但两两之间均为正交关系, 故不需要对显著性水平进行校正, 依旧采用α = 0.05。所有线性分析中的方差同质性均检测且满足。此外, 考虑到不同的萌发方法, 还对两个年份的萌发率和幼苗芽长进行了成对t检测。结果中全部使用原始数据(算术平均数±标准误差)。
2 结果
2.1 不同交配距离对野慈姑繁殖的影响
单父本情形下, 将自交、近距离异交、远距离异交的后代繁殖指标进行比较后发现, 3种处理下的各项繁殖指标均无差异(表1), 暗示野慈姑并无明显的自交衰退或远交衰退现象。Table 1
表1
表1野慈姑不同交配距离下的繁殖指标比较(平均值±标准误差)
Table 1
| 繁殖指标 Reproductive indices | 自交 Selfing | 近距离异交 Outcrossing of short distance | 远距离异交 Outcrossing of long distance | 比较 Comparison |
|---|---|---|---|---|
| 坐果概率 Fruiting probability (%) | 90.38 ± 4.13 | 88.46 ± 4.47 | 90.00 ± 4.29 | LRT = 0.136, p = 0.93 |
| 单果种子数 Seed number per fruit | 236.96 ± 21.78 | 237.49 ± 25.02 | 253.98 ± 24.00 | F2,125.0 = 0.22, p = 0.81 |
| 种子面积 Seed size (mm2) | 5.50 ± 0.20 | 5.90 ± 0.24 | 5.68 ± 0.20 | F2,123.8 = 0.84, p = 0.44 |
| 2018年萌发率 Germination rate of year 2018 (%) | 5.82 ± 0.95 | 4.64 ± 1.05 | 4.47 ± 0.70 | F2,126.0 = 0.07, p = 0.93 |
| 2019年萌发率 Germination rate of year 2019 (%) | 8.48 ± 1.38 | 7.62 ± 1.23 | 9.16 ± 1.63 | F2,104.0 = 0.14, p = 0.87 |
| 2018年幼苗芽长 Seedling length of year 2018 (cm) | 3.24 ± 0.28 | 3.09 ± 0.21 | 3.36 ± 0.24 | F2,93.3 = 0.67, p = 0.52 |
| 2019年幼苗芽长 Seedling length of year 2019 (cm) | 2.13 ± 0.17 | 2.33 ± 0.20 | 2.23 ± 0.20 | F2,85.0 = 0.26, p = 0.77 |
新窗口打开|下载CSV
然而, 双父本处理下, 远距离的异交较近距离的异交显著提升了坐果概率(似然比检验(LRT) = 5.63, p = 0.018; 图2A)。不过, 两种交配距离的单果种子数(F1,87.0 = 0.004, p = 0.95; 图2B)、种子面积(F1,87.2 = 0.58, p = 0.45; 图2C)、萌发率(2018年: F1,85.7 = 0.78, p = 0.38; 2019年: F1,69.3 = 0.30, p = 0.59; 图3A、3B)以及幼苗芽长(2018年: F1,80.0 = 0.25, p = 0.62; 2019年: F1,48.3 = 0.21, p = 0.65; 图3C、3D)都未呈现显著区别。
Fig. 2
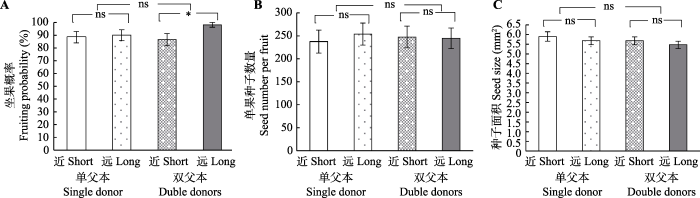 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Fruiting status of Sagittaria trifolia under different mating treatments (mean ± SE). Short, outcrossing of short distance; Long, outcrossing of long distance. ns, not significant (p > 0.05); *, p < 0.05.
2.2 不同父本数量对野慈姑繁殖的影响
单父本与双父本授粉条件下, 野慈姑的结实情况无明显差异(坐果概率: LRT = 0.69, p = 0.4; 单果种子数: F1,175.94 = 0.05, p = 0.82; 种子面积: F1,174.73 = 1.13, p = 0.29; 图2)。但在种子萌发方面, 单双父本2018年的萌发率差异接近显著(F1,172.52 = 3.30, p = 0.07; 图3A), 双父本交配后代的萌发率略高; 但2019年的第二次萌发并未呈现显著区别(F1,138 = 0.33, p = 0.57; 图3B)。在幼苗芽长方面, 单双父本2018年与2019年的幼苗芽长均无显著区别(2018年: F1,149.26 = 0.98, p = 0.32; 2019年: F1,105.4 = 0.94, p = 0.34; 图3C、3D), 但两年中双父本交配后代的幼苗芽长比单父本交配后代的幼苗芽长略长。以上结果表明父本数量在本实验中未影响野慈姑的结实数量, 但在后代质量上, 双父本交配的萌发率略有优势。Fig. 3
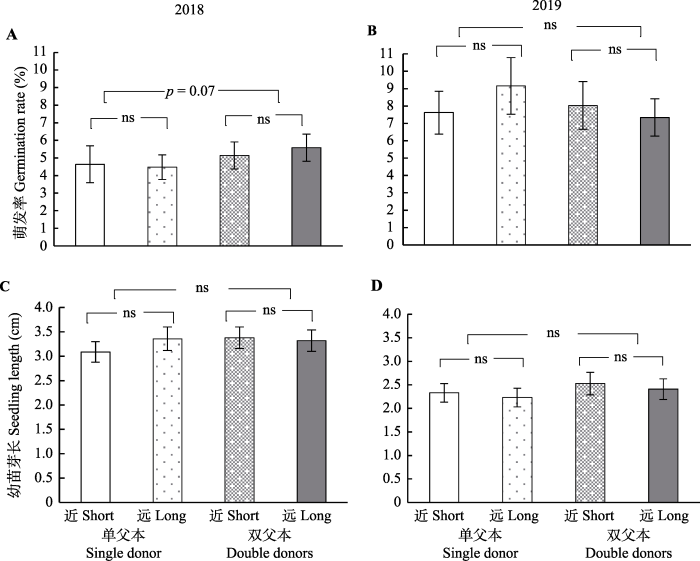 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Seed germination status of Sagittaria trifolia under different mating treatments (mean ± SE). Short, outcrossing of short distance; Long, outcrossing of long distance. ns, not significant (p > 0.05).
2.3 不同萌发方法的结果比较
本实验中两次萌发的区别体现在, 2018年将采集回来的鲜种子直接萌发, 而2019年将烘干过且储存了一年的种子浸泡两个月后再进行萌发。对比发现, 2019年的种子萌发率有显著提升(成对t检测t = 6.28, df = 189, p < 0.001; 图3A、3B), 但是2019年的幼苗芽长却显著下降(t = 7.06, df = 126, p < 0.001; 图3C、3D)。3 讨论
研究结果发现, 野慈姑没有明显的自交衰退现象(表1), 这不符合前期的实验猜想。一般认为, 中低水平自交的植物存在一定的遗传负荷(张大勇和姜新华, 2001), 研究指出野慈姑的自交率为20.9%- 22.7% (Dai et al., 2018b), 属于以异交为主的混合交配系统, 但经过人工自交后, 其繁殖表现和后代质量并未出现下降。类似的表现也出现在以异交为主的紫萼路边青(Geum rivale)的研究中(Ruhsam et al., 2010)。相反, 一些自交率很高的植物, 如秋海棠属(Begonia)植物B. hirsuta和B. semiovata、苦参属(Sophora)植物S. microphylla、毒子榄属(Baillonella)植物B. toxisperma等, 却又存在严重的自交衰退现象(Agren & Schemske et al., 1993; van Etten et al., 2015; Duminil et al., 2016)。由此可见, 不能单凭物种现阶段的自交率来判定是否会出现自交衰退现象。因为自交衰退程度会因物种、环境等多方面的差异而有所不同(王峥峰, 2003), 即便是同一物种, 也会因不同种群的发展阶段和长期交配格局而改变(Husband & Schemske, 1997; Kittelson & Maron, 2000; Hull- Sanders et al., 2005)。本实验中野慈姑未表现出自交衰退现象, 但前期一个自然种群的自交实验导致种子萌发下降(覃道凤等, 2015), 这可能与单一自然种群的建群来源及其交配历史有关。本实验的野慈姑种群来源广泛, 那些具有频繁自交历史(或来自频繁自交种群)的基因型在长期的交配中有害突变得以清除, 自交后代的适合度较高, 有助于平衡某些种群的自交劣势, 使得平均的繁殖水平保持稳定。此外, 与条件恶劣的自然环境比, 同质园的环境条件相对稳定, 这些优良的环境条件可能在一定程度上降低了自交衰退的表达(何亚平和刘建全, 2003; Armbruster & Reed, 2005)。另一方面, 野慈姑也未表现出远交衰退(表1), 这与前期的预测相符。野慈姑的长距离基因散布与克隆生长, 降低了空间分离种群的遗传分化, 同时水生环境的相似性使得不同种群间的歧化选择与局部适应不明显。另外野慈姑这一物种在第四纪冰川撤退后不断向北延伸, 分布范围不仅仅局限于单一的气候区域(Santamaría, 2002), 所以本实验200 km的距离相对野慈姑广泛的分布范围来讲可能尺度并不太大, 导致远交衰退现象不明显。值得注意的是, 双父本处理下, 远交的坐果概率较近交有显著提升(图2A)。可能是因为双父本提供了更多机会使得雌性配子与更匹配的雄性配子结合(Marshall, 1988); 或少数个体的花粉质量特别突出(Lankinen et al., 2009), 在双父本处理中由于使用次数加倍, 其优异性得以展现。一些研究提到, 杂合优势的程度与亲本间的遗传差异成正比(East, 1936), 也就是说: 杂合优势的水平随亲本遗传距离的增加而增加(Kaeppler, 2012; Marcón et al., 2019)。而域间遗传分化较低的野慈姑(Chen et al., 2008)是否存在杂合优势或是远交衰退, 还需要延长实验观测时间(Waser & Price, 1994; Quilichini et al., 2001)或增加亲本间的交配距离, 可能才会有更显著的表现。
本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(图2, 图3)。这可能是因为: (1)种子质量及数量多受低至中等程度的花粉量的影响(Hildesheim et al., 2019), 且只有在花粉量低的情况下, 多父本的种子质量提升才会凸显(Bertin, 1986)。而本实验以两朵雄花的所有花粉为雌花授粉, 花粉量比较充分饱和, 不论是单父本还是双父本都能保证充分优质的结实。(2)不同父本产生的果实争夺资源的能力相差无几, 或母本植株没有选择性的偏好, 致使多父本混合授粉的后代表现与单父本后代表现相似。比如, 在给山扁豆属(Chamaecrista)植物C. fasciculata授多个父本的混合花粉后, 其坐果率、果实与种子产量、种子重量和萌发率都没有显著提升(Sork & Schemske, 1992)。父本数量的增加对黄蓉花属(Dalechampia)植物D. scandens的种子质量也无显著影响(Pélabon et al., 2015)。本研究中, 2018年双父本处理的萌发率比单父本的略高(图3A), 暗示父本数量的增多在一定程度上提升了种子质量, 但该趋势在第二次萌发中并未展现。也许添加更多的父本数量才能有助于发挥竞争的筛选作用, 从而充分显示其对繁殖和子代质量的积极影响(Marshall & Ellstrand, 1986; Paschke et al., 2002; Karron et al., 2006; 赵学杰和谭敦炎, 2007; Koffi et al., 2013)。
不论交配距离或父本数量如何改变, 野慈姑种子的萌发指标都没有表现出显著变化(表1; 图3), 说明其种子和子代幼苗质量的稳定性, 暗示野慈姑的繁殖策略会优先满足单个种子的养分供给和生命活力, 当条件发生变化时通过调整果实或种子的数量进行响应(Dai et al., 2018b)。这一结果也提示今后类似的实验应重点关注野慈姑坐果与结实数量的变异而非质量。不过, 我们发现不同的萌发处理对野慈姑种子的萌发效果有显著影响: 与2018年第一次萌发处理相比, 2019年的萌发率显著提升, 而幼苗芽长却显著下降(图3)。本实验的萌发处理在人工气候箱内完成, 两次萌发设置的条件相同, 唯一不同的是种子的新鲜程度与烘干处理: 2018年将鲜种子直接浸水萌发, 以模拟自然状态; 剩下的种子烘干且储存了一年, 于2019年浸泡吸水2个月后萌发。2019年的萌发率提升说明烘干和一年的存储期对野慈姑的种子活性没有明显的负面影响, 适当时间的浸泡或许对萌发有益(杨忠仁等, 2007; 席沁等, 2014)。至于幼苗芽长下降, 可能是因为2019年的种子全部集中放置于一个气候箱内萌发, 由于密度太大, 幼苗光照强度不足所致。
综上所述, 以异交为主的野慈姑经过人工自交后, 其适合度和F1表现并未降低, 200 km距离的种群间交配也未发现远交衰退现象, 频繁的自交历史、远距离的基因扩散和相似的物理生境可能扮演了至关重要的作用。不过, 鉴于野慈姑广泛的分布范围, 建议在类似物种的研究中, 适当拓宽交配距离或延长观测周期, 以深入探讨交配距离对繁殖的影响。此外, 本研究基于野慈姑20余个自然种群的基因型, 相较于单一种群的研究, 实验样本具有更高的基因多样性。从实验设计上, 每个种群的基因型都有均等的机会作为父本或母本参与交配, 避免了以单一或少量种群作为母本交配的遗传背景影响(Vandepitte et al., 2010)或母本效应(Galloway & Etterson, 2007), 因此, 本论文的结果从物种水平上揭示了野慈姑有性繁殖的优异和稳定性, 更具说服力。
致谢 感谢湖北大学资源环境学院邓琳、牟亚莉在实验过程中的帮助, 丁新颖、牛可可、解季明、解雪婷对文章修改给予的建议。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1558-5646.1993.tb01204.xURLPMID:28568096 [本文引用: 2]

DOI:10.1890/09-0050.1URLPMID:20380195 [本文引用: 1]
Isolation by distance (IBD) models are widely used to predict levels of genetic connectivity as a function of Euclidean distance, and although recent studies have used GIS-landscape ecological approaches to improve the predictability of spatial genetic structure, few if any have addressed the effect of habitat continuity on gene flow. Landscape effects on genetic connectivity are even less understood in marine populations, where habitat mapping is particularly challenging. In this study, we model spatial genetic structure of a habitat-structuring species, the giant kelp Macrocystis pyrifera, using highly variable microsatellite markers. GIS mapping was used to characterize habitat continuity and distance between sampling sites along the mainland coast of the Santa Barbara Channel, and their roles as predictors of genetic differentiation were evaluated. Mean dispersal distance (sigma) and effective population size (Ne) were estimated by comparing our IBD slope with those from simulations incorporating habitat continuity and spore dispersal characteristics of the study area. We found an allelic richness of 7-50 alleles/locus, which to our knowledge is the highest reported for macroalgae. The best regression model relating genetic distance to habitat variables included both geographic distance and habitat continuity, which were respectively, positively and negatively related to genetic distance. Our results provide strong support for a dependence of gene flow on both distance and habitat continuity and elucidate the combination of Ne and a that explained genetic differentiation.
DOI:10.1038/sj.hdy.6800721URLPMID:16077737 [本文引用: 1]
Understanding the consequences of inbreeding has important implications for a wide variety of topics in population biology. Although it is often stated in the literature that the deleterious effects of inbreeding (inbreeding depression) are expected to be more pronounced under stressful than benign conditions, this issue remains unresolved and controversial. We review the current literature on the relationship between the magnitude of inbreeding depression and environmental stress and calculate haploid lethal equivalents expressed under relatively benign and stressful conditions based on data from 34 studies. Inbreeding depression increases under stress in 76% of cases, although this increase is only significant in 48% of the studies considered. Estimates of lethal equivalents are significantly greater under stressful (mean = 1.45, median = 1.02) than relatively benign (mean = 0.85, median = 0.61) conditions. This amounts to an approximately 69% increase in inbreeding depression in a stressful vs a benign environment. However, we find strong lineage effects to be ubiquitous among studies that examine inbreeding depression in multiple environments, and a prevalence of conditionally expressed deleterious effects within lineages that are uncorrelated across environments. These results have important implications for both evolutionary and conservation biology.
DOI:10.1111/jec.2006.94.issue-1URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00377104URLPMID:28311280 [本文引用: 2]
I examined the effects of pollen loads containing pollen from one, three and five donors on fruit production and fruit quality in Campsis radicans. Number of pollen donors had no significant effect on % fruit production, seed number, seed weight or seed germination. In singledonor pollinations the identity of the donor did have a strong effect on the above parameters. Furthermore, the best single donor sired fruits with more seeds and heavier seeds than any mixture containing this donor. This pattern indicates interference of pollens or preemption of some ovules by the inferior pollen. In Campsis, therefore, the number of pollen donors contributing to a pollen load is less important than the identity of these donors in determining fruit production and fruit quality. Seeds from fruits resulting from mixed pollination were slightly more variable than seeds from fruits resulting from single-donor pollinations.
[本文引用: 1]
DOI:10.1038/347380a0URL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ympev.2008.03.008URL [本文引用: 1]
AbstractSagittaria trifolia L. is a perennial, erect herb that is confined to ponds, rice fields, ditches, and freshwater wetlands. Using chloroplast DNA (cpDNA) atpB–rbcL intergenic spacer sequences, we studied the phylogeographic pattern and demographic history of S. trifolia with 108 samples from 42 populations representing the entire geographic range in China. Twenty-seven haplotypes were characterized and two of them were widely distributed in the populations. In the minimum-spanning network, all tip haplotypes were unique to a particular population, while the interior nodes represented widespread haplotypes. Nested clade analysis (NCA) of cpDNA haplotypes indicated that long distance dispersal characterized the post-glacial recolonization of S. trifolia in China. No specific refugia areas were suggested because genetic differentiation was low among the sampled regions and among populations within regions although a large number of the haplotypes were unique to a single population. The present data support that the unique haplotypes in individual population most likely represent recent mutational derivatives after long distance dispersal rather than the relics in refugia. These results for S. trifolia represent the first phylogeographic analysis of a widespread marsh herb in China and support the importance of long distance dispersal events in the post-glacial migrations of plants.]]>
DOI:10.1111/j.0014-3820.2002.tb00160.xURLPMID:12583575 [本文引用: 1]
Inbreeding depression, the reduction in fitness that accompanies inbreeding, is one of the most important topics of research in evolutionary and conservation genetics. In the recent literature, much attention has been paid to the possibility of purging the genetic load. If inbreeding depression is due to deleterious alleles, whose effect on fitness are negative when in a homozygous state, then successive generations of inbreeding may result in a rebound in fitness due to the selective decrease in frequency of deleterious alleles. Here we examine the experimental evidence for purging of the genetic load by collating empirical tests of rebounds in fitness-related traits with inbreeding in animals and plants. We gathered data from 28 studies including five mammal, three insect, one mollusc, and 13 plant species. We tested for purging by examining three measures of fitness-component variation with serial generations of inbreeding: (1) changes in inbreeding depression, (2) changes in fitness components of inbred lines relative to the original outbred line, and (3) purged population (outcrossed inbred lines) trait means as a function of ancestral outbred trait means. Frequent and substantial purging was found using all three measures, but was particularly pronounced when tracking changes in inbreeding depression. Despite this, we found little correspondence between the three measures of purging within individual studies, indicating that the manner in which a researcher chooses to estimate purging will affect interpretation of the results obtained. The discrepancy suggests an alternative hypothesis: rebounds in fitness with inbreeding may have resulted from adaptation to laboratory conditions and not to purging when using outcrossed inbred lines. However, the pronounced reduction in inbreeding depression for a number of studies provides evidence for purging, as the measure is likely less affected by selection for laboratory conditions. Unlike other taxon-specific reviews on this topic, our results provide support for the purging hypothesis, but firm predictions about the situations in which purging is likely or the magnitude of fitness rebound possible when populations are inbred remain difficult. Further research is required to resolve the discrepancy between the results obtained using different experimental approaches.
DOI:10.1111/psbi.2018.33.issue-4URL [本文引用: 5]
DOI:10.1002/ajb2.1053URLPMID:29710426 [本文引用: 3]
PREMISE OF THE STUDY: Understanding resource allocation to reproduction, a key factor in life history tradeoffs, has long intrigued plant ecologists. Despite the recognized importance of understanding the movement of resources among flowers following variable pollination, the patterns of resource reallocation to plant reproductive organs have not been thoroughly addressed. In this study, we aimed to empirically explore how resources redistribute within inflorescences in response to differential pollination intensities. METHODS: Using a common herb, Sagittaria trifolia, we conducted supplemental and controlled pollination for single, some, or all flowers in simple and complex inflorescences, and compared their resulting fruiting probabilities, seed production, and average seed masses. KEY RESULTS: Pollen supplementation of a single flower significantly increased its fruiting probability; however, the same manipulation of an inflorescence did not increase its overall reproduction. Single pollen-supplemented flowers had a higher percentage fruit set than inflorescences receiving supplemental pollination. In complex inflorescences, supplemental pollination had no effect on the reproductive success of flowers on the lateral or main branches. CONCLUSIONS: We provided evidence of resource reallocation from controlled to pollen-supplemented flowers in simple inflorescences; however, resources were unlikely to be reallocated between the main and lateral branches in the complex inflorescences, suggesting that flowering branches represent integrated physiological units in S. trifolia. The results also demonstrated that single-flower supplemental pollination would exaggerate pollen limitation and lead to a biased understanding of a plant's reproductive status.
DOI:10.1016/j.foreco.2016.08.003URL
URLPMID:17246801 [本文引用: 1]
DOI:10.1016/S0169-5347(02)02585-5URL [本文引用: 1]
DOI:10.1111/j.1365-294X.2006.03148.xURLPMID:17257106 [本文引用: 1]
As populations become increasingly fragmented, managers are often faced with the dilemma that intentional hybridization might save a population from inbreeding depression but it might also induce outbreeding depression. While empirical evidence for inbreeding depression is vastly greater than that for outbreeding depression, the available data suggest that risks of outbreeding, particularly in the second generation, are on par with the risks of inbreeding. Predicting the relative risks in any particular situation is complicated by variation among taxa, characters being measured, level of divergence between hybridizing populations, mating history, environmental conditions and the potential for inbreeding and outbreeding effects to be occurring simultaneously. Further work on consequences of interpopulation hybridization is sorely needed with particular emphasis on the taxonomic scope, the duration of fitness problems and the joint effects of inbreeding and outbreeding. Meanwhile, managers can minimize the risks of both inbreeding and outbreeding by using intentional hybridization only for populations clearly suffering from inbreeding depression, maximizing the genetic and adaptive similarity between populations, and testing the effects of hybridization for at least two generations whenever possible.
URLPMID:21708572 [本文引用: 2]
Isolation and small size of populations as a result of habitat destruction and fragmentation may negatively affect plant fitness through pollinator limitation and increased levels of inbreeding. To increase genetic variation in small populations of rare plants artificial gene flow has been suggested as a management tool. We investigated whether pollinator limitation and inbreeding depression could reduce fitness in Gentianella germanica, an endangered biennial of increasingly fragmented calcareous grasslands in Central Europe. We experimentally excluded pollinators and generated progenies by hand-pollinating flowers with pollen from different distances. G. germanica was highly selfing. Pollinator exclusion strongly reduced seed set, indicating that pollinator limitation could potentially reduce plant fitness. Germination rate as well as number of leaves and rosette size of progeny from 10-m crosses was higher than that of progeny from open pollinations, self-, 1-m, and interpopulation crosses. After 6 mo of growth differences in the number of surviving plants persisted, whereas differences in plant size did not. The results suggest that inbreeding depression may reduce plant performance in G. germanica. Outbreeding depression in the performance of progeny from interpopulation crosses indicates that caution is necessary in using artificial interpopulation gene flow as a management tool.
[本文引用: 1]
DOI:10.1111/j.1523-1739.2011.01662.xURL [本文引用: 1]
DOI:10.1126/science.1148766URLPMID:18006745 [本文引用: 1]
Plants exhibit adaptive responses to light, but it is not known whether parental plants transmit environmental cues that elicit adaptive responses in offspring. We show that offspring life history (annual versus biennial) is influenced by the maternal light environment (understory versus light gap). This transgenerational plasticity is adaptive when offspring are grown in their maternal light environment, where seeds typically disperse. Projections of population growth show that plants that are appropriately cued for their light environment through maternal effects have 3.4 times greater fitness than otherwise. Transgenerational plasticity has evolved in response to natural variation in light and provides a flexible mechanism by which sedentary organisms cope with heterogeneous environments.
DOI:10.1111/j.0014-3820.2003.tb01512.xURLPMID:14761049 [本文引用: 1]
Accumulation of deleterious mutations has important consequences for the evolution of mating systems and the persistence of small populations. It is well established that consanguineous mating can purge a part of the mutation load and that lethal mutations can also be purged in small populations. However, the efficiency of purging in natural populations, due to either consanguineous mating or to reduced population size, has been questioned. Consequences of consanguineous mating systems and small population size are often equated under
[本文引用: 1]
URL [本文引用: 1]
植物繁育系统是当今进化生物学研究中最为活跃的领域。繁育系统是指代表所有影响后代遗传组成的有性特征的总和,主要包括花形态特征、花的开放式样、花各部位的寿命、传粉者种类和频率、自交亲和程度和交配系统,其中交配系统是核心。我们重点综述了植物有性繁育系统研究中1)传粉模式的多样性,2)从历史发生角度利用系统发育方法检验花性状的演化和繁育系统的生态转变过程,3)近交衰退对植物生殖史的影响及其机制和4)混和交配系统的时空动态、维持机理以及进化趋势的最新研究结果。总结了繁育系统在研究花适应性、物种形成机制和濒危植物
DOI:10.1002/ece3.5858URLPMID:31938516
Greater pollination intensity can enhance maternal plant fitness by increasing seed set and seed quality as a result of more intense pollen competition or enhanced genetic sampling. We tested experimentally these effects by varying the pollen load from a single pollen donor on stigmas of female flowers of Dalechampia scandens (Euphorbiaceae) and measuring the effects on seed number and seed mass. Seed set increased rapidly with pollen number at low to moderate pollen loads, and a maximum set of three seeds occurred with a mean pollen load of 19 pollen grains. We did not detect a trade-off between the number of seeds and seed mass within a fruit. Seed mass increased with increasing pollen load, supporting the hypothesis of enhanced seed quality via increased pollen-competition intensity or genetic sampling. These results suggest that maternal fitness increases with larger pollen loads, even when the fertilization success is already high. Our results further highlight the importance of high rates of pollen arrival onto stigmas, as mediated by reliable pollinators. Comparing the pollen-to-seed response curve obtained in this experiment with those observed in natural populations suggests that pollen limitation may be more severe in natural populations than predicted from greenhouse studies. These results also indicate that declines in pollinator abundance may decrease plant fitness through lowered seed quality before an effect on seed set is detected.
DOI:10.1086/597266URL [本文引用: 1]
DOI:10.1002/ece3.302URLPMID:23139884 [本文引用: 1]
The benefits of composite rather than local seed provenances for ecological restoration have recently been argued, largely on the basis of maximizing evolutionary potential. However, these arguments have downplayed the potentially negative consequences of outbreeding depression once mixed provenances interbreed. In this study, we compared intraspecific F1 hybrid performance and molecular marker differentiation among four populations of Stylidium hispidum, a species endemic to Southwestern Australia. Multivariate ordination of 134 AFLP markers analyzed genetic structure and detected two clusters of paired sites that diverged significantly for marker variation along a latitudinal boundary. To test for outbreeding depression and to determine the consequences of molecular population divergence for hybrid fitness, we conducted controlled pollinations and studied germination and survival for three cross categories (within-population crosses, short- and long-distance F1 hybrids) for paired sites distributed within and between the two genetically differentiated regions. We found evidence of outbreeding depression in long-distance hybrids (111-124 km), and inbreeding depression among progeny of within-population crosses, relative to short-distance (3-10 km) hybrids, suggesting an intermediate optimal outcrossing distance in this species. These results are discussed in light of the evolutionary consequences of mixing seed sources for biodiversity restoration.
DOI:10.1016/S0169-5347(03)00002-8URL [本文引用: 1]
DOI:10.3732/ajb.92.11.1871URLPMID:21646104 [本文引用: 1]
Inbreeding depression and selfing rate were investigated in the self-compatible vine Ipomoea hederacea to assess the variability of the breeding system. Inbreeding depression differed between populations and the magnitude varied at germination, growth (as measured by aboveground biomass), and reproductive potential. Plants from Macon County, Alabama, USA, had significant inbreeding depression (31%) at germination, but no significant inbreeding depression for aboveground biomass or number of reproductive structures (buds and flowers) at 45 d post germination in the greenhouse or in the field. Plants from Morgan County, Alabama, however, had significant inbreeding depression (>50%) for all three stages in the greenhouse. In allozyme comparisons, five of the 11 I. hederacea populations surveyed had high selfing rates (66.66-92.53%) and high levels of homozygosity (F(IS) = 0.500-0.861) in 2003, and three of four populations surveyed in 2004 had selfing rates that exceeded 50%. High selfing rates, high levels of homozygosity, and low levels of inbreeding depression suggest that inbreeding depression may not present a significant barrier to the transmission of selfing alleles in some populations of I. hederacea, but does not account for the maintenance of a mixed mating system in other populations.
DOI:10.1111/j.1558-5646.1997.tb03657.xURLPMID:28568601 [本文引用: 2]
The partial dominance model for the evolution of inbreeding depression predicts that tetraploids should exhibit less inbreeding depression than their diploid progenitors. We tested this prediction by comparing the magnitude of inbreeding depression in tetraploid and diploid populations of the herbaceous perennial Epilobium angustifolium (Onagraceae). Inbreeding depression was estimated in the greenhouse for three tetraploid and two diploid populations at four life stages. The mating system of a tetraploid population was estimated and compared to a previous estimate for diploids. Tetraploids showed less inbreeding depression than diploids at all life history stages, and these differences were significant for seed-set and cumulative fitness, but not for germination, survival, or plant dry mass at nine weeks. This result suggests that the genetic basis of inbreeding depression may differ among life stages. The primary selfing rate of the tetraploid population was r = 0.43, which is nearly identical to that of a diploid population (r = 0.45), indicating that differences in inbreeding depression between diploids and tetraploids are probably not due to differences in the mating system. Cumulative inbreeding depression, calculated from the four life history stages, was significantly higher for diploids (x-=0.95) than for tetraploids (x-=0.67), supporting the partial dominance model of inbreeding depression.
DOI:10.1111/j.1558-5646.1996.tb03921.xURLPMID:28565711 [本文引用: 1]
The relation between inbreeding depression and rate of self-fertilization was studied in nine natural populations of the annual genus Amsinckia. The study included two clades (phylogenetic lineages) in which small-flowered, homostylous populations or species are believed to have evolved from large-flowered, heterostylous, self-compatible ones. In one lineage the small-flowered species is tetraploid with disomic inheritance. Rates of self-fertilization were 25% to 55% in the four large-flowered, heterostylous populations; 72% in a large-flowered but homostylous population; and greater than 99.5% in the four small-flowered, homostylous populations, which produce seed autonomously. When present, inbreeding depression occurred in the fertility but not the survival components of fitness. Using a cumulative fitness measure incorporating both survival and fertility (flower number), we found inbreeding depression to be lower in the four very highly self-fertilizing populations than in the five intermediate ones. The Spearman rank correlation between inbreeding depression and selfing rate for the nine populations was -0.50, but was not statistically significant (P = 0.12). Inbreeding depression was greater in the two tetraploid populations than in the very highly self-fertilizing, diploid ones. Phenotypic stability of progeny from self-fertilization tended to be higher in populations with lower inbreeding depression. We conclude that levels of self-fertilization and inbreeding depression in Amsinckia are determined more by other factors than by each other. Estimates of mutation rates and dominance coefficients of deleterious alleles, obtained from a companion study of the four highly self-fertilizing populations, suggest that a strong relationship may not be expected. We discuss the relationship of the present results to current theory of the coevolution of self-fertilization and inbreeding depression.
[本文引用: 1]
DOI:10.3732/ajb.93.9.1306URLPMID:21642195 [本文引用: 1]
URLPMID:10811789 [本文引用: 1]
Little is known about the breeding systems of perennial Lupinus species. We provide information about the breeding system of the perennial yellow bush lupine, Lupinus arboreus, specifically determining self-compatibility, outcrossing rate, and level of inbreeding depression. Flowers are self-compatible, but autonomous self-fertilization rarely occurs; thus selfed seed are a product of facilitated selfing. Based on four isozyme loci from 34 maternal progeny arrays of seeds we estimated an outcrossing rate of 0.78. However, when we accounted for differential maturation of selfed seeds, the outcrossing rate at fertilization was lower, approximately 0.64. Fitness and inbreeding depression of 11 selfed and outcrossed families were measured at four stages: seed maturation, seedling emergence, seedling survivorship, and growth at 12 wk. Cumulative inbreeding depression across all four life stages averaged 0.59, although variation existed between families for the magnitude of inbreeding depression. Inbreeding depression was not manifest uniformly across all four life stages. Outcrossed flowers produced twice as many seeds as selfed flowers, but the mean performance of selfed and outcrossed progeny was not different for emergence, seedling survivorship, and size at 12 wk. Counter to assumptions about this species, L. arboreus is both self-compatible and outcrosses approximately 78% of the time.
DOI:10.1016/j.scienta.2013.10.007URL [本文引用: 2]
DOI:10.2307/2418541URL [本文引用: 1]
[本文引用: 2]
DOI:10.1111/j.1558-5646.1985.tb04077.xURLPMID:28563655 [本文引用: 1]
The amounts of inbreeding depression upon selfing and of heterosis upon outcrossing determine the strength of selection on the selfing rate in a population when this evolves polygenically by small steps. Genetic models are constructed which allow inbreeding depression to change with the mean selfing rate in a population by incorporating both mutation to recessive and partially dominant lethal and sublethal alleles at many loci and mutation in quantitative characters under stabilizing selection. The models help to explain observations of high inbreeding depression (> 50%) upon selfing in primarily outcrossing populations, as well as considerable heterosis upon outcrossing in primarily selfing populations. Predominant selfing and predominant outcrossing are found to be alternative stable states of the mating system in most plant populations. Which of these stable states a species approaches depends on the history of its population structure and the magnitude of effect of genes influencing the selfing rate.
DOI:10.1093/aob/mcp014URLPMID:19202136 [本文引用: 1]
BACKGROUND AND AIMS: Evolutionary change in response to natural selection will occur only if a trait confers a selective advantage and there is heritable variation. Positive connections between pollen traits and fitness have been found, but few studies of heritability have been conducted, and they have yielded conflicting results. To understand better the evolutionary significance of pollen competition and its potential role in sexual selection, the heritability of pollen tube-growth rate and the relationship between this trait and sporophytic offspring fitness were investigated in Collinsia heterophylla. METHODS: Because the question being asked was if female function benefited from obtaining genetically superior fathers by enhancing pollen competition, one-donor (per flower) crosses were used in order to exclude confounding effects of post-fertilization competition/allocation caused by multiple paternity. Each recipient plant was crossed with an average of five pollen donors. Pollen-tube growth rate and sporophytic traits were measured in both generations. KEY RESULTS: Pollen-tube growth rate in vitro differed among donors, and the differences were correlated with in vivo growth rate averaged over two to four maternal plants. Pollen-tube growth rate showed significant narrow-sense heritability and evolvability in a father-offspring regression. However, this pollen trait did not correlate significantly with sporophytic-offspring fitness. CONCLUSIONS: These results suggest that pollen-tube growth rate can respond to selection via male function. The data presented here do not provide any support for the hypothesis that intense pollen competition enhances maternal plant fitness through increased paternity by higher-quality sporophytic fathers, although this advantage cannot be ruled out. These data are, however, consistent with the hypothesis that pollen competition is itself selectively advantageous, through both male and female function, by reducing the genetic load among successful gametophytic fathers (pollen), and reducing inbreeding depression associated with self-pollination in plants with mix-mating systems.
[本文引用: 1]
DOI:10.11913/PSJ.2095-0837.2015.20133URL [本文引用: 1]
Thamnolia vermicularis (Sw.)Ach. ex Schaer. and Thamnolia subuliformis (Ehrh.)W. L. Culb. were carried out, and variant under these two species have also been studied. Very few differences were observed between the original species and variant, though intermediate forms were found, and chemical compounds were the same, thus, T.vermicularis f.qomolangmana and T.subuliformis f.minor were synonymized. Taxonomic characteristics of Thamnolia are clarified, species boundaries are newly defined, and distribution ranges of the genus Thamnolia in China are discussed in this paper.]]>
DOI:10.11913/PSJ.2095-0837.2015.20133URL [本文引用: 1]
Thamnolia vermicularis(Sw.)Ach. ex Schaer.)和雪地茶(Thamnolia subuliformis(Ehrh.)W. L. Culb.)及其种下变型进行了形态分类和特征化合物研究。结果显示,2个变形在形态上均与原变形之间无明显界限,存在过渡形态,并且化学成分也均与原变形一致,因此合并了2个种下变型。通过分析地茶属地衣的分类特征,对地茶属2个种的分类界限进行了重新界定,讨论了我国地茶属的地理分布。]]>
[本文引用: 1]
DOI:10.5846/stxb201704100621URL [本文引用: 1]
Sagittaria trifolia L.)为研究材料,将相同基因型组成的植株分别种植在光照差异明显的两个同质园(林荫区与日照区),于盛花期对野慈姑的开花数量、昆虫访花进行了观察,确定其主要访花昆虫、记录访花行为以及野慈姑的繁殖产出水平。研究表明,日照区野慈姑每天开放的花朵和植株数量显著高于林荫区。两个区域内野慈姑的主要访花者有四大类:蚜蝇类、蜂类、蝶类和蝇类;其中林荫区的主要传粉昆虫为蚜蝇类,而日照区为蜂类。日照区昆虫单位时间内的访花频率、昆虫每回合访问的雄花数、总花数和花序数均显著高于林荫区。日照区野慈姑的座果率显著高于林荫区,而单果种子数量与种子面积与林荫区相当。总体而言,日照区野慈姑的繁殖产出达到了林荫区的三倍以上,这是环境的直接作用与环境介导的传粉作用在同一方向上叠加的结果,且前者占主导地位。在关注植物、物理环境和生物因子三者之间互作的前提下,量化每个繁殖阶段的水平,特别是传粉昆虫在不同生境下的表现,阐述了环境异质性导致植物繁殖差异的根本原因。]]>
DOI:10.1007/s11032-018-0907-xURL
DOI:10.1111/j.1558-5646.1988.tb04185.xURLPMID:28581087 [本文引用: 1]
After pollen arrives on a stigma, the paternity of seeds may be influenced by microgametophyte competition, maternal choice, genetic complementation between parents, and embryo competition. While microgametophyte competition has been well accepted, the other mechanisms are more difficult to demonstrate, and their effects are often confounded. Here, wild radish plants were pollinated with single and mixed pollen loads, and some plants were stressed such that reproduction was reduced. Effects of pollen donors, maternal families, maternal x paternal interaction, pollen donor number, and stress on fruit abortion, seed number per fruit, seed weight, and total seed weight per fruit were measured. Maternal-plant x pollen-donor interaction effects were found for all variables, indicating that genetic complementation or maternal choice occurred. Values of the components of reproduction were generally higher for multiply sired fruits than for singly sired fruits, indicating that either competition among embryos changed under multiple paternity or maternal choice for multiply sired fruits occurred. Finally, when maternal plants were stressed, the components of reproduction were more strongly affected by seed and fruit paternity. This result indicates that either competitive regimes among embryos were affected by stress or maternal plants become more selective under stress. In both cases where embryo competition might have been an explanation of the results, variation in seed weight within fruits was unaffected, suggesting that competitive regimes were unchanged. Clearly, mechanisms in addition to microgametophyte competition are important in sorting the pollen that arrives on stigmas of wild radish. These data suggest that maternal choice is likely to be important. In addition, these processes are likely to occur in the field, since the effects are stronger in stressed than in control plants.
DOI:10.1086/284494URL
DOI:10.1111/j.1558-5646.2007.00167.xURLPMID:17683434 [本文引用: 1]
For sexual selection to be important in plants, it must occur at pollen load sizes typical of field populations. However, studies of the impact of pollen load size on pollen competition have given mixed results, perhaps because so few of these studies directly examined the outcome of mating when pollen load size was varied. We asked whether seed paternity after mixed pollination of wild radish was affected by pollen load sizes ranging from 22 to 220 pollen grains per stigma. We examined the seed siring abilities of 12 pollen donors across 11 maternal plants. Seed paternity was statistically indistinguishable across the pollen load sizes even though, overall, the pollen donors sired different numbers of seeds. This lack of effect of pollen load size on seed paternity may have occurred because fruit abortion and early abortion or failure of fertilization of seeds increased as load size decreased. Thus, failures of fruits and seeds sired by poorer pollen donors may keep seed paternity constant across pollen load sizes.
DOI:10.1093/aob/mcy167URLPMID:30535041 [本文引用: 1]
Background: The male fitness pathway, from pollen production to ovule fertilization, is thought to strongly influence reproductive trait evolution in animal-pollinated plants. This pathway is characterized by multiple avenues of pollen loss which may lead to reductions in male fitness. However, empirical data on the mechanistic processes leading to pollen loss during transport are limited, and we therefore lack a comprehensive understanding of how male fitness is influenced by each step in the pollination process. Scope: This review assesses the history of studying male function in plants and identifies critical gaps in our understanding of the ecology and evolution of pollen transport. We explore male reproductive function along the steps of the pathway to paternity and discuss evolutionary options to overcome barriers to siring success. In particular, we present a newly emerging idea that bodies of pollinators function as a dynamic arena facilitating intense male-male competition, where pollen of rival males is constantly covered or displaced by competitors. This perspective extends the pollen-competitive arena beyond the confines of the stigma and style, and highlights the opportunity for important new breakthroughs in the study of male reproductive strategies and floral evolution.
DOI:10.3732/ajb.89.8.1250URLPMID:21665726 [本文引用: 1]
In small, fragmented populations of self-incompatible plant species, genetic drift and increasingly close relationships between plants may restrict the number of genetically different pollen donors, the availability of compatible mates, and the opportunity for pollen competition and selection. These restrictions may reduce the siring success or increase the probability of inbreeding depression in the offspring. To test if this was the case, we hand-pollinated maternal plants in small and large populations of the rare, endemic plant Cochlearia bavarica (Brassicaceae) with pollen from one, three, or nine donors from the same population or with nine donors from a different population. In one additional population of intermediate size, maternal plants were hand-pollinated with ten donors located at a distance of 1, 10, 100, or 1000 m. We then recorded seed and offspring characters. On average, offspring from small populations were smaller than normal and fewer survived to maturity. Increasing the number of pollen donors had a positive effect on reproductive success in small and large populations, but at the highest pollen diversity this occurred at the expense of slightly reduced offspring fitness. Because the total amount of transferred pollen was held constant, these effects could not be attributed to increasing pollen load. Rather, the increasing pollen diversity may have increased the chances of selecting a particularly
DOI:10.1111/jeb.12692URLPMID:26174371 [本文引用: 1]
Flowers fertilized by multiple fathers may be expected to produce heavier seeds than those fertilized by a single father. However, the adaptive mechanisms leading to such differences remain unclear, and the evidence inconsistent. Here, we first review the different hypotheses predicting an increase in seed mass when multiple paternity occurs. We show that distinguishing between these hypotheses requires information about average seed mass, but also about within-fruit variance in seed mass, bias in siring success among pollen donors, and whether siring success and seed mass are correlated. We then report the results of an experiment on Dalechampia scandens (Euphorbiaceae), assessing these critical variables in conjunction with a comparison of seed mass resulting from crosses with single vs. multiple pollen donors. Siring success differed among males when competing for fertilization, but average seed mass was not affected by the number of fathers. Furthermore, paternal identity explained only 3.8% of the variance in seed mass, and siring success was not correlated with the mass of the seeds produced. Finally, within-infructescence variance in seed mass was not affected by the number of fathers. These results suggest that neither differential allocation nor sibling rivalry has any effect on the average mass of seeds in multiply sired fruits in D. scandens. Overall, the limited paternal effects observed in most studies and the possibility of diversification bet hedging among flowers (but not within flowers), suggest that multiple paternity within fruits or infructescence is unlikely to affect seed mass in a large number of angiosperm species.
DOI:10.3732/ajb.1500126URLPMID:26451034 [本文引用: 1]
PREMISE OF THE STUDY: Competition among pollen grains from a single donor is expected to increase the quality of the offspring produced because of the recessive deleterious alleles expressed during pollen-tube growth. However, evidence for such an effect is inconclusive; a large number of studies suffer from confounding variation in pollen competition with variation in pollen load. METHODS: In this study, we tested the effect of pollen competition on offspring performance independently of pollen-load variation. We compared seed mass and early seedling performance in Dalechampia scandens (Euphorbiaceae) between crosses in which variation in pollen competition was achieved, without variation in pollen load, by manipulating the dispersion of pollen grains on the stigmas. KEY RESULTS: Despite a large sample size (211 crosses on 20 maternal plants), we failed to find an effect of pollen competition on seed characteristics or early seedling performance. Paternal effects were always limited, and pollen competition never reduced the within-father (residual) variance. CONCLUSION: These results suggest that limited within-donor variation in genetic quality of pollen grains reduces the potential benefits of pollen competition in the study population. The lack of paternal effects on early sporophyte performance further suggests that benefits of pollen competition among pollen from multiple donors should be limited as well, and it raises questions about the significance of pollen competition as a mechanism of sexual selection.
DOI:10.1038/277294a0URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1046/j.1365-2540.2001.00897.xURLPMID:11703509 [本文引用: 3]
[本文引用: 1]
DOI:10.3732/ajb.91.2.192URLPMID:21653375 [本文引用: 1]
In monoecious plants, gametes can be exchanged in three ways: among unrelated genets (outbreeding), with close relatives (inbreeding), or within individuals (geitonogamous selfing). These different mating systems may have consequences for population demography and fitness. The experiment presented herein used artificial crosses to examine the mating system of Chesapeake Bay, Virginia, USA eelgrass (Zostera marina L; Zosteraceae), a bisexual submerged aquatic plant that can outbreed, inbreed, and self. Genetic data indicate severe heterozygosity deficiencies and patchy genotype distribution in these beds, suggesting that plants therein reproduce primarily by vegetative propagation, autogamy, or geitonogamy. To clarify eelgrass reproductive strategies, flowers from three genetically and geographically distinct beds were hand-pollinated in outbred, inbred, and selfed matings. Fertilization success and seed production, life history stages which contribute greatly to the numeric maintenance of populations, were monitored. We found no evidence that inbreeding had negative consequences for seed production. On the contrary, selfed matings produced seeds significantly more frequently than outcrossed matings and produced significantly larger numbers of seeds than either inbred or outbred matings. These results contrast with patterns for eelgrass in other regions but might be expected for similar populations in which pollen limitation or a short reproductive season renders selfing advantageous.
DOI:10.1111/bij.2010.101.issue-4URL
DOI:10.1016/S1146-609X(02)01146-3URL [本文引用: 3]
DOI:10.1007/BF00985462URL [本文引用: 1]
Lathyrus vernus (L.) Bernh. is a diploid, long-lived perennial and insect-pollinated herb with no special adaptation to long-distance dispersal. It occurs on neutral soil in deciduous forests throughout western Eurasia. Due to specific habitat preferences,L. vernus has a fragmented distribution with isolated populations. We investigated allozyme variation at eleven loci in 20 populations ofL. vernus from one geographically central region (the Czech Republic and the Slovak Republic) and two geographically marginal regions (southern and central Sweden) in the species present-day distribution. There was a clear differentiation between the three regions and the genetic distance between the populations was highly correlated with geographic distance. The total genetic diversity (HT) was 0.354. The proportion of genetic diversity due to differentiation between regions, and to differentiation between populations within regions, accounted for 10% each. There was no difference in level of genetic diversity between the three regions. No significant difference in level of genetic diversity was found between small and large populations. The genetic diversity inL. vernus may either be a result of the long generation-time of the species or peculiarities in the post-glacial migration species, e.g. survival only in refugia far east of the sampled populations and/or migration as a continuous process not involving founder-events.]]>
[本文引用: 1]
DOI:10.1002/j.1537-2197.1992.tb14586.xURL [本文引用: 1]
[本文引用: 2]
DOI:10.1007/BF00365567URLPMID:28306918 [本文引用: 1]
In thisstudy we combine field experiments, designed to test the predictions of optimal outcrossing theory in Agave schottii, with molecular genetic studies, using RAPD (random amplified polymorphic DNA), polymerase chain reaction to assess the underlying genetic hypothesis of optimal outcrossing theory. Initially, 48
DOI:10.1093/aob/mcv118URLPMID:26229065 [本文引用: 2]
BACKGROUND AND AIMS: Interactions between species are especially sensitive to environmental changes. The interaction between plants and pollinators is of particular interest given the potential current global decline in pollinators. Reduced pollinator services can be compensated for in some plant species by self-pollination. However, if inbreeding depression is high, selfed progeny could die prior to reaching adulthood, leading to cryptic recruitment failure. METHODS: To examine this scenario, pollinator abundance, pollen limitation, selfing rates and inbreeding depression were examined in 12 populations of varying disturbance levels in Sophora microphylla (Fabaceae), an endemic New Zealand tree species. KEY RESULTS: High pollen limitation was found in all populations (average of 58 % reduction in seed production, nine populations), together with high selfing rates (61 % of offspring selfed, six populations) and high inbreeding depression (selfed offspring 86 % less fit, six populations). Pollen limitation was associated with lower visitation rates by the two endemic bird pollinators. CONCLUSIONS: The results suggest that for these populations, over half of the seeds produced are genetically doomed. This reduction in the fitness of progeny due to reduced pollinator service is probably important to population dynamics of other New Zealand species. More broadly, the results suggest that measures of seed production or seedling densities may be a gross overestimate of the effective offspring production. This could lead to cryptic recruitment failure, i.e. a decline in successful reproduction despite high progeny production. Given the global extent of pollinator declines, cryptic recruitment failure may be widespread.
DOI:10.1007/s10682-010-9395-0URL [本文引用: 1]
Geum urbanum. Assessment of mating system parameters using microsatellite markers inferred high selfing rates (92.5%), confirming the predominantly self-fertilizing character of the study species. We conducted experimental pollinations with self and outcross pollen collected from populations at different distances from the target populations. There were no significant signs of inbreeding depression, even in very small target populations. Except for a minor negative effect on the germination rate for the long-distance crosses, we found no effects of outbreeding on fitness estimates.]]>
DOI:10.1038/hdy.2010.79URL [本文引用: 3]
Populations of predominantly selfing plant species often show spatial genetic structure but little is known whether epistatic gene interactions are spatially structured. To detect a possible epistatic effect and a spatial scale at which it operates, we created artificial crosses between plants spanning a range of fixed distances from 1 to 400 m in three populations of wild barley. The self-pollinated and crossed progeny (F(1)) and two generations of segregated progeny (F(2) and F(3)) were tested in experimentally simulated population environments for relative performance (RP). The measured fitness traits included number of seeds, total seed weight and seed germination. For any of these traits, there was no association between RP of F(1), F(2) and F(3) plants and either pairwise kinship coefficients or crossing distance. In contrast, in all three populations, we found lower seed viability of outcrossed as compared with self-pollinated genotypes in the first generation of segregation. However, in the F(3) generation this outbreeding effect disappeared in the two populations and greatly decreased in the third population. For seed production, heterosis in F(1) and outbreeding depression in F(2) were observed only in the population with unusually high number of heterozygotes. Our findings support the view that in selfing species a spatial mosaic of various locally abundant genotypes represents not randomly fixed combinations of alleles but the co-adapted gene complexes that were sieved by selection, while heterozygotes are characteristic for the transient phase of this process, when segregation and purging of maladaptive genotypes have not yet occurred. Heredity (2011) 106, 300-309; doi:10.1038/hdy.2010.79; published online 16 June 2010
DOI:10.1111/j.1558-5646.1994.tb01366.xURLPMID:28568280 [本文引用: 2]
Depending on its genetic causes, outbreeding depression in quantitative characters may occur first in the free-living F1 generation produced by a wide cross. In 1981-1985, we generated F1 progenies by hand-pollinating larkspurs (Delphinium nelsonii) with pollen from 1-m, 3-m, 10-m, or 30-m distances. From the spatial genetic structure indicated by previous electrophoretic and reciprocal transplantation studies, we estimate that these crosses range from being inbred (f approximately 0.06) to outbred. We planted 594 seeds from 66 maternal sibships under natural conditions. As of 1992, there was strong evidence for both inbreeding depression and outbreeding depression. Progeny from intermediate crossing distances grew approximately twice as large as more inbred or outbred progeny in the first 5 yr after planting (P = 0.013, repeated measures ANOVA), and survived almost 1 yr longer on average (contrast of 3-m and 10-m treatments versus 1 m and 30 m; P = 0.028, ANOVA). Twenty maternal sibships produced flowering individuals; only four and two of these represented 1-m and 30-m crossing distances, respectively (P = 0.021, G-test). The cumulative fitness of intermediate distance sibships averaged about twice that of 1-m sibships, and five to eight times that of 30-m sibships (P = 0.017, ANOVA). Thus, even though progeny of 1-m crosses were inbred to a degree only about one-eighth that of selling, inbreeding depression approximated 50%, and outbreeding depression equaled or exceeded 50% for all fitness components.
DOI:10.3724/SP.J.1142.2014.40427URL
Pinus pumila seed viability, five treatments, namely, water soak, NaOH solution soak, PEG6000 (PEG, polyethylene glycol) solution soak, low-temperature, absorption of water and desiccation treatments, were used to explore factors that affect the viability of Pinus pumila seeds. Groups of each factor were tested under different concentrations, durations and temperatures, with indigo dyeing applied to measure seed viabilities. Results showed that the PEG6000 solution soak at 10% concentration, initial temperature of 60℃, and 36 h of soaking showed the most improved seed viability. Water soaking also enhanced the viability of the Pinus pumila seeds, as well as cryogenics and absorption of water and desiccation treatments. NaOH solution with concentrations of 4%-12% lowered seed viability. Testing revealed that the PEG6000 treatment was the most effective method for improving the viability of Pinus pumila, which can be applied to the cultivation and better utilization of Pinus pumila.]]>
DOI:10.3724/SP.J.1142.2014.40427URL
为了提高偃松种子生活力,采用水浸、NaOH溶液浸泡、PEG6000(PEG,聚乙二醇)溶液浸泡、低温冷冻、吸湿/回干5种方法对偃松种子进行处理,研究不同处理中各因素(如溶液浓度、处理时间与温度)对偃松种子生活力的影响,并采用靛蓝染色法测定偃松种子的生活力。结果表明:10%PEG6000溶液、初始温度60℃、浸泡36 h时,偃松种子生活力最强;水浸、低温处理和吸湿/回干处理方法也能提高偃松种子生活力;然而4% ~ 12% NaOH溶液浸泡处理会导致偃松种子生活力下降。因此可将PEG6000溶液处理种子的方法应用于偃松种子萌发及人工种苗培育的实践中,使偃松资源得到更好的可持续开发与利用。
URL [本文引用: 3]
[本文引用: 3]
[本文引用: 2]
URL [本文引用: 2]
URL
发育中的幼果或种子选择性败育的现象。植株可以选择性地败育位于果序顶部或基部的果实以及位于果实基部、中部或柱头端的种子。此现象 在被子植物中比较普遍,特别是在豆科、十字花科和紫草科中最为常见。导致植物选择性败育的主要原因主要有资源限制和遗传因子两个方面 。植物通过选择性败育部分自交或基因型较差的果实或种子,不仅可以提高母本和后代的适合度,而且还可以提高果实或种子的扩散效率。因 此,对选择性败育的研究在深入了解植物的结实结籽格局、探讨其 进化式样与机制等方面具有重要意义。该文系统总结了国际上有关植物选择 性败育的研究工作,重点介绍了选择性败育发生的式样、导致选择性败育的因素、选择性败育的进化生态意义,以及目前研究选择性败育现象 的主要方法,并对该领域今后研究前景进行了展望。]]>
DOI:10.3724/SP.J.1142.2014.30259URL [本文引用: 1]
Xanthoceras sorbifolium, a self-incompatible species. Results showed that increasing the diversity of compatible pollen enhanced the mean number of fruit in an inflorescence and seed number per fruit and mean seed weight. The highest mean number of fruits in an inflorescence, the highest mean number of seeds per fruit, and the highest mean weight of seeds were produced by four, three and three compatible donors, respectively. If mixed pollen donors contained self-pollen, all fruits were aborted. In addition to diversity of pollen genotypes, recipient genotypes also affected female reproductive success. The present results suggest that greater attention to high diversity varieties may help maintain or enhance fruit and seed production in X.sorbifolium. The efffect of pollen genetic diversity, maternal mate choice and selective fruit abortion were discussed in this paper.]]>
DOI:10.3724/SP.J.1142.2014.30259URL [本文引用: 1]
Xanthoceras sorbifolium)进行1、2、3、4种花粉供体传粉实验,调查了不同花粉供体数量对文冠果雌性生殖的影响。结果表明,增加亲和花粉供体的多样性,一定程度上提高了花序上果实的坐果数量,同时也增加了果实中种子的数量及单粒种子重量。对文冠果授以4种亲和性花粉供体后,花序上成熟果实的平均数最多;3种亲和性花粉供体传粉后,每个果实中的种子数最多,单粒种子也最重。如果混合花粉中含有自交花粉,那么果实将败育。除花粉多样性影响雌性生殖产出外,母本基因型也具有重要影响,即增加文冠果种植品系的多样性,将有助于提高果实和种子产量。本研究还讨论了花粉遗传多样性效应、母本交配选择及果实选择性败育假说。]]>
URLPMID:17246494 [本文引用: 3]
The effect of the rate of partial self-fertilization and viability selection on the magnitude of inbreeding depression was investigated for the overdominance genetic model. The influence of these factors was determined for populations with equilibrium genotypic frequencies. Inbreeding depression was measured as the normalized disadvantage in mean viability of selfed progeny as compared to outcrossed progeny. When caused by symmetric homozygous disadvantage at a single locus it is shown always to be less than one-third. Moreover, for fixed rates of self-fertilization, its maximum value is found at intermediate levels of homozygous disadvantage. As the rate of self-fertilization increases, inbreeding depression increases and the homozygote viability that results in maximum depression tends toward one-half the heterozygote viability. Symmetric selection against homozygotes at multiple loci can lead to substantially higher values than selection at a single-locus. As the number of independent loci involved increases, inbreeding depression can reach high levels even though the selfing rate is low. Viability distributions for progenies produced from both random mating and self-fertilization were derived for the case of symmetric selection at independently assorting multiple loci. Distributions of viabilities in progenies resulting from mixtures of selfing and outcrossing were shown to be bimodal when inbreeding depression is high.
Outcrossing rate and inbreeding depression in two annual monoecious herbs, Begonia hirsuta and B. semiovata
2
1993
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Habitat continuity and geographic distance predict population genetic differentiation in giant kelp
1
2010
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
Inbreeding depression in benign and stressful environments
1
2005
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
The effects of inbreeding, outbreeding and long-distance gene flow on survivorship in North American populations of Silene vulgaris
1
2006
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Fitting linear mixed-effects models using lme4
1
2015
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
Consequences of mixed pollinations in Campsis radicans
2
1986
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
Do plant populations purge their genetic load? Effects of population size and mating history on inbreeding depression. Annual Review of Ecology, Evolution,
1
1999
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Genetic loads and estimates of mutation rates in highly inbred plant populations
1
1990
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Inbreeding depression and its evolutionary consequences. Annual Review of Ecology, Evolution,
2
1987
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... )和失去了某些生存发育能力相关位点的杂合优势(heterozygote advantage at viability loci)(
1
1989
... 野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(
1
1989
... 野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(
Phylogeography of a marsh herb Sagittaria trifolia (Alismataceae) in China inferred from cpDNA atpB-rbcL intergenic spacers
1
2008
... 野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(
Perspective: purging the genetic load: a review of the experimental evidence
1
2002
... 另一方面, 野慈姑也未表现出远交衰退(
Sequential decline in fruit resource allocation within inflorescences of Sagittaria trifolia: a test of non-uniform pollination hypothesis
5
2018a
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
... 野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(
... 野慈姑是多年生挺水植物, 既可以通过地下球茎进行营养繁殖, 亦可以通过蜜蜂、食蚜蝇等昆虫授粉完成有性生殖, 且自交亲和(
... 为探究野慈姑是否存在自交或远交衰退现象, 本实验设置自交、近距离异交和远距离异交3个水平, 拟以近距离异交的子代性状为参照, 将自交后代、远距离异交后代与其进行对比.在本实验中, 综合考虑了水系连通性和传粉昆虫的飞行能力分别影响的种子与花粉散布, 把种群之间直线距离小于50 km的交配定义为近距离异交, 大于200 km的交配定义为远距离异交.具体方法为以每一个种群为中心, 列出它与其他种群之间的距离, 把植株来源小于50 km的作为可选近距离父本, 大于200 km的作为可选远距离父本(
... 人工授粉以每朵雌花为重复单元, 采用雄花花药与雌花心皮直接接触的方法, 每朵雌花用两朵雄花授粉, 直至球形心皮上的花粉均匀饱和.为了控制不同基因型的母本效应, 5个处理都尽量选择在同一个花序上进行, 每种处理对应的轮次位置随机, 以避免位置效应(
Resource reallocation patterns within Sagittaria trifolia inflorescences following differential pollination
3
2018b
... 野慈姑是多年生挺水植物, 既可以通过地下球茎进行营养繁殖, 亦可以通过蜜蜂、食蚜蝇等昆虫授粉完成有性生殖, 且自交亲和(
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
... 不论交配距离或父本数量如何改变, 野慈姑种子的萌发指标都没有表现出显著变化(
High selfing rate, limited pollen dispersal and inbreeding depression in the emblematic African rain forest tree Baillonella toxisperma Management implications
2016
Heterosis
1
1936
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Does parental divergence predict reproductive compatibility?
1
2002
... 另一方面, 野慈姑也未表现出远交衰退(
Between a rock and a hard place: evaluating the relative risks of inbreeding and outbreeding for conservation and management
1
2007
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
Mating structure and inbreeding and outbreeding depression in the rare plant Gentianella germanica (Gentianaceae)
2
1997
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
An {R} Companion to Applied Regression
1
2019
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Predicting the probability of outbreeding depression
1
2011
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
Transgenerational plasticity is adaptive in the wild
1
2007
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
How are deleterious mutations purged? Drift versus nonrandom mating
1
2003
... 综上所述, 以异交为主的野慈姑经过人工自交后, 其适合度和F1表现并未降低, 200 km距离的种群间交配也未发现远交衰退现象, 频繁的自交历史、远距离的基因扩散和相似的物理生境可能扮演了至关重要的作用.不过, 鉴于野慈姑广泛的分布范围, 建议在类似物种的研究中, 适当拓宽交配距离或延长观测周期, 以深入探讨交配距离对繁殖的影响.此外, 本研究基于野慈姑20余个自然种群的基因型, 相较于单一种群的研究, 实验样本具有更高的基因多样性.从实验设计上, 每个种群的基因型都有均等的机会作为父本或母本参与交配, 避免了以单一或少量种群作为母本交配的遗传背景影响(
植物繁育系统研究的最新进展和评述
1
2003
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
植物繁育系统研究的最新进展和评述
1
2003
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Quantitative and qualitative consequences of reduced pollen loads in a mixed-mating plant
2019
Effects of pollen load and donor diversity on seed and fruit mass in the Columnar Cactus, Pachycereus schottii (Cactaceae)
1
2009
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Inbreeding and outbreeding depression in Stylidium hispidum: implications for mixing seed sources for ecological restoration
1
2012
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
Plant ecotypes: genetic differentiation in the age of ecological restoration
1
2003
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
Inbreeding depression and selfing rate of Ipomoea hederacea var. integriuscula (Convolvulaceae)
1
2005
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
The effect of inbreeding in diploid and tetraploid populations of Epilobium angustifolium (Onagraceae): implications for the genetic basis of inbreeding depression
2
1997
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Correlated evolution of self-fertilization and inbreeding depression: an experimental study of nine populations of Amsinckia (Boraginaceae)
1
1996
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Heterosis: many genes, many mechanism End the search for an undiscovered unifying theory
1
2012
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Multiple pollinator visits to Mimulus ringens (Phrymaceae) flowers increase mate number and seed set within fruits
1
2006
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Outcrossing rate and inbreeding depression in the perennial yellow bush lupine, Lupinus arboreus (Fabaceae)
1
2000
... 另一方面, 野慈姑也未表现出远交衰退(
Effect of pollen load, source and mixture on reproduction success of four cultivars of Citrullus lanatus (Thunb.) Matsumara and Nakai (Cucurbitaceae)
2
2013
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
Sibling competition and evolution of pollen unit, ovule number, and pollen vector in angiosperms
1
1981
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
lmerTest package: tests in linear mixed effects models
2
2017
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
The evolution of self-fertilization and inbreeding depression in plants. I: Genetic models
1
1985
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
Pollen-tube growth rates in Collinsia heterophylla (Plantaginaceae): one-donor crosses reveal heritability but no effect on spoophytic-offspring fitness
1
2009
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
Least-Squares Means: the R package lsmeans
1
2016
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
利用SSR荧光标记对野慈姑异交率的估测
1
2015
... 另一方面, 野慈姑也未表现出远交衰退(
利用SSR荧光标记对野慈姑异交率的估测
1
2015
... 另一方面, 野慈姑也未表现出远交衰退(
两种生境下野慈姑繁殖差异及其机制
1
2018
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
两种生境下野慈姑繁殖差异及其机制
1
2018
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
Genetic distance and the relationship with heterosis and reproductive behavior in tetraploid bahiagrass hybrids
2019
Postpollination effects on seed paternity: mechanisms in addition to microgametophyte competition operate in wild radish
1
1988
... 实验于2018年8月在中国科学院武汉植物园(114.42° E, 30.54° N)内野慈姑的同质园进行(
Sexual selection in Raphanus sativus: experimental data on nonrandom fertilization, maternal choice, and consequences of multiple paternity
1986
Effects of pollen load size on seed paternity in wild radish: the roles of pollen competition and mate choice
1
2007
... 实验于2018年8月在中国科学院武汉植物园(114.42° E, 30.54° N)内野慈姑的同质园进行(
Plant-pollinator interactions along the pathway to paternity
1
2019
... 另一方面, 野慈姑也未表现出远交衰退(
Effects of population size and pollen diversity on reproductive success and offspring size in the narrow endemic Cochlearia bavarica (Brassicaceae)
1
2002
... 另一方面, 野慈姑也未表现出远交衰退(
Does multiple paternity affect seed mass in angiosperms? An experimental test in Dalechampia scandens
1
2015
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
Does stronger pollen competition improve offspring fitness when pollen load does not vary?
1
2016
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
Pollen dispersal and optimal outcrossing in Delphinium nelsoni
1
1979
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
野慈姑花粉限制评估的影响因素
2
2015
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
野慈姑花粉限制评估的影响因素
2
2015
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
Evidence for local outbreeding depression in the Mediterranean island endemic Anchusa crispa Viv. (Boraginaceae)
3
2001
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... ;
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
psych: Procedures for Personality and Psychological Research
1
2018
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
Seed production from the mixed mating system of Chesapeake Bay (USA) eelgrass (Zostera marina; Zosteraceae)
1
2004
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
Signficant differences in outcrossing rate, self-incmpatibility, and inbreeding depression between two widely hybridizing species of Geum
2010
Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment
3
2002
... 野慈姑(Sagittaria trifolia)为泽泻科慈姑属多年生的草本植物, 在亚洲范围内广泛分布, 常见于池塘、水稻田、水渠等地, 可以通过实生苗和球茎进行有性和无性繁殖、维系种群(
... 2019年10月对剩余的种子进行了第二次萌发实验.由于这批种子进行了烘干处理, 因此在这次萌发之前对种子进行了两个月的浸泡(
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Allozyme diversity in relation to geographic distribution and population size in Lathyrus vernus (L.) Bernh. (Fabaceae)
1
2000
... 另一方面, 野慈姑也未表现出远交衰退(
Mating system, outcrossing distance effects and pollen availability in the wind-pollinated treeline species Polylepis australis BITT. (Rosaceae)
1
2009
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
Fitness consequences of mixed-donor pollen loads in the annual legume Chamaecrista fasciculata
1
1992
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
R: a Language and Environment for Statistical Computing
2
2016
... 本实验包括5种处理, 7个变量: 坐果概率、单果种子数量、种子面积、种子萌发率(2018和2019年)以及幼苗芽长(2018和2019年).除坐果概率(0、1的二元分布), 对其他变量进行了正态性分析, 其中种子面积和幼苗芽长满足正态分布.单果种子数量经过开平方转换、萌发率经过开方后的反正弦转换, 正态性均有所提高, 故统计中使用转换后数据.全部统计用R软件(
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Field and genetic studies testing optimal outcrossing in Agave schottii, a long-lived clonal plant
1
1995
... 另一方面, 野慈姑也未表现出远交衰退(
The compounding effects of high pollen limitation, selfing rates and inbreeding depression leave a New Zealand tree with few viable offspring
2
2015
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Effects of outcrossing in fragmented populations of the primarily selfing forest herb Geum urbanum
1
2010
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
Epistasis in natural populations of a predominantly selfing plant
3
2011
... 除了交配距离, 花粉多样性在很大程度上也会影响繁殖的结果.多项研究指出, 随着父本多样性的提高, 果实和种子产量均得到了不同程度的提升(
... ).也有研究指出, 当纳入实验的多个父本之间竞争能力及其与母本个体的兼容性没有明显差异时, 繁殖水平与后代表现也不会优于单个父本的交配(
... 本实验中, 与单父本处理相比, 双父本处理对野慈姑的种子产量和质量的促进作用有限(
植物保护遗传学
2003
植物保护遗传学
2003
Crossing-distance effects in Delphinium nelsonii: outbreeding and inbreeding depression in progeny fitness
2
1994
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... ;
若干种子处理方法对偃松种子生活力的影响
2014
若干种子处理方法对偃松种子生活力的影响
2014
沙葱种子的萌发特性和几种贮藏物质含量的变化
3
2007
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
... ;
... 综上所述, 以异交为主的野慈姑经过人工自交后, 其适合度和F1表现并未降低, 200 km距离的种群间交配也未发现远交衰退现象, 频繁的自交历史、远距离的基因扩散和相似的物理生境可能扮演了至关重要的作用.不过, 鉴于野慈姑广泛的分布范围, 建议在类似物种的研究中, 适当拓宽交配距离或延长观测周期, 以深入探讨交配距离对繁殖的影响.此外, 本研究基于野慈姑20余个自然种群的基因型, 相较于单一种群的研究, 实验样本具有更高的基因多样性.从实验设计上, 每个种群的基因型都有均等的机会作为父本或母本参与交配, 避免了以单一或少量种群作为母本交配的遗传背景影响(
沙葱种子的萌发特性和几种贮藏物质含量的变化
3
2007
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
... ;
... 综上所述, 以异交为主的野慈姑经过人工自交后, 其适合度和F1表现并未降低, 200 km距离的种群间交配也未发现远交衰退现象, 频繁的自交历史、远距离的基因扩散和相似的物理生境可能扮演了至关重要的作用.不过, 鉴于野慈姑广泛的分布范围, 建议在类似物种的研究中, 适当拓宽交配距离或延长观测周期, 以深入探讨交配距离对繁殖的影响.此外, 本研究基于野慈姑20余个自然种群的基因型, 相较于单一种群的研究, 实验样本具有更高的基因多样性.从实验设计上, 每个种群的基因型都有均等的机会作为父本或母本参与交配, 避免了以单一或少量种群作为母本交配的遗传背景影响(
植物交配系统的进化、资源分配对策与遗传多样性
2
2001
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
植物交配系统的进化、资源分配对策与遗传多样性
2
2001
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... 交配距离影响有性繁殖的现象十分普遍, 但每个物种的表现不一, 主要有以下4种模式.(1)自交衰退与远交衰退都存在, 即植物可能有最适的异交距离.如假龙胆属(Gentianella)植物G. germanica相距10 m个体间异交所产子代的萌发率、叶片数量、莲座大小均高于自然授粉、自交、1 m异交及25 km外的种群间异交(
种子植物的选择性败育及其进化生态意义
2007
种子植物的选择性败育及其进化生态意义
2007
文冠果花粉供体的多样性对果实产出的影响
1
2014
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
文冠果花粉供体的多样性对果实产出的影响
1
2014
... 研究结果发现, 野慈姑没有明显的自交衰退现象(
Inbreeding depression due to overdominance in partially self-fertilizing plant populations
3
1989
... 利用各种传粉媒介把花粉转移到柱头上从而完成有性繁殖是有花植物的一大进化优势(
... ), 因此许多研究者通过设置不同的交配距离来观测物种的自交或远交衰退情况(
... 另一方面, 野慈姑也未表现出远交衰退(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}