 ,1,2,*, 赵玉金3, 郑朝菊1, 熊杰1,2, 赵旦1
,1,2,*, 赵玉金3, 郑朝菊1, 熊杰1,2, 赵旦1Forest species diversity mapping based on clustering algorithm
Hai-Yan YI1,2, Yuan ZENG,1,2,*, Yu-Jin ZHAO3, Zhao-Ju ZHENG1, Jie XIONG1,2, Dan ZHAO1通讯作者: *zengyuan@aircas.ac.cn
编委: 沈国春
责任编辑: 赵航
收稿日期:2019-12-12接受日期:2020-02-7网络出版日期:2020-06-20
| 基金资助: |
Corresponding authors: *zengyuan@aircas.ac.cn
Received:2019-12-12Accepted:2020-02-7Online:2020-06-20
| Fund supported: |

摘要
该研究基于机载激光雷达(LiDAR)和高光谱数据, 从森林物种叶片的生理化学源头探寻生化特征与光谱特征的内在关联, 探讨生化多样性、光谱多样性与物种多样性之间的响应机制, 选择最优植被指数并结合最优结构参数, 通过聚类方法构建森林物种多样性遥感估算模型, 在古田山自然保护区开展森林乔木物种多样性监测。研究结果表明: (1)从16种叶片生化组分中, 筛选出叶绿素a、叶绿素b、类胡萝卜素、叶片含水量、比叶面积、纤维素、木质素、氮、磷和碳可通过偏最小二乘法用叶片光谱有效模拟(R2 = 0.60-0.79, p < 0.01), 并选择有效的植被指数: 转换型吸收反射指数/优化型土壤调整指数(TCARI/OSAVI)、类胡萝卜素反射指数(CRI)、水波段指数(WBI)、比值植被指数(RVI)、生理反射指数(PRI)和冠层叶绿素浓度指数(CCCI)表征相应的最优生化组分; (2)基于机载LiDAR数据利用结合形态学冠层控制的分水岭算法获得高精度单木分离结果(R 2 = 0.77, RMSE = 16.48), 同时采用逐步回归方法从常用的森林结构参数中选取树高和偏度作为最优结构参数(R 2 = 0.32, p < 0.01); (3)基于6个最优植被指数和2个最优结构参数, 以20 m × 20 m为窗口通过自适应模糊C均值方法进行聚类, 实现了研究区森林乔木物种丰富度(Richness, R 2= 0.56, RMSE = 1.81)和多样性指数Shannon-Wiener (R 2 = 0.83, RMSE = 0.22)与Simpson (R 2 = 0.85, RMSE = 0.09)的成图。该研究在冠层尺度上获取了与物种多样性相关的生化、光谱和结构参数, 将单木个体作为最小单元, 利用聚类算法直接估算物种类别差异, 无需判定具体的树种属性, 是利用遥感数据进行区域尺度森林物种多样性监测与成图的实践, 可为亚热带地区常绿阔叶林的物种多样性监测提供借鉴。
关键词:
Abstract
Aims Monitoring forest species diversity continuously and efficiently is important to maintain ecosystem services and achieve sustainability and conservation goals. In this paper, we explored the relationship between leaf biochemical and spectral properties and their inner linkage with species diversity, then estimated the forest species diversity based on a clustering algorithm using airborne imaging spectroscopy and Light Detection and Ranging (LiDAR) data in the Gutianshan National Nature Reserve of China.
Methods Firstly, we isolated individual tree crowns (ITCs) with the watershed algorithm from the LiDAR data. Then we calculated the optimal vegetation indices (VIs) representing the key biochemical properties from the hyperspectral data and selected optimal structural parameters from commonly used LiDAR-derived structural parameters based on correlation and stepwise regression analysis with the field samples. Finally, a self-adaptive Fuzzy C-Means (FCM) clustering algorithm was applied to map the species diversity (i.e. Richness, Shannon-Wiener index and Simpson index) in the study area for each 20 m × 20 m moving window.
Important findings The results indicated that biochemical components (chlorophyll a & b, total carotenoids, equivalent water thickness, specific leaf area, cellulose, lignin, nitrogen, phosphorus and carbon) could be well quantified by leaf spectrum using partial least squares regression (R2 = 0.60-0.79, p < 0.01), and represented by hyperspectral VIs, namely, Transformed Chlorophyll Ratio Index/Optimization of Soil-adjusted Vegetation Index (TCARI/OSAVI), Carotenoid Reflectance Index (CRI), Water Band Index (WBI), Ratio Vegetation Index (RVI), Photochemical Reflectance Index (PRI) and Canopy Chlorophyll Concentration Index (CCCI). The individual tree isolation showed high accuracy (R 2 = 0.77, RMSE = 16.48). The correlation and stepwise regression analysis showed tree height and skewness were the optimal structural parameters among seven commonly used forest structural parameters (R 2 = 0.32, p < 0.01). The species diversity indices calculated from the self-adaptive FCM clustering algorithm based on the six VIs and two optimal structural parameters correlated well with the field measurements (species richness, R 2 = 0.56, RMSE = 1.81; Shannon-Wiener index, R 2 = 0.83, RMSE = 0.22; Simpson index, R 2 = 0.85, RMSE = 0.09). With the clustering method combined with crown-by-crown variations in hyperspectral biochemical VIs and LiDAR-derived structural parameters, we created continuous maps of forest species diversity in the examined subtropical forest without the need to identify specific tree species. Our case study in Gutianshan showed the potential of airborne hyperspectral and LiDAR data in mapping species diversity of the subtropical evergreen broad-leaved forest. It could also provide a pathway for monitoring the state and changes of forest biodiversity at regional scales.
Keywords:
PDF (14036KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
衣海燕, 曾源, 赵玉金, 郑朝菊, 熊杰, 赵旦. 利用聚类算法监测森林乔木物种多样性. 植物生态学报, 2020, 44(6): 598-615. DOI: 10.17521/cjpe.2019.0347
YI Hai-Yan, ZENG Yuan, ZHAO Yu-Jin, ZHENG Zhao-Ju, XIONG Jie, ZHAO Dan.
森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(Díaz et al., 2006; Harrison et al., 2014)。物种多样性是研究生物多样性的关键, 它既可反映不同环境与群落之间的关系, 又反映了生境中生物资源的丰富度。然而, 由于人类活动和气候变化, 目前全球森林物种数量正在锐减(郑华和欧阳志云, 2014), 从生态保护和管理的角度出发, 监测森林物种多样性至关重要。传统的森林物种多样性监测主要基于野外样方调查(Kerr & Ostrovsky, 2003), 重点关注在样方尺度的森林物种分布(Myers et al., 2000; Duro et al., 2007)。遥感具有探测范围广、获取数据周期短、动态性强等优势, 在时间和空间尺度上延拓了森林物种多样性监测的可能。目前, 利用遥感方法监测森林物种多样性, 主要有直接和间接两种方法(Turner et al., 2003)。直接法主要通过识别物种或群落的类型, 以获取物种个体名称和数量。该方法依赖于不同林分的纹理信息特征, 对于遥感数据的空间分辨率和光谱分辨率要求较高。同时, 由于森林生态系统环境的复杂性, 林分的空间异质性增加了直接识别物种的难度。间接法是基于遥感数据衍生的参数或变量, 利用相关分析或聚类理论构建模型来估算物种多样性指数。间接法的关键在于选择合适的参数来表示物种特征, 植被的生化组分、植被指数、结构参数以及环境参数等对应着物种-光谱、物种-结构和物种-生境关系, 是进行物种多样性研究的重要指标。
生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(Running, 1990; Goetz & Prince, 1996)。森林冠层的生化组分差异, 在很大程度上取决于物种组成。多样性高的植被群落, 其叶片生化组分的差异明显(Townsend et al., 2008)。叶片生化组分含量调控着叶片波谱的吸收程度, 进而影响遥感观测到的冠层光谱(Ustin et al., 2004)。国内外对于叶片光谱和生化组分的关联已有大量研究, 尤其是随着高光谱技术的发展, 尝试从叶片生化组分的机理出发, 通过模拟和建立光谱特征与生化特性的关系来进行物种识别和多样性评估(王纪华等, 2003; Carlson et al., 2007)。Asner和Martin (2008, 2009)基于高光谱数据利用偏最小二乘法反演不同物种叶片的生化组分(R2 = 0.47-0.81), 印证了物种光谱与生化组分之间的内在关联, 揭示了森林物种的生化足迹具有唯一性, 基于此提出“光谱组学(spectranomics)”方法用于热带雨林物种多样性监测。Zhao等(2016)将该方法应用于亚热带常绿阔叶林研究区, 利用高光谱数据基于叶片生化与光谱的关联筛选出8种生化组分用于森林乔木物种多样监测 (R2 = 0.58-0.85)。
激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(Lim et al., 2003; Coops et al., 2016)。研究表明基于结构参数与物种多样性指数的相关关系可实现物种多样性的估算, 例如Lopatin等(2016)基于机载LiDAR数 据提取了海拔高度、坡度标准偏差和平均冠层高度用于亚热带地中海地区次生林的物种多样性监测 (R2 = 0.66, nRMSE = 16.29%); Simonson等(2012)在同一地区的乔木物种多样性研究中发现, 平均树高与物种多样性最为相关(R2 = 0.50, p < 0.001), 利用9种结构参数对于2类优势树种的分类准确度可达89%; 董文雪等(2018)对我国亚热带常绿阔叶林物种多样性的研究表明, 95%分位数高度、冠层盖度和植被穿透率与物种多样性指数的相关性相对较高(R2 = 0.39-0.42)。然而, 结构参数并不是决定树种类型的关键因素, 仅利用LiDAR数据进行树种分类, 会受到树冠形态、物种复杂程度、植被发育状况、树龄等因素影响, 无法获得较高精度的估算结果。但将激光雷达获取的结构特征与生化组分和高光谱表征的光谱特征相结合, 在区域尺度物种多样性监测中将发挥巨大作用(Rocchini et al., 2018; Wang & Gamon, 2019)。
聚类分析可以搜索并识别一个有限的数据集 合, 发现数据的全局分布模式, 聚类方法的基础在于能被聚为一类的各样本具有相似的属性集合。对于森林物种来说, 同一物种的光谱由于具有本质相同的生化组分而具有高度相似性, 在区域分布上同一物种还在树高、冠层结构等方面表现出相近性, 这种一致使得某一物种呈现出能够被遥感识别的有异于其他物种的特征, 从而可直接估算物种类别差异, 无需判定具体的树种属性。因此, 在冠层尺度如获取了与物种多样性相关的生化、光谱和结构参数, 将单木个体作为最小单元并通过聚类分析, 可估算物种类别的差异度, 即为物种丰富度。Zhao等(2018)在我国神农架自然保护区基于机载LiDAR和高光谱数据, 在单木尺度上利用7个生化组分和树高, 通过自适应模糊C均值聚类方法, 估算了研究区的乔木物种丰富度(RMSE = 2.20)和Shannon-Wiener指数(R2 = 0.83, RMSE = 0.25), 证实了在生化-光谱-物种多样性的关联机制下通过聚类方法估算区域尺度多样性指数的可行性。但是该研究仍存在一些问题, 例如: 可预测的物种数会随着生化组分的增加而达到饱和, 在聚类参数选取时, 不同研究区会受到生化组分与光谱表达的制约; 同一树种的生化组分与结构特征会随着个体发育情况、地貌地形、人为管理措施和数据获取时间的不同而产生较大的种内差异, 在该研究区所选取的生化与结构参数仅为该生态情境下的最优组合, 在其他区域物种多样性监测中的适用性尚不明确。
为进一步探讨以上问题, 本研究选择亚热带常绿阔叶林生物多样性的典型研究基地古田山自然保护区作为研究区, 同步获取机载LiDAR和高光谱遥感数据, 基于生化特征、光谱差异及物种多样性三者的关系, 提出适用于本研究区乔木物种多样性监测的最优叶片生化组分、冠层植被指数和结构参数, 在单木尺度上选择最佳移动窗口利用自适应模糊C均值聚类方法, 构建乔木物种多样性估算模型, 并讨论成图结果的合理性与方法的适用性, 以期为亚热带地区常绿阔叶林的森林物种多样性监测提供参考。
1 材料和方法
1.1 研究区概况
研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(祝燕等, 2008; Legendre et al., 2009; 钱海源等, 2018), 同时, 古田山还是中国森林生物多样性监测网(Sino Bon-CForBio)中亚热带常绿阔叶林的重要研究基地(马克平, 2015; 米湘成等, 2016)。本研究以古田山保护区的核心地带作为研究 区, 地理坐标为118.08°-118.18° E, 29.19°-29.28° N, 总面积约104 km2 (图1)。古田山属南岭山系怀玉山脉的一部分, 主峰青尖海拔1 258 m, 整个山峰呈东北-西南走向。该地区属我国中亚热带湿润季风气候区, 四季分明, 雨量充沛, 年平均气温15.3 ℃, 年降水量1 963.7 mm。古田山母岩以花岗岩为主, 土壤类型属中亚热带山地红黄壤类型, 局部地区有沼泽土分布。所选研究区涵盖了古田山保护区内完整的植被垂直带谱, 典型的地带性植被包括以甜槠(Castanopsis eyrei)、木荷(Schima superba)和青冈(Cyclobalanopsis glauca)等为优势种的中亚热带常绿阔叶林, 位于海拔较高地区以黄山松(Pinus taiwanensis)为优势种的针阔混交林(700-1 100 m)和针叶林(1 100 m以上), 山地、沟谷地带分布有常绿落叶阔叶混交林, 还有大片原始状态的天然次生林(于明坚等, 2001)。
图1
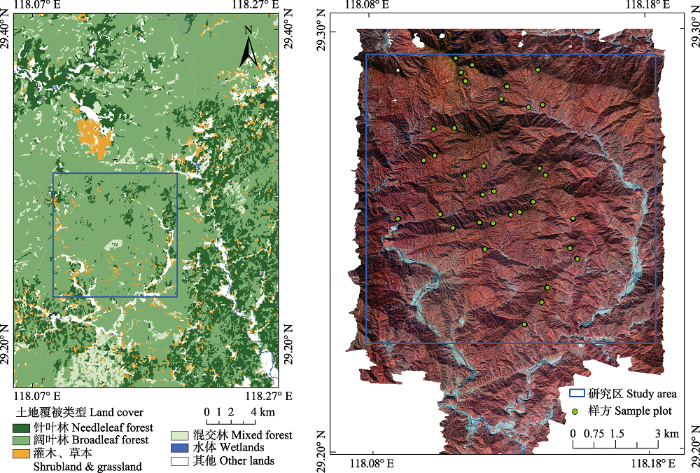 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1古田山研究区土地覆被类型及地面观测样方分布。
Fig. 1Land cover and locations of forest field measured sample plots in Gutianshan study area.
1.2 数据及预处理
1.2.1 遥感数据机载LiDAR及高光谱数据于2017年10月由搭载在运5飞机上的LiCHy系统获取(Pang et al., 2016)。航飞区内共获取航带24条, 平均飞行高度为2 000 m。其中, LiDAR数据由激光扫描仪(LMS-Q560, Riegl, Horn, Austria)获得, 扫描角为±30o, 点云平均密度为3.5个·m-2; 高光谱数据由高光谱传感器(AISA Eagle Ⅱ, Specim, Oulu, Finland)获得, 光谱范围为400-970 nm, 光谱分辨率为3.3 nm, 空间分辨率为1 m。
LiDAR数据的预处理依据国家测绘地理信息局发布的《机载激光雷达数据处理技术规范》和《机载激光雷达数据后处理技术规范》进行, 利用TerraSolid软件将点云自动分为地面点(Max building size = 60, Iteration angle = 6, Iteration distance = 1.4)和植被点(除地面及建筑物), 随后由点云的首次回波点插值生成数字表面模型(DSM), 再由点云中的地面点通过构建不规则三角网(TIN)插值生成数字高程模型(DEM), 最后由两者通过差值运算得到冠层高度模型(CHM)(Zhao et al., 2013)。在开展森林结构参数提取时, 还需将点云数据的高程值减去对应位置DEM像元值, 得到点云距离地面的相对高度, 即去除地形影响后的归一化点云数据。
高光谱数据预处理包括辐射定标、几何校正、大气校正和地形辐射校正, 由LiCHy系统配套的CaliGeoPro软件实现。几何校正首先利用同平台的外部GPS、IMU和高精度DEM数据进行坐标重采样后得到影像的地理坐标, 随后进行Boresight检校; 大气校正基于ATCOR 4软件采用MODTRAN 5模型进行; 地形辐射校正结合综合辐射校正模型(IRC)与ATCOR 4大气校正模型共同改善山区阴面地物光谱失真现象。为消除太阳位置影响, 利用ATCOR 4软件提供的经验BRDF校正模型使高光谱影像地表反射率统一至传感器天顶角为0°, 太阳天顶角为45°。最后进行航带拼接, 利用直方图匹配消除航带色差。
此外, 本研究区的土地覆被数据来自30 m空间分辨率的中国土地覆被数据集(ChinaCover)。该数据集以国产环境卫星HJ数据和美国陆地卫星Landsat数据为数据源, 依据在FAO LCCS分类工具支持下定义的适合中国土地覆盖类型特点与应用需求的分类系统, 将中国土地覆被分为森林、草地、耕地、湿地、人工表面及其他6个一级类和40个二级类(Wu et al., 2017)。本研究区的土地覆被是在ChinaCover二级类的基础上进行了合并, 共包括针叶林、阔叶林、混交林、灌草、水体及其他六大类(图1)。
1.2.2 样方观测数据
地面观测数据的获取与航飞同步进行, 共获得了20 m × 20 m样方34个(图1)。样方的四角点坐标利用差分GPS (定位精度± 2 m)确定。样方内获取的单木参数包括树种名称、胸径(DBH ≥ 5 cm)、树高、枝下高、冠幅等。其中, 胸径用胸径尺测量, 树高用激光测高仪(TruPulse200, TruPulse, Texas, USA)测量, 冠幅采用目估法分别记录东西和南北两个走向的长度。同时还获取了样方的海拔、坡度、坡向、土壤类型和林下环境等信息, 样方的森林郁闭度和叶面积指数(LAI)分别利用鱼眼相机和植物冠层分析仪(LAI2000, LI-COR, Lincoln, USA)测量获得。此外, 在34个样方中, 随机选择了6个样方利用全站仪测量了单木树干根部位置(精度±0.5 m), 主要用于冠层结构参数提取的精度验证。
基于地面观测数据, 分别计算了34个样方的物种丰富度、Shannon-Wiener指数(H)和Simpson指数(D)。物种丰富度即物种数目, 可通过样方内的物种数来表示。Shannon-Wiener和Simpson指数既能体现物种的丰富程度, 又能体现物种分布的均匀度, 是反映物种多样性程度的综合指标。其中, Shannon- Wiener指数对于物种数目更为敏感, Simpson指数对于富集种的均匀度更为敏感。两种指数可通过物种丰富度及每个物种所包含的单木个数计算得到。计算公式如下:
式中, pi表示含有n个物种的群落中第i个物种个体数占所有物种总个体数的比例(Pielou, 1975)。
1.2.3 生化组分测量数据
基于地面观测的样方单木调查记录和文献资 料, 根据森林树种的代表性和分布范围, 选择在数量上占优势、出现频率高且分布均匀的17个树种作为研究区的优势树种(于明坚等, 2001; 陈彬等, 2017)。同时, 根据植被生化组分的作用和特性, 选取16种常用的生化组分, 对这17个优势树种的叶片进行生化组分测定。
利用高枝剪收集完全光照条件下的冠层叶片将其分为两组。其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(李云梅等, 2003)。另一组根据每个优势树种叶片大小和质量各采集200-300枚叶片, 在6小时内测定湿质量, 之后随机选择3组, 在20 cm × 20 cm单位面积下利用扫描法测定真实叶面积, 最后将全部叶片置于85 ℃环境下烘干至恒质量, 保证每个优势树种叶片的总干质量大于5 g。其中, 叶片含水量(EWT)和比叶面积(SLA)与光捕获、化学物质分布密切相关(Wright et al., 2004), EWT用叶片湿质量与干质量之差与其单叶面积的比值表示, SLA为叶片单叶面积与其干质量的比值; 碳、氮(N)、磷(P)是植物体内组成糖类、蛋白质、核酸及叶绿素等物质的必需元素, 影响植物光合作用、呼吸作用等内部生化过程及外在形态(Long et al., 2006), N含量可利用元素分析仪(Vario EL cube, Elementar, Hanau, Germany)测定, 碳含量利用重铬酸钾-外加热法测定, P含量利用酸溶-钼锑抗比色法测定; 纤维素(Cel)和木质素(Lig)等叶片结构化合物起到支撑、维持和防御作用, 增强了植物对不良外界环境的抵抗性(Melillo et al., 1982), 其含量可利用范式法测定(Van Soest, 1963); 钾(K)能够增强细胞内酶活性, 其含量利用电感耦合等离子发射光谱仪(ICAP-6500, Thermo, Waltham, USA)测定; 微量元素, 如钙(Ca)、镁(Mg)、锌(Zn)、锰(Mn)、硼(B)同样利用元素分析仪测定。考虑到树种分布对生化组分的影响, 每一树种采集3组位于不同海拔位置的叶片, 使用3组叶片生化组分测定值的平均值, 并将其与比叶面积做比值计算, 最终换算为单位面积的生化组分含量(表1)。
Table 1
表1
表1古田山研宄区优势树种叶片生化组分
Table 1
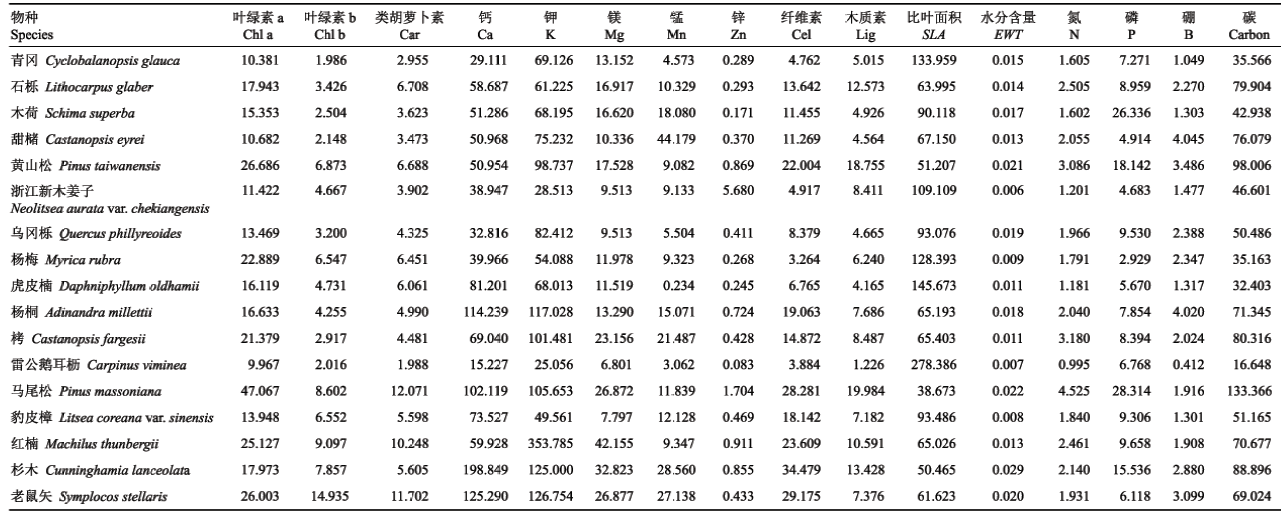 |
新窗口打开|下载CSV
1.2.4 叶片光谱观测数据
在实验室利用标准型地物光谱仪(FieldSpec 4 Std-Res, Analytica Spectra Devices, Colorado, USA)对地面同步采集的17个优势树种的叶片分别进行光谱测定, 每个树种随机选择10个叶片计算平均值(图2)。考虑到仪器噪声的影响, 移除波长小于400 nm和大于2 400 nm的数据, 并对400-2 400 nm之间的反射率进行Savitzky-Golay滤波平滑处理(Savitzky & Golay, 1964)。
图2
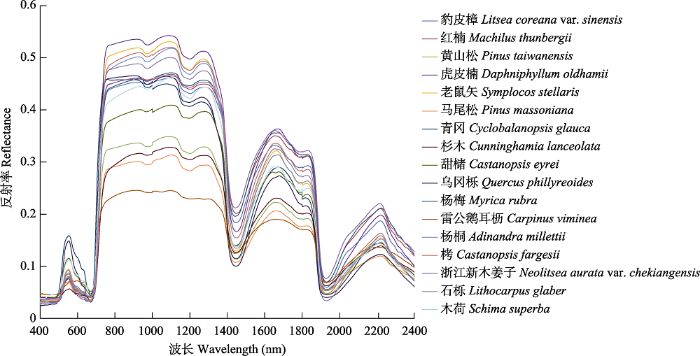 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2古田山研究区17个优势树种叶片光谱曲线。
Fig. 2Spectral signatures of 17 dominant tree species in Gutianshan study area.
1.3 多样性监测方法
本研究的总体技术路线如图3所示。物种多样性估算的关键在于选取与多样性指数相关性高的生化组分、植被指数和结构参数。首先基于叶片光谱和叶片生化组分通过回归分析选取能够表征物种多样性的最优生化组分, 利用高光谱数据计算与之对应的最优植被指数; 其次根据结构参数与物种多样性的回归分析选取最优结构参数, 引入单木分离算法计算单木尺度上的最优植被指数和最优结构参数; 最后利用自适应模糊C均值聚类方法, 按20 m × 20 m窗口获取最佳聚类数, 即物种丰富度, 并计算Shannon-Wiener和Simpson指数, 利用地面观测数据进行验证, 实现在研究区的森林乔木物种多样性监测与成图。图3
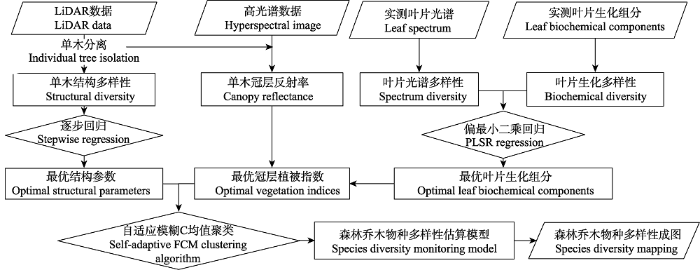 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3古田山研究区森林乔木物种多样性监测技术路线。
Fig. 3Technique flow chart of forest species diversity mapping in Gutianshan study area.
1.3.1 单木分离算法
以基于LiDAR数据生成的经过无效值填充的冠层高度模型CHM为基础, 利用结合形态学冠层控制的分水岭算法实现单木的分离(Zhao et al., 2013, 2014)。首先在CHM上应用形态学闭运算, 以样方内单木枝下高的平均值(1.8 m)作为阈值来确定树冠区域, 该阈值代表研究区乔木枝下高的平均水平; 随后对原始CHM应用中值滤波得到整型灰度影像以减少树冠的过识别, 在树冠内部采用局部极值法探测可能的树冠顶点位置; 接着对经形态学冠层控制的CHM进行两次分水岭变换以勾绘树冠形状, 在两次分水岭变换之间加入一次图像重建以移除过分割树冠; 最后利用形态学开运算对树冠进行优化从而还原真实树冠形状。此时得到的单木树冠根据地面调查数据设置了最小树冠阈值(9个0.2 m分辨率像元), 因此已去除了小图斑的干扰, 从而使地面调查数据与遥感数据能够更好地匹配。该算法利用IDL编程语言实现。
1.3.2 基于最优叶片生化组分的植被指数选取
野外采集的森林物种叶片生化组分含量差异明显, 不同的生化组分具有不同的量纲和单位, 要分析17种优势物种生化组分的唯一性, 则需对叶片的16种生化组分进行标准化(归一化)处理。Min-Max标准化法通过对原始数据进行线性变换, 可使不同评价指标在同一数量级下进行比较分析, 转换函数如下:
${{M}_{ij}}=({{X}_{ij}}-\text{min}({{X}_{ij}}))/(\text{max}({{X}_{ij}})-\text{min}({{X}_{ij}}))$
其中,${{M}_{ij}}$为i物种的生化组分j的归一化结果,${{X}_{ij}}$为i物种的生化组分j的实测值,$\text{max}({{X}_{ij}})$和$\text{min}({{X}_{ij}})$
分别为生化组分实测值的最大值和最小值。该方法可在Excel 2010软件中实现。
在16个常用生化组分中选取能够表征森林物种的最优生化组分。首先基于叶片实测光谱利用偏最小二乘回归(Partial Least Squares Regression, PLSR)方法预测16种叶片生化组分含量, 通过与实测叶片生化组分含量的对比分析选取相关性较高的生化组分作为最优生化组分。偏最小二乘回归方法是一种多对多的线性回归方法, 汇集了主成分分析、典型相关分析和多元线性回归分析的优点, 能够在生化组分少、光谱波段多且相关性强的情况下建立回归模型, 可通过JMP 11.0软件实现。随后, 将预测的与实测的叶片生化组分含量进行对比分析, 选择相关性相对高的作为最优生化组分, 同时还可以借鉴前人研究的分析结果, 如Asner等(2009)选择了光谱预测精度R2 = 0.54-0.83的5种生化组分; 赵玉金(2016)选择了精度范围在R2 = 0.57-0.85的8种生化组分。
确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数。其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(Daughtry et al., 2000; Wu et al., 2008); Car利用类胡萝卜素反射率指数(CRI)表示(Gitelson et al., 2002); EWT通过水波段指数(WBI)反演得到(Penuelas et al., 1993); SLA利用比值植被指数(RVI))表示(Jordan, 1969)。一些生化组分由于相互之间相关性较强或者不能被光谱直接表达, 需要通过转化以寻求适当的植被指数予以表征, 如Cel和Lig的光谱吸收特征重叠(Kokaly et al., 2009), 绿色植物中选用叶黄素代替纤维素, 可选择与之对应的生理反射指数(PRI)(Gamon et al., 1992)来表示以上两种生化组分; N和P光谱吸收特征重叠, 且N与植物体的叶绿素密切相关, 可用冠层叶绿素浓度指数(CCCI)表示(El-Shikha et al., 2007)。此外, 碳在研究中很少进行叶片层次的光谱特征分析, 微量元素通常借助波段变换或与其他植被指数建立相关关系来反映, 缺乏与之直接对应的植被指数。在选定了代表最优生化组分的植被指数后, 基于LiDAR和高光谱数据计算出单木尺度的最优植被指数, 作为物种多样性估算模型的输入参数时, 可将其与单木LAI进行比值处理, 削弱冠层结构对植被指数的影响(Zarco-Tejada et al., 2001)。植被指数的计算可利用ENVI 5.3软件实现。
1.3.3 基于LiDAR数据的冠层结构参数选取
常用的森林结构参数主要有95%分位数高度、平均植被高度、首次回波标准偏差、首次回波偏度、平均绝对偏差、植被穿透率、叶高度多样性等(表2), 分别在森林的垂直结构和内部结构上反映着森林物种的多样性特征。在研究区基于LiDAR数据计算34个样方单木尺度的7个结构参数, 通过相关分析评价结构参数对物种多样性指数的解释程度, 在此基础上利用逐步回归分析(Stepwise Regression, SR)方法选取与物种多样性最相关的结构参数作为最优结构参数。逐步回归分析建立在线性回归分析基础上, 将具有显著意义的自变量逐一引入方程中, 并对每一个自变量进行显著性检验, 从而保留影响最为显著的变量, 剔除对因变量不起作用或作用极小的变量, 使回归方程更合理地反映自变量与因变量的关系, 可通过SPSS 22.0软件实现。
Table 2
表2
表2常用冠层结构参数
Table 2
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | Ria?o et al., 2004 |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | Lefsky et al., 2002 |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | Moffiet et al., 2005 |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | Clawges et al., 2008 |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | Nelson et al., 1988 |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | Zhao et al., 2018 |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis | Antonarakis et al., 2008 |
新窗口打开|下载CSV
1.3.4 自适应模糊C均值聚类
聚类分析通过对数据集合进行合理分割以确定类别数, 使同一类中数据尽可能相近, 不同类之间数据尽可能相异(Wang et al., 2006)。其中, 模糊C均值聚类算法(Fuzzy C-Means clustering algorithm, FCM)能够将聚类过程转化为一个非线性优化问题, 通过迭代来求解类别数, 是非监督模式识别的重要方法。模糊C均值聚类对于给定的样本数据集X = {x1, x2, …, xn}, 将其划分为c类(2 ≤ c ≤ n)并确定聚类中心V = {v1, v2, …, vn}, 样点属于某一类的隶属度用模糊矩阵U = [uij]表示, 目标函数J(U,V)及其约束条件如下:
式中, uij表示第j个样本点属于第i类的隶属度, ${{d}_{ij}}=$$||{{x}_{j}}-{{v}_{i}}||$为第j个样本点与第i个聚类中心之间的欧氏距离, m ≥ 1是模糊加权指数。
模糊C均值聚类的基本原则为通过迭代调整隶属度矩阵U和聚类中心V, 使得目标函数J(U,V)取得极小值, 通过拉格朗日乘法求得J(U,V)的条件极值, 分别得到U和V的迭代形式, 其中k表示迭代步数, ε为迭代阈值, $V_{0}^{k}$为初始聚类中心。
自适应模糊C均值聚类算法(Self-adaptive Fuzzy C-Means clustering algorithm)发展于模糊C均值聚类, 通过运用有效性函数来自动获取聚类数目, 无需依赖预先设定的类别数, 解决了聚类算法对于初始值的敏感问题。Li和Yu (2009)提出的改进有效性函数L(c)如下:
L(c)的分子表示类间距离之和, 分母表示所有类内距离之和。L(c)越大, 则聚类结果越可靠。聚类结果即为样方内的物种丰富度, 基于最佳聚类数和每个聚类组包含的个体数可以计算Shannon-Wiener和Simpson指数。该算法利用IDL编程语言实现。
2 结果
2.1 单木分离
34个样方单木分离估算结果与实测单木数量的对比分析如图4A所示, 以有全站仪测量数据的25号样方为例(图4B), 表示实测单木根部位置与LiDAR数据经单木分离得到的树冠位置进行匹配的结果。单木分离结果在数量上接近实测单木数(R2 = 0.77, RMSE = 16.48), 但是由于点云密度、树冠重叠、树冠过小等原因的影响, 导致出现过分割和欠分割现象, 树冠位置不能与实测单木位置完全匹配(N?sset & ?kland, 2002; Koch et al., 2006; Zhao et al., 2014)。过分割常出现在树冠较大, 树顶不突出的阔叶单木中; 欠分割则是由于森林郁闭度较高造成树冠重叠, 多个重叠树冠被认为是一个树冠而不被分割。25号样方的单木分离结果为90, 实测单木数目为106, 因欠分割导致单木分离数量略低于实测单木数量。图4
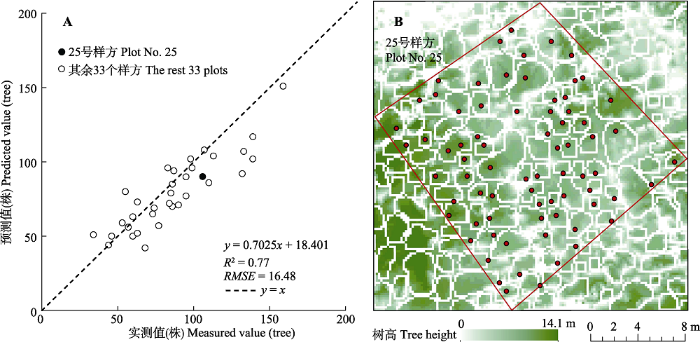 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4古田山研究区样方单木分离精度验证散点图(A)和25号样方的单木分离结果图(B)。红框为样方边界, 白框为分离出的单木树冠, 红点为实测单木根部位置。
Fig. 4Validation result of individual tree isolation (A) and individual tree isolation result of plot No. 25 (B) in Gutianshan study area. The red polygon represents plot border, the white polygons represent canopy locations, and the red dots represent field-measured positions of the base of tree trunk.
2.2 最优生化组分
野外采集的17个优势树种的16种叶片生化组分的归一化处理结果如图5所示。各条带代表了不同树种的生化组分组合, 同一条带内各色带的长度代表了不同生化组分对该优势种的相对贡献大小, 同一色带在不同条带中的长度代表了该生化组分含量在不同种间的差异。生化组分越多, 越能体现物种间的差异。例如马尾松和黄山松同属于针叶林, 针叶的水分含量相近, 但叶绿素和氮含量相差很大。16个生化组分的归一化结果表明了17个优势树种均具有唯一的生化特征, 不同物种的生化特征在元素的组成和含量上差异明显。图5
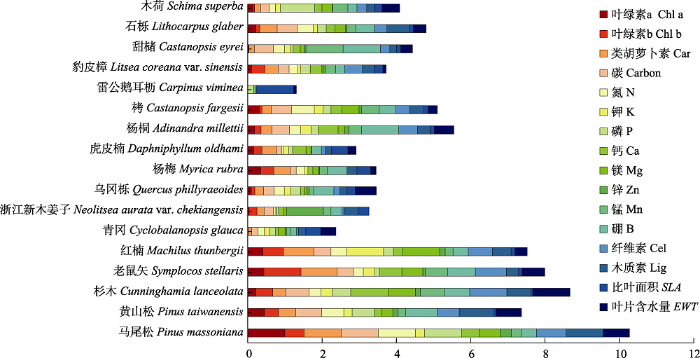 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5古田山研究区17个优势树种的16种叶片生化组分归一化结果。
Fig. 516 standardized biochemical components of 17 dominant tree species in Gutianshan study area. Car, carotenoids; Cel, cellulose; Chl a, chlorophyll a; Chl b, chlorophyll b; Lig, lignin; SLA, specific leaf area; EWT, equivalent water thickness.
将叶片生化组分预测值与实测值进行比较(表3)发现: 比叶面积、叶绿素a、木质素、碳的预测效果较好(R2 = 0.70-0.79), 其次为叶绿素b、类胡萝卜素、纤维素、水分含量、N、P (R2 = 0.60-0.69), 其他组分的预测能力明显低于前面几种组分(R2 ≤ 0.51)。综合16个常用生化组分的预测精度及前人的研究, 最终选择了以上10种生化组分(R2 = 0.60-0.79)作为最优叶片生化组分。
Table 3
表3
表3叶片生化组分预测精度
Table 3
| 生化组分 Biochemical component | 叶绿素a Chl a | 叶绿素b Chl b | 类胡萝卜素 Car | 比叶面积 SLA | 水分含量 EWT | 纤维素 Cel | 木质素 Lig | 碳 Carbon | 氮 N | 磷 P | 钙 Ca | 钾 K | 镁 Mg | 锰 Mn | 锌 Zn | 硼 B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | 0.70 | 0.60 | 0.69 | 0.79 | 0.67 | 0.66 | 0.76 | 0.72 | 0.63 | 0.63 | 0.51 | 0.23 | 0.48 | 0.10 | 0.09 | 0.21 |
新窗口打开|下载CSV
在选择10种最优生化组分对应的植被指数时, 基于高光谱数据结合各组分的光谱特征和内在关联对其进行了合并和筛选, 最终选择TCARI/OSAVI、CRI、WBI、CCCI、RVI和PRI共6个植被指数作为聚类模型的输入参数(表4)。其中, 叶绿素a和叶绿素b合并用TCARI/OSAVI表示, 类胡萝卜素用CRI表示, 水分含量用WBI表示, N和P合并用CCCI表示, 纤维素和木质素用叶黄素对应的PRI表示, 比叶面积用RVI表示, 而碳因没有明显的光谱吸收波段予以去除。
Table 4
表4
表4最优生化组分所对应的植被指数及其计算公式
Table 4
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | Daughtry et al., 2000; Wu et al., 2008 |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | Gitelson et al., 2002 |
| 水分 EWT | WBI | WBI = R895/R972 | Penuelas et al., 1993 |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | El-Shikha et al., 2007 |
| 比叶面积 SLA | RVI | RVI = R750/R705 | Jordan, 1969 |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) | Gamon et al., 1992 |
新窗口打开|下载CSV
2.3 最优结构参数
对34个样方的7个结构参数的自相关性及其与实测多样性指数的相关性进行分析(表5)。垂直结构参数中, 95%分位数高度(近似于树高)与物种丰富度和物种多样性指数的相关性最高; 内部结构参数中, 偏度与物种丰富度和物种多样性指数的相关性最高。逐步回归分析结果显示, Simpson指数回归模型同样将95%分位数高度和偏度选为最优结构参数, 预测模型为D = 1.237 - 0.036 × 95%分位数高度- 0.094 ×偏度(R2 = 0.32, p < 0.01), 该结果表示利用两个结构参数可解释约32%的物种多样性。本研究选择的95%分位数高度和偏度均与高度信息相关, 能够在一定程度上反映树种的多样性特征, 故将其纳入聚类模型的输入参数中。Table 5
表5
表5基于激光雷达提取的结构参数相关性分析
Table 5
| 结构多样性参数 Structural parameter | Simpson 指数 Simpson index | Shannon- Wiener 指数 Shannon- Wiener index | 物种丰富度 Species richness | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 95%分位 数高度 95% Quantile height | 平均植 被高度 Mean vegetation height | 平均绝对偏差 Mean absolute deviation | 标准偏差 Standard deviation | 偏度 Skewness | 叶高度 多样性 Foliage height diversity | 植被 穿透率 Vegetation permeability | |||||
| 垂直结构 Vertical structure | 95%分位数高度 | 1.000 | 0.884** | 0.757** | 0.811** | -0.320 | -0.233 | 0.233 | 0.455** | 0.320 | 0.088 |
| 平均植被高度 | 0.884** | 1.000 | 0.428* | 0.511** | -0.578** | 0.171 | 0.318 | -0.352* | -0.187 | 0.012 | |
| 平均绝对偏差 | 0.757** | 0.428* | 1.000 | 0.988** | -0.055 | -0.763** | 0.134 | -0.314 | -0.236 | 0.002 | |
| 标准偏差 | 0.811** | 0.511** | 0.988** | 1.000 | -0.167 | -0.700** | 0.214 | -0.328 | -0.244 | 0.010 | |
| 内部结构 Inner structure | 偏度 | -0.320 | -0.578** | -0.055 | -0.167 | 1.000 | -0.281 | -0.546** | -0.171 | -0.225 | 0.152 |
| 叶高度多样性 | -0.233 | 0.171 | -0.763** | -0.700** | -0.281 | 1.000 | 0.227 | 0.020 | 0.020 | 0.123 | |
| 植被穿透率 | 0.233 | 0.318 | 0.134 | 0.214 | -0.546** | 0.227 | 1.000 | -0.095 | -0.075 | 0.149 | |
新窗口打开|下载CSV
此外, 为验证结构参数的提取精度, 在6个具有单木位置的样方中, 将基于LiDAR数据提取的单木树高与样方实测树高进行对比分析(表6)。结果表明, 点云数据可以较为准确地估算树高(R2 = 0.715-0.836, RMSE = 0.684-1.323), 同时估算精度与以往研究相比具有一致性, 如Véga和Durrieu (2011)估算的树高RMSE为1.83 m; Apostol等(2020)提取挪威云杉(Picea abies)和山毛榉(Fagus longipetiolata)树高的均方根误差分别为1.09和1.17 m。
Table 6
表6
表6基于激光雷达提取的单木树高精度验证结果
Table 6
| 单木样方 Individual tree plot | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| R2 | 0.784 | 0.836 | 0.741 | 0.774 | 0.715 | 0.785 |
| RMSE | 1.196 | 0.894 | 0.684 | 1.083 | 1.323 | 1.132 |
新窗口打开|下载CSV
2.4 基于单木的聚类结果
基于单木的最优植被指数和最优结构参数, 采用自适应模糊C均值聚类算法, 在研究区内按照样方大小20 m × 20 m的窗口进行聚类, 并利用34个样方的物种丰富度和多样性指数实测值对森林乔木多样性估算结果进行精度验证。如表7所示, 物种丰富度的预测值明显低于实测值(R2 = 0.29, RMSE = 7.75), 当选择样方内树种的个体数大于1 (R2 = 0.37, RMSE = 2.41)和大于2 (R2 = 0.56, RMSE = 1.82)的树种参与实际物种丰富度的计算时, 精度有明显提高。Shannon-Wiener指数(R2 = 0.83, RMSE = 0.22)和Simpson指数(R2 = 0.85, RMSE = 0.09)与物种丰富度相比, 预测结果更好(图6)。Table 7
表7
表7古田山研究区物种丰富度预测及精度验证结果
Table 7
| 样方号 Plot No. | 实测物种丰富度 Measured species richness | 实测物种 丰富度* Measured species richness* | 实测物种 丰富度** Measured species richness** | 聚类数 (预测值) Prediction | 样方号 Plot No. | 实测物种丰富度 Measured species richness | 实测物种 丰富度* Measured species richness* | 实测物种 丰富度** Measured species richness** | 聚类数 (预测值) Prediction |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 14 | 9 | 8 | 10 | 18 | 13 | 7 | 6 | 8 |
| 2 | 10 | 6 | 5 | 8 | 19 | 19 | 10 | 6 | 9 |
| 3 | 17 | 9 | 9 | 8 | 20 | 17 | 11 | 8 | 8 |
| 4 | 9 | 6 | 5 | 7 | 21 | 16 | 10 | 6 | 8 |
| 5 | 11 | 9 | 7 | 8 | 22 | 16 | 10 | 9 | 7 |
| 6 | 20 | 14 | 11 | 13 | 23 | 8 | 5 | 5 | 5 |
| 7 | 19 | 14 | 10 | 11 | 24 | 9 | 8 | 5 | 5 |
| 8 | 11 | 6 | 5 | 8 | 25 | 13 | 6 | 3 | 6 |
| 9 | 6 | 4 | 3 | 6 | 26 | 13 | 9 | 7 | 7 |
| 10 | 14 | 8 | 5 | 7 | 27 | 14 | 6 | 4 | 5 |
| 11 | 18 | 9 | 6 | 6 | 28 | 7 | 5 | 5 | 7 |
| 12 | 10 | 7 | 6 | 9 | 29 | 16 | 8 | 5 | 7 |
| 13 | 16 | 7 | 5 | 5 | 30 | 20 | 12 | 8 | 8 |
| 14 | 16 | 10 | 8 | 8 | 31 | 15 | 7 | 6 | 8 |
| 15 | 20 | 12 | 8 | 9 | 32 | 16 | 8 | 7 | 9 |
| 16 | 18 | 14 | 8 | 8 | 33 | 15 | 8 | 3 | 5 |
| 17 | 16 | 8 | 5 | 6 | 34 | 17 | 13 | 9 | 8 |
| R2 | 0.29 | 0.37 | 0.56 | RMSE | 7.75 | 2.41 | 1.82 |
新窗口打开|下载CSV
图6
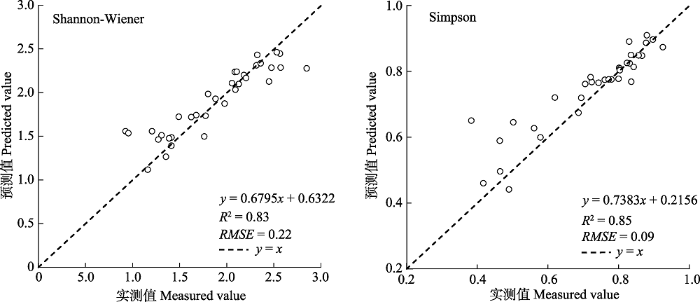 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6古田山研究区Shannon-Wiener指数和Simpson指数结果精度验证。
Fig. 6Validation result of Shannon-Wiener index and Simpson index in Gutianshan study area.
将研究区的物种丰富度和物种多样性估测结果按照直方图分级显示, 由图7可知, 三类指数的空间分布具有一致性: 中部和北部地区的山脊、山坡地带物种多样性较低, 主要由于该区域海拔在1 000 m以上, 森林类型是以黄山松为主的针阔混交林和针叶林, 环境条件限制了阔叶林的发育, 物种较为单一; 东南部和西南部地势较低、地形平坦, 多为耕地和以油茶林等小乔木为主的经济林, 建模时不作为森林类型考虑, 故物种多样性呈现零值和较低值。结合该地区的海拔信息, 物种丰富度和多样性随着海拔高度的增加整体上呈现出先增加后减小的特点。
图7
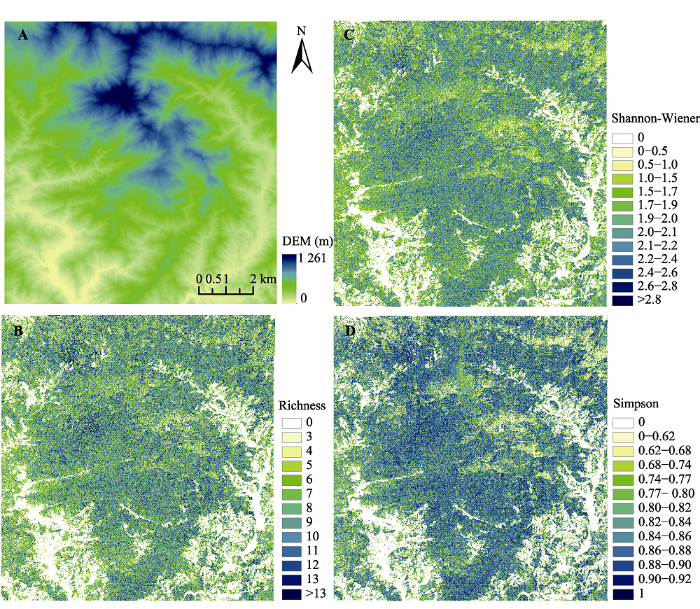 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7古田山研究区数字高程模型(DEM)(A)、物种丰富度(B)、Shannon-Wiener指数(C)和Simpson指数(D)分布图。
Fig. 7Digital Elevation Model (DEM)(A), species richness (B), Shannon-Wiener index (C) and Simpson index (D) mapping in Gutianshan study area.
3 讨论
本研究与刘鲁霞等(2019)在同一研究区基于高分2号数据采用随机森林(Random Forest, RF)方法进行Simpson (R2 = 0.53, RMSE = 0.042)和Shannon-Wiener指数(R2 = 0.53, RMSE = 0.30)的估算相比, 精度明显更高, 主要原因在于方法和数据的差异。其中, 随机森林是普遍适用的基于决策树的分类方法, 按照变量的重要性筛选最优特征, 但并未考虑森林物种多样性的成因和机理。而本研究基于生化-光谱-物种多样性的关联寻找最优聚类参数, 更符合植物生理特性。此外, 本研究使用的高光谱数据相比高分数据对于地物的光谱细节表达更加敏感, 有助于捕捉植被的生化特征信息, 同时LiDAR数据提供了树冠的三维信息, 在垂直维度上补充了丰富的结构特征(庞勇等, 2005; Coops et al., 2016)。本研究与传统的样地调查结果相比具有一致性。目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查。例如, 胡正华和于明坚(2006)应用相邻格子样方法调查了该地区16个处于不同演替阶段典型样地的乔木层Shannon- Wiener和Simpson多样性指数, 阔叶林分别为2.44和0.85、针叶林分别为1.97和0.76、针阔混交林分别为2.50和0.88, 而本研究基于ChinaCover数据集的植被类型计算得到的Shannon-Wiener和Simpson指数的平均值阔叶林分别为2.45和0.91、针叶林分别为2.32和0.85、针阔混交林分别为2.47和0.87, 与样方调查结果相符。森林乔木物种丰富度的变异往往与环境因子密切相关, 研究表明海拔、坡度等地形因子与物种分布和多样性之间具有明显关联(Palmer & Dixon, 1990)。翁昌露等(2019)在该地区网格化布设了79个样方研究10种森林群落类型的多样性指数分布格局及其影响因素, 发现随着海拔升高, 以Shannon-Wiener指数为代表的α多样性逐渐增大, 本研究中Shannon-Wiener指数分级图也呈现出相同趋势。
结构参数能否应用于森林物种多样性监测是当前研究讨论的热点。已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(Mayfield & Levine, 2010; Spasojevic & Suding, 2012)。董文雪等(2018)选取了95%分位数高度、冠层盖度和植被穿透率3个结构参数, 与Shannon-Wiener指数的相关性为R2 = 0.39-0.42, p < 0.01; 赵玉金(2016)的研究中, 植被穿透率、平均绝对偏差和偏度3个结构参数与Shannon-Wiener指数的相关性为R2 = 0.46-0.53, p < 0.05, 结合偏度、植被穿透率和平均植被高度的Shannon-Wiener指数的逐步回归模型预测精度为 R2 = 0.55, RMSE = 0.40。以上研究表明, 在森林类型复杂的地区, 结构参数与物种多样性指数的相关性普遍不高, 预测能力也受到多种因素制约。本研究选择的95%分位数高度和偏度对于Simpson指数的相关性R2分别为0.46和0.17, 逐步回归模型的预测精度R2 = 0.32, p < 0.01, 该逐步回归方程同样表明仅利用结构参数对物种多样性指数的模拟能力有所欠缺, 但其主要目的是在众多结构参数中选取与物种多样性相关性最高且能够代表其他结构参数的最优结构参数用于多样性监测。
为进一步探讨不同类型的特征参数在提升聚类模型预测能力上的作用, 对分别考虑光谱差异和结构特征的多样性监测研究进行对比分析。Shi等(2018)发现将植物功能性状与光谱和结构指标共同用于树种分类时的总体精度(83.7%)明显高于仅利用结构(65.1%)或光谱(69.3%)指标的分类精度; Zhao等(2018)结合光谱指数和结构参数估算的乔木物种多样性(R2 = 0.83, RMSE = 0.25)比仅利用光谱信息(R2 = 0.60, RMSE = 0.38)的估算结果精度更高。本研究前期探讨了仅利用6个植被指数参与聚类分析时物种丰富度(R2 = 0.18, RMSE = 3.08)、Shannon- Wiener指数(R2 = 0.61, RMSE = 0.34)和 Simpson指数(R2 = 0.61, RMSE = 0.11)的预测精度, 发现在引入树高和偏度两个结构参数后, 3个指数的预测精度均有明显提高, 与前人研究的结论具有一致性, 说明综合考虑光谱特征和结构特征后, 模型对于物种多样性的预测能力有显著提升。
聚类分析是根据数据相似性进行分组的无监督学习过程, 通过聚类有效性函数将样本自然划分成不同的簇。利用聚类分析开展物种多样性监测的最大优势在于无需先验知识指导, 并且通过改进的聚类算法甚至可不预先定义类别数。该方法的缺点是聚类过程在黑箱中进行, 无法给出具体的聚类模型表达形式, 因而无法得出每个参数对于聚类结果的影响程度。本研究样本所选择的属性信息, 如植被指数、结构参数共同参与聚类过程, 其中, 植被指数反映着调控物种生命活动的生理化学根源, 结构参数反映着物种的外在形态, 从不同角度提供了物种识别特征, 并可弥补由光谱饱和带来的物种饱和问题, 进一步增加了聚类中心的差异性, 有助于提升模型的聚类能力。本研究因获得了高精度LiDAR数据, 使单木树冠的获取成为可能, 故可在单木尺度上开展物种多样性监测, 这也与传统的基于样方每木调查获得的物种多样性含义一致。以往利用遥感手段开展的多样性监测研究由于LiDAR数据获取难度大、成本高, 因此多聚焦在像元尺度上。LiDAR数据的引入将多样性的估算从像元层次上升至生物个体层次, 通过对比基于像元与基于单木尺度的多样性聚类模型, 可进一步探讨聚类方法在不同场景生物多样性监测的适用性。
4 结论
本研究在古田山自然保护区探讨了基于遥感数据利用聚类技术开展中亚热带森林乔木物种多样性监测的方法。利用高光谱和激光雷达数据提取的特征参数作为自适应模糊C均值聚类算法的输入参数, 直接估算单木尺度的乔木物种类别差异, 无需判定具体的树种。本研究与基于随机森林方法的监测相比在精度上具有优势, 与基于样方实测物种多样性的结果相比, 在物种多样性指数的量级和变化趋势上具有一致性。该研究整体上基于植物的生化调控机制, 相比利用光谱变换提取特征信息的物种多样性监测, 机理更明确, 能充分体现叶片生化、冠层光谱和物种多样性之间的内在联系。然而, 由于森林物种多样性的影响因素极其复杂, 使得对生化-光谱-物种多样性机制的解译仍然存在较大难度。另外, 考虑生境过滤机制对多样性格局的作用, 在后续研究中还可引入纹理信息和海拔等地形因子, 进一步完善森林物种特征识别库, 从多角度探究森林物种的识别标签, 发展新的森林物种多样性指数, 并深入研究适用于不同生态情境的物种多样性估算模型。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/08-0023.1URLPMID:19323186 [本文引用: 1]

Leaf chemical and spectral properties of 162 canopy species were measured at 11 tropical forest sites along a 6024 mm precipitation/yr and 8.7 degrees C climate gradient in Queensland, Australia. We found that variations in foliar nitrogen, phosphorus, chlorophyll a and b, and carotenoid concentrations, as well as specific leaf area (SLA), were expressed more strongly among species within a site than along the entire climate gradient. Integrated chemical signatures consisting of all leaf properties did not aggregate well at the genus or family levels. Leaf chemical diversity was maximal in the lowland tropical forest sites with the highest temperatures and moderate precipitation levels. Cooler and wetter montane tropical forests contained species with measurably lower variation in their chemical signatures. Foliar optical properties measured from 400 to 2500 nm were also highly diverse at the species level, and were well correlated with an ensemble of leaf chemical properties and SLA (r2 = 0.54-0.83). A probabilistic diversity model amplified the leaf chemical differences among species, revealing that lowland tropical forests maintain a chemical diversity per unit richness far greater than that of higher elevation forests in Australia. Modeled patterns in spectral diversity and species richness paralleled those of chemical diversity, demonstrating a linkage between the taxonomic and remotely sensed properties of tropical forest canopies. We conclude that species are the taxonomic unit causing chemical variance in Australian tropical forest canopies, and thus ecological and remote sensing studies should consider the role that species play in defining the functional properties of these forests.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1371/journal.pbio.0040277URLPMID:16895442 [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
DOI:10.1016/0034-4257(92)90059-SURL [本文引用: 2]
DOI:10.1562/0031-8655(2002)075<0272:accipl>2.0.co;2URLPMID:11950093 [本文引用: 2]
Spectral reflectance of maple, chestnut and beech leaves in a wide range of pigment content and composition was investigated to devise a nondestructive technique for total carotenoid (Car) content estimation in higher plant leaves. Reciprocal reflectance in the range 510 to 550 nm was found to be closely related to the total pigment content in leaves. The sensitivity of reciprocal reflectance to Car content was maximal in a spectral range around 510 nm; however, chlorophylls (Chl) also affect reflectance in this spectral range. To remove the Chl effect on the reciprocal reflectance at 510 nm, a reciprocal reflectance at either 550 or 700 nm was used, which was linearly proportional to the Chl content. Indices for nondestructive estimation of Car content in leaves were devised and validated. Reflectances in three spectral bands, 510+/-5 nm, either 550+/-15 nm or 700+/-7.5 nm and the near infrared range above 750 nm are sufficient to estimate total Car content in plant leaves nondestructively with a root mean square error of less than 1.75 nmol/cm2.
DOI:10.1016/0168-1923(95)02268-6URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:19341137 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-3040.2005.01493.xURLPMID:17080588 [本文引用: 1]
The yield potential (Yp) of a grain crop is the seed mass per unit ground area obtained under optimum growing conditions without weeds, pests and diseases. It is determined by the product of the available light energy and by the genetically determined properties: efficiency of light capture (epsilon i), the efficiency of conversion of the intercepted light into biomass (epsilon c) and the proportion of biomass partitioned into grain (eta). Plant breeding brings eta7 and epsilon i close to their theoretical maxima, leaving epsilon c, primarily determined by photosynthesis, as the only remaining major prospect for improving Yp. Leaf photosynthetic rate, however, is poorly correlated with yield when different genotypes of a crop species are compared. This led to the viewpoint that improvement of leaf photosynthesis has little value for improving Yp. By contrast, the many recent experiments that compare the growth of a genotype in current and future projected elevated [CO2] environments show that increase in leaf photosynthesis is closely associated with similar increases in yield. Are there opportunities to achieve similar increases by genetic manipulation? Six potential routes of increasing epsilon c by improving photosynthetic efficiency were explored, ranging from altered canopy architecture to improved regeneration of the acceptor molecule for CO2. Collectively, these changes could improve epsilon c and, therefore, Y p by c. 50%. Because some changes could be achieved by transgenic technology, the time of the development of commercial cultivars could be considerably less than by conventional breeding and potentially, within 10-15 years.
DOI:10.1016/j.rse.2015.11.029URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
URLPMID:20576030 [本文引用: 1]
[本文引用: 1]
DOI:10.17520/biods.2015313URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.isprsjprs.2005.05.002URL [本文引用: 1]
AbstractUnderstanding your data through exploratory data analysis is a necessary first stage of data analysis particularly for observational data. The checking of data integrity and understanding the distributions, correlations and relationships between potentially important variables is a fundamental part of the analysis process prior to model development and hypothesis testing. In this paper, exploratory data analysis is used to assess the potential of laser return type and return intensity as variables for classification of individual trees or forest stands according to species.For narrow footprint lidar instruments that record up to two return amplitudes for each output pulse, the usual pre-classification of return data into first and last intensity returns camouflages the fact that a number of the return signals have only “single amplitude” (singular) returns. The importance of singular returns for species discrimination has received little discussion in the remote sensing literature. A map view of the different types of returns overlaid on field species data indicated that it is possible to visually distinguish between vegetation types that produce a high proportion of singular returns, compared to vegetation types that produce a lower proportion of singular returns, at least when using a specific laser footprint size. Using lidar data and the corresponding field data derived from a subtropical woodland area of South East Queensland, Australia, map scatterplots of return types combined with field data enabled, in some cases, visual discrimination at the individual tree level between White Cypress Pine (Callitris glaucophylla) and Poplar Box (Eucalyptus populnea). While a clear distinction between these two species was not always visually obvious at the individual tree level, due to other extraneous sources of variation in the dataset, the observation was supported in general at the site level. Sites dominated by Poplar Box generally exhibited a lower proportion of singular returns compared to sites dominated by Cypress Pine.While return intensity statistics for this particular dataset were not found to be as useful for classification as the proportions of laser return types, an examination of the return intensity data leads to an explanation of how return intensity statistics are affected by forest structure. Exploratory data analysis indicated that a large component of variation in the intensity of the return signals from a forest canopy is associated with reflections of only part of the laser footprint. Consequently, intensity return statistics for the forest canopy, such as average and standard deviation, are related not only to the reflective properties of the vegetation, but also to the larger scale properties of the forest such as canopy openness and the spacing and type of foliage components within individual tree crowns.]]>
DOI:10.1038/35002501URLPMID:10706275 [本文引用: 1]
Conservationists are far from able to assist all species under threat, if only for lack of funding. This places a premium on priorities: how can we support the most species at the least cost? One way is to identify 'biodiversity hotspots' where exceptional concentrations of endemic species are undergoing exceptional loss of habitat. As many as 44% of all species of vascular plants and 35% of all species in four vertebrate groups are confined to 25 hotspots comprising only 1.4% of the land surface of the Earth. This opens the way for a 'silver bullet' strategy on the part of conservation planners, focusing on these hotspots in proportion to their share of the world's species at risk.
[本文引用: 1]
[本文引用: 1]
DOI:10.2307/3236053URL [本文引用: 1]
DOI:10.11707/j.1001-7488.20050322URL [本文引用: 1]
Lidar (light detection and ranging) remote sensing is a breakthrough of active remote sensing technology in recent years. It has shown enormous potential for forest parameters retrieval. Lidar remote sensing has the unique advantage of providing horizontal and vertical information at high accuracies. Especially it can be used to measure forest height directly with unprecedented accuracy. This article introduces the basic principle of lidar remote sensing and the lidar systems of large footprint and small footprint. The status of the technology, the retrieval methods of forest parameters such as tree height, biomass, canopy vertical structure and crown closure, and the prospect of the application of lidar technology in forestry have been reviewed in detail in this paper.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1523-1739.2012.01869.xURLPMID:22731687 [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.tree.2008.04.009URLPMID:18582987 [本文引用: 1]
Tropical forests are renowned for their biological diversity, but also harbor variable combinations of soil age, chemistry and susceptibility to erosion or tectonic uplift. Here we contend that the combined effects of this biotic and abiotic diversity promote exceptional biogeochemical heterogeneity at multiple scales. At local levels, high plant diversity creates variation in chemical and structural traits that affect plant production, decomposition and nutrient cycling. At regional levels, myriad combinations of soil age, soil chemistry and landscape dynamics create variation and uncertainty in limiting nutrients that do not exist at higher latitudes. The effects of such heterogeneity are not well captured in large-scale estimates of tropical ecosystem function, but we suggest new developments in remote sensing can help bridge the gap.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.rse.2019.111218URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nature02403URLPMID:15103368 [本文引用: 1]
Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Object- based land cover classification using airborne LiDAR
1
2008
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
Species discrimination and individual tree detection for predicting main dendrometric characteristics in mixed temperate forests by use of airborne laser scanning and ultra-high-resolution imagery
1
2020
... 此外, 为验证结构参数的提取精度, 在6个具有单木位置的样方中, 将基于LiDAR数据提取的单木树高与样方实测树高进行对比分析(
Spectral and chemical analysis of tropical forests: scaling from leaf to canopy levels
1
2008
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Airborne spectranomics: mapping canopy chemical and taxonomic diversity in tropical forests
1
2009
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Leaf chemical and spectral diversity in Australian tropical forests
1
2009
... 在16个常用生化组分中选取能够表征森林物种的最优生化组分.首先基于叶片实测光谱利用偏最小二乘回归(Partial Least Squares Regression, PLSR)方法预测16种叶片生化组分含量, 通过与实测叶片生化组分含量的对比分析选取相关性较高的生化组分作为最优生化组分.偏最小二乘回归方法是一种多对多的线性回归方法, 汇集了主成分分析、典型相关分析和多元线性回归分析的优点, 能够在生化组分少、光谱波段多且相关性强的情况下建立回归模型, 可通过JMP 11.0软件实现.随后, 将预测的与实测的叶片生化组分含量进行对比分析, 选择相关性相对高的作为最优生化组分, 同时还可以借鉴前人研究的分析结果, 如
Hyperspectral remote sensing of canopy biodiversity in Hawaiian lowland rainforests
1
2007
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
1
2017
... 基于地面观测的样方单木调查记录和文献资 料, 根据森林树种的代表性和分布范围, 选择在数量上占优势、出现频率高且分布均匀的17个树种作为研究区的优势树种(
1
2017
... 基于地面观测的样方单木调查记录和文献资 料, 根据森林树种的代表性和分布范围, 选择在数量上占优势、出现频率高且分布均匀的17个树种作为研究区的优势树种(
The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest
1
2008
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
A forest structure habitat index based on airborne laser scanning data
2
2016
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
... 本研究与
Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance
2
2000
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
Biodiversity loss threatens human well-being
1
2006
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
机载激光雷达及高光谱的森林乔木物种多样性遥感监测
2
2018
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
机载激光雷达及高光谱的森林乔木物种多样性遥感监测
2
2018
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
Development of a large area biodiversity monitoring system driven by remote sensing
1
2007
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
Ground-based remote sensing for assessing water and nitrogen status of broccoli
2
2007
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency
2
1992
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
Assessing carotenoid content in plant leaves with reflectance spectroscopy
2
2002
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
Remote sensing of net primary production in boreal forest stands
1
1996
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Linkages between biodiversity attributes and ecosystem services: a systematic review
1
2014
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
浙江古田山常绿阔叶林演替序列研究: 群落物种多样性
1
2006
... 本研究与传统的样地调查结果相比具有一致性.目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查.例如,
浙江古田山常绿阔叶林演替序列研究: 群落物种多样性
1
2006
... 本研究与传统的样地调查结果相比具有一致性.目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查.例如,
Derivation of leaf-area index from quality of light on the forest floor
2
1969
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
From space to species: ecological applications for remote sensing
1
2003
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
Detection of individual tree crowns in airborne lidar data
1
2006
... 34个样方单木分离估算结果与实测单木数量的对比分析如
Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies
1
2009
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
Lidar remote sensing of aboveground biomass in three biomes
1
2002
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
Partitioning beta diversity in a subtropical broad-leaved forest of China
1
2009
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
A new validity function for fuzzy clustering
1
2009
... 自适应模糊C均值聚类算法(Self-adaptive Fuzzy C-Means clustering algorithm)发展于模糊C均值聚类, 通过运用有效性函数来自动获取聚类数目, 无需依赖预先设定的类别数, 解决了聚类算法对于初始值的敏感问题.
线性回归模型估算水稻叶片叶绿素含量的适宜性分析
1
2003
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
线性回归模型估算水稻叶片叶绿素含量的适宜性分析
1
2003
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
LiDAR remote sensing of forest structure
1
2003
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
基于高分2号遥感数据估测中亚热带天然林木本植物物种多样性
1
2019
... 本研究与
基于高分2号遥感数据估测中亚热带天然林木本植物物种多样性
1
2019
... 本研究与
Can improvement in photosynthesis increase crop yields?
1
2006
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
Comparing generalized linear models and random forest to model vascular plant species richness using LiDAR data in a natural forest in central Chile
1
2016
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
中国生物多样性监测网络建设:从CForBio到Sino BON
1
2015
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
中国生物多样性监测网络建设:从CForBio到Sino BON
1
2015
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
Opposing effects of competitive exclusion on the phylogenetic structure of communities
1
2010
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
Nitrogen and lignin control of hardwood leaf litter decomposition dynamics
1
1982
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
中国森林生物多样性监测: 科学基础与执行计划
1
2016
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
中国森林生物多样性监测: 科学基础与执行计划
1
2016
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
Airborne laser scanning: exploratory data analysis indicates potential variables for classification of individual trees or forest stands according to species
1
2005
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
Biodiversity hotspots for conservation priorities
1
2000
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
Estimating tree height and tree crown properties using airborne scanning laser in a boreal nature reserve
1
2002
... 34个样方单木分离估算结果与实测单木数量的对比分析如
Estimating forest biomass and volume using airborne laser data
1
1988
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
Small-scale environmental heterogeneity and the analysis of species distributions along gradients
1
1990
... 本研究与传统的样地调查结果相比具有一致性.目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查.例如,
激光雷达技术及其在林业上的应用
1
2005
... 本研究与
激光雷达技术及其在林业上的应用
1
2005
... 本研究与
LiCHy: the CAF’s LiDAR, CCD and hyperspectral integrated airborne observation system
1
2016
... 机载LiDAR及高光谱数据于2017年10月由搭载在运5飞机上的LiCHy系统获取(
The reflectance at the 950-970 nm region as an indicator of plant water status
2
1993
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
Ecological Diversity
1
1975
... 式中, pi表示含有n个物种的群落中第i个物种个体数占所有物种总个体数的比例(
古田山自然保护区阔叶林与两种人工林的群落结构和生物多样性
1
2018
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
古田山自然保护区阔叶林与两种人工林的群落结构和生物多样性
1
2018
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
Estimation of leaf area index and covered ground from airborne laser scanner (Lidar) in two contrasting forests
1
2004
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
Measuring β-diversity by remote sensing: a challenge for biodiversity monitoring
1
2018
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
Estimating terrestrial primary productivity by combining remote sensing and ecosystem simulation
1
1990
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Smoothing and differentiation of data by simplified least squares procedures
1
1964
... 在实验室利用标准型地物光谱仪(FieldSpec 4 Std-Res, Analytica Spectra Devices, Colorado, USA)对地面同步采集的17个优势树种的叶片分别进行光谱测定, 每个树种随机选择10个叶片计算平均值(
Tree species classification using plant functional traits from LiDAR and hyperspectral data
1
2018
... 为进一步探讨不同类型的特征参数在提升聚类模型预测能力上的作用, 对分别考虑光谱差异和结构特征的多样性监测研究进行对比分析.
Use of an airborne lidar system to model plant species composition and diversity of mediterranean oak forests
1
2012
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
Inferring community assembly mechanisms from functional diversity patterns: the importance of multiple assembly processes
1
2012
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
The biogeochemical heterogeneity of tropical forests
1
2008
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Remote sensing for biodiversity science and conservation
1
2003
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
Using imaging spectroscopy to study ecosystem processes and properties
1
2004
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin
1
1963
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
Multi-level filtering segmentation to measure individual tree parameters based on Lidar data: application to a mountainous forest with heterogeneous stands
1
2011
... 此外, 为验证结构参数的提取精度, 在6个具有单木位置的样方中, 将基于LiDAR数据提取的单木树高与样方实测树高进行对比分析(
利用光谱反射率估算叶片生化组分和籽粒品质指标研究
1
2003
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
利用光谱反射率估算叶片生化组分和籽粒品质指标研究
1
2003
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Remote sensing of terrestrial plant biodiversity
1
2019
... 激光雷达(LiDAR)通过发射高频脉冲穿透冠层从而提供森林的三维空间信息, 获取的点云数据可提取树高、叶面积指数、冠幅等森林结构参数(
The global fuzzy C-means clustering algorithm
1
2006
... 聚类分析通过对数据集合进行合理分割以确定类别数, 使同一类中数据尽可能相近, 不同类之间数据尽可能相异(
古田山10种主要森林群落类型的α和β多样性格局及影响因素
1
2019
... 本研究与传统的样地调查结果相比具有一致性.目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查.例如,
古田山10种主要森林群落类型的α和β多样性格局及影响因素
1
2019
... 本研究与传统的样地调查结果相比具有一致性.目前生态学上对森林物种多样性的调查是通过布设样方逐株调查的方式来获得物种数量和分布情况, 已有研究利用样方法对古田山地区物种多样性指数及其变异规律等进行了调查.例如,
The worldwide leaf economics spectrum
1
2004
... 利用高枝剪收集完全光照条件下的冠层叶片将其分为两组.其中一组每个优势树种采集约200枚叶片, 利用基于色素吸收峰的分光光度法测定叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量, 这些生化组分是植被光合作用能力、生理胁迫状况、固碳能力及氮利用效率的良好指示器(
1
2017
... 此外, 本研究区的土地覆被数据来自30 m空间分辨率的中国土地覆被数据集(ChinaCover).该数据集以国产环境卫星HJ数据和美国陆地卫星Landsat数据为数据源, 依据在FAO LCCS分类工具支持下定义的适合中国土地覆盖类型特点与应用需求的分类系统, 将中国土地覆被分为森林、草地、耕地、湿地、人工表面及其他6个一级类和40个二级类(
Estimating chlorophyll content from hyperspectral vegetation indices: modeling and validation
2
2008
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
... Vegetation indices corresponding to the optimal biochemical components
| 生化组分 Biochemical component | 植被指数 Vegetation index | 计算公式 Formula | 引用 Reference |
|---|---|---|---|
| 叶绿素a/b Chl a/b | TCARI/OSAVI | TCARI/OSAVI [705,750] = 3[(R750.66 - R704.6) - 0.2(R750.66 - R550.67)(R750.66/R704.6)]/[(1 + 0.16)(R750.66 - R704.6)/(R750.66 + R704.6 + 0.16)] | |
| 类胡萝卜素 Car | CRI | CRI = 1/R510 - 1/R550 | |
| 水分 EWT | WBI | WBI = R895/R972 | |
| 氮/磷 N/P | CCCI | CCCI = (0.7415R790 - 0.6965R720)/(0.0319R790 - 0.281R720) | |
| 比叶面积 SLA | RVI | RVI = R750/R705 | |
| 纤维素/木质素 Cel/Lig | PRI | PRI = (R531 - R570)/(R531 + R570) |
浙江古田山自然保护区森林植被类型
2
2001
... 本研究以古田山保护区的核心地带作为研究 区, 地理坐标为118.08°-118.18° E, 29.19°-29.28° N, 总面积约104 km2 (
... 基于地面观测的样方单木调查记录和文献资 料, 根据森林树种的代表性和分布范围, 选择在数量上占优势、出现频率高且分布均匀的17个树种作为研究区的优势树种(
浙江古田山自然保护区森林植被类型
2
2001
... 本研究以古田山保护区的核心地带作为研究 区, 地理坐标为118.08°-118.18° E, 29.19°-29.28° N, 总面积约104 km2 (
... 基于地面观测的样方单木调查记录和文献资 料, 根据森林树种的代表性和分布范围, 选择在数量上占优势、出现频率高且分布均匀的17个树种作为研究区的优势树种(
Scaling-up and model inversion methods with narrowband optical indices for chlorophyll content estimation in closed forest canopies with hyperspectral data
1
2001
... 确定最优生化组分后, 可通过已有的植被指数模型选择与之相对应的植被指数.其中, Chl a和Chl b可利用转换型吸收反射率指数(TCARI)和优化型土壤调整植被指数(OSAVI)的比值来表示(
Isolating individual trees in a closed coniferous forest using small footprint lidar data
2
2014
... 以基于LiDAR数据生成的经过无效值填充的冠层高度模型CHM为基础, 利用结合形态学冠层控制的分水岭算法实现单木的分离(
... 34个样方单木分离估算结果与实测单木数量的对比分析如
Filling invalid values in a lidar-derived canopy height model with morphological crown control
2
2013
... LiDAR数据的预处理依据国家测绘地理信息局发布的《机载激光雷达数据处理技术规范》和《机载激光雷达数据后处理技术规范》进行, 利用TerraSolid软件将点云自动分为地面点(Max building size = 60, Iteration angle = 6, Iteration distance = 1.4)和植被点(除地面及建筑物), 随后由点云的首次回波点插值生成数字表面模型(DSM), 再由点云中的地面点通过构建不规则三角网(TIN)插值生成数字高程模型(DEM), 最后由两者通过差值运算得到冠层高度模型(CHM)(
... 以基于LiDAR数据生成的经过无效值填充的冠层高度模型CHM为基础, 利用结合形态学冠层控制的分水岭算法实现单木的分离(
基于高光谱和LiDAR的森林冠层物种多样性遥感监测方法研究
2
2016
... 在16个常用生化组分中选取能够表征森林物种的最优生化组分.首先基于叶片实测光谱利用偏最小二乘回归(Partial Least Squares Regression, PLSR)方法预测16种叶片生化组分含量, 通过与实测叶片生化组分含量的对比分析选取相关性较高的生化组分作为最优生化组分.偏最小二乘回归方法是一种多对多的线性回归方法, 汇集了主成分分析、典型相关分析和多元线性回归分析的优点, 能够在生化组分少、光谱波段多且相关性强的情况下建立回归模型, 可通过JMP 11.0软件实现.随后, 将预测的与实测的叶片生化组分含量进行对比分析, 选择相关性相对高的作为最优生化组分, 同时还可以借鉴前人研究的分析结果, 如
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
基于高光谱和LiDAR的森林冠层物种多样性遥感监测方法研究
2
2016
... 在16个常用生化组分中选取能够表征森林物种的最优生化组分.首先基于叶片实测光谱利用偏最小二乘回归(Partial Least Squares Regression, PLSR)方法预测16种叶片生化组分含量, 通过与实测叶片生化组分含量的对比分析选取相关性较高的生化组分作为最优生化组分.偏最小二乘回归方法是一种多对多的线性回归方法, 汇集了主成分分析、典型相关分析和多元线性回归分析的优点, 能够在生化组分少、光谱波段多且相关性强的情况下建立回归模型, 可通过JMP 11.0软件实现.随后, 将预测的与实测的叶片生化组分含量进行对比分析, 选择相关性相对高的作为最优生化组分, 同时还可以借鉴前人研究的分析结果, 如
... 结构参数能否应用于森林物种多样性监测是当前研究讨论的热点.已有研究表明结构参数能够在一定程度上反映物种多样性, 但是从森林三维特征出发的结构多样性, 由于环境异质性等因素的影响, 同一树种的树冠结构在不同生境下存在差异, 使得结构参数难以完全反映复杂森林类型的多样性特 点, 因而不能作为区分物种的决定因素(
The optimal leaf biochemical selection for mapping species diversity based on imaging spectroscopy
1
2016
... 生化组分是植物生理过程的关键决定因子, 影响着植被的生理功能和表征性状, 与植被的生长、发育、维持和防御有关(
Forest species diversity mapping using airborne LiDAR and hyperspectral data in a subtropical forest in China
3
2018
... 聚类分析可以搜索并识别一个有限的数据集 合, 发现数据的全局分布模式, 聚类方法的基础在于能被聚为一类的各样本具有相似的属性集合.对于森林物种来说, 同一物种的光谱由于具有本质相同的生化组分而具有高度相似性, 在区域分布上同一物种还在树高、冠层结构等方面表现出相近性, 这种一致使得某一物种呈现出能够被遥感识别的有异于其他物种的特征, 从而可直接估算物种类别差异, 无需判定具体的树种属性.因此, 在冠层尺度如获取了与物种多样性相关的生化、光谱和结构参数, 将单木个体作为最小单元并通过聚类分析, 可估算物种类别的差异度, 即为物种丰富度.
... Canopy structural parameters derived from LiDAR
| 结构参数 Structural parameter | 描述 Description | 引用 Reference |
|---|---|---|
| 95%分位数高度 95% quantile height | 近似于森林冠层峰值高度, 由首次回波统计获得 Approximates the peak height in meters of the forest canopy, obtained from the point cloud of the first echo | |
| 平均植被高度 Mean vegetation height | 植被的平均高度, 由各植被分层首次回波和末次回波统计获得 The mean height of vegetation, obtained from the point cloud of the first and last echo of each vegetation layer | |
| 植被穿透率 Vegetation permeability | 植被首次回波在二次回波中的比例, 由植被的首次回波和所有二次回波计算得到 Proportion of first vegetation returns (see above) for which there is a second return, obtained from the point cloud of the first echo and all secondary echoes of vegetation | |
| 叶高度多样性 Foliage height diversity | 描述植被剖面的叶密度和高度分布, 公式: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, 其中, pi表示不同高度间隔内的点云返回值与所有返回值的比例, h表示树高 Metric intended to characterize the density and height distribution of foliage in a vegetation profile. Formula: $h=-\mathop{\sum }^{}{{p}_{i}}\ln {{p}_{i}}$, where pi is the ratio of return value of point cloud in different height intervals to all return values, h is the tree height | |
| 标准偏差(首次回波) Standard deviation (first return) | 反映单木树高的离散程度 Metric the dispersion of each individual tree | |
| 平均绝对偏差 Mean absolute deviation | 所有单木树高值与其算术平均值的偏差的绝对值的平均 Mean of absolute value of the deviation of tree height from the mean | |
| 偏度(首次回波) Skewness (first return) | 与峰态(首次回波)高度相关 Highly correlated with kurtosis |
... 为进一步探讨不同类型的特征参数在提升聚类模型预测能力上的作用, 对分别考虑光谱差异和结构特征的多样性监测研究进行对比分析.
生态红线的实践与思考
1
2014
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
生态红线的实践与思考
1
2014
... 森林生物多样性对维持森林生态系统服务、功能和稳定性具有重大作用(
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
1
2008
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
1
2008
... 研究区位于浙江省开化县古田山国家级自然保护区, 地理环境复杂, 自然资源丰富, 在低海拔地区保存有发育良好的天然常绿阔叶林地带性植被, 是亚热带生物多样性研究的绝佳场所, 许多****在该研究区针对物种组成、演替过程等方面开展过大量研究(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}