李帅锋
1 , 郎学东
1 , 黄小波
1 , 王艳红
1 , 刘万德
1 , 徐崇华
2 , 苏建荣
, , 1 , * 1 中国林业科学研究院资源昆虫研究所, 国家林业和草原局云南普洱森林生态系统国家定位观测研究站, 昆明 6502242 云南省太阳河自然保护区管理局, 云南普洱 665000Association classification of a 30 hm2 dynamics plot in the monsoon broad-leaved evergreen forest in Pu’er, Yunnan, China Shuai-Feng LI
1 , Xue-Dong LANG
1 , Xiao-Bo HUANG
1 , Yan-Hong WANG
1 , Wan-De LIU
1 , Chong-Hua XU
2 , Jian-Rong SU
, , 1 , * 1 Research Institute of Resource Insects, Chinese Academy of Forestry, The Pu?er Forest Ecosystem Research Station, National Forestry and Grassland Administration of China, Kunming 650224, China2 Taiyanghe Provincinal Nature Reserve, Pu’er, Yunnan 665000, China通讯作者: * 苏建荣,ORCID:0000-0001-5667-5670
jianrongsu@vip.sina.com 编委: 曹敏
责任编辑: 赵航
收稿日期: 2019-10-15
接受日期: 2020-01-7
网络出版日期: 2020-03-20
基金资助: 中央级公益性科研院所基本科研业务费专项资助 (CAFYBB2017ZX002-4 )林业公益性行业科研专项 (201404211 )云南省基础研究计划 (2019FB058 )
Corresponding authors: *
jianrongsu@vip.sina.com Received: 2019-10-15
Accepted: 2020-01-7
Online: 2020-03-20
Fund supported: Fundamental Research Funds of Chinese Academy of Forestry (CAFYBB2017ZX002-4 )Forestry Industry Research Special Funds for Public Welfare Projects (201404211 )Applied Basic Research Foundation of Yunnan Province (2019FB058 )
摘要 季风常绿阔叶林是我国南亚热带典型的地带性植被, 建立森林动态监测样地是研究生物多样性维持和群落构建机制的重要平台。该文以普洱30 hm
2 森林动态监测样地为研究对象, 采用多元回归树、重要值、主成分分析与指示种相结合的方法对样地内750个样方进行群丛数量分类, 以获取季风常绿阔叶林森林植被群丛类型。结果表明: 森林动态监测样地共发现木本植物271种, 隶属于78科178属, 群落类型为短刺锥(
Castanopsis echidnocarpa )群系, 可划分4个群丛类型, 分别是珍珠花+毛银柴-短刺锥+泥柯群丛(
Lyonia ovalifolia +
Aporosa villosa -
Castanopsis echidnocarpa +
Lithocarpus fenestratus Association)、云南瘿椒树+耳叶柯-短刺锥+西南木荷群丛(
Tapiscia yunnanensis +
Lithocarpus grandifolius -
Castanopsis echidnocarpa +
Schima wallichii Association)、大果杜英+黄药大头茶-短刺锥+西南木荷群丛(
Elaeocarpus sikkimensis +
Polyspora chrysandra -
Castanopsis echidnocarpa +
Schima wallichii Association)、西桦+尼泊尔桤木-短刺锥+枹丝锥群丛(
Betula alnoides +
Alnus nepalensis -
Castanopsis echidnocarpa +
Castanopsis calathiformis Association)。群丛间物种分布存在较多过渡重叠, 指示物种是区分群丛类型的主要依据, 海拔与坡向对群丛分类有较大的影响, 坡度对群丛分类影响较小。
关键词: 植被分类 ;
物种组成 ;
地形因子 ;
多元回归树 ;
重要值 ;
指示种 ;
季风常绿阔叶林 Abstract Aims The monsoon broad-leaved evergreen forest is a typical zonal vegetation of south subtropical China, where forest dynamics plot is a study platform of mechanisms of biodiversity maintenance and community assembly. Our objectives were to obtain plant association types in a 30 hm
2 forest dynamic monitoring plot in the Pu’er City, Yunnan Province.
Methods We identified typical association types by the methods of multivariate regression trees, importance value, principal component analysis and indicator species based on 750 subplots (20 m × 20 m), and analyzed the differences of topography factors and community characteristics among different plant associations.
Important findings The results showed that the specie richness of woody species (diameter at breast height (
DBH ) ≥ 1 cm) was 271, which belonged to 78 families and 178 genera in the forest dynamic plot. The community formation was Formation
Castanopsis echidnocarpa. Forest was divided into four associations: (1)
Lyonia ovalifolia +
Aporosa villosa -
Castanopsis echidnocarpa +
Lithocarpus fenestratus Association, (2)
Tapiscia yunnanensis +
Lithocarpus grandifolius -
Castanopsis echidnocarp +
Schima wallichii Association, (3)
Elaeocarpus sikkimensis +
Polyspora chrysandra -
Castanopsis echidnocarp +
Schima wallichii Association, and (4)
Betula alnoides +
Alnus nepalensis -
Castanopsis echidnocarp +
Castanopsis calathiformis Association. There were many transitional overlaps in the distribution of associations, while indicator species was the main reference for distinguishing the types of associations. In addition, altitude and slope aspect had a great influence on the plant association types, while slope
per se had a relatively weak impact on the association types.
Keywords: vegetation classification ;
species composition ;
topography factor ;
multivariate regression tree ;
importance value ;
indicator species ;
monsoon broad-leaved evergreen forest PDF (2025KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 李帅锋, 郎学东, 黄小波, 王艳红, 刘万德, 徐崇华, 苏建荣. 云南普洱30 hm
2 季风常绿阔叶林动态监测样地群丛数量分类 . 植物生态学报, 2020, 44(3): 236-247. DOI:
10.17521/cjpe.2019.0268 LI Shuai-Feng, LANG Xue-Dong, HUANG Xiao-Bo, WANG Yan-Hong, LIU Wan-De, XU Chong-Hua, SU Jian-Rong.
Association classification of a 30 hm2 dynamics plot in the monsoon broad-leaved evergreen forest in Pu’er, Yunnan, China .
Chinese Journal of Plant Ecology , 2020, 44(3): 236-247. DOI:
10.17521/cjpe.2019.0268 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 )。常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 )。植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 )。随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 )。近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局。
季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 )。云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 )。由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义。
随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 )。2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场。目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及。本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子。本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据。
1 材料和方法 1.1 研究区域与样地概况 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区实验区内。保护区成立于1986年, 总面积为7 035 hm
2 , 地理坐标22.50°-22.63° N, 101.12°-101.25° E, 地处滇南热带与南亚热带的过渡位置, 保护对象为保存完整的森林生态系统类型。保护区位于无量山末端, 受中山地貌切割, 区内沟谷纵横交错, 形成多种小气候环境, 该区气候受印度洋西南季风暖湿气流影响, 亚热带高原季风气候明显, 干湿季分明, 年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
李帅锋等, 2012 ;
苏建荣等, 2015 )。
季风常绿阔叶林占保护区面积的93.1%, 分布在海拔1 100-1 700 m之间的沟谷与山地, 是云南省南亚热带季风常绿阔叶林分布的代表性地段。由于受热带季风影响, 保护区内还间断分布有沟谷季节雨林, 与季风常绿阔叶林存在重叠分布。保护区内发现维管束植物2 104种, 区系组成属于热带亚洲与东亚植物区系的过渡带上(
朱华等, 2006 ), 植物多样性非常丰富, 蕴藏众多珍稀濒危保护植物, 如国家Ⅰ级保护植物藤枣(
Eleutharrhena macrocarpa )。森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤, 分别对应的植被类型为季节雨林、山地雨林和季风常绿阔叶林, 其中季风常绿阔叶林土壤通透性好于山地雨林(
何蓉, 2003 ), 土壤pH为4.65, 全氮含量为2.94 g·kg
-1 , 全磷含量为0.43 g·kg
-1 , 全钾含量为5.84 g·kg
-1 , 有机质含量为91 g·kg
-1 , 水解性氮含量为313.08 mg·kg
-1 , 有效磷含量为15.71 mg·kg
-1 , 速效钾含量153.54 mg·kg
-1 。
2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 )。样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m。样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1普洱30 hm2 森林动态监测样地地形图。A, 等高线图。B, 三维地形图。 Fig. 1Topography of the 30 hm2 forest dynamics plot of Pu’er. A, Contour map. B, Three dimensional topographic map. 1.2 样地设置及调查方法 普洱30 hm
2 森林动态监测样地从2016年10月开始选点, 并完成地形测量, 2018年10月至2019年3月完成野外植被群落调查。用全站仪将30 hm
2 森林动态监测样地划分为750个20 m × 20 m的样方, 每个样方四角均用水泥桩固定, 并标记样地编号。样方按1-750编号, 起点位于西南角, 将其定义为第一个样方, 编号为001号, 东北角最后一个样方编号为750号, 东西一行共计30个样方, 南北1列共计25个样方。用全站仪测定样地内806个位置点, 基于相对高程结合全球定位系统(GPS)测定动态监测样地四个边角及中心点海拔, 采用Kriging差值法绘制出整个样地地形图(
图1A )。样地植被调查以每个20 m × 20 m样方为单位, 每隔5年复查一次。群落调查时, 将每个样方划分为16个5 m × 5 m的小样方, 对胸径≥1 cm的所有木本植物(包括乔木、藤本、灌木及枯立木)鉴定到种, 每木检尺, 并挂牌标记以备长期监测, 记录其标号、种名、胸径(
DBH )、树高、坐标、分支或萌条状况、生长状况及生境特征。
1.3 数据分析 1.3.1 重要值
以每块20 m × 20 m的样方为单位, 物种在每个样方重要值(
IV )的计算公式(
赖江山等, 2010 ;
孙小伟等, 2018 )为:
(1) IV = (RD + RP )/2 式中,
RD 为相对多度,
RP 为相对显著度。
RD =某种植物在样方中的个体数/样方中所有物种的个体数× 100,
RP =某种植物在样方中所有个体胸高断面积和/样方中所有物种个体的胸高断面积和× 100。
1.3.2 地形因子
包括3个地形因子: 海拔、坡度和坡向。每个样方海拔为4个样方顶点海拔平均值; 坡度为每个样方4个顶点任取3个顶点可以组成一个平面, 可以获得4个不同平面, 利用这4个平面与样方投影面夹角平均值可计算样方坡度(
赖江山等, 2010 ;
王家鸣等, 2018 ); 坡向则结合地理信息系统与野外实测对照获取。
1.3.3 群丛数量分类
鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类。因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 )。MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析。
1.3.4 指示值计算
常绿阔叶林的群丛命名原则采用乔木层优势种为主, 以乔木层下层标志种作为辅助(
宋永昌, 2004 ), 即群丛乔木层优势种通过重要值确定, 将重要值最高的两个乔木种作为群丛名称的主体, 放在最后, 乔木下层则依据指示值确定区别种或诊断种, 放在优势种前面, 用“-”相连, 对群丛名称进行限定区分。物种指示值是一个物种在某个群丛内平均多度和出现频度的组合, 指示值高表示该物种在某个群丛的绝大多数样方都存在, 通过R 3.3.2语言labdsv程序包中的indval函数进行计算(
赖江山等, 2010 ), 物种分布韦恩图通过VennDiagram软件包进行运算。物种中文名与拉丁名参照《中国植物志》电子版(http://www.iplant.cn/frps)。
2 结果 2.1 物种组成 样地内
DBH ≥ 1 cm的木本植物共计271种154 372株, 其中乔木140 702株, 灌木5 271株, 藤本8 399株(
表1 ), 隶属于78科178属, 物种数量随着样方数量累积呈渐近增加趋势(
图2 )。从科的数量统计来看, 大戟科植物最为丰富, 共有10属18种; 其次是樟科, 共有9属18种; 豆科有13属17种, 茜草科有13属17种, 桑科有3属15种, 壳斗科有3属15种。从生活型数量统计来看, 乔木共计181种, 灌木39种, 藤本51种(
表1 )。
Table 1 表1 表1 普洱30 hm
2 森林动态监测样地不同生活型林分特征
Table 1
Stand characteristics of different life form in the 30 hm2 forest dynamics plot of Pu’er 类型 物种丰富度 株数 胸径 树高 乔木 Tree 181 140 702 6.49 ± 7.25 6.98 ± 5.10 灌木 Shrub 39 5 271 2.85 ± 1.90 3.82 ± 1.77 藤本 Liana 51 8 399 3.84 ± 2.97 15.17 ± 6.17 合计 Total 271 154 372 6.22 ± 7.02 7.32 ± 5.46
新窗口打开 |
下载CSV 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2普洱30 hm2 森林动态监测样地物种-样方数量累计 曲线。 Fig. 2Species accumulation curves based on the cumulative subplot number in the 30 hm2 forest dynamics plot of Pu’er. 2.2 群落数量分类 采用MRT将750个样方进行分类, 交叉验证结果显示分为8组具有最小的预测误差, 根据(1 -
SE )规则, 也可以选择分为7组, 海拔、坡向及坡位都参与构建回归树(
图3 ), 而在本研究中发现, 普洱30 hm
2 森林动态监测样地为短刺锥(
Castanopsis echidnocarpa )群系, 群落建群种与优势种明显, 各样方之间存在较多过渡重叠部分, 群丛划分应考虑这一因素, 本文结合地形因子, 通过人机交互的方式选择群丛的分类组数, 最终确定为4个群丛类型(
图4A ), 并将群落类型在样地中显示出来(
图5A )。通过MRT分析发现, 第一次分割以海拔1 522 m为节点, 将750个样方分为两组, 第一组405个样方, 第二组345个样方; 第二次分割以海拔1 486 m为节点, 将345个样方分为两组, 样方数分别为100和245个; 第三次分割以坡向200.8°为节点, 将245个样方分为2组, 样方数分别为186和59个。由
图4A 可以看出, 地形因子中的坡度不参与构建回归树, 这是由于不同坡度过于集中导致群落不宜区分; 物种分布韦恩图(
图5B )显示, 4类群丛物种存在较多的重叠。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3普洱30 hm2 森林动态监测样地群落分类相对误差及交叉验证相对误差变化图。Min + 1SE, 交叉验证相对误差最小值加上一个标准差误差线; cp, 复杂度参数; Inf, 无穷大。 Fig. 3Resubstitution and cross-validation relative error for the multivariate regression tree in the 30 hm2 forest dynamics plot of Pu’er. Min + 1SE, one standard error above the minimum cross-validated relative error; cp, complexity parameter; Inf, infinite. 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4普洱30 hm2 森林动态监测样地群丛分类多元回归树与主成分分析。红点表示群丛1 (珍珠花+毛银柴-短刺锥+泥柯群丛), 绿点表示群丛2 (云南瘿椒树+耳叶柯-短刺锥+西南木荷群丛), 深蓝点表示群丛3 (大果杜英+黄药大头茶-短刺锥+西南木荷群丛), 浅蓝点表示群丛4 (西桦+尼泊尔桤木-短刺锥+枹丝锥群丛)。 Fig. 4Multivariate regression tree and principal component analysis for association classification in the 30 hm2 forest dynamics plot of Pu’er. Red dots indicate Association 1 (Lyonia ovalifolia + Aporosa villosa - Castanopsis echidnocarpa + Lithocarpus fenestratus Association), green dots indicate Association 2 (Tapiscia yunnanensis + Lithocarpus grandifolius - Castanopsis echidnocarpa + Schima wallichii Association), dark blue dots indicate Association 3 (Elaeocarpus sikkimensis + Polyspora chrysandra - Castanopsis echidnocarpa + Schima wallichii Association), and light blue dots indicate Association 4 (Betula alnoides + Alnus nepalensis - Castanopsis echidnocarpa + Castanopsis calathiformis Association). 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5普洱30 hm2 森林动态监测样地4类群丛20 m × 20 m的样方分布及物种分布韦恩图。群丛1, 珍珠花+毛银柴-短刺锥+泥柯群丛; 群丛2, 云南瘿椒树+耳叶柯-短刺锥+西南木荷群丛; 群丛3, 大果杜英+黄药大头茶-短刺锥+西南木荷群丛; 群丛4, 西桦+尼泊尔桤木-短刺锥+枹丝锥群丛。B中数字为物种数量。 Fig. 5Four associations and venn diagram of species distribution at the scale of 20 m × 20 m in the 30 hm2 forest dynamics plot of Pu’er. Association 1, Lyonia ovalifolia + Aporosa villosa - Castanopsis echidnocarpa + Lithocarpus fenestratus Association; Association 2, Tapiscia yunnanensis + Lithocarpus grandifolius - Castanopsis echidnocarpa + Schima wallichii Association; Association 3, Elaeocarpus sikkimensis + Polyspora chrysandra - Castanopsis echidnocarpa + Schima wallichii Association; Association 4, Betula alnoides + Alnus nepalensis - Castanopsis echidnocarpa + Castanopsis calathiformis Association. Number in B represent species mumber. 基于各群丛内物种的重要值(
表2 )和指示值(
表3 )排序对4类群丛进行命名。群丛1中短刺锥和泥柯(
Lithocarpus fenestratus )的重要值要高于其他物种, 为群落中乔木层上层物种, 因此选短刺锥和泥柯为该群丛的优势物种, 该群丛是普洱森林动态监测样地群落类型的主要组成部分; 根据指示值排序, 珍珠花(
Lyonia ovalifolia )和毛银柴(
Aporosa villosa )是群丛1中易与其他群丛类型区别的指示种, 因而将群丛1命名为珍珠花+毛银柴-短刺锥+泥柯群丛(
Lyonia ovalifolia +
Aporosa villosa -
Castanopsis echidnocarpa +
Lithocarpus fenestratus Association)。按照相同的群丛命名原则, 群丛2、3和4根据
表2 重要值和
表3 指示值排序, 分别命名为云南瘿椒树+耳叶柯-短刺锥+西南木荷群丛(
Tapiscia yunnanensis +
Lithocarpus grandifolius -
Castanopsis echidnocarpa +
Schima wallichii Association)、大果杜英+黄药大头茶-短刺锥+西南木荷群丛(
Elaeocarpus sikkimensis +
Polyspora chrysandra -
Castanopsis echidnocarpa +
Schima wallichii Association)、西桦+尼泊尔桤木-短刺锥+枹丝锥群丛(
Betula alnoides +
Alnus nepalensis -
Castanopsis echidnocarpa +
Castanopsis calathiformis Association)。
Table 2 表2 表2 普洱30 hm
2 森林动态监测样地4类群丛内重要值排序
Table 2
Order of four associations by importance value in the 30 hm2 forest dynamics plot of Pu’er 排序 群丛1 Association 1 群丛2 Association 2 群丛3 Association 3 群丛4 Association 4 物种 Species 重要值IV 物种 Species 重要值IV 物种 Species 重要值IV 物种 Species 重要值IV 1 短刺锥Castanopsis echidnocarpa 27.80 短刺锥Castanopsis echidnocarpa 10.28 短刺锥Castanopsis echidnocarpa 25.75 短刺锥Castanopsis echidnocarpa 12.02 2 泥柯Lithocarpus fenestratus 9.99 西南木荷Schima wallichii 6.96 西南木荷Schima wallichii 9.55 枹丝锥Castanopsis calathiformis 7.43 3 西南木荷Schima wallichii 6.67 红梗润楠Machilus rufipes 5.73 泥柯Lithocarpus fenestratus 8.29 西南木荷Schima wallichii 7.30 4 截果柯Lithocarpus truncatus 5.88 云南瘿椒树Tapiscia yunnanensis 5.38 枹丝锥Castanopsis calathiformis 6.15 泥柯Lithocarpus fenestratus 7.26 5 茶梨 Anneslea fragrans 4.23 泥柯Lithocarpus fenestratus 4.73 红梗润楠Machilus rufipes 5.87 红梗润楠Machilus rufipes 5.95 6 西桦 Betula alnoides 4.06 齿叶黄杞Engelhardtia serrata 4.01 截果柯Lithocarpus truncatus 5.24 截果柯Lithocarpus truncatus 5.38 7 红梗润楠Machilus rufipes 4.03 耳叶柯Lithocarpus grandifolius 3.56 红叶木姜子Litsea rubescens 3.90 西桦 Betula alnoides 5.14 8 隐距越桔Vaccinium exaristatum 3.95 厚皮香Ternstroemia gymnanthera 2.97 茶梨Anneslea fragrans 3.83 红叶木姜子Litsea rubescens 3.35 9 红叶木姜子Litsea rubescens 3.70 北酸脚杆Medinilla septentrionalis 2.89 隐距越桔Vaccinium exaristatum 1.98 云南瘿椒树Tapiscia yunnanensis 2.74 10 枹丝锥Castanopsis calathiformis 3.06 枹丝锥Castanopsis calathiformis 2.89 深绿山龙眼Helicia nilagirica 1.79 茶梨 Anneslea fragrans 2.56
IV , importance value.新窗口打开 |
下载CSV Table 3 表3 表3 普洱30 hm
2 森林动态监测样地4类群丛内物种指示值排序
Table 3
Order for four associations by species indicator value in the 30 hm2 forest dynamics plot of Pu’er 物种 指示值 生活型 物种 指示值 生活型 群丛1 Association 1 毛杜茎山 Maesa permollis 0.208 T 红皮水锦树 Wendlandia tinctoria subsp. intermedia 0.529 T 构棘 Maclura cochinchinensis 0.206 L 隐距越桔 Vaccinium exaristatum 0.492 T 野柿 Diospyros kaki var. silvestris 0.195 T 深绿山龙眼 Helicia nilagirica 0.418 T 云南狗牙花 Ervatamia yunnanensis 0.172 T 巴豆藤 Craspedolobium unijugum 0.400 L 假山龙眼 Heliciopsis henryi 0.171 T 短刺锥 Castanopsis echidnocarpa 0.376 T 齿叶枇杷 Eriobotrya serrata 0.160 T 泥柯 Lithocarpus fenestratus 0.334 T 坚核桂樱 Laurocerasus jenkinsii 0.153 T 茶梨 Anneslea fragrans 0.320 T 粗梗胡椒 Piper macropodum 0.151 L 截果柯 Lithocarpus truncatus 0.308 T 苹果榕 Ficus oligodon 0.142 T 毛叶黄杞 Engelhardia spicata var. colebrookeana 0.264 T 珍珠伞 Ardisia maculosa 0.139 T 毛杨梅 Myrica esculenta 0.264 T 岭罗麦 Tarennoidea wallichii 0.138 T 珍珠花 Lyonia ovalifolia 0.243 T 刺篱木 Flacourtia indica 0.136 T 毛银柴 Aporosa villosa 0.194 T 台湾粗叶木 Lasianthus formosensis 0.135 T 猴耳环 Archidendron clypearia 0.188 T 假柿木姜子 Litsea monopetala 0.116 T 密花豆 Spatholobus suberectus 0.188 L 思茅锥 Castanopsis ferox 0.113 T 小果锥 Castanopsis fleuryi 0.168 T 分叉露兜 Pandanus urophyllus 0.112 T 密花树 Myrsine seguinii 0.147 T 焰序山龙眼 Helicia pyrrhobotrya 0.107 T 金叶子 Craibiodendron stellatum 0.113 T 棒果榕 Ficus subincisa 0.106 T 合果木 Michelia baillonii 0.107 T 越南安息香 Styrax tonkinensis 0.104 T 称杆树 Maesa ramentacea 0.077 T 四川冬青 Ilex szechwanensis 0.095 T 群丛2 Association 2 桫椤 Alsophila spinulosa 0.090 T 普文楠 Phoebe puwenensis 0.566 T 粗叶榕 Ficus hirta 0.088 S 阔叶蒲桃 Syzygium megacarpum 0.559 T 飞龙掌血 Toddalia asiatica 0.078 L 齿叶黄杞 Engelhardia serrata 0.514 T 望谟崖摩 Aglaia lawii 0.075 T 耳叶柯 Lithocarpus grandifolius 0.493 T 长叶柞木 Xylosma longifolia 0.074 T 北酸脚杆 Medinilla septentrionalis 0.466 T 砚壳花椒 Zanthoxylum dissitum 0.073 L 山香圆 Turpinia montana 0.462 T 鹧鸪花 Heynea trijuga 0.069 T 云南瘿椒树 Tapiscia yunnanensis 0.435 T 劲直刺桐 Erythrina stricta 0.065 T 艾胶树 Glochidion lanceolarium 0.423 T 鸡爪簕 Benkara sinensis 0.063 T 滇南木姜子 Litsea martabanica 0.393 T 剑叶木姜子 Litsea lancifolia 0.061 T 买麻藤 Gnetum montanum 0.358 L 尼泊尔水东哥 Saurauia napaulensis 0.060 T 钝叶桂 Cinnamomum bejolghota 0.356 T 腺萼木 Mycetia glandulosa 0.060 S 胭脂 Artocarpus tonkinensis 0.343 T 多蕊木 Tupidanthus calyptratus 0.059 T 独子藤 Celastrus monospermus 0.321 L 当归藤 Embelia parviflora 0.059 L 云南狗骨柴 Diplospora mollissima 0.320 T 毛叶合欢 Albizia mollis 0.052 T 纽子果 Ardisia virens 0.314 T 藤金合欢 Acacia concinna 0.051 L 李榄琼楠 Beilschmiedia linocieroides 0.302 T 红果樫木 Dysoxylum gotadhora 0.051 T 山杜英 Elaeocarpus sylvestris 0.295 T 玉兰叶木姜子 Litsea semecarpifolia 0.048 T 大叶斑鸠菊 Vernonia volkameriifolia 0.281 S 钩藤 Uncaria rhynchophylla 0.047 L 黑老虎 Kadsura coccinea 0.275 L 羊脆木 Pittosporum kerrii 0.047 T 山油柑 Acronychia pedunculata 0.242 T 榕叶掌叶树 Euaraliopsis ficifolia 0.042 S 异形南五味子 Kadsura heteroclita 0.240 L 小叶女贞 Ligustrum quihoui 0.042 T 瓦山锥 Castanopsis ceratacantha 0.222 T 中平树 Macaranga denticulata 0.041 T 山蕉 Mitrephora macclurei 0.216 T 微毛布惊 Vitex quinata var. puberula 0.040 T 越南石栎 Lithocarpus annamensis 0.214 T 歪叶榕 Ficus cyrtophylla 0.037 T 泰梭罗 Reevesia pubescens var. siamensis 0.032 T 群丛4 Association 4 森林榕 Ficus neriifolia 0.030 T 五瓣子楝树 Decaspermum parviflorum 0.408 T 矮小天仙果 Ficus erecta var. beecheyana 0.030 T 枹丝锥 Castanopsis calathiformis 0.320 T 南酸枣 Choerospondias axillaris 0.029 T 岗柃 Eurya groffii 0.280 T 爬树龙 Rhaphidophora decursiva 0.026 L 穗序鹅掌柴 Schefflera delavayi 0.260 T 鱼尾葵 Caryota maxima 0.024 T 鲫鱼胆 Maesa perlarius 0.253 T 扁担藤 Tetrastigma planicaule 0.024 L 三桠苦 Melicope pteleifolia 0.193 S 假苹婆 Sterculia lanceolata 0.023 T 圆锥悬钩子 Rubus paniculatus 0.192 L 浆果楝 Cipadessa baccifera 0.022 T 云南臀果木 Pygeum henryi 0.185 T 红枝崖爬藤 Tetrastigma erubescens 0.021 L 西桦 Betula alnoides 0.177 T 白花龙船花 Ixora henryi 0.020 T 黄心树 Machilus gamblei 0.170 T 多体蕊黄檀 Dalbergia polyadelpha 0.020 T 奶桑 Morus macroura 0.148 T 割舌树 Walsura robusta 0.020 T 喜马拉雅崖爬藤 Tetrastigma rumicispermum 0.146 L 水麻 Debregeasia orientalis 0.020 S 檀梨 Pyrularia edulis 0.115 T 泰国黄叶树 Xanthophyllum flavescens 0.020 T 红叶藤 Rourea minor 0.089 L 瓦理棕 Wallichia gracilis 0.020 T 云南黄杞 Engelhardia spicata 0.086 T 牛枓吴萸 Tetradium trichotomum 0.018 T 玉叶金花 Mussaenda pubescens 0.083 L 樱叶杜英 Elaeocarpus prunifolioides 0.018 T 托叶黄檀 Dalbergia stipulacea 0.078 L 细花秀丽火把花 0.017 L 大血藤 Sargentodoxa cuneata 0.077 L Colquhounia elegans var. tenuiflora 无腺吴萸 Tetradium fraxinifolium 0.074 T 群丛3 Association 3 尼泊尔桤木 Alnus nepalensis 0.070 T 红叶木姜子 Litsea rubescens 0.310 T 高盆樱桃 Cerasus cerasoides 0.065 T 西南木荷 Schima wallichii 0.301 T 小萼瓜馥木 Fissistigma polyanthoides 0.057 L 红花木犀榄 Olea rosea 0.218 T 七小叶崖爬藤 Tetrastigma delavayi 0.038 L 腺叶木犀榄 Olea paniculata 0.198 T 一担柴 Colona floribunda 0.025 T 猪肚木 Canthium horridum 0.187 T 楹树 Albizia chinensis 0.022 T 大果杜英 Elaeocarpus sikkimensis 0.165 T 白瑞香 Daphne papyracea 0.020 S 黄药大头茶 Polyspora chrysandra 0.104 T 爪哇黄杞 Engelhardia spicata var. aceriflora 0.017 T 青藤仔 Jasminum nervosum 0.017 L
T, tree; S, shrub; L, liana. T, 乔木; S, 灌木; L, 藤本。 新窗口打开 |
下载CSV 主成分分析(
图4B )进一步显示, 前两轴特征值分别为0.599和0.336, 前两个排序轴的贡献率分别为88.34%和6.35%, 累计贡献率为94.69%, 可以反映群落的基本信息。从
图4B 可知, 普洱森林动态监测样地4类群丛的样方分布有较多重叠, 这与主要优势物种在多数样方均有出现相关。同时, 群丛1中短刺锥既是指示物种, 也是优势物种, 此外珍珠花、毛银柴、金叶子(
Craibiodendron stellatum )等物种也集中出现群丛1中; 群丛2指示物种最多, 达45种, 其中, 指示物种有5种藤本植物, 1种附生乔木; 群丛3中出现的西南木荷、大果杜英、猪肚木(
Canthium horridum )及黄药大头茶等物种指示作用较强; 群丛4中出现的西桦及尼泊尔桤木次生性较强, 可以与其他群丛区分。
2.3 群丛间差异分析 普洱森林动态监测样地4类群丛群落特征差异分析见
表4 , 群丛1面积最大, 而其他3个群丛面积接近; 群丛2的物种丰富度最高, 其次是群丛1、3和4; 群丛2的胸径要显著高于其他3个群丛类型。群丛1的海拔分布范围要显著高于其他类型, 群丛3的林分密度最大, 与群丛4的海拔之间无显著差异, 群丛2的分布海拔最低; 群丛2和3的坡度要显著高于群丛1和4; 群丛4的坡向位于半阴坡, 群丛2位于半阳坡, 群丛1和3则多位于阳坡。
Table 4 表4 表4 普洱30 hm
2 森林动态监测样地4类群丛差异(平均值±标准误差)
Table 4
Variance among four associations in the 30 hm2 forest dynamics plot of Pu’er (mean ± SE ) 类型 Type 面积2 ) 物种丰富度 林分密度-2 ) 胸径 海拔 坡度 坡向 群丛1 Association 1 16.20 195 5 381.79 6.29 ± 0.71b 1 544.24 ± 14.92a 18.7 ± 5.07b 196.01 ± 88.33c 群丛2 Association 2 4.00 214 3 851.50 6.91 ± 1.62a 1 477.81 ± 5.32c 20.22 ± 6.84a 268.73 ± 70.58b 群丛3 Association 3 5.04 186 7 847.00 6.22 ± 0.79b 1 506.53 ± 9.64b 20.17 ± 3.93a 163.88 ± 46.86d 群丛4 Association 4 4.76 172 2 370.00 6.2 ± 0.89b 1 506.22 ± 10.39b 18.42 ± 5.58b 293.76 ± 43.71a
Different lowercase letters showed significant differences among four associations (p < 0.05). 同行不同小写字母表示差异显著(p < 0.05)。 新窗口打开 |
下载CSV 3 讨论 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 )。鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 )。在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 )。本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致。短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类。普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林。西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 )。这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 )。
Table 5 表5 表5 普洱与毗邻区域动态监测样地比较
Table 5
Comparison of forest dynamic plots between Pu’er and adjacent districts 类型 Type 森林动态监测样地 Forest dynamic plot 普洱 Pu’er 西双版纳 Xishuangbanna 哀牢山 Ailaoshan 建成时间 Establishment time (year) 2019 2007 2014 植被类型 Vegetation types 季风常绿阔叶林 热带雨林 中山湿性常绿阔叶林 面积 Area (hm2 ) 30 20 20 物种丰富度 Species richness 271 468 104 海拔范围 Altitude range (m) 1 467-1 587 709-869 2 472-2 628 优势物种 Dominant species 短刺锥、西南木荷、枹丝锥、泥柯、截果柯、红梗润楠、茶梨、西桦、耳叶柯Castanopsis echidnocarpa, Schima wallichii, Castanopsis calathiformis, Lithocarpus fenestratus, Lithocarpus truncates, Machilus rufipes, Anneslea fragrans, Betula alnoides, Lithocarpus grandifolius 假海桐、望天树、云树、短刺锥、蚁花、毛猴欢喜、木奶果、红光树、绒毛番龙眼Pittosporopsis kerrii, Parashorea chinensis, Garcinia cowa, Castanopsis echidnocarpa, Orophea laui, Sloanea tomentosa, Baccaurea ramiflora, Knema tenuinervia, Pometia pinnata 蒙自连蕊茶、云南越桔、多花山矾、硬壳柯、变色锥、南亚枇杷、木果柯、山矾、滇润楠Camellia forrestii, Vaccinium duclouxii, Symplocos ramosissima, Lithocarpus hancei, Castanopsis wattii, Eriobotrya bengalensis, Lithocarpus xylocarpus, Symplocos sumuntia, Machilus yunnanensis 参考文献 Reference - Lan et al ., 2008 Wen et al ., 2018
新窗口打开 |
下载CSV 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 )。MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 )。森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 )。本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局。这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 )。
群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 )。群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显。群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段。群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 )。群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种。
普洱30 hm
2 森林动态监测样地群丛划分主要依据MRT和指示物种, 同时结合样地实际情况, 获得较好的群丛聚类结果, 尽管MRT在划分大样地连续样方或具有过渡样方时更具优势(
张文静等, 2015 ), 但是本文研究表明, 由于群落数量分类侧重于揭示群落的间断性, 从而增加排序的有效性, 因此严格按照MRT相对误差和交叉验证相对误差的结论划分最佳的分类方案(即1 -
SE 规则), 在包含多个优势物种的森林动态监测样地中并不能真实反映群丛分布特征, 会导致群丛分类分组过多与过渡样方自成一体的现状, 其原因需要在今后研究中对群丛分类进行深入探索。本文可以对季风常绿阔叶林生物多样性维持与群落构建机制的深入研究提供重要依据, 但是植物与环境、植物与植物的关系在自然环境中极为复杂, 草本层植物在群丛分类与名称中也十分重要, 本研究所用样方数据仅限于乔木层数据, 同时还有附生植物也是季风常绿阔叶林的重要指示物种, 在野外调查中发现, 群丛2分布的箐沟水分与热量充沛, 分布有掌叶秋海棠(
Begonia hemsleyana )、尖苞柊叶(
Phrynium placentarium )等适应湿热环境的物种, 在后续研究中, 应进一步探究植物群落结构和空间分布与环境因子之间的关系。
[1] Burke A 2001 ). Classification and ordination of plant communities of the Naukluft Mountains, NamibiaJournal of Vegetation Science, 53 -60 . DOI:
10.1111/jvs.2001.12.issue-1 URL [本文引用: 1] [2] Condit R 1998 ). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and A Comparison with Other Plots. [本文引用: 1] [3] Dang CL Wu ZL 1992 ). Studies on the biomass for Castanopsis echidnocarpa community of monsoon evergreen broad-leaved forestJournal of Yunnan University (Natural Sciences )95 -107 . [本文引用: 2] [ 党承林 , 吴兆录 (1992 ). 季风常绿阔叶林短刺栲群落的生物量研究云南大学学报(自然科学版) , 14, 95 -107 .] [本文引用: 2] [4] De’Ath G 2002 ). Multivariate regression trees: a new technique for modeling species-environment relationshipsEcology, 1105 -1117 . [本文引用: 1] [5] Fang JY Shen ZH Cui HT 2004 ). Ecological characteristics of mountains and research issues of mountain ecologyBiodiversity Science, 10 -19 . [本文引用: 1] [ 方精云 , 沈泽昊 , 崔海亭 (2004 ). 试论山地的生态特征及山地生态学的研究内容生物多样性 , 12, 10 -19 .] [本文引用: 1] [6] He R 2003 ). Soil characteristics of three forest types of Caiyanghe nature reserveYunnan Forestry Science and Technology, 2 ), 25 -30 . [本文引用: 1] [ 何蓉 (2003 ). 菜阳河自然保护区3种森林类型的土壤特性云南林业科技 , 103(2 ), 25 -30 .] [本文引用: 1] [7] Lai JS Mi XC Ren HB Ma KP 2009 ). Species-habitat associations change in a subtropical forest of ChinaJournal of Vegetation Science, 415 -423 . DOI:
10.1111/jvs.2009.20.issue-3 URL [本文引用: 2] [8] Lai JS Mi XC Ren HB Ma KP 2010 ). Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees—A case study 24 hm 2 Gutianshan forest plot in China Chinese Journal of Plant Ecology, 761 -769 . [本文引用: 7] [ 赖江山 , 米湘成 , 任海保 , 马克平 (2010 ). 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例植物生态学报 , 34, 761 -769 .] [本文引用: 7] [9] Lan GY Hu YH Cao M Zhu H Wang H Zhou SS Deng XB Cui JY Huang JG Liu LY Xu HL Song JP He YC 2008 ). Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patternsJournal of Plant Ecology (Chinese Version )287 -298 . [本文引用: 1] [ 兰国玉 , 胡跃华 , 曹敏 , 朱华 , 王洪 , 周仕顺 , 邓晓保 , 崔景云 , 黄建国 , 刘林云 , 许海龙 , 宋军平 , 何有才 (2008 ). 西双版纳热带森林动态监测样地——树种组成与空间分布格局植物生态学报 , 32, 287 -298 .] [本文引用: 1] [10] Li SF Liu WD Su JR Zhang ZJ Liu QY 2012 ). Dynamics of seed banks in soil at the different restoration stages of a monsoon broad-leaved evergreen forestScientia Silvae Sinicae, 9 ), 56 -61 . [本文引用: 1] [ 李帅锋 , 刘万德 , 苏建荣 , 张志钧 , 刘庆云 (2012 ). 季风常绿阔叶林土壤种子库的恢复动态林业科学 , 48(9 ), 56 -61 .] [本文引用: 1] [11] Liu WD Su JR Li SF Lang XD Zhang ZJ Huang XB 2015 ). Leaf carbon, nitrogen and phosphorus stoichiometry at different growth stages in dominant tree species of a monsoon broad-leaved evergreen forest in Pu’er, Yunnan Province, ChinaChinese Journal of Plant Ecology, 52 -62 . DOI:
10.17521/cjpe.2015.0006 URL [本文引用: 4] [ 刘万德 , 苏建荣 , 李帅锋 , 郎学东 , 张志钧 , 黄小波 (2015 ). 云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征植物生态学报 , 39, 52 -62 .] DOI:
10.17521/cjpe.2015.0006 URL [本文引用: 4] [12] Ma KP 2017 ). Forest dynamics plot is a crosscutting research platform for biodiversity scienceBiodiversity Science, 227 -228 . DOI:
10.17520/biods.2017113 URL [本文引用: 1] [ 马克平 (2017 ). 森林动态大样地是生物多样性科学综合研究平台生物多样性 , 25, 227 -228 .] DOI:
10.17520/biods.2017113 URL [本文引用: 1] [13] Song YC 2004 ). Tentative classification scheme of evergreen broad-leaved forests of ChinaActa Phytoecologica Sinica, 435 -448 . [本文引用: 5] [ 宋永昌 (2004 ). 中国常绿阔叶林分类试行方案植物生态学报 , 28, 435 -448 .] [本文引用: 5] [14] Su JR Liu WD Li SF Lang XD 2015 ). Restoration Ecology of Monsoon Broad-leaved Evergreen Forest in West China [本文引用: 2] [ 苏建荣 , 刘万德 , 李帅锋 , 郎学东 (2015 ). 西部季风常绿阔叶林恢复生态学 . 科学出版社, 北京.] [本文引用: 2] [15] Sun XW Yang QS Liu HM Wang XH 2018 ). Classification of plant associations based on a 20 hm 2 dynamics plot of evergreen broad-leaved forest in Mt. Tiantong, Zhejiang, China Chinese Journal of Plant Ecology, 550 -561 . DOI:
10.17521/cjpe.2017.0263 URL [本文引用: 6] [ 孙小伟 , 杨庆松 , 刘何铭 , 王希华 (2018 ). 基于浙江天童20 hm 2 常绿阔叶林动态监测样地的群丛划分 植物生态学报 , 42, 550 -561 .] DOI:
10.17521/cjpe.2017.0263 URL [本文引用: 6] [16] Wang JM Xu H Li YD Lin MX Zhou Z Luo TS Chen DX 2018 ). Effects of topographic heterogeneity on community structure and diversity of woody plants in Jianfengling tropical montane rainforestScientia Silvae Sinicae, 1 ), 1 -11 . [本文引用: 1] [ 王家鸣 , 许涵 , 李意德 , 林明献 , 周璋 , 骆土寿 , 陈德祥 (2018 ). 地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响林业科学 , 54(1 ), 1 -11 .] [本文引用: 1] [17] Wang XP Jiang GM 2000 ). The study of classification and geographical distribution of evergreen broadleaved forest in GuangxiJournal of Wuhan Botanical Research, 195 -205 . [本文引用: 1] [ 王献溥 , 蒋高明 (2000 ). 广西常绿阔叶林的分类和地理分布研究武汉植物学研究 , 18, 195 -205 .] [本文引用: 1] [18] Wen HD Lin LX Yang J Hu YH Cao M Liu YH Lu ZY Xie YN 2018 ). Species composition and community structure of a 20 hm 2 plot of mid-mountain moist evergreen broad-leaved forest on the Mts. Ailaoshan, Yunnan Province, China Chinese Journal of Plant Ecology, 419 -429 . DOI:
10.17521/cjpe.2017.0272 URL [本文引用: 1] [ 温韩东 , 林露湘 , 杨洁 , 胡跃华 , 曹敏 , 刘玉洪 , 鲁志云 , 谢有能 (2018 ). 云南哀牢山中山湿性常绿阔叶林20 hm 2 动态样地的物种组成与群落结构 植物生态学报 , 42, 419 -429 .] DOI:
10.17521/cjpe.2017.0272 URL [本文引用: 1] [19] Wu ZY 1987 ). Yunnan Flora [本文引用: 4] [ 吴征镒 (1987 ). 云南植被 . 科学出版社, 北京.] [本文引用: 4] [20] Ye WH Cao HL Huang ZL Lian JY Wang ZG Li L Wei SG Wang ZM 2008 ). Community structure of a 20 hm 2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China Journal of Plant Ecology (Chinese Version )274 -286 . [本文引用: 3] [ 叶万辉 , 曹洪麟 , 黄忠良 , 练琚愉 , 王志高 , 李林 , 魏识广 , 王章明 (2008 ). 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究植物生态学报 , 32, 274 -286 .] [本文引用: 3] [21] Zhang R Dong TF Deng XB Liu JY 2018 ). Vegetation numerical classification and ordination of a 20-hectare tropical forest plot in Xishuangbanna, Southwest YunnanChinese Journal of Ecology, 347 -352 . [本文引用: 6] [ 张容 , 董廷发 , 邓晓保 , 刘俊雁 (2018 ). 西双版纳20公顷样地热带森林植被数量分类与排序生态学杂志 , 37, 347 -352 .] [本文引用: 6] [22] Zhang WJ Zhang QD Wang J Feng F Bi RC 2015 ). A comparison of multivariate regression tree and two-way indicator species analysis in plant community classificationChinese Journal of Plant Ecology, 586 -592 . DOI:
10.17521/cjpe.2015.0056 URL [本文引用: 6] [ 张文静 , 张钦弟 , 王晶 , 冯飞 , 毕润成 (2015 ). 多元回归树与双向指示种分析在群落分类中的应用比较植物生态学报 , 39, 586 -592 .] DOI:
10.17521/cjpe.2015.0056 URL [本文引用: 6] [23] Zhu H 2007 ). On the classification of forest vegetation in Xishuangbanna, Southern YunnanActa Botanica Yunnanica, 377 -387 . [本文引用: 1] [ 朱华 (2007 ). 论滇南西双版纳的森林植被分类云南植物研究 , 29, 377 -387 .] [本文引用: 1] [24] Zhu H 2017 ). A biogeographical study on tropical flora of southern ChinaEcology and Evolution, 10398 -10408 . DOI:
10.1002/ece3.3561 URL [本文引用: 1] [25] Zhu H 2019 ). Floristic divergence of the evergreen broad- leaved forests in Yunnan, southwestern ChinaPhytotaxa, 1 -20 . DOI:
10.11646/phytotaxa.393.1 URL [本文引用: 3] [26] Zhu H Zhao CJ Wang H Zhou SS Shi JP Li BG 2006 ). A study on the flora of Caiyanghe Nature Reserve in Simao, Yunnan with references to the transition from tropical Asian flora to Eastern Asian floraBulletin of Botanical Research, 38 -52 . [本文引用: 1] [ 朱华 , 赵崇奖 , 王洪 , 周时顺 , 施济普 , 李保贵 (2006 ). 思茅菜阳河自然保护区植物区系研究——兼论热带亚洲植物区系向东亚植物区系的过渡植物研究 , 26, 38 -52 .] [本文引用: 1] [27] Zhu H Zhou SS Yan LC Shi JP Shen YX 2019 ). Studies on the evergreen broad-leaved forests of Yunnan, southwestern ChinaThe Botanical Review, 131 -148 . DOI:
10.1007/s12229-019-09210-1 URL [本文引用: 3] [28] Zou S Zhou GY Zhang QM Xu S Xiong X Xia YJ Liu SZ Meng Z Chu GW 2018 ). Long-term (1992-2015) dynamics of community composition and structure in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere ReserveChinese Journal of Plant Ecology, 442 -452 . DOI:
10.17521/cjpe.2017.0171 URL [本文引用: 1] [ 邹顺 , 周国逸 , 张倩媚 , 徐珊 , 熊鑫 , 夏艳菊 , 刘世忠 , 孟泽 , 褚国伟 (2018 ). 1992-2015年鼎湖山季风常绿阔叶林群落结构动态植物生态学报 , 42, 442 -452 .] DOI:
10.17521/cjpe.2017.0171 URL [本文引用: 1] Classification and ordination of plant communities of the Naukluft Mountains, Namibia
1
2001
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
1
1998
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
季风常绿阔叶林短刺栲群落的生物量研究
2
1992
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
季风常绿阔叶林短刺栲群落的生物量研究
2
1992
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
Multivariate regression trees: a new technique for modeling species-environment relationships
1
2002
... 鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类.因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
试论山地的生态特征及山地生态学的研究内容
1
2004
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
试论山地的生态特征及山地生态学的研究内容
1
2004
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
菜阳河自然保护区3种森林类型的土壤特性
1
2003
... 季风常绿阔叶林占保护区面积的93.1%, 分布在海拔1 100-1 700 m之间的沟谷与山地, 是云南省南亚热带季风常绿阔叶林分布的代表性地段.由于受热带季风影响, 保护区内还间断分布有沟谷季节雨林, 与季风常绿阔叶林存在重叠分布.保护区内发现维管束植物2 104种, 区系组成属于热带亚洲与东亚植物区系的过渡带上(
朱华等, 2006 ), 植物多样性非常丰富, 蕴藏众多珍稀濒危保护植物, 如国家Ⅰ级保护植物藤枣(
Eleutharrhena macrocarpa ).森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤, 分别对应的植被类型为季节雨林、山地雨林和季风常绿阔叶林, 其中季风常绿阔叶林土壤通透性好于山地雨林(
何蓉, 2003 ), 土壤pH为4.65, 全氮含量为2.94 g·kg
-1 , 全磷含量为0.43 g·kg
-1 , 全钾含量为5.84 g·kg
-1 , 有机质含量为91 g·kg
-1 , 水解性氮含量为313.08 mg·kg
-1 , 有效磷含量为15.71 mg·kg
-1 , 速效钾含量153.54 mg·kg
-1 . ...
菜阳河自然保护区3种森林类型的土壤特性
1
2003
... 季风常绿阔叶林占保护区面积的93.1%, 分布在海拔1 100-1 700 m之间的沟谷与山地, 是云南省南亚热带季风常绿阔叶林分布的代表性地段.由于受热带季风影响, 保护区内还间断分布有沟谷季节雨林, 与季风常绿阔叶林存在重叠分布.保护区内发现维管束植物2 104种, 区系组成属于热带亚洲与东亚植物区系的过渡带上(
朱华等, 2006 ), 植物多样性非常丰富, 蕴藏众多珍稀濒危保护植物, 如国家Ⅰ级保护植物藤枣(
Eleutharrhena macrocarpa ).森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤, 分别对应的植被类型为季节雨林、山地雨林和季风常绿阔叶林, 其中季风常绿阔叶林土壤通透性好于山地雨林(
何蓉, 2003 ), 土壤pH为4.65, 全氮含量为2.94 g·kg
-1 , 全磷含量为0.43 g·kg
-1 , 全钾含量为5.84 g·kg
-1 , 有机质含量为91 g·kg
-1 , 水解性氮含量为313.08 mg·kg
-1 , 有效磷含量为15.71 mg·kg
-1 , 速效钾含量153.54 mg·kg
-1 . ...
Species-habitat associations change in a subtropical forest of China
2
2009
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
7
2010
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
... 以每块20 m × 20 m的样方为单位, 物种在每个样方重要值(
IV )的计算公式(
赖江山等, 2010 ;
孙小伟等, 2018 )为: ...
... 包括3个地形因子: 海拔、坡度和坡向.每个样方海拔为4个样方顶点海拔平均值; 坡度为每个样方4个顶点任取3个顶点可以组成一个平面, 可以获得4个不同平面, 利用这4个平面与样方投影面夹角平均值可计算样方坡度(
赖江山等, 2010 ;
王家鸣等, 2018 ); 坡向则结合地理信息系统与野外实测对照获取. ...
... 鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类.因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... 常绿阔叶林的群丛命名原则采用乔木层优势种为主, 以乔木层下层标志种作为辅助(
宋永昌, 2004 ), 即群丛乔木层优势种通过重要值确定, 将重要值最高的两个乔木种作为群丛名称的主体, 放在最后, 乔木下层则依据指示值确定区别种或诊断种, 放在优势种前面, 用“-”相连, 对群丛名称进行限定区分.物种指示值是一个物种在某个群丛内平均多度和出现频度的组合, 指示值高表示该物种在某个群丛的绝大多数样方都存在, 通过R 3.3.2语言labdsv程序包中的indval函数进行计算(
赖江山等, 2010 ), 物种分布韦恩图通过VennDiagram软件包进行运算.物种中文名与拉丁名参照《中国植物志》电子版(http://www.iplant.cn/frps). ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
7
2010
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
... 以每块20 m × 20 m的样方为单位, 物种在每个样方重要值(
IV )的计算公式(
赖江山等, 2010 ;
孙小伟等, 2018 )为: ...
... 包括3个地形因子: 海拔、坡度和坡向.每个样方海拔为4个样方顶点海拔平均值; 坡度为每个样方4个顶点任取3个顶点可以组成一个平面, 可以获得4个不同平面, 利用这4个平面与样方投影面夹角平均值可计算样方坡度(
赖江山等, 2010 ;
王家鸣等, 2018 ); 坡向则结合地理信息系统与野外实测对照获取. ...
... 鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类.因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... 常绿阔叶林的群丛命名原则采用乔木层优势种为主, 以乔木层下层标志种作为辅助(
宋永昌, 2004 ), 即群丛乔木层优势种通过重要值确定, 将重要值最高的两个乔木种作为群丛名称的主体, 放在最后, 乔木下层则依据指示值确定区别种或诊断种, 放在优势种前面, 用“-”相连, 对群丛名称进行限定区分.物种指示值是一个物种在某个群丛内平均多度和出现频度的组合, 指示值高表示该物种在某个群丛的绝大多数样方都存在, 通过R 3.3.2语言labdsv程序包中的indval函数进行计算(
赖江山等, 2010 ), 物种分布韦恩图通过VennDiagram软件包进行运算.物种中文名与拉丁名参照《中国植物志》电子版(http://www.iplant.cn/frps). ...
西双版纳热带森林动态监测样地——树种组成与空间分布格局
1
2008
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
西双版纳热带森林动态监测样地——树种组成与空间分布格局
1
2008
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
季风常绿阔叶林土壤种子库的恢复动态
1
2012
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区实验区内.保护区成立于1986年, 总面积为7 035 hm
2 , 地理坐标22.50°-22.63° N, 101.12°-101.25° E, 地处滇南热带与南亚热带的过渡位置, 保护对象为保存完整的森林生态系统类型.保护区位于无量山末端, 受中山地貌切割, 区内沟谷纵横交错, 形成多种小气候环境, 该区气候受印度洋西南季风暖湿气流影响, 亚热带高原季风气候明显, 干湿季分明, 年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
李帅锋等, 2012 ;
苏建荣等, 2015 ). ...
季风常绿阔叶林土壤种子库的恢复动态
1
2012
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区实验区内.保护区成立于1986年, 总面积为7 035 hm
2 , 地理坐标22.50°-22.63° N, 101.12°-101.25° E, 地处滇南热带与南亚热带的过渡位置, 保护对象为保存完整的森林生态系统类型.保护区位于无量山末端, 受中山地貌切割, 区内沟谷纵横交错, 形成多种小气候环境, 该区气候受印度洋西南季风暖湿气流影响, 亚热带高原季风气候明显, 干湿季分明, 年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
李帅锋等, 2012 ;
苏建荣等, 2015 ). ...
云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征
4
2015
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区实验区内.保护区成立于1986年, 总面积为7 035 hm
2 , 地理坐标22.50°-22.63° N, 101.12°-101.25° E, 地处滇南热带与南亚热带的过渡位置, 保护对象为保存完整的森林生态系统类型.保护区位于无量山末端, 受中山地貌切割, 区内沟谷纵横交错, 形成多种小气候环境, 该区气候受印度洋西南季风暖湿气流影响, 亚热带高原季风气候明显, 干湿季分明, 年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
李帅锋等, 2012 ;
苏建荣等, 2015 ). ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征
4
2015
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... 研究区域位于云南省普洱市思茅区南部太阳河省级自然保护区实验区内.保护区成立于1986年, 总面积为7 035 hm
2 , 地理坐标22.50°-22.63° N, 101.12°-101.25° E, 地处滇南热带与南亚热带的过渡位置, 保护对象为保存完整的森林生态系统类型.保护区位于无量山末端, 受中山地貌切割, 区内沟谷纵横交错, 形成多种小气候环境, 该区气候受印度洋西南季风暖湿气流影响, 亚热带高原季风气候明显, 干湿季分明, 年平均气温17.7 ℃, 年日照时间2 122.9 h, ≥10 ℃的积温为6 353.5 ℃, 年降水量1 547.6 mm, 主要集中在5-10月的雨季(
李帅锋等, 2012 ;
苏建荣等, 2015 ). ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
森林动态大样地是生物多样性科学综合研究平台
1
2017
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
森林动态大样地是生物多样性科学综合研究平台
1
2017
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
中国常绿阔叶林分类试行方案
5
2004
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 常绿阔叶林的群丛命名原则采用乔木层优势种为主, 以乔木层下层标志种作为辅助(
宋永昌, 2004 ), 即群丛乔木层优势种通过重要值确定, 将重要值最高的两个乔木种作为群丛名称的主体, 放在最后, 乔木下层则依据指示值确定区别种或诊断种, 放在优势种前面, 用“-”相连, 对群丛名称进行限定区分.物种指示值是一个物种在某个群丛内平均多度和出现频度的组合, 指示值高表示该物种在某个群丛的绝大多数样方都存在, 通过R 3.3.2语言labdsv程序包中的indval函数进行计算(
赖江山等, 2010 ), 物种分布韦恩图通过VennDiagram软件包进行运算.物种中文名与拉丁名参照《中国植物志》电子版(http://www.iplant.cn/frps). ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
中国常绿阔叶林分类试行方案
5
2004
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 常绿阔叶林的群丛命名原则采用乔木层优势种为主, 以乔木层下层标志种作为辅助(
宋永昌, 2004 ), 即群丛乔木层优势种通过重要值确定, 将重要值最高的两个乔木种作为群丛名称的主体, 放在最后, 乔木下层则依据指示值确定区别种或诊断种, 放在优势种前面, 用“-”相连, 对群丛名称进行限定区分.物种指示值是一个物种在某个群丛内平均多度和出现频度的组合, 指示值高表示该物种在某个群丛的绝大多数样方都存在, 通过R 3.3.2语言labdsv程序包中的indval函数进行计算(
赖江山等, 2010 ), 物种分布韦恩图通过VennDiagram软件包进行运算.物种中文名与拉丁名参照《中国植物志》电子版(http://www.iplant.cn/frps). ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
2
2015
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
2
2015
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
基于浙江天童20 hm
2 常绿阔叶林动态监测样地的群丛划分
6
2018
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
... 以每块20 m × 20 m的样方为单位, 物种在每个样方重要值(
IV )的计算公式(
赖江山等, 2010 ;
孙小伟等, 2018 )为: ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
基于浙江天童20 hm
2 常绿阔叶林动态监测样地的群丛划分
6
2018
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
... 以每块20 m × 20 m的样方为单位, 物种在每个样方重要值(
IV )的计算公式(
赖江山等, 2010 ;
孙小伟等, 2018 )为: ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响
1
2018
... 包括3个地形因子: 海拔、坡度和坡向.每个样方海拔为4个样方顶点海拔平均值; 坡度为每个样方4个顶点任取3个顶点可以组成一个平面, 可以获得4个不同平面, 利用这4个平面与样方投影面夹角平均值可计算样方坡度(
赖江山等, 2010 ;
王家鸣等, 2018 ); 坡向则结合地理信息系统与野外实测对照获取. ...
地形异质性对尖峰岭热带山地雨林木本植物群落结构及多样性的影响
1
2018
... 包括3个地形因子: 海拔、坡度和坡向.每个样方海拔为4个样方顶点海拔平均值; 坡度为每个样方4个顶点任取3个顶点可以组成一个平面, 可以获得4个不同平面, 利用这4个平面与样方投影面夹角平均值可计算样方坡度(
赖江山等, 2010 ;
王家鸣等, 2018 ); 坡向则结合地理信息系统与野外实测对照获取. ...
广西常绿阔叶林的分类和地理分布研究
1
2000
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
广西常绿阔叶林的分类和地理分布研究
1
2000
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
云南哀牢山中山湿性常绿阔叶林20 hm
2 动态样地的物种组成与群落结构
1
2018
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
云南哀牢山中山湿性常绿阔叶林20 hm
2 动态样地的物种组成与群落结构
1
2018
... 2019年3月, 云南省普洱30 hm
2 季风常绿阔叶林动态监测样地在太阳河省级自然保护区建成(22.58° N, 101.13° E), 技术规范参照了Center for Tropical Forest Science (CTFS)建设标准(
Condit, 1998 ;
赖江山等, 2010 ;
孙小伟等, 2018 ;
温韩东等; 2018 ).样地为长方形, 东西长600 m, 南北宽500 m, 最高海拔1 587 m, 最低海拔1 467 m, 平均海拔为1 523 m.样地总体上东北高、西南低, 样地内包含两条箐沟, 一条山脊西南走向上下分割, 地形较为复杂(
图1B ). ...
4
1987
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
4
1987
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 群落分类的依据即要求物种组成和结构相对一致, 同时对占据的生境也有严格的要求, 这是因为不同植物具有相差较大的生境偏好(
Lai et al ., 2009 ), 地形因子在生境异质性较大的区域是形成生境差异的最根本因素之一(
方精云等, 2004 ).群丛2主要分布在海拔1 467-1 486 m的区域内, 大地形为陡峭箐沟, 为森林动态监测样地海拔分布最低的区域, 该群丛以短刺锥、西南木荷为主要优势种, 同时优势种还有耳叶柯与云南瘿椒树等其他群丛中较少出现的种类, 但是在群丛指示种分析中发现, 该群落类型指示种达45种, 远高于其他群丛, 其中有些物种, 具有较为明显的沟谷热带雨林成分, 如耳叶柯、阔叶蒲桃(
Syzygium megacarpum )、苹果榕(
Ficus oligodon )、分叉露兜(
Pandanus urophyllus )、桫椤(
Alsophila spinulosa )、尼泊尔水东哥(
Saurauia napaulensis )、鱼尾葵(
Caryota maxima )、瓦理棕(
Wallichia gracilis )等物种(
吴征镒, 1987 ), 其热带雨林向季风常绿阔叶林过渡特征明显.群丛3与4则与坡向的分布具有直接的关系, 前者林分密度最大, 集中分布在向阳的区域, 群落中出现的西南木荷与红叶木姜子既是优势物种也是指示物种, 而后者则集中分布在背阴的区域, 群落中出现的指示植物有较多的落叶植物, 如西桦、尼泊尔桤木, 这些物种为高大乔木, 反映出群落在时间上处于演替的中间阶段.群丛4在西北角分布的区域在20世纪80年代进行过小面积毁林开荒(
图5 ), 说明人为干扰是影响植被群落空间分布格局的重要因素(
Burke, 2001 ).群丛1分布面积最大, 主要集中在海拔1 522-1 587 m的山脊两侧, 林分密度较大, 短刺锥的优势地位突出, 群落中出现的珍珠花可以分布在海拔更高的区域, 而海拔较低的区域则较少见到该物种. ...
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
3
2008
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
3
2008
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
西双版纳20公顷样地热带森林植被数量分类与排序
6
2018
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
西双版纳20公顷样地热带森林植被数量分类与排序
6
2018
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 随着物种共存或群落构建机制研究在植被生态研究中取得明显进展, 大型森林动态监测样地成为森林生物多样性研究重要的研究平台(
马克平, 2017 ).2019年3月, 中国林业科学研究院资源昆虫研究所在云南省普洱市太阳河省级自然保护区建成30 hm
2 季风常绿阔叶林森林动态监测样地, 与广东省鼎湖山南亚热带常绿阔叶林20 hm
2 样地(
叶万辉等, 2008 ), 成为研究我国季风常绿阔叶林植被数量分类的天然试验场.目前针对季风常绿阔叶林植被数量分类的研究, 多集中在群系(
宋永昌, 2004 ;
张容等, 2018 ), 而群丛数量分类还较少涉及.本文以普洱30 hm
2 森林动态监测样地第一次调查数据为基础, 在采用多元回归树、主成分分析、重要值及指示值划分群丛类型, 分析了季风常绿阔叶林的物种组成、群落分布规律和影响群落分布的主要地形因子.本研究有助于深入理解南亚热带森林群落类型的空间分布及各优势种的生态适应特性, 为南亚热带季风常绿阔叶林植被生态保护和生态系统功能管理提供理论依据. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
多元回归树与双向指示种分析在群落分类中的应用比较
6
2015
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类.因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... 普洱30 hm
2 森林动态监测样地群丛划分主要依据MRT和指示物种, 同时结合样地实际情况, 获得较好的群丛聚类结果, 尽管MRT在划分大样地连续样方或具有过渡样方时更具优势(
张文静等, 2015 ), 但是本文研究表明, 由于群落数量分类侧重于揭示群落的间断性, 从而增加排序的有效性, 因此严格按照MRT相对误差和交叉验证相对误差的结论划分最佳的分类方案(即1 -
SE 规则), 在包含多个优势物种的森林动态监测样地中并不能真实反映群丛分布特征, 会导致群丛分类分组过多与过渡样方自成一体的现状, 其原因需要在今后研究中对群丛分类进行深入探索.本文可以对季风常绿阔叶林生物多样性维持与群落构建机制的深入研究提供重要依据, 但是植物与环境、植物与植物的关系在自然环境中极为复杂, 草本层植物在群丛分类与名称中也十分重要, 本研究所用样方数据仅限于乔木层数据, 同时还有附生植物也是季风常绿阔叶林的重要指示物种, 在野外调查中发现, 群丛2分布的箐沟水分与热量充沛, 分布有掌叶秋海棠(
Begonia hemsleyana )、尖苞柊叶(
Phrynium placentarium )等适应湿热环境的物种, 在后续研究中, 应进一步探究植物群落结构和空间分布与环境因子之间的关系. ...
多元回归树与双向指示种分析在群落分类中的应用比较
6
2015
... 植被群落分类作为基础学科的重要工具, 能促进研究者对生态格局理解以及收集和解释生态背景信息, 在理论与实践研究中具有重要意义(
孙小伟等, 2018 ), 由于植物群落分布存在连续性和间断性, 使得植被分类成为植被生态学研究中最复杂和充满争议的一个问题(
吴征镒, 1987 ).常绿阔叶林复杂性仅次于热带雨林, 其过渡性群落极多, 因而植被分类难度更大, 至今没有公认的分类标准和原则被大家接受(
宋永昌, 2004 ).植被分类基本单位为植被型、群系与群丛, 其中群丛作为植被分类的基本单位, 是由很多植物个体交织而成的群落联合体, 其群落结构的相似导致群丛分类比植物物种分类更为困难(
吴征镒, 1987 ;
赖江山等, 2010 ).随着植被生态学研究的深入以及计算机技术的高速发展, 植被分类已由早期的人为分类方法向数量分类方法转变, 群落排序、聚类、双向指示种分析(TWINSPAN)及多元回归树(MRT)已成为常用的群落数量分类方法(
赖江山等, 2010 ;
孙小伟等, 2018 ;
张容等, 2018 ).近期研究表明在划分森林动态监测样地连续样方或具有过渡性质样方时, 多元回归树更具优势(
张文静等, 2015 ), 成为研究植被群落物种分布与环境因子之间关系的数量分类方法, 该方法是结合环境因子作为分类节点, 利用递归划分法, 将样方尽可能分为同质的类群, 避免了人为因素的干扰, 分类结果更为客观, 能够揭示植被空间分布格局. ...
... 鉴于MRT在划分森林动态监测样地连续样方或具有过渡性质样方更具优势(
赖江山等, 2010 ;
张文静等, 2015 ), 本文采用以环境梯度为分类依据的MRT对普洱森林动态监测样地进行群落分类.因变量为样地内271种木本植物在750个样方内的重要值, 建立271 × 750的物种-样方矩阵, 自变量选择每个样方的海拔、坡度与坡向作为地形因子, 建立3 × 750的地形因子-样方矩阵, 采用交叉验证对分类结果进行剪枝, 参考剪枝理论(1 -
SE )规则, 获得合适的分类树(
De’Ath, 2002 ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... ;
张文静等, 2015 ).MRT基于R 3.3.2语言mvpart程序包进行运算, 并基于该程序包对群落分类结果进行主成分分析. ...
... 本研究基于MRT和指示物种对普洱30 hm
2 季风常绿阔叶林动态监测样地进行群丛数量分类, 可以清晰看出样方分布与环境变量的关系, 比TWINSPAN更直观(
张文静等, 2015 ).MRT将750个样方分为4类群丛, 群落乔木建群种相同, 存在较多的物种重叠(
图4B ), 同为短刺锥, 在群落的各个层次都有一定数量的个体, 而且有较多的幼树和幼苗, 是该地区季风常绿阔叶林的优势种, 其生物量在中龄林中可达60%以上, 是构建群落生物量的主体(
党承林和吴兆录, 1992 ;
刘万德等, 2015 ).森林动态监测样地由于样方分布具有连续性和过渡性特征, 其界限往往难以区分, 群丛的划分则要求种类组成和结构相对一致, 且占据相同的生境(
孙小伟等, 2018 ), 而在类似生境下, 常绿阔叶林群丛种类组成差异较小, 区别在于小生境和群落发展阶段不同产生的建群成分及其优势程度与伴生种的组合和数量的差异(
王献溥和蒋高明, 2000 ).本文的群丛数量分类主要是基于物种重要值及地形因子基础进行划分, 研究发现群丛分布与海拔和坡向联系较大, 共同影响了植被的分布格局.这也证实了地形因子是影响植被数量分类的重要因素(
Lai et al ., 2009 ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... ;
张文静等, 2015 ;
孙小伟等, 2018 ;
张容等, 2018 ). ...
... 普洱30 hm
2 森林动态监测样地群丛划分主要依据MRT和指示物种, 同时结合样地实际情况, 获得较好的群丛聚类结果, 尽管MRT在划分大样地连续样方或具有过渡样方时更具优势(
张文静等, 2015 ), 但是本文研究表明, 由于群落数量分类侧重于揭示群落的间断性, 从而增加排序的有效性, 因此严格按照MRT相对误差和交叉验证相对误差的结论划分最佳的分类方案(即1 -
SE 规则), 在包含多个优势物种的森林动态监测样地中并不能真实反映群丛分布特征, 会导致群丛分类分组过多与过渡样方自成一体的现状, 其原因需要在今后研究中对群丛分类进行深入探索.本文可以对季风常绿阔叶林生物多样性维持与群落构建机制的深入研究提供重要依据, 但是植物与环境、植物与植物的关系在自然环境中极为复杂, 草本层植物在群丛分类与名称中也十分重要, 本研究所用样方数据仅限于乔木层数据, 同时还有附生植物也是季风常绿阔叶林的重要指示物种, 在野外调查中发现, 群丛2分布的箐沟水分与热量充沛, 分布有掌叶秋海棠(
Begonia hemsleyana )、尖苞柊叶(
Phrynium placentarium )等适应湿热环境的物种, 在后续研究中, 应进一步探究植物群落结构和空间分布与环境因子之间的关系. ...
论滇南西双版纳的森林植被分类
1
2007
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
论滇南西双版纳的森林植被分类
1
2007
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
A biogeographical study on tropical flora of southern China
1
2017
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
Floristic divergence of the evergreen broad- leaved forests in Yunnan, southwestern China
3
2019
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
思茅菜阳河自然保护区植物区系研究——兼论热带亚洲植物区系向东亚植物区系的过渡
1
2006
... 季风常绿阔叶林占保护区面积的93.1%, 分布在海拔1 100-1 700 m之间的沟谷与山地, 是云南省南亚热带季风常绿阔叶林分布的代表性地段.由于受热带季风影响, 保护区内还间断分布有沟谷季节雨林, 与季风常绿阔叶林存在重叠分布.保护区内发现维管束植物2 104种, 区系组成属于热带亚洲与东亚植物区系的过渡带上(
朱华等, 2006 ), 植物多样性非常丰富, 蕴藏众多珍稀濒危保护植物, 如国家Ⅰ级保护植物藤枣(
Eleutharrhena macrocarpa ).森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤, 分别对应的植被类型为季节雨林、山地雨林和季风常绿阔叶林, 其中季风常绿阔叶林土壤通透性好于山地雨林(
何蓉, 2003 ), 土壤pH为4.65, 全氮含量为2.94 g·kg
-1 , 全磷含量为0.43 g·kg
-1 , 全钾含量为5.84 g·kg
-1 , 有机质含量为91 g·kg
-1 , 水解性氮含量为313.08 mg·kg
-1 , 有效磷含量为15.71 mg·kg
-1 , 速效钾含量153.54 mg·kg
-1 . ...
思茅菜阳河自然保护区植物区系研究——兼论热带亚洲植物区系向东亚植物区系的过渡
1
2006
... 季风常绿阔叶林占保护区面积的93.1%, 分布在海拔1 100-1 700 m之间的沟谷与山地, 是云南省南亚热带季风常绿阔叶林分布的代表性地段.由于受热带季风影响, 保护区内还间断分布有沟谷季节雨林, 与季风常绿阔叶林存在重叠分布.保护区内发现维管束植物2 104种, 区系组成属于热带亚洲与东亚植物区系的过渡带上(
朱华等, 2006 ), 植物多样性非常丰富, 蕴藏众多珍稀濒危保护植物, 如国家Ⅰ级保护植物藤枣(
Eleutharrhena macrocarpa ).森林土壤为粗骨性赤红壤、紫色赤红壤和黄色赤红壤, 分别对应的植被类型为季节雨林、山地雨林和季风常绿阔叶林, 其中季风常绿阔叶林土壤通透性好于山地雨林(
何蓉, 2003 ), 土壤pH为4.65, 全氮含量为2.94 g·kg
-1 , 全磷含量为0.43 g·kg
-1 , 全钾含量为5.84 g·kg
-1 , 有机质含量为91 g·kg
-1 , 水解性氮含量为313.08 mg·kg
-1 , 有效磷含量为15.71 mg·kg
-1 , 速效钾含量153.54 mg·kg
-1 . ...
Studies on the evergreen broad-leaved forests of Yunnan, southwestern China
3
2019
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
... 普洱30 hm
2 森林动态监测样地群系类型为短刺锥群系, 与西双版纳20 hm
2 森林动态监测样地中的热带山地常绿阔叶林为同一类型(
张容等, 2018 ), 与鼎湖山森林动态监测样地的厚壳桂(
Cryptocarya chinensis ) -栲类群系区别较大(
叶万辉等, 2008 ).鼎湖山森林动态监测样地属于东部(大陆)亚热带季风常绿阔叶林, 普洱动态监测样地属于西部亚热带季风常绿阔叶林(
宋永昌, 2004 ).在以往研究中, 其群系命名为刺栲(
Castanopsis hystrix )、印栲(
Castanopsis indica )群系(
吴征镒, 1987 ;
宋永昌, 2004 ).本研究发现普洱森林动态监测样地群落类型中优势物种主要以短刺锥、西南木荷、枹丝锥与泥柯为主, 这与对西双版纳季风常绿阔叶林的研究结论(
朱华, 2007 ;
兰国玉等, 2008 ;
张容等, 2018 ;
Zhu, 2019 )一致.短刺锥群系应该为滇南地区主要的群系类型, 尤其在普洱地区分布较广(
党承林和吴兆录, 1992 ), 本研究丰富了云南省植被分类.普洱太阳河、西双版纳与哀牢山等3个森林动态监测样地概况及优势物种比较见
表5 , 季风常绿阔叶林的物种丰富度要低于热带雨林, 而高于中山湿性常绿阔叶林.西双版纳热带雨林动态监测样地中分布有9.76 hm
2 的季风常绿阔叶林, 接近样地面积一半, 主要分布在海拔770 m以上的山脊地区, 与热带雨林存在交错分布现象(
张容等, 2018 ); 同时, 哀牢山中山湿性常绿阔叶林的优势物种与季风常绿阔叶林相比, 发生极大变化(表5), 一方面出现了物种替代, 如云南越桔(
Vaccinium duclouxii )替代了隐距越桔, 硬壳柯(
Lithocarpus hancei )和木果柯(
Lithocarpus xylocarpus )替代了以泥柯与截果柯(
Lithocarpus truncatus )为代表的壳斗科优势物种, 另一方面季风常绿阔叶林出现数量较少的变色锥(
Castanopsis wattii ), 成为哀牢山中山湿性常绿阔叶林的优势物种, 这是因为季风常绿阔叶林具有热带森林生态外貌和显著的热带亚洲植物区系组成, 而与哀牢山的植被类型相区分(
Zhu, 2019 ;
Zhu et al ., 2019 ).这也证实了季风常绿阔叶林具有热带向亚热带过渡的性质(
叶万辉等, 2008 ;
苏建荣等, 2015 ;
Zhu, 2017 ;
Zhu et al ., 2019 ). ...
... ;
Zhu et al ., 2019 ). ...
1992-2015年鼎湖山季风常绿阔叶林群落结构动态
1
2018
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
1992-2015年鼎湖山季风常绿阔叶林群落结构动态
1
2018
... 季风常绿阔叶林是我国最复杂、生产力最高、生物多样性最丰富的一种地带性植被, 对保护环境和维持全球碳平衡具有极重要的作用, 在我国南亚热带的生态环境建设中占据举足轻重的地位(
刘万德等, 2015 ;
苏建荣等, 2015 ;
邹顺等, 2018 ).云南分布的季风常绿阔叶林物种组成极其丰富, 具有热带森林的生态外貌和显著的热带亚洲区系亲缘, 与东南亚的热带低山常绿阔叶林为同一类型(
Zhu, 2019 ;
Zhu et al ., 2019 ).由于受茶叶、咖啡等种植活动的影响, 目前季风常绿阔叶林分布面积逐年下降, 多呈片段孤岛分布, 严重影响到森林生态系统功能与服务的发挥, 导致森林生产力等诸多功能出现衰退现象(
苏建荣等, 2015 ), 因而季风常绿阔叶林群落生态过程及其生物多样性维持机制研究对天然林保护具有重要意义. ...
 ,
, ,1,*
,1,*
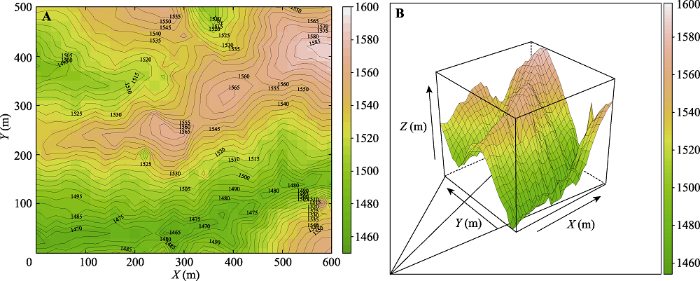 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT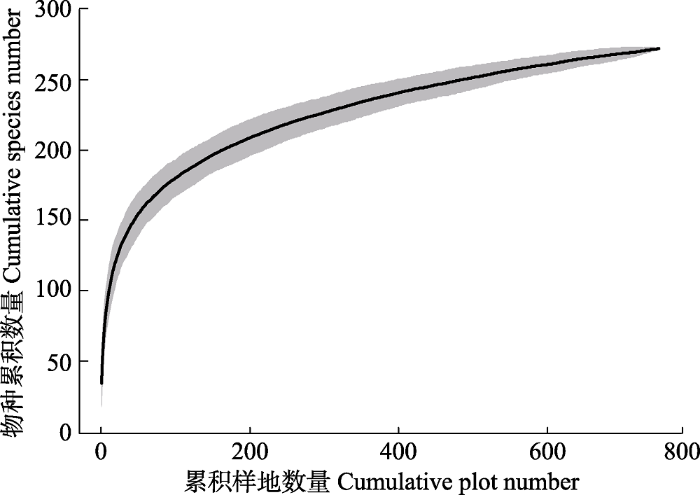 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT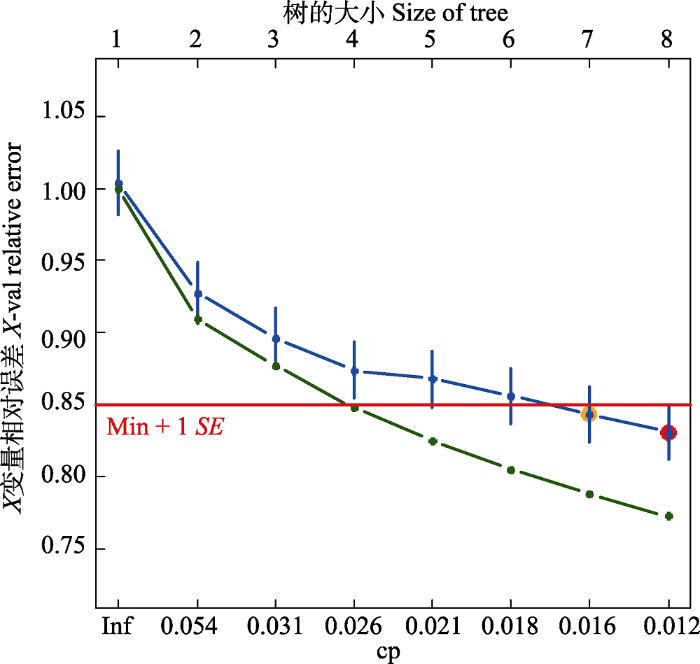 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT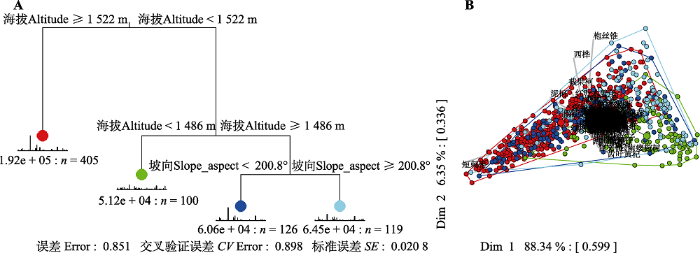 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT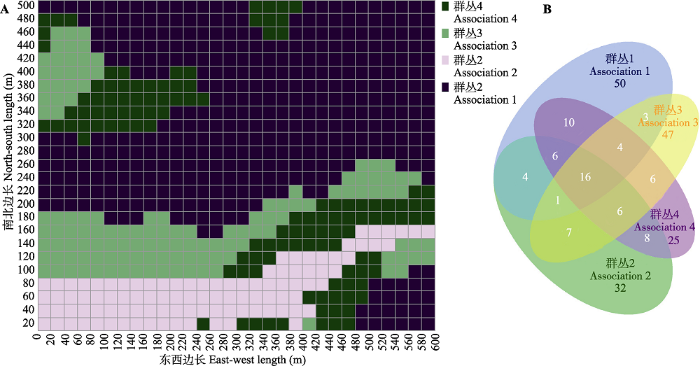 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}