申佳艳
1 , 李帅锋
1 , 黄小波
1 , 雷志全
2 , 施兴全
2 , 苏建荣
, 1 , * 1 中国林业科学研究院资源昆虫研究所, 昆明 650224 2 曲靖市国有海寨林场, 云南曲靖 655000 Radial growth responses to climate warming and drying in Pinus yunnanensis in Nanpan River Basin Jia-Yan SHEN
1 , Shuai-Feng LI
1 , Xiao-Bo HUANG
1 , Zhi-Quan LEI
2 , Xing-Quan SHI
2 , Jian-Rong SU
, 1 , * 1 Research Institute of Resource Insects, Chinese Academy of Forestry, Kunming 650224, China 2 State-owned Haizhai Forest Farm of Qujing, Qujing, Yunnan 655000, China 通讯作者: *
jianrongsu@vip.sina.com 编委: 曹敏
责任编辑: 李敏, 赵航(实习)
收稿日期: 2019-07-2
接受日期: 2019-10-22
网络出版日期: 2019-11-20
基金资助: 云南省科技创新人才计划目 (2018HC013 )
Corresponding authors: *
jianrongsu@vip.sina.com Received: 2019-07-2
Accepted: 2019-10-22
Online: 2019-11-20
Fund supported: Supported by the Yunnan Science and Technology Innovation Talent Program (2018HC013 )
摘要 全球气候变暖背景下, 西南地区气候呈现出明显的暖干化特征, 但区域优势树种云南松(
Pinus yunnanensis )对气候暖干化的响应存在不确定性。该研究根据树木年代学方法选择研究区域87株云南松样本进行树芯采集, 构建云南松树轮年表, 结合1952-2016年的气温和降水等气象资料, 利用响应分析、多元回归分析以及滑动相关分析等方法研究了影响南盘江流域云南松径向生长的关键气候因子及其对气候暖干化的响应规律。研究结果表明: 1985年以来, 研究区域气候暖干化特征明显, 气温上升和降水量下降的速率是1984年前的5和6倍, 年平均气温、年平均最高气温、年平均最低气温的上升速率为0.044、0.041和0.050 ℃·a
-1 , 年降水量的下降速率为 6.02 mm·a
-1 。气候暖干化使云南松的生长对温度响应的敏感度降低, 对水分响应的敏感度增强, 气温的解释率由暖干化前的44.95%下降到21.97%, 水分的解释率由暖干化前的55.05%上升到78.03%。暖干化增强了当年气候因子对径向生长的影响, 减弱了上年气候因子的影响, 与径向生长显著相关的当年气候因子增加了3个, 当年气候因子对径向生长的解释率增加了16.05%。暖干化减弱了云南松生长的“滞后效应”, 气候变化对树木生长影响的时效性增强。在5-7月和9-11月, 气候变暖使径向生长与气温、水分的响应关系变得不稳定。该研究可为气候暖干化区域云南松林的经营、管理以及区域气候重建提供理论依据和基础数据。
关键词: 云南松 ;
年轮 ;
气候暖干化 ;
气候响应 ;
稳定性 Abstract Aims Forests in Nanpan River Basin are under severe effects by drought because of the warming and drying climate. As the typical dominant coniferous forests in this region,
Pinus yunnanensis forests occur over a broad range and present a good potential for dendroclimatological studies. However, little is known about the relationship between radial growth and climate in this tree species. Our objectives were to determine the constraining factors of radial growth and examine the stability of growth-climate relationships in
P. yunnanensis in response to climate warming and drying.
Methods We used standardized dendrochronological method, combined with trend analysis of meteorological data from 1952 to 2016, to study the effects of climate variabilities on the radial growth in
P. yunnanensis. We extracted increment cores from 87 trees and measured annual ring-width. Response analysis and multiple regression analysis were used to determine the constraining factors of radial growth. Moving correlation was used to detect the stability of growth-climate response.
Important findings The annual mean temperature (
T mp ), mean maximum temperature (
T mx ), and mean minimum temperature (
T mn ) in the research region have increased at a rate of 0.044, 0.041 and 0.050 °C·a
-1 , respectively. The annual total precipitation showed a significant decreasing trend at a rate of 6.02 mm·a
-1 during 1985-2016. The growth of
P. yunnanensis trees was less sensitive to temperature but more sensitive to moisture; prior to and after warming and drying period, the explainable variance of radial growth by temperature decreased from 44.95% to 21.97%, and that by moisture-related factors increased from 55.05% to 78.03%. Warming and drying climate enhanced the influence of climatic factors on radial growth of the current year and weakened the influence of climatic factors of the previous year, while increasing the climatic factors of the current year significantly affecting radial growth by three and explanation rate by 16.05%. The “hysteresie effect” weakened and the effects of climate change on growth became more promptly. The growth-climate relationship in
P. yunnanensis could become more moisture sensitive under a warming and drying climate. Tree growth and forest productivity of
P. yunnanensis in the Nanpan River Basin is likely to decline if the warming and drying climate trend continues.
Keywords: Pinus yunnanensis ;
;
climate warming and drying ;
climate response ;
stability PDF (8786KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 申佳艳, 李帅锋, 黄小波, 雷志全, 施兴全, 苏建荣. 南盘江流域云南松径向生长对气候暖干化的响应. 植物生态学报, 2019, 43(11): 946-958. DOI:
10.17521/cjpe.2019.0169 SHEN Jia-Yan, LI Shuai-Feng, HUANG Xiao-Bo, LEI Zhi-Quan, SHI Xing-Quan, SU Jian-Rong.
Radial growth responses to climate warming and drying in Pinus yunnanensis in Nanpan River Basin .
Chinese Journal of Plant Ecology , 2019, 43(11): 946-958. DOI:
10.17521/cjpe.2019.0169 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题。树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 )。树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息。在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 )。基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 )。近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础。
我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 )。当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 )。近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 )。近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究。
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征。
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应。
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应。
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作。目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道。
云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 )。在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 )。云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部。本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据。
1 材料和方法 1.1 研究区域概况 本研究在南盘江流域的云南省曲靖市海寨林场进行。海寨林场位于曲靖沾益坝子周围台地上, 地理位置103.70°-104.08° E, 25.15°-25.70° N, 地处滇中高原中部、珠江源头南盘江上游两侧。地势北高南低, 地形平缓, 坡度6°-15°, 最高海拔2 577 m, 最低海拔1 915 m, 相对高差622 m。研究区冬春干旱少雨, 夏秋湿润多雨。年平均气温14.5 ℃, 最冷月(1月)平均气温7.3 ℃, 最热月(7月)平均气温19.9 ℃, 极端最高气温33 ℃, 极端最低气温-9 ℃; 年降水量1 008.9 mm, 其中6-10月降水占全年降水量的73% (
图1 ), 年蒸发量较大, 为2 140.3 mm。土壤主要为红壤、褐红壤及黄棕壤, 山坡下部土层深厚湿润, 山坡中上部和山脊土层较为瘠薄, 多裸露岩石。优势树种为云南松、华山松(
Pinus armandii ), 伴生麻栎(
Quercus acutissima )、桉树(
Eucalyptus robusta )、杉木(
Cunninghamia lanceolata )、柏木(
Cupressus funebris )、桤木(
Alnus cremastogyne )、杨树(
Populus spp.)等。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1云南曲靖沾益区气象站月平均气温、月平均最高气温、月平均最低气温和月降水量年内变化(1952-2016年)。Pre, 月降水量; T mn , 月平均最低气温; T mp , 月平均气温; T mx , 月平均最高气温。 Fig. 1Monthly mean temperature, monthly mean maximum temperature, monthly mean minimum temperature and precipitation in Zhanyi District Meteorological Station, Qujing,Yunnan (1952-2016). Pre , monthly precipitation; T mn , monthly mean minimum temperature; T mp , monthly mean temperature; T mx , monthly mean maximum temperature. 1.2 样品采集与年表建立 按照国际树木年轮库(ITRDB)标准以及树木年轮学采样原则与方法(
Fritts, 1976 ), 在海拔1 950-2 200 m范围随机选取生长良好、树木间相互遮蔽较小、立地条件较差的云南松钻取树芯。共选取87株云南松样本, 样树胸径23.0-42.6 cm, 树高9.0-19.6 m。每株样本在胸高处(1.3 m)不同方位钻取2根年轮样芯(取样方向首选南向, 另一样芯方向与第一根垂直), 共采集172根样芯。
样芯按树木年轮样品处理流程处理。干燥后用白乳胶固定在样本槽内, 用不同粗细程度的砂纸逐级打磨至光、滑、亮状态, 能在40倍显微镜下清晰观察到年轮轮廓为止。在双筒显微镜下进行目测交叉定年后, 用LINTAB 6.0年轮宽度测量仪(精度为0.001 mm)(Rinntech, Heidelberg, Germany)测量树轮原始宽度。在COFECHA程序中进行交叉定年, 精确确定每个年轮的生长年份。对去除树木生长的自相关性以及低频信号的结果进行检验, 剔除与主序列相关性差、年轮序列不完整以及难以交叉定年的样芯, 最终剩余62株样本的95根样芯进入主序列(
表1 )。用R语言dplR扩展包(
Bunn, 2010 )中的负指数函数或样条函数拟合去除因遗传因素或个体干扰产生的抑制、释放等生长趋势后, 最终得到云南松标准年表(STD)(
图2 )、差值年表(RES)和自回归年表(ARS)。STD公共区间(1954-2018年)统计特征如
表1 所示, 样本总体代表性(EPS)为0.94, 大于阈值0.85, 能较好地代表研究区云南松树轮宽度变化的基本特征; 较高的平均敏感度(MS, 0.18)和信噪比(SNR, 14.31)表明云南松对环境变化敏感, 年表质量较好, 保存了较高的气候信号。因此, 选用标准年表进行相关性分析。
Table 1 表1 表1 南盘江流域云南松树木年轮宽度标准年表(STD)统计特征及公共区间统计量
Table 1
Statistics of tree-ring width standard chronology (STD) and common interval analysis of Pinus yunanensis in Nanpan River Basin 统计特征 Statistic feature 标准年表 STD 样本量(树/样芯) Sample size (trees/radii) 62/95 年表时段 Chronology span 1954-2018 平均敏感度 Mean sensitivity 0.18 公共区间 Common period (year) 1968-2015 公共区间统计量 Statistics of common interval analysis 第一特征向量百分比 Variance in first eigenvector (%) 28.26 标准偏差 Standard deviation 0.39 一阶自相关系数 First order autocorrelation 0.125 信噪比 Signal-to-noise ratio 14.31 样本总体代表性 Expressed population signal 0.94 样芯间相关系数 Correlations between cores 0.254
新窗口打开 |
下载CSV 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2南盘江流域云南松树木年轮宽度指数(RWI )及样本量。 Fig. 2Tree ring-width index (RWI ) and sample size of Pinus yunanensis in Nanpan River Basin. 1.3 气象数据 气象资料选取距离采样点最近的曲靖沾益气象站器测气象资料。资料时段为1952-2016年(
图1 ), 其中月平均气温(
T mp )、月平均最高气温(
T mx )、月平均最低气温(
T mn )、月降水量(
Pre )、月平均相对湿度(
RH )来源于中国气象科学数据共享服务网(http://data.cma.cn/data/weatherBk.html)。帕尔默干旱指数(
PDSI, 1901-2017)来源于荷兰皇家气象局服务网(https://climexp.knmi.nl/start.cgi), 分辨率为0.5° × 0.5°。鉴于树木生长对气候响应存在“滞后效应”, 即前一年气候状况会对当年树木生长产生影响(
Fritts et al ., 1965b ), 故选取上年5月至当年11月的气象数据, 共计108个气候变量与STD年表数据进行响应分析。采用
PDSI 描述研究区干湿状况,
PDSI 是结合降水、蒸散以及干旱持续时间的综合气象干旱指标, 被广泛用于干旱评估(
卫捷和马柱国, 2003 )。-0.5 <
PDSI ≤ 0.5表示正常、-1 <
PDSI ≤ -0.5表示初旱、-2 <
PDSI ≤ -1表示轻旱、-3 <
PDSI ≤ -2表示中旱、-4 <
PDSI ≤ -3表示重旱、
PDSI ≤ -4表示极旱,
PDSI 值越小, 气候越干旱。
1.4 数据处理 运用Dendroclim 2002软件中的响应函数分析年轮宽度指数与气候数据(温度、降水、
RH 、
PDSI )的响应关系。以年轮宽度指数为预测变量、与其显著相关(
p < 0.05)的气候因子作为解释变量, 利用R软件的lm函数进行多元回归分析, 用step函数通过后向逐步回归方法优化获得简化回归模型(
van der Maaten-Theunissen et al ., 2015 )。为保证模型参数的可比性, 分析前利用scale函数对数据进行标准化。用简化回归模型中各解释变量标准化回归系数绝对值占所有解释变量回归系数绝对值总和的百分率作为气候因子对年轮生长的解释率(
Le Bagousse-Pinguet et al ., 2019 )。基于测量的树木年轮宽度计算云南松断面积增长量(
BAI ),BAI=$\pi\times(R_n^2-R_n-1^2)$, 其中
R 为云南松样树半径,
n 为年份(
Jump et al ., 2006 )。利用R语言中bootRes程序包中的mdcc函数计算滑动响应相关系数, 并利用mdcplot函数作出滑动响应色温图(
Zang & Biondi, 2013 ), 分析随气候暖干化发展与云南松径向生长显著相关的气候因子与年轮宽度响应关系的稳定性。作图用Sigmplot 10.0软件完成。
2 结果 2.1 研究区域气候变化趋势 1984年以前, 研究区温度、降水的变化较平缓; 但它们在1985年以后发生了较大的变化(
图3 )。1984年前, 年平均气温、年平均最高气温、年平均最低气温仅以小于0.008 ℃·a
-1 的速率上升, 年降水量以 0.12 mm·a
-1 的速率下降。1985年后, 年平均气温、年平均最高气温、年平均最低气温都出现了明显上升趋势, 其中年平均气温以0.044 ℃·a
-1 , 年平均最高气温以0.041 ℃·a
-1 的速率上升, 年平均最低气温上升速率达0.050 ℃·a
-1 ; 年降水量呈明显减少趋势, 以6.02 mm·a
-1 的速率下降。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3南盘江研究区域两个时期(1952-1984年, 1985-2016年)气温和降水量变化趋势。T mn , 月平均最低气温; T mp , 月平均气温; T mx , 月平均最高气温。 Fig. 3Long-term trends of temperature and precipitation during 1952-1984 and 1985-2016 in Nanpan River Basin research region. T mn , monthly mean minimum temperature; T mp , monthly mean temperature; T mx , monthly mean maximum temperature. 图4 表明, 研究区
PDSI 值在-3.35-3.61之间, 气候由湿润向干旱转变; 特殊年份
PDSI 值小于-3, 旱情达到重旱程度。1984年以前, 研究区的
PDSI 值呈升高趋势, 以每10年0.50的速率上升; 1985年以后
PDSI 值以每10年0.18的速率下降, 气候越来越干。总体上看, 研究区1985-2016年间的气温大幅升高, 降水大幅减少,
PDSI 值越来越小, 呈现出明显的暖干化特征。随着研究区气候的暖干化, 云南松断面积增长量(
BAI )以0.019 cm
2 ·a
-1 的速率减小, 云南松径向生长呈现衰退趋势。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4南盘江研究区域帕尔默干旱指数(PDSI )以及断面积增长量(BAI )随时间变化趋势。 Fig. 4Palmer Drought Standard Index (PDSI ) and basal area increment (BAI ) over time in Nanpan River Basin research region. 2.2 云南松年轮宽度与主要气候因子的关系 云南松年轮宽度与上年5、7月的平均气温、平均最高气温以及上年9月和当年9、11月的平均最低气温显著正相关(
p < 0.05), 与上年12月平均最高气温显著负相关(
p < 0.05)(
图5A ); 年轮宽度与当年春、夏的
PDSI 指数显著正相关(
p < 0.05), 而与上年7月的降水量、平均相对湿度显著负相关(
p < 0.05)(
图5B )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5南盘江云南松年轮宽度年表与主要气候因子的响应关系。PDSI, 帕尔默干旱指数; Pre, 月降水量; RH, 平均相对湿度; T mn , 月平均最低气温; T mp , 月平均气温; T mx , 月平均最高气温。 Fig. 5Relationships of ring-width chronology of Pinus yunanensis with major climatic factors in Nanpan River Basin. PDSI, Palmer Drought Standard Index; Pre, monthly precipitation; RH, average air relative humidity; T mn , monthly mean minimum temperature; T mp , monthly mean temperature; T mx , monthly mean maximum temperature. 2.3 云南松径向生长对气候暖干化的响应 相关分析(
图6 )表明, 暖干化前, 年轮宽度与上年7月平均气温、平均最低气温、上年10月平均相对湿度、当年1和6月的平均相对湿度显著正相关(
p < 0.05), 而与上年12月的平均气温、平均最低气温和平均最高气温显著负相关(
p < 0.05)。暖干化后, 年轮宽度与上年7月平均气温、平均最低气温、上年9月平均最低气温、当年11月平均最低气温以及当年6、7、10、11月的
PDSI 显著正相关(
p < 0.05); 而与上年7月平均相对湿度显著负相关(
p < 0.05)。气候暖干化增强了云南松径向生长对水分因子的响应, 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子, 原来与年轮宽度不显著相关的当年7、10、11月
PDSI 转变为显著相关因子。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6南盘江树轮宽度年表与不同时段逐月气候因子的响应关系。PDSI, 帕尔默干旱指数; Pre, 月降水量; RH, 平均相对湿度; T mn , 月平均最低气温; T mp , 月平均气温; T mx , 月平均最高气温。 Fig. 6Response relationships of tree ring-width chronology with monthly climatic factors during different time periods in Nanpan River Basin. Pre, monthly precipitation; PDSI, Palmer Drought Standard Index; RH, average air relative humidity; T mn , monthly mean minimum temperature; T mp , monthly mean temperature; T mx , monthly mean maximum temperature. 多元回归分析表明, 云南松对气候变化的响应方式发生了一定程度变化。暖干化前, 年轮宽度指数与气候因子的简化回归模型为:
RWI STD = 0.352 × p
T mn 7 - 0.235 × p
T mx 12 + 0.207 × c
RH 1 + 0.252 × c
RH 6 + 0.261 × p
RH 10 (
R 2 = 0.543 2,
p < 0.001)
暖干化后, 年轮宽度指数与气候因子的最优回归模型为:
RWI STD = 0.289 × p
T mp 7 - 0.354 × p
RH 7 + 0.316 × c
PDSI 6 + 0.358 × c
PDSI 11 (
R 2 = 0.533 9,
p < 0.001)
式中,
RWI STD 为标准年表年轮宽度指数, p
T mn 7为上年7月平均最低气温, p
T mx 12为上年12月平均最高气温, c
RH 1和c
RH 6分别为当年1、6月平均相对湿度, p
RH 7和p
RH 10分别为上年7、10月平均相对湿度, c
PDSI 6和c
PDSI 11为当年6和11月的
PDSI 。
暖干化前后, 入选简化回归模型的气候因子及其对云南松径向生长的解释率如
图7 所示。暖干化前, 上年7月平均最低气温、12月平均最高气温, 当年1、6月和上年10月平均相对湿度对云南松径向生长的解释率分别为26.92%、18.03%、15.82%、19.26%和19.97%。暖干化后, 上年7月平均气温、平均相对湿度和当年6、11月的
PDSI 对云南松径向生长的解释率分别为21.97%、26.90%、23.96%、27.17%。气候暖干化后, 温度因子对径向生长的解释率由暖干化前的44.95%下降到21.97%, 水分因子的解释率由55.05%上升到78.03%。暖干化后, 上年度气候因子对径向生长的解释率由64.92%降到48.87%, 当年气候因子的解释率由35.08%上升到51.13%。暖干化前后, 上年7月平均气温和平均最低气温、当年6月平均相对湿度和
PDSI 对径向生长的解释率较稳定, 变化幅度仅在±5%左右。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7简化回归模型中气候因子对云南松径向生长的解释率。cPDSI 11, 当年11月帕尔默干旱指数; cRH 1, 当年1月平均相对湿度; cRH 6, 当年6月平均相对湿度; pRH 7, 上年7月平均相对湿度; pRH 10, 上年10月平均相对湿度; pT mn 7, 上年7月平均最低气温; pT mp 7, 上年7月平均气温; pT mx 12, 上年12月平均温最高气温。 Fig. 7Pie chart of variance in radial growth in Pinus yunanensis explainable by different climatic factors in optimized regression model. CPDSI 11, Palmer Drought Standard Index of current November; cRH 1, mean relative humidity of current January; cRH 6, mean relative humidity of current June; pRH 7, mean relative humidity of last July; pRH 10, mean relative humidity of last October; pT mn 7, mean minimum temperature of last July; pT mp 7, mean temperature of last July; pT mx 12, mean maximum temperature of last December. 2.4 云南松径向生长对气候暖干化的响应稳定性 滑动响应分析图(8A)表明, 云南松径向生长与上年5、6月平均气温的响应关系在1971-1995年分别呈现明显的减弱、增强趋势, 存在较强的不稳定性。云南松径向生长与上年7月平均气温的正响应关系在1969-2004年间有所增强, 但总体稳定性较好。云南松径向生长与上年12月、当年1、5月平均气温的负响应关系在1973-1997年明显减弱, 与当年3月平均气温的负响应关系在2004年之后明显增强, 呈现较强的不稳定性。云南松径向生长与当年6、7月平均气温的负响应关系呈现年际波动。
图8 新窗口打开 |
下载原图ZIP |
生成PPT 图8云南松年轮宽度年表与上年5月至当年11月主要气候因子的滑动响应分析, 滑动窗口为25年, 偏移1年。A, 云南松径向生长与月平均气温(T mp )的滑动响应分析。B, 云南松径向生长与月降水量(Pre )的滑动响应分析。C, 云南松径向生长与帕尔默干旱指数(PDSI )的滑动响应分析。 Fig. 8Sliding response analysis of ring width chronology in Pinus yunanensis with main climatic factors from May of the previous year to November of the current year. The window size is 25 years, and the windows have been offset by one year. A, Sliding response analysis of radial growth and monthly mean temperature (T mp ). B, Sliding response analysis of radial growth and monthly precipitation (Pre ). C, Sliding response analysis of radial growth and Palmer Drought Standard Index (PDSI ). 滑动响应分析图(8B)表明, 云南松径向生长与上年6月降水量的正响应关系在1999年后增强, 与上年7月降水量的负响应关系在1968-1998年间明显减弱, 与上年9、10月降水量的响应关系在1969- 1993年不稳定, 分别发生由负响应向正响应、正响应向负响应的转变。云南松径向生长与上年12月降水量的正响应关系, 与当年8月降水量的负响应关系在1969-1993年间明显增强, 呈现明显的不稳定性。
滑动响应分析图(8C)显示, 云南松径向生长与当年4月水热条件的正响应关系在1976-2000年间明显减弱, 与当年5月水热条件的正响应关系基本稳定, 与当年6月水热条件的正响应关系在2000年后明显增强, 与当年8月水热条件的负响应关系在1997年之后明显增强, 与当年9、10月水热条件的响应关系在1992年之后发生由正响应向负响应的转变, 与当年11月水热条件的响应关系在1997年发生了从负响应向正响应的转变。
3 讨论 3.1 云南松径向生长对气候因子的响应 研究区5、7和12月的水热条件是云南松径向生长的关键因素。7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用。此时, 高温就可促进云南松的生长。研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 )。这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子。夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 )。12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 )。此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害。本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份。这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致。原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长。这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式。这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径。
3.2 暖干化对云南松径向生长的影响 暖干化前后, 6、7月气候因子的影响比较稳定, 它们对径向生长的解释率都在45%左右, 由此可见,暖干化并没有改变研究区6和7月的气候特点, 云南松生长旺盛期的径向生长也相对稳定。在水分条件不受限制的情况下(夏季6、7、8月)增温作用不会对树木生长产生抑制作用, 反而能够缓解水分过多产生的胁迫作用, 提高光合作用效率(
李宗善等, 2010 )。这与研究区6、7月水热条件优越, 云南松生长迅速的实地观测结果(
图1 )一致。
暖干化后, 与云南松径向生长显著相关的水分因子增加较多, 说明随着研究区温度的升高, 水分因子对径向生长越来越重要。暖干化后, 水分和温度因子对径向生长解释率的增减均达23%, 云南松对水分因子的响应也越来越敏感(
Bryukhanova et al ., 2013 )。这与
Babst等(2019) 得出的全球气候变暖会导致北方森林树木对温度敏感转变为对水分更加敏感的结论一致, 但与全球气候变暖会增强树木生长对温度敏感性的结论(
Andreu et al ., 2007 )不符。其原因可能是, 研究区气候暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出; 气候暖干化使云南松径向生长对气候变化的响应模式发生了从响应温度胁迫到响应水分胁迫的转变。这说明, 在评估气候变暖对林木生长的影响时, 还应考虑升温导致的水分变化, 否则可能会产生较大的偏差。
暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降。主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 )。(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%。(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现。云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%。可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性。
3.3 云南松径向生长对暖干化响应的稳定性 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间。说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变。
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势。
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈。
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定。
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性。在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变。本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段。树木生长与气候因子的动态关系一直是树木年轮学研究的热点。针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 )。本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考。
4 结论 综上表明, 近30年来南盘江流域气候呈现出明显的暖干化特征。气候暖干化使云南松径向生长对气候变化的响应模式发生了从响应温度胁迫到响应水分胁迫的转变。气候变暖可以促进云南松的径向生长, 但促进效应随着云南松生长对温度的响应减弱而越来越弱。由升温和低降水引发的干旱胁迫是限制研究区云南松径向生长的主要因素。在区域干旱化趋势下, 云南松的生长可能会减慢, 对森林生产力产生不利影响。本研究表明, 树木生长与气候因子间的关系会随气候变化发生一定程度的改变, 这可能是树木适应环境变化的一种方式。另一方面, 各个气候要素对树木生长产生的作用不是单一的, 研究树木生长与气候因子关系时应该考虑气候要素间的综合效应, 增强研究的可靠性。本研究证实气候变暖会改变树木生长与气候要素间关系的稳定性, 在利用树轮进行气候重建研究时应先检验树木生长与气候响应关系的稳定性, 以提高气候重建的可信度。本研究区域是云南松在云南高原的典型分布区, 代表了滇中地区海拔1 500-2 800 m范围内云南松的情况, 研究结果可为该区域云南松林的气候适应性管理和区域气候重建提供理论依据和参考数据。
致谢 感谢中国林业科学院基本科研业务费专项(CAFYBB2017ZX002)资助。
[1] Andreu L Gutiérrez E Macias M Ribas M Bosch O Camarero JJ 2007 ). Climate increases regional tree-growth variability in Iberian pine forestsGlobal Change Biology 804 -815 . [本文引用: 1] [2] Babst F Bouriaud O Poulter B Trouet V Girardin MP Frank DC 2019 ). Twentieth century redistribution in climatic drivers of global tree growthScience Advances 10.1126/sciadv.aat4313 . [本文引用: 1] [3] Bryukhanova M Fonti P Kirdyanov A Saurer M Siegwolf R Pochebit N Sidorova O Prokushkin A 2013 ). Growth of trees on permafrost: Habitat driven response to climateGeophysical Research Abstracts [本文引用: 1] [4] Bunn AG 2010 ). Statistical and visual crossdating in R using the dplR libraryDendrochronologia 251 -258 . [本文引用: 1] [5] Chang YX Chen ZJ Zhang XL Bai XP Zhao XP Li JX Lu X 2017 ). Responses of radial growth to temperature in Larix gmelinii of the Da Hinggan Ling under climate warmingChinese Journal of Plant Ecology, 279 -289 . [本文引用: 1] [ 常永兴 , 陈振举 , 张先亮 , 白学平 , 赵学鹏 , 李俊霞 , 陆旭 ( 2017 ). 气候变暖下大兴安岭落叶松径向生长对温度的响应植物生态学报 279 -289 .] [本文引用: 1] [6] Chen XQ Hu B Yu R 2005 ). Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern ChinaGlobal Change Biology 1118 -1130 . [本文引用: 1] [7] Cleaveland MK Stahle DW Therrell MD Villanueva-Diaz J Burns BT 2003 ). Tree-ring reconstructed winter precipitation and tropical teleconnections in Durango, MexicoClimatic Change 369 -388 . [本文引用: 1] [8] Cullen LE Palmer JG Duncan RP Stewart GH 2001 ). Climate change and tree-ring relationships of Nothofagus menziesii tree-line forestsCanadian Journal of Forest Research, 1981 -1991 . [本文引用: 1] [9] Deng XQ Huang BL Wen QZ Hua CL Tao J 2013 ). A research on the distribution of Pinus yunnanensis forest in Yunnan ProvinceJournal of Yunnan University (Natural Science Edition )843 -848 . [本文引用: 1] [ 邓喜庆 , 皇宝林 , 温庆忠 , 华朝朗 , 陶晶 ( 2013 ). 云南松林在云南的分布研究云南大学学报(自然科学版) 843 -848 .] [本文引用: 1] [10] Deng XQ Huang BL Wen QZ Hua CL Tao J Zheng JX 2014 ). Dynamic of Pinus yunnanensis forest resources in YunnanJournal of Natural Resources, 1411 -1419 . [本文引用: 1] [ 邓喜庆 , 皇宝林 , 温庆忠 , 华朝朗 , 陶晶 , 郑进烜 ( 2014 ). 云南松林资源动态研究自然资源学报 1411 -1419 .] [本文引用: 1] [11] Deng Y Gou XH Gao LL Zhang F Xu XY Yang MX 2016 ). Tree-ring recorded drought variability in the northern Min Mountains of northwestern ChinaInternational Journal of Climatology 3550 -3560 . [本文引用: 1] [12] Du S Yamanaka N Yamamoto F Otsuki K Wang SQ Hou QC 2007 ). The effect of climate on radial growth of Quercus liaotungensis forest trees in Loess Plateau, ChinaDendrochronologia, 29 -36 . [本文引用: 1] [13] Fan ZX Br?uning A Cao KF Zhu SD 2009 ). Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern ChinaForest Ecology and Management 306 -313 . [本文引用: 1] [14] Fan ZX Br?uning A Thomas A Li JB Cao KF 2011 ). Spatial and temporal temperature trends on the Yunnan Plateau (southwest China) during 1961-2004International Journal of Climatology 2078 -2090 . [本文引用: 1] [15] Fang KY Gou XH Chen FH Peng JF Liu PX Wang M Zhu YM 2008 ). The advance in dendroecologyJournal of Glaciology and Geocryology 825 -834 . [本文引用: 1] [ 方克艳 , 勾晓华 , 陈发虎 , 彭剑峰 , 刘普幸 , 王茂 , 朱有明 ( 2008 ). 树轮生态学研究进展冰川冻土 825 -834 .] [本文引用: 1] [16] Forest Resources Management Department of the State Forestry Administration ( 2010 ). The 7th national forest resources inventory and forest resources statusForest Resources Management 1 ), 1 -8 . [本文引用: 1] [ 国家林业局森林资源管理司 ( 2010 ). 第七次全国森林资源清查及森林资源状况林业资源管理 1 ), 1 -8 .] [本文引用: 1] [17] Fritts HC 1966 ). Growth-rings of trees: Their correlation with climateScience 973 -979 . [本文引用: 2] [18] Fritts HC 1976 ). Tree-Rings and Climate Elsevier, New York . [本文引用: 2] [19] Fritts HC Smith DG Cardis JW Budelsky CA 1965 b). Tree-ring characteristics along a vegetation gradient in Northern ArizonaEcology 393 -401 . [本文引用: 2] [20] Fritts HC Smith DG Stokes MA 1965 a). The biological model for paleoclimatic interpretation of mesa verde tree-ring seriesMemoirs of the Society for American Archaeology 101 -121 . [本文引用: 1] [21] Graumlich LJ 1991 ). Subalpine tree growth, climate, and increasing CO2 : An assessment of recent growth trendsEcology 1 -11 . [本文引用: 1] [22] Guo BD 2016 ). Differences in Response to Climate Change of Picea purpurea and Abies faxoniana at Different Elevations and Slopes in Western Sichuan Plateau, China PhD dissertation, Northeast Forestry University, Harbin. [本文引用: 1] [ 郭滨德 ( 2016 ). 川西高原不同坡向和海拔云冷杉树轮对气候变化的响应差异博士学位论文, 东北林业大学, 哈尔滨 .] [本文引用: 1] [23] Jia CQ Dong YJ Peng JT Rong YS Wang L 2014 ). Spatial- temporal laws of droughts and floods cross Nanpan River BasinJournal of Arid Land Resources and Environment 104 -108 . [本文引用: 1] [ 贾春强 , 董延军 , 彭俊台 , 荣艳淑 , 王琳 ( 2014 ). 南盘江流域旱涝时空分布规律研究干旱区资源与环境 104 -108 .] [本文引用: 1] [24] Jiang P Liu H Wu XC Wang HY 2017 ). Tree-ring-based SPEI reconstruction in central Tianshan Mountains of China since A.D. 1820 and links to westerly circulationInternational Journal of Climatology 2863 -2872 . [本文引用: 1] [25] Jump AS Hunt JM Penuelas J 2006 ). Rapid climate change- related growth decline at the southern range edge of Fagus sylvaticaGlobal Change Biology 2163 -2174 . [本文引用: 1] [26] Komer , Christian ( 2001 ). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems Springer, Berlin . [本文引用: 2] [27] Kujansuu J Yasue K Koike T Abaimov AP Kajimoto T Takeda T Tokumoto M Matsuura Y 2007 ). Climatic responses of tree-ring widths of Larix gmelinii on contrasting north-facing and south-facing slopes in central SiberiaJournal of Wood Science, 87 -93 . [本文引用: 1] [28] Le Bagousse-Pinguet Y Soliveres S Gross N Torices R Berdugo M Maestre FT 2019 ). Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionalityProceedings of the National Academy of Sciences of the United States of America 8419 -8424 . [本文引用: 1] [29] Li T He XY Chen ZJ 2014 ). Response of radial growth of Quercus mongolica to climate change in the south of northeast China—Taking Qianshan as an exampleChinese Journal of Applied Ecology 1841 -1848 . [本文引用: 1] [ 李腾 , 何兴元 , 陈振举 ( 2014 ). 东北南部蒙古栎径向生长对气候变化的响应——以千山为例应用生态学报 1841 -1848 .] [本文引用: 1] [30] Li ZS Liu GH Zhang QB Hu CJ Luo SZ Liu XL He F 2010 ). Tree ring reconstruction of summer temperature variations over the past 159 years in Wolong National Natural Reserve, western Sichuan, ChinaChinese Journal of Plant Ecology 628 -641 . [本文引用: 2] [ 李宗善 , 刘国华 , 张齐兵 , 胡婵娟 , 罗淑政 , 刘兴良 , 何飞 ( 2010 ). 利用树木年轮宽度资料重建川西卧龙地区过去159年夏季温度的变化植物生态学报 628 -641 .] [本文引用: 2] [31] Liu YJ Gu ZX Wang XC 2017 ). Impact of simulated climate warming on the radial growth of Larix gmelinii in northeast ChinaActa Ecologica Sinica, 2684 -2693 . [本文引用: 1] [ 刘玉佳 , 顾卓欣 , 王晓春 ( 2017 ). 模拟气候变暖对东北兴安落叶松径向生长的影响生态学报 2684 -2693 .] [本文引用: 1] [32] Ma ZG Ren XB 2007 ). Drying trend over China from 1951 to 2006Advances in Climate Change Research 195 -201 . [本文引用: 1] [ 马柱国 , 任小波 ( 2007 ). 1951-2006年中国区域干旱化特征气候变化研究进展 195 -201 .] [本文引用: 1] [33] Nie CY Zhang QB Lyu L 2017 ). Millennium-long tree-ring chronology reveals megadroughts on the southeastern Tibetan PlateauTree-Ring Research 1 -10 . [本文引用: 1] [34] Panthi S Br?uning A Zhou ZK Fan ZX 2018 ). Growth response of Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern ChinaDendrochronologia, 1 -9 . [本文引用: 3] [35] Rathgeber C Nicault A Guiot J Keller T Guibal F Roche P 2000 ). Simulated responses of Pinus halepensis forest productivity to climatic change and CO2 increase using a statistical modelGlobal and Planetary Change, 405 -421 . [本文引用: 1] [36] Shao XM Wu XD 1994 ). Tree-ring chronologies for Pinus armandi franch from Huashan, ChinaActa Geographica Sinica, 174 -181 . [本文引用: 1] [ 邵雪梅 , 吴祥定 ( 1994 ). 华山树木年轮年表的建立地理学报 174 -181 .] [本文引用: 1] [37] Shao XM Wu XD 1997 ). Reconstruction of climate change on Changbai Mountain, northeast China using tree-ring dataQuaternary Studies 76 -85 . [本文引用: 2] [ 邵雪梅 , 吴祥定 ( 1997 ). 利用树轮资料重建长白山区过去气候变化第四纪研究 76 -85 .] [本文引用: 2] [38] Shen CC Wang L Li MY 2016 ). The altitudinal variability and temporal instability of the climate-tree-ring growth relationships for Changbai larch ( Larix olgensis Henry) in the Changbai mountains area, Jilin, Northeastern ChinaTrees, 901 -912 . [本文引用: 1] [39] Su XC Wang L Li QL 2014 ). Study on surface dry and wet conditions in southwest China in recent 50 yearsJournal of Natural Resources 104 -116 . [本文引用: 1] [ 苏秀程 , 王磊 , 李奇临 ( 2014 ). 近50a中国西南地区地表干湿状况研究自然资源学报 104 -116 .] [本文引用: 1] [40] van der Maaten-Theunissen M van der Maaten E Bouriaud O 2015 ). PointRes: An R package to analyze pointer years and components of resilienceDendrochronologia 34 -38 . [本文引用: 1] [41] Wang T Yu D Li JF Ma KP 2003 ). Advances in research on the relationship between climatic change and tree-ring widthActa Phytoecologica Sinica 23 -33 . [本文引用: 1] [ 王婷 , 于丹 , 李江风 , 马克平 ( 2003 ). 树木年轮宽度与气候变化关系研究进展植物生态学报 23 -33 .] [本文引用: 1] [42] Wang WZ Jia M Wang GX Zhu WZ McDowell NG 2017 ). Rapid warming forces contrasting growth trends of subalpine fir ( Abies fabri ) at higher- and lower-elevations in the eastern Tibetan PlateauForest Ecology and Management, 135 -144 . [本文引用: 1] [43] Wang XC Song LP Zhang YD 2011 ). Climate-tree growth relationships of Pinus sylvestris var. mongolica in the northern Daxing’an Mountains, ChinaChinese Journal of Plant Ecology, 294 -302 . [本文引用: 2] [ 王晓春 , 宋来萍 , 张远东 ( 2011 ). 大兴安岭北部樟子松树木生长与气候因子的关系植物生态学报 294 -302 .] [本文引用: 2] [44] Wang Y Yan XD 2006 ). The response of the forest ecosystem in China to global climate changeChinese Journal of Atmospheric Sciences 1009 -1018 . [本文引用: 1] [ 王叶 , 延晓冬 ( 2006 ). 全球气候变化对中国森林生态系统的影响大气科学 1009 -1018 .] [本文引用: 1] [45] Wei J Ma ZG 2003 ). Comparison of palmer drought severity index, percentage of precipitation anomaly and surface humid indexActa Geographica Sinica 117 -124 . [本文引用: 1] [ 卫捷 , 马柱国 ( 2003 ). Palmer干旱指数、地表湿润指数与降水距平的比较地理学报 117 -124 .] [本文引用: 1] [46] Wu XD 1990 ). Application of tree ring analysis to the study on environment variationQuaternary Sciences 188 -196 . [本文引用: 1] [ 吴祥定 ( 1990 ). 树木年轮分析在环境变化研究中的应用第四纪研究 188 -196 .] [本文引用: 1] [47] Wu XD Shao XM 1993 ). Status of dendroclimatological study and its prospects in ChinaAdvance in Earth Sciences 31 -35 . [本文引用: 1] [ 吴祥定 , 邵雪梅 ( 1993 ). 中国树木年轮气候学研究动态与展望地球科学进展 31 -35 .] [本文引用: 1] [48] Yang B He MH Shishov V Tychkov I Vaganov E Rossi S Ljungqvist FC Br?uning A Grie?inger J 2017 ). New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring dataProceedings of the National Academy of Sciences of the United States of America 6966 -6971 . [本文引用: 2] [49] Yang ML Wang L Gao R Yang R Fu B 2013 ). Temporal and spatial distribution characteristics of seasonal drought in the Nanpan River BasinPeople’s Yangtze River 11 ), 5 -8 . [本文引用: 1] [ 杨茂灵 , 王龙 , 高瑞 , 杨蕊 , 付奔 ( 2013 ). 南盘江流域季节性干旱时空分布特征研究人民长江 11 ), 5 -8 .] [本文引用: 1] [50] Yang RQ Fan ZX Li ZS Wen QZ 2018 ). Radial growth of Pinus yunnanensis at different elevations and their responses to climatic factors in the Yulong Snow Mountain, Northwest Yunnan, ChinaActa Ecologica Sinica, 8983 -8991 . [本文引用: 1] [ 杨绕琼 , 范泽鑫 , 李宗善 , 温庆忠 ( 2018 ). 滇西北玉龙雪山不同海拔云南松径向生长对气候因子的响应生态学报 8983 -8991 .] [本文引用: 1] [51] Yao YB Zhang Q Wang JS Shang JL Wang Y Shi J Han LY 2014 ). Response characteristics of drought in southwest China to climate warmingJournal of Ecology and Environment 1409 -1417 . [本文引用: 1] [ 姚玉璧 , 张强 , 王劲松 , 尚军林 , 王莺 , 石界 , 韩兰英 ( 2014 ). 中国西南干旱对气候变暖的响应特征生态环境学报 1409 -1417 .] [本文引用: 1] [52] Yin DC Sun M Zhang WG Yue HT Zhang Y Tian K Xiao DR Zhang Y 2019 ). Effects of climate warming on radial growth of Picea brachytyla in Shangri-La, southwestern ChinaJournal of Northeast Forestry University 3 ), 3 -9 . [本文引用: 1] [ 尹定财 , 孙梅 , 张卫国 , 岳海涛 , 张勇 , 田昆 , 肖德荣 , 张贇 ( 2019 ). 气候变暖对香格里拉油麦吊云杉径向生长的影响东北林业大学学报 3 ), 3 -9 .] [本文引用: 1] [53] Yin H Li YH 2013 ). Summary of advance on drought study in southwest ChinaJournal of Arid Meteorology 182 -193 . [本文引用: 1] [ 尹晗 , 李耀辉 ( 2013 ). 我国西南干旱研究最新进展综述干旱气象 182 -193 .] [本文引用: 1] [54] Yu DP Liu JQ Benard J L Zhou L Zhou WM Fang XM Wei YW Jiang SW Dai LM 2013 ). Spatial variation and temporal instability in the climate-growth relationship of Korean pine in the Changbai Mountain region of Northeast ChinaForest Ecology and Management 96 -105 . [本文引用: 2] [55] Yu DP Wang SZ Tang LN Dai LM Wang QL Wang SX 2005 ). Relationship between tree-ring chronology of Larix olgensis in Changbai Mountains and the climae changeChinese Journal of Applied Ecology, 14 -20 . [本文引用: 1] [ 于大炮 , 王顺忠 , 唐立娜 , 代力民 , 王庆礼 , 王绍先 ( 2005 ). 长白山北坡落叶松年轮年表及其与气候变化的关系应用生态学报 14 -20 .] [本文引用: 1] [56] Yu J Liu QJ Zhou G Meng SW Zhou H Xu ZZ Shi JN Du WX 2017 ). Response of radial growth of Pinus koraiensis and Picea jezoensis to climate change in Xiaoxing’anling Mountains, Northeast ChinaChinese Journal of Applied Ecology 3451 -3460 . [本文引用: 2] [ 于健 , 刘琪璟 , 周光 , 孟盛旺 , 周华 , 徐振招 , 史景宁 , 杜文先 ( 2017 ). 小兴安岭红松和鱼鳞云杉径向生长对气候变化的响应应用生态学报 3451 -3460 .] [本文引用: 2] [57] Yu JL Zhang WG Tian K Song WH Li QP Yang R Zhang Y 2017 ). Response of radial growth of three conifer trees to climate change at their upper distribution limits in Potatso National Park, Shangri-La, southwestern ChinaJournal of Beijing Forestry University 1 ), 43 -51 . [本文引用: 2] [ 余佳霖 , 张卫国 , 田昆 , 松卫红 , 李秋平 , 杨荣 , 张赟 ( 2017 ). 普达措国家公园海拔上限3个针叶树种径向生长对气候变化的响应北京林业大学学报 1 ), 43 -51 .] [本文引用: 2] [58] Yuan YJ Li JF 1999 ). Reconstruction and analysis of 450 years’ winter temperature series in the Urumqi River source of Tianshan MountainsJournal of Glaciolgy and Geocryology 1 ), 64 -70 . [本文引用: 1] [ 袁玉江 , 李江风 ( 1999 ). 天山乌鲁木齐河源450 a冬季温度序列的重建与分析冰川冻土 1 ), 64 -70 .] [本文引用: 1] [59] Zang C Biondi F 2013 ). Dendroclimatic calibration in R: The bootRes package for response and correlation function analysisDendrochronologia 68 -74 . [本文引用: 1] [60] Zhang WG Xiao DR Tian K Chen GL He RH Zhang Y 2017 ). Response of radial growth of three conifer species to climate at their respective upper distributional limits on Yulong snow MountainActa Ecologica Sinica 3796 -3804 . [本文引用: 1] [ 张卫国 , 肖德荣 , 田坤 , 陈广磊 , 和荣华 , 张赟 ( 2017 ). 玉龙雪山3个针叶树种在海拔上限的径向生长及气候响应生态学报 3796 -3804 .] [本文引用: 1] [61] Zhang XL Cui MX Ma YJ Wu T Chen ZJ Ding WH 2010 ). Larix gmelinii tree-ring width chronology and its responses to climate change in Kuduer, Great Xing’an MountainsChinese Journal of Applied Ecology 2501 -2507 . [本文引用: 1] [ 张先亮 , 崔明星 , 马艳军 , 吴涛 , 陈振举 , 丁玮航 ( 2010 ). 大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系应用生态学报 2501 -2507 .] [本文引用: 1] [62] Zhang Y Yin DC Sun M Li LP Tian K Zhang WG 2018 a). Radial growth response of two conifers to temperature and precipitation at upper forest limits in Shika Snow Mountain, Northwestern Yunnan PlateauActa Ecologica Sinica 2442 -2449 . [本文引用: 2] [ 张赟 , 尹定财 , 孙梅 , 李丽萍 , 田昆 , 张卫国 ( 2018 a). 滇西北石卡雪山2个针叶树种森林上限径向生长对温度和降水的响应生态学报 2442 -2449 .] [本文引用: 2] [63] Zhang Y Yin DC Tian K He RH He MZ Li YC Sun DC Zhang WG 2018 b). Relationship between radial growth of Abies georgei and climate factors at different altitudes on the eastern slope of Yulong Snow Mountain, ChinaChinese Journal of Applied Ecology, 2355 -2361 . [本文引用: 2] [ 张贇 , 尹定财 , 田昆 , 和荣华 , 和茂珍 , 李玉春 , 孙大成 , 张卫国 ( 2018 b). 玉龙雪山东坡不同海拔长苞冷杉径向生长与气候因子的关系应用生态学报 2355 -2361 .] [本文引用: 2] [64] Zhang Y Yin DC Zhang WG Yue HT Du JCD Li QP Yang R Tian K 2018 c). Response of radial growth of two conifers to temperature and precipitation in Potatso National Park, Southwest ChinaActa Ecologica Sinica 5383 -5392 . [本文引用: 1] [ 张贇 , 尹定财 , 张卫国 , 岳海涛 , 杜杰次丹 , 李秋平 , 杨荣 , 田昆 ( 2018 c). 普达措国家公园2个针叶树种径向生长对温度和降水的响应生态学报 5383 -5392 .] [本文引用: 1] [65] Zhao ZP Wu XP Li G Li JS 2015 ). Drought in southwestern China and its impact on the net primary productivity of vegetation from 2009-2011Acta Ecologica Sinica 350 -360 . [本文引用: 1] [ 赵志平 , 吴晓莆 , 李果 , 李俊生 ( 2015 ). 2009-2011年我国西南地区旱灾程度及其对植被净初级生产力的影响生态学报 350 -360 .] [本文引用: 1] [66] Zheng JY Ge QS Hao ZX Wang WC 2006 ). Spring phenophases in recent decades over Eastern China and its possible link to climate changesClimatic Change 449 -462 . [本文引用: 1] [67] Zhu BL Zhao EX Zhao G Wan YX 2015 ). Interannual/ interdecadal variabilities and trend forecasting on Nanpanjiang Basin in the major flooding seasonJournal of Yunnan University (Natural Science Edition )861 -869 . [本文引用: 1] [ 朱保林 , 赵尔旭 , 赵刚 , 万云霞 ( 2015 ). 南盘江流域主汛期降雨量年(代)际变率研究及趋势预测云南大学学报(自然科学版) 861 -869 .] [本文引用: 1] [68] Zhu JH Hou ZH Zhang ZJ Luo YJ Zhang XQ 2007 ). Climate change and forest ecosystem: Impacts, vulnerability and adaptationScientia Silvae Sinicae 11 ), 138 -145 . [本文引用: 1] [ 朱建华 , 侯振宏 , 张治军 , 罗云建 , 张小全 ( 2007 ). 气候变化与森林生态系统:影响、脆弱性与适应性林业科学 11 ), 138 -145 .] [本文引用: 1] Climate increases regional tree-growth variability in Iberian pine forests
1
2007
... 暖干化后, 与云南松径向生长显著相关的水分因子增加较多, 说明随着研究区温度的升高, 水分因子对径向生长越来越重要.暖干化后, 水分和温度因子对径向生长解释率的增减均达23%, 云南松对水分因子的响应也越来越敏感(
Bryukhanova et al ., 2013 ).这与
Babst等(2019) 得出的全球气候变暖会导致北方森林树木对温度敏感转变为对水分更加敏感的结论一致, 但与全球气候变暖会增强树木生长对温度敏感性的结论(
Andreu et al ., 2007 )不符.其原因可能是, 研究区气候暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出; 气候暖干化使云南松径向生长对气候变化的响应模式发生了从响应温度胁迫到响应水分胁迫的转变.这说明, 在评估气候变暖对林木生长的影响时, 还应考虑升温导致的水分变化, 否则可能会产生较大的偏差. ...
Twentieth century redistribution in climatic drivers of global tree growth
1
2019
... 暖干化后, 与云南松径向生长显著相关的水分因子增加较多, 说明随着研究区温度的升高, 水分因子对径向生长越来越重要.暖干化后, 水分和温度因子对径向生长解释率的增减均达23%, 云南松对水分因子的响应也越来越敏感(
Bryukhanova et al ., 2013 ).这与
Babst等(2019) 得出的全球气候变暖会导致北方森林树木对温度敏感转变为对水分更加敏感的结论一致, 但与全球气候变暖会增强树木生长对温度敏感性的结论(
Andreu et al ., 2007 )不符.其原因可能是, 研究区气候暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出; 气候暖干化使云南松径向生长对气候变化的响应模式发生了从响应温度胁迫到响应水分胁迫的转变.这说明, 在评估气候变暖对林木生长的影响时, 还应考虑升温导致的水分变化, 否则可能会产生较大的偏差. ...
Growth of trees on permafrost: Habitat driven response to climate
1
2013
... 暖干化后, 与云南松径向生长显著相关的水分因子增加较多, 说明随着研究区温度的升高, 水分因子对径向生长越来越重要.暖干化后, 水分和温度因子对径向生长解释率的增减均达23%, 云南松对水分因子的响应也越来越敏感(
Bryukhanova et al ., 2013 ).这与
Babst等(2019) 得出的全球气候变暖会导致北方森林树木对温度敏感转变为对水分更加敏感的结论一致, 但与全球气候变暖会增强树木生长对温度敏感性的结论(
Andreu et al ., 2007 )不符.其原因可能是, 研究区气候暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出; 气候暖干化使云南松径向生长对气候变化的响应模式发生了从响应温度胁迫到响应水分胁迫的转变.这说明, 在评估气候变暖对林木生长的影响时, 还应考虑升温导致的水分变化, 否则可能会产生较大的偏差. ...
Statistical and visual crossdating in R using the dplR library
1
2010
... 样芯按树木年轮样品处理流程处理.干燥后用白乳胶固定在样本槽内, 用不同粗细程度的砂纸逐级打磨至光、滑、亮状态, 能在40倍显微镜下清晰观察到年轮轮廓为止.在双筒显微镜下进行目测交叉定年后, 用LINTAB 6.0年轮宽度测量仪(精度为0.001 mm)(Rinntech, Heidelberg, Germany)测量树轮原始宽度.在COFECHA程序中进行交叉定年, 精确确定每个年轮的生长年份.对去除树木生长的自相关性以及低频信号的结果进行检验, 剔除与主序列相关性差、年轮序列不完整以及难以交叉定年的样芯, 最终剩余62株样本的95根样芯进入主序列(
表1 ).用R语言dplR扩展包(
Bunn, 2010 )中的负指数函数或样条函数拟合去除因遗传因素或个体干扰产生的抑制、释放等生长趋势后, 最终得到云南松标准年表(STD)(
图2 )、差值年表(RES)和自回归年表(ARS).STD公共区间(1954-2018年)统计特征如
表1 所示, 样本总体代表性(EPS)为0.94, 大于阈值0.85, 能较好地代表研究区云南松树轮宽度变化的基本特征; 较高的平均敏感度(MS, 0.18)和信噪比(SNR, 14.31)表明云南松对环境变化敏感, 年表质量较好, 保存了较高的气候信号.因此, 选用标准年表进行相关性分析. ...
气候变暖下大兴安岭落叶松径向生长对温度的响应
1
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
气候变暖下大兴安岭落叶松径向生长对温度的响应
1
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China
1
2005
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
Tree-ring reconstructed winter precipitation and tropical teleconnections in Durango, Mexico
1
2003
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
Climate change and tree-ring relationships of
Nothofagus menziesii tree-line forests
1
2001
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
云南松林在云南的分布研究
1
2013
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
云南松林在云南的分布研究
1
2013
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
云南松林资源动态研究
1
2014
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
云南松林资源动态研究
1
2014
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
Tree-ring recorded drought variability in the northern Min Mountains of northwestern China
1
2016
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
The effect of climate on radial growth of
Quercus liaotungensis forest trees in Loess Plateau, China
1
2007
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern China
1
2009
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
Spatial and temporal temperature trends on the Yunnan Plateau (southwest China) during 1961-2004
1
2011
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
树轮生态学研究进展
1
2008
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
树轮生态学研究进展
1
2008
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
第七次全国森林资源清查及森林资源状况
1
2010
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
第七次全国森林资源清查及森林资源状况
1
2010
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
Growth-rings of trees: Their correlation with climate
2
1966
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
Tree-Rings and Climate 2
1976
... 按照国际树木年轮库(ITRDB)标准以及树木年轮学采样原则与方法(
Fritts, 1976 ), 在海拔1 950-2 200 m范围随机选取生长良好、树木间相互遮蔽较小、立地条件较差的云南松钻取树芯.共选取87株云南松样本, 样树胸径23.0-42.6 cm, 树高9.0-19.6 m.每株样本在胸高处(1.3 m)不同方位钻取2根年轮样芯(取样方向首选南向, 另一样芯方向与第一根垂直), 共采集172根样芯. ...
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
Tree-ring characteristics along a vegetation gradient in Northern Arizona
2
1965
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 气象资料选取距离采样点最近的曲靖沾益气象站器测气象资料.资料时段为1952-2016年(
图1 ), 其中月平均气温(
T mp )、月平均最高气温(
T mx )、月平均最低气温(
T mn )、月降水量(
Pre )、月平均相对湿度(
RH )来源于中国气象科学数据共享服务网(http://data.cma.cn/data/weatherBk.html).帕尔默干旱指数(
PDSI, 1901-2017)来源于荷兰皇家气象局服务网(https://climexp.knmi.nl/start.cgi), 分辨率为0.5° × 0.5°.鉴于树木生长对气候响应存在“滞后效应”, 即前一年气候状况会对当年树木生长产生影响(
Fritts et al ., 1965b ), 故选取上年5月至当年11月的气象数据, 共计108个气候变量与STD年表数据进行响应分析.采用
PDSI 描述研究区干湿状况,
PDSI 是结合降水、蒸散以及干旱持续时间的综合气象干旱指标, 被广泛用于干旱评估(
卫捷和马柱国, 2003 ).-0.5 <
PDSI ≤ 0.5表示正常、-1 <
PDSI ≤ -0.5表示初旱、-2 <
PDSI ≤ -1表示轻旱、-3 <
PDSI ≤ -2表示中旱、-4 <
PDSI ≤ -3表示重旱、
PDSI ≤ -4表示极旱,
PDSI 值越小, 气候越干旱. ...
The biological model for paleoclimatic interpretation of mesa verde tree-ring series
1
1965
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
Subalpine tree growth, climate, and increasing CO
2 : An assessment of recent growth trends
1
1991
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
川西高原不同坡向和海拔云冷杉树轮对气候变化的响应差异
1
2016
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
川西高原不同坡向和海拔云冷杉树轮对气候变化的响应差异
1
2016
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
南盘江流域旱涝时空分布规律研究
1
2014
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
南盘江流域旱涝时空分布规律研究
1
2014
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
Tree-ring-based SPEI reconstruction in central Tianshan Mountains of China since A.D. 1820 and links to westerly circulation
1
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
Rapid climate change- related growth decline at the southern range edge of Fagus sylvatica
1
2006
... 运用Dendroclim 2002软件中的响应函数分析年轮宽度指数与气候数据(温度、降水、
RH 、
PDSI )的响应关系.以年轮宽度指数为预测变量、与其显著相关(
p < 0.05)的气候因子作为解释变量, 利用R软件的lm函数进行多元回归分析, 用step函数通过后向逐步回归方法优化获得简化回归模型(
van der Maaten-Theunissen et al ., 2015 ).为保证模型参数的可比性, 分析前利用scale函数对数据进行标准化.用简化回归模型中各解释变量标准化回归系数绝对值占所有解释变量回归系数绝对值总和的百分率作为气候因子对年轮生长的解释率(
Le Bagousse-Pinguet et al ., 2019 ).基于测量的树木年轮宽度计算云南松断面积增长量(
BAI ),BAI=$\pi\times(R_n^2-R_n-1^2)$, 其中
R 为云南松样树半径,
n 为年份(
Jump et al ., 2006 ).利用R语言中bootRes程序包中的mdcc函数计算滑动响应相关系数, 并利用mdcplot函数作出滑动响应色温图(
Zang & Biondi, 2013 ), 分析随气候暖干化发展与云南松径向生长显著相关的气候因子与年轮宽度响应关系的稳定性.作图用Sigmplot 10.0软件完成. ...
Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems 2
2001
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
... ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
Climatic responses of tree-ring widths of
Larix gmelinii on contrasting north-facing and south-facing slopes in central Siberia
1
2007
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality
1
2019
... 运用Dendroclim 2002软件中的响应函数分析年轮宽度指数与气候数据(温度、降水、
RH 、
PDSI )的响应关系.以年轮宽度指数为预测变量、与其显著相关(
p < 0.05)的气候因子作为解释变量, 利用R软件的lm函数进行多元回归分析, 用step函数通过后向逐步回归方法优化获得简化回归模型(
van der Maaten-Theunissen et al ., 2015 ).为保证模型参数的可比性, 分析前利用scale函数对数据进行标准化.用简化回归模型中各解释变量标准化回归系数绝对值占所有解释变量回归系数绝对值总和的百分率作为气候因子对年轮生长的解释率(
Le Bagousse-Pinguet et al ., 2019 ).基于测量的树木年轮宽度计算云南松断面积增长量(
BAI ),BAI=$\pi\times(R_n^2-R_n-1^2)$, 其中
R 为云南松样树半径,
n 为年份(
Jump et al ., 2006 ).利用R语言中bootRes程序包中的mdcc函数计算滑动响应相关系数, 并利用mdcplot函数作出滑动响应色温图(
Zang & Biondi, 2013 ), 分析随气候暖干化发展与云南松径向生长显著相关的气候因子与年轮宽度响应关系的稳定性.作图用Sigmplot 10.0软件完成. ...
东北南部蒙古栎径向生长对气候变化的响应——以千山为例
1
2014
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
东北南部蒙古栎径向生长对气候变化的响应——以千山为例
1
2014
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
利用树木年轮宽度资料重建川西卧龙地区过去159年夏季温度的变化
2
2010
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 暖干化前后, 6、7月气候因子的影响比较稳定, 它们对径向生长的解释率都在45%左右, 由此可见,暖干化并没有改变研究区6和7月的气候特点, 云南松生长旺盛期的径向生长也相对稳定.在水分条件不受限制的情况下(夏季6、7、8月)增温作用不会对树木生长产生抑制作用, 反而能够缓解水分过多产生的胁迫作用, 提高光合作用效率(
李宗善等, 2010 ).这与研究区6、7月水热条件优越, 云南松生长迅速的实地观测结果(
图1 )一致. ...
利用树木年轮宽度资料重建川西卧龙地区过去159年夏季温度的变化
2
2010
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 暖干化前后, 6、7月气候因子的影响比较稳定, 它们对径向生长的解释率都在45%左右, 由此可见,暖干化并没有改变研究区6和7月的气候特点, 云南松生长旺盛期的径向生长也相对稳定.在水分条件不受限制的情况下(夏季6、7、8月)增温作用不会对树木生长产生抑制作用, 反而能够缓解水分过多产生的胁迫作用, 提高光合作用效率(
李宗善等, 2010 ).这与研究区6、7月水热条件优越, 云南松生长迅速的实地观测结果(
图1 )一致. ...
模拟气候变暖对东北兴安落叶松径向生长的影响
1
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
模拟气候变暖对东北兴安落叶松径向生长的影响
1
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
1951-2006年中国区域干旱化特征
1
2007
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
1951-2006年中国区域干旱化特征
1
2007
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
Millennium-long tree-ring chronology reveals megadroughts on the southeastern Tibetan Plateau
1
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
Growth response of
Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern China
3
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
Simulated responses of
Pinus halepensis forest productivity to climatic change and CO
2 increase using a statistical model
1
2000
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
华山树木年轮年表的建立
1
1994
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
华山树木年轮年表的建立
1
1994
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
利用树轮资料重建长白山区过去气候变化
2
1997
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
利用树轮资料重建长白山区过去气候变化
2
1997
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
The altitudinal variability and temporal instability of the climate-tree-ring growth relationships for Changbai larch (
Larix olgensis Henry) in the Changbai mountains area, Jilin, Northeastern China
1
2016
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
近50a中国西南地区地表干湿状况研究
1
2014
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
近50a中国西南地区地表干湿状况研究
1
2014
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
PointRes: An R package to analyze pointer years and components of resilience
1
2015
... 运用Dendroclim 2002软件中的响应函数分析年轮宽度指数与气候数据(温度、降水、
RH 、
PDSI )的响应关系.以年轮宽度指数为预测变量、与其显著相关(
p < 0.05)的气候因子作为解释变量, 利用R软件的lm函数进行多元回归分析, 用step函数通过后向逐步回归方法优化获得简化回归模型(
van der Maaten-Theunissen et al ., 2015 ).为保证模型参数的可比性, 分析前利用scale函数对数据进行标准化.用简化回归模型中各解释变量标准化回归系数绝对值占所有解释变量回归系数绝对值总和的百分率作为气候因子对年轮生长的解释率(
Le Bagousse-Pinguet et al ., 2019 ).基于测量的树木年轮宽度计算云南松断面积增长量(
BAI ),BAI=$\pi\times(R_n^2-R_n-1^2)$, 其中
R 为云南松样树半径,
n 为年份(
Jump et al ., 2006 ).利用R语言中bootRes程序包中的mdcc函数计算滑动响应相关系数, 并利用mdcplot函数作出滑动响应色温图(
Zang & Biondi, 2013 ), 分析随气候暖干化发展与云南松径向生长显著相关的气候因子与年轮宽度响应关系的稳定性.作图用Sigmplot 10.0软件完成. ...
树木年轮宽度与气候变化关系研究进展
1
2003
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
树木年轮宽度与气候变化关系研究进展
1
2003
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
Rapid warming forces contrasting growth trends of subalpine fir (
Abies fabri ) at higher- and lower-elevations in the eastern Tibetan Plateau
1
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
大兴安岭北部樟子松树木生长与气候因子的关系
2
2011
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
大兴安岭北部樟子松树木生长与气候因子的关系
2
2011
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
全球气候变化对中国森林生态系统的影响
1
2006
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
全球气候变化对中国森林生态系统的影响
1
2006
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
Palmer干旱指数、地表湿润指数与降水距平的比较
1
2003
... 气象资料选取距离采样点最近的曲靖沾益气象站器测气象资料.资料时段为1952-2016年(
图1 ), 其中月平均气温(
T mp )、月平均最高气温(
T mx )、月平均最低气温(
T mn )、月降水量(
Pre )、月平均相对湿度(
RH )来源于中国气象科学数据共享服务网(http://data.cma.cn/data/weatherBk.html).帕尔默干旱指数(
PDSI, 1901-2017)来源于荷兰皇家气象局服务网(https://climexp.knmi.nl/start.cgi), 分辨率为0.5° × 0.5°.鉴于树木生长对气候响应存在“滞后效应”, 即前一年气候状况会对当年树木生长产生影响(
Fritts et al ., 1965b ), 故选取上年5月至当年11月的气象数据, 共计108个气候变量与STD年表数据进行响应分析.采用
PDSI 描述研究区干湿状况,
PDSI 是结合降水、蒸散以及干旱持续时间的综合气象干旱指标, 被广泛用于干旱评估(
卫捷和马柱国, 2003 ).-0.5 <
PDSI ≤ 0.5表示正常、-1 <
PDSI ≤ -0.5表示初旱、-2 <
PDSI ≤ -1表示轻旱、-3 <
PDSI ≤ -2表示中旱、-4 <
PDSI ≤ -3表示重旱、
PDSI ≤ -4表示极旱,
PDSI 值越小, 气候越干旱. ...
Palmer干旱指数、地表湿润指数与降水距平的比较
1
2003
... 气象资料选取距离采样点最近的曲靖沾益气象站器测气象资料.资料时段为1952-2016年(
图1 ), 其中月平均气温(
T mp )、月平均最高气温(
T mx )、月平均最低气温(
T mn )、月降水量(
Pre )、月平均相对湿度(
RH )来源于中国气象科学数据共享服务网(http://data.cma.cn/data/weatherBk.html).帕尔默干旱指数(
PDSI, 1901-2017)来源于荷兰皇家气象局服务网(https://climexp.knmi.nl/start.cgi), 分辨率为0.5° × 0.5°.鉴于树木生长对气候响应存在“滞后效应”, 即前一年气候状况会对当年树木生长产生影响(
Fritts et al ., 1965b ), 故选取上年5月至当年11月的气象数据, 共计108个气候变量与STD年表数据进行响应分析.采用
PDSI 描述研究区干湿状况,
PDSI 是结合降水、蒸散以及干旱持续时间的综合气象干旱指标, 被广泛用于干旱评估(
卫捷和马柱国, 2003 ).-0.5 <
PDSI ≤ 0.5表示正常、-1 <
PDSI ≤ -0.5表示初旱、-2 <
PDSI ≤ -1表示轻旱、-3 <
PDSI ≤ -2表示中旱、-4 <
PDSI ≤ -3表示重旱、
PDSI ≤ -4表示极旱,
PDSI 值越小, 气候越干旱. ...
树木年轮分析在环境变化研究中的应用
1
1990
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
树木年轮分析在环境变化研究中的应用
1
1990
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
中国树木年轮气候学研究动态与展望
1
1993
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
中国树木年轮气候学研究动态与展望
1
1993
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data
2
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
南盘江流域季节性干旱时空分布特征研究
1
2013
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
南盘江流域季节性干旱时空分布特征研究
1
2013
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
滇西北玉龙雪山不同海拔云南松径向生长对气候因子的响应
1
2018
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
滇西北玉龙雪山不同海拔云南松径向生长对气候因子的响应
1
2018
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
中国西南干旱对气候变暖的响应特征
1
2014
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
中国西南干旱对气候变暖的响应特征
1
2014
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
气候变暖对香格里拉油麦吊云杉径向生长的影响
1
2019
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
气候变暖对香格里拉油麦吊云杉径向生长的影响
1
2019
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
我国西南干旱研究最新进展综述
1
2013
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
我国西南干旱研究最新进展综述
1
2013
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
Spatial variation and temporal instability in the climate-growth relationship of Korean pine in the Changbai Mountain region of Northeast China
2
2013
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
... ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
长白山北坡落叶松年轮年表及其与气候变化的关系
1
2005
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
长白山北坡落叶松年轮年表及其与气候变化的关系
1
2005
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
小兴安岭红松和鱼鳞云杉径向生长对气候变化的响应
2
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
小兴安岭红松和鱼鳞云杉径向生长对气候变化的响应
2
2017
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
普达措国家公园海拔上限3个针叶树种径向生长对气候变化的响应
2
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
普达措国家公园海拔上限3个针叶树种径向生长对气候变化的响应
2
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
天山乌鲁木齐河源450 a冬季温度序列的重建与分析
1
1999
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
天山乌鲁木齐河源450 a冬季温度序列的重建与分析
1
1999
... 研究区5、7和12月的水热条件是云南松径向生长的关键因素.7月是云南松生长的旺盛时期, 研究区7月降水丰富, 但连续阴雨会降低温度和光照, 削弱光合作用.此时, 高温就可促进云南松的生长.研究发现, 当降水不再是树木生长的限制因子时, 降水就与树木生长不相关或负相关(
Fritts, 1966 ,
1976 ).这说明降水对树木生长的影响取决于降水的分配以及它是否是树木生长的限制因子.夏季末期以及秋季的平均最低气温与云南松径向生长明显相关, 可能原因是温度升高延长了生长季, 为来年早材生长储备更多养分(
Du et al ., 2007 ;
袁玉江和李江风, 1999 ).12月升温会消耗前期储存的养分, 减少生长季节可利用的营养物质(
Graumlich,1991 ;
王晓春等, 2011 ).此时, 研究区正处于旱季, 如有降水补充, 则可缓解干旱的危害.本研究中10月的气候因子与云南松径向生长相关性不显著, 并且温度的促进作用推迟到11月份.这与
杨绕琼等(2018) 对滇西北玉龙雪山云南松的研究结论不一致.原因可能是, 研究区的纬度较低, 10月的降水和气温均高于滇西北区域, 云南松生长季较长.这些差异也表明, 开展林木对气候响应的研究, 应注意研究区内的气候差异, 才能全面揭示林木对气候的响应模式.这种差异也为通过以气候空间变化替代气候时间变化进行林木对不同气候变化情景响应的研究提供了可能的方法途径. ...
Dendroclimatic calibration in R: The bootRes package for response and correlation function analysis
1
2013
... 运用Dendroclim 2002软件中的响应函数分析年轮宽度指数与气候数据(温度、降水、
RH 、
PDSI )的响应关系.以年轮宽度指数为预测变量、与其显著相关(
p < 0.05)的气候因子作为解释变量, 利用R软件的lm函数进行多元回归分析, 用step函数通过后向逐步回归方法优化获得简化回归模型(
van der Maaten-Theunissen et al ., 2015 ).为保证模型参数的可比性, 分析前利用scale函数对数据进行标准化.用简化回归模型中各解释变量标准化回归系数绝对值占所有解释变量回归系数绝对值总和的百分率作为气候因子对年轮生长的解释率(
Le Bagousse-Pinguet et al ., 2019 ).基于测量的树木年轮宽度计算云南松断面积增长量(
BAI ),BAI=$\pi\times(R_n^2-R_n-1^2)$, 其中
R 为云南松样树半径,
n 为年份(
Jump et al ., 2006 ).利用R语言中bootRes程序包中的mdcc函数计算滑动响应相关系数, 并利用mdcplot函数作出滑动响应色温图(
Zang & Biondi, 2013 ), 分析随气候暖干化发展与云南松径向生长显著相关的气候因子与年轮宽度响应关系的稳定性.作图用Sigmplot 10.0软件完成. ...
玉龙雪山3个针叶树种在海拔上限的径向生长及气候响应
1
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
玉龙雪山3个针叶树种在海拔上限的径向生长及气候响应
1
2017
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系
1
2010
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系
1
2010
... 本研究发现, 云南松径向生长与上年5、6、7、12月, 当年1、3、5月平均气温响应关系的不稳定性集中于1971-2004年间, 与上年6、7、9、10、12月, 当年8月降水的响应关系不稳定时间段均集中于1968-1999年间, 对各月水热条件(
PDSI )响应的不稳定性主要集中在1968-2000年间.说明云南松径向生长与气候因子的响应关系受气候变化的影响, 响应关系的稳定性会随气候变化发生改变.
于健等(2017) 对小兴安岭鱼鳞云杉(
Picea koraiensis )与气候关系稳定性的研究发现, 鱼鳞云杉与气候的响应关系在20世纪后期随着气候变暖导致的暖干化加剧变得不稳定, 且不稳定性呈增强趋势.
Yu等(2013) 关于长白红松对气候响应的研究也表明, 气候因子对红松生长影响的时间不稳定性在1970年以后随着气候明显变暖而变得强烈.
Shen等(2016) 对长白落叶松(
Korean Larch )与气候响应关系的研究也证明, 长白落叶松生长与温度的响应关系在1980年以后随气温升高变得极不稳定.
尹定财等(2019) 对香格里拉麦吊云杉对气候变暖响应的研究发现树木径向生长与主要气候因子关系的稳定性与不同时段生长-气候响应关系变化具有一致性, 进一步证明了气候变化会影响树木生长与气候因子关系的稳定性.在滇西北地区对长苞冷杉的研究中也得出了类似的结论(
Panthi et al ., 2018 ), 即气候突变会导致树木生长与气候因子的关系发生改变.本研究中, 云南松径向生长与温度、水分以及水热组合条件的响应关系均在1968-2004年间发生明显波动, 气候响应关系的不稳定性明显增强, 而这段时期正是研究区域增温最为明显, 暖干化程度加剧的阶段.树木生长与气候因子的动态关系一直是树木年轮学研究的热点.针对我国东北地区树种的研究资料表明, 树木生长对气候变化响应稳定性关系可为研究区气候重建提供参考(
邵雪梅和吴祥定, 1997 ;
于大炮等, 2005 ;
张先亮等, 2010 ;
李腾等, 2014 ), 为未来气候生长模型参数设置方面的改进提供有用参考信息(
Yu et al ., 2013 ).本研究也可在这两方面为云南松林区气候的重建及建立气候生长模型提供参考. ...
滇西北石卡雪山2个针叶树种森林上限径向生长对温度和降水的响应
2
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
滇西北石卡雪山2个针叶树种森林上限径向生长对温度和降水的响应
2
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
玉龙雪山东坡不同海拔长苞冷杉径向生长与气候因子的关系
2
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
玉龙雪山东坡不同海拔长苞冷杉径向生长与气候因子的关系
2
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
普达措国家公园2个针叶树种径向生长对温度和降水的响应
1
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
普达措国家公园2个针叶树种径向生长对温度和降水的响应
1
2018
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
2009-2011年我国西南地区旱灾程度及其对植被净初级生产力的影响
1
2015
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
2009-2011年我国西南地区旱灾程度及其对植被净初级生产力的影响
1
2015
... 我国西南地区气候总体较为湿润, 且存在“东湿西干”的区域差异(
姚玉璧等, 2014 ).当前, 西南地区正处于一个干旱化过程(
马柱国和任小波, 2007 ), 2009-2010年的“西南大旱”就是其间的极端干旱事件(
尹晗和李耀辉, 2013 ;
赵志平等, 2015 ).近50年来, 西南地区的气候呈“暖干化”发展, 在21世纪后呈现出进一步加剧的趋势(
苏秀程等, 2014 ).近年来, ****围绕全球气候变化对滇西北和川西高原高山、亚高山典型针叶树种树轮气候响应特征开展了大量研究.
Fan等(2009) 和
Panthi等(2018) 研究了横断山区不同海拔长苞冷杉(
Abies georgei )及高海拔主要针叶树种径向生长对气候变化的响应, 利用1655年以来滇西北云杉(
Picea asperata )、铁杉(
Tsuga chinensis )、冷杉(
Abies fabri )的树轮资料揭示了横断山区干旱的时空变化特征.
余佳霖等(2017) , 张赟等(
2018a ,
2018b )研究了滇西北石卡雪山、玉龙雪山不同海拔及林线地段优势针叶树种长苞冷杉、高山松(
Pinus densata )、大果红杉(
Larix potaninii )、麦吊云杉(
Picea brachytyla )对气候因子的响应.
郭滨德(2016) 研究了川西高原不同坡向和海拔云冷杉径向生长对气候变化的响应.
李宗善等(2010) 利用岷江冷杉的树轮资料完成了川西卧龙地区气候重建工作.目前, 针对气候暖干化对树木生长影响的研究主要集中于东北地区(
王晓春等, 2011 ;
常永兴等, 2017 ;
刘玉佳等, 2017 ;
于健等, 2017 ), 关于西南地区气候暖干化对树木生长影响的研究尚未见相关报道. ...
Spring phenophases in recent decades over Eastern China and its possible link to climate changes
1
2006
... 暖干化后, 当年气候因子对云南松生长的影响增强, 上年气候因子的影响有所下降.主要原因可能是: (1)暖干化导致的12月增温避免了低温对云南松的危害(
Komer & Christian, 2001 ), 原来与年轮宽度显著相关的上年12月平均气温和平均最低气温转变为不显著相关因子(
图6 ), 避免了对云南松来年径向生长的影响, 降低了上年气候因子的影响作用(
Komer & Christian, 2001 ).(2)暖干化使云南松生长季后期(9-11月)仍具有较高的温度, 延长了生长期(
Chen et al ., 2005 ;
Zheng et al ., 2006 ;
Yang et al ., 2017 ), 促进了当年的径向生长(
Cleaveland et al ., 2003 ), 使径向生长解释率增加了7.20%.(3)暖干化后, 温度不再是云南松生长的限制因子, 水分的限制作用越来越突出, 原来与年轮宽度相关性不显著的当年7、10、11月
PDSI 转变为显著相关因子(
图6 ), 并在当年体现.云南松生长旺盛期(6、7月)的气候因子对径向生长的影响在暖干化前后都较稳定, 暖干化后与径向生长显著相关的当年气候因子增加了3个, 对径向生长的解释率增加了16.05%.可见, 暖干化减弱了气候因子对云南松生长影响的“滞后效应” (
Fritts et al ., 1965a ;
吴祥定, 1990 ), 增强了当年气候因子影响的时效性. ...
南盘江流域主汛期降雨量年(代)际变率研究及趋势预测
1
2015
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
南盘江流域主汛期降雨量年(代)际变率研究及趋势预测
1
2015
... 云南松(
Pinus yunnanensis )林是西南地区重要的森林类型和典型的偏干性暖性针叶林(
邓喜庆等, 2013 ,
2014 ).在我国优势乔木树种中, 云南松的面积和蓄积量均列前10位, 分布面积460.59万hm
2 、蓄积量46 872.15万m³, 分别占全国森林资源面积和蓄积量的2.96%和3.51% (
国家林业局森林资源管理司, 2010 ).云南松是西南的乡土树种, 主要分布在气候暖干化明显的西南地区西部.本研究以生长在暖干化特征明显的南盘江流域(
杨茂灵等, 2013 ;
贾春强等, 2014 ;
朱保林等, 2015 )的云南松为研究对象, 通过树木年轮学方法建立径向生长与气候因子的关系, 开展径向生长对气候暖干化响应的研究, 旨在回答: (1)限制云南松径向生长的主要气候因子是什么? (2)在西南气候暖干化背景下, 影响云南松径向生长的主要气候因子及其影响强度是否有变化? (3)云南松径向生长对西南气候暖干化响应关系是否稳定? 本研究可为气候暖干化区域森林生产力和生态系统功能的评估提供基础数据, 为制定应对气候变化的森林管理对策提供参考依据. ...
气候变化与森林生态系统:影响、脆弱性与适应性
1
2007
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
气候变化与森林生态系统:影响、脆弱性与适应性
1
2007
... 全球气候变化对森林生态系统的结构、功能产生了严重影响(
王叶和延晓冬, 2006 ;
朱建华等, 2007 ), 使林木对气候变化响应的研究成为当前重要的科学问题.树木生长受环境因子及遗传因素的影响(
吴祥定和邵雪梅, 1993 ;
邵雪梅和吴祥定, 1994 ,
1997 ), 其中气候的影响尤为显著(
Fritts et al ., 1965b ;
Fritts, 1966 ).树木径向生长率是多种生态因子综合作用的结果, 树木年轮记录了树木的年龄、生长特征以及生长过程中的气候和环境变化信息.在去除树木自身生长趋势及非气候因子的影响后, 通过年轮可获取树木生长过程的气候信息, 揭示气候的演变过程以及影响树木生长的关键气候因子(
王婷等, 2003 ;
方克艳等, 2008 ).基于树轮记录的连续性和准确性, 树木年轮已经成为研究树木生长对气候变化响应的主要方法(
Rathgeber et al ., 2000 ;
Cullen et al ., 2001 ;
Kujansuu et al ., 2007 ;
Nie et al ., 2017 ).近十几年来, 树木年轮学在我国迅速发展, 已对高寒山区(
Jiang et al ., 2017 ;
Yang et al ., 2017 ;
Panthi et al ., 2018 ), 干旱区(
Fan et al ., 2011 ;
Deng et al ., 2016 )以及受季风气候影响下的主要优势树种做了大量的研究(
Wang et al ., 2017 ;
余佳霖等, 2017 ;
张卫国等, 2017 ;
张贇等, 2018a ,
2018b ,
2018c ), 为全球气候变化与树木的互作研究奠定了坚实基础. ...
 ,1,*
,1,*
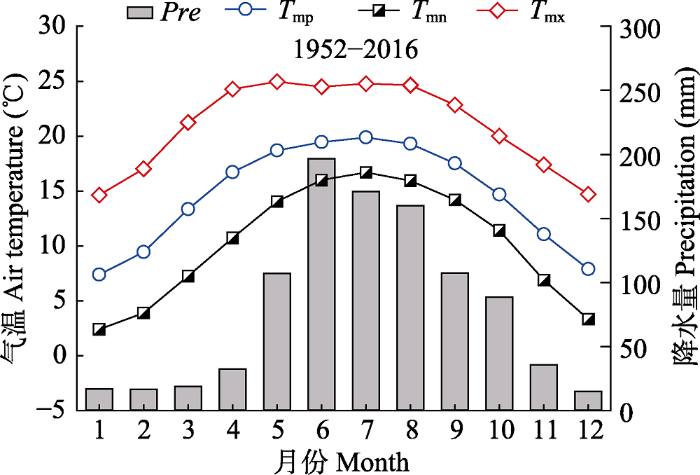 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT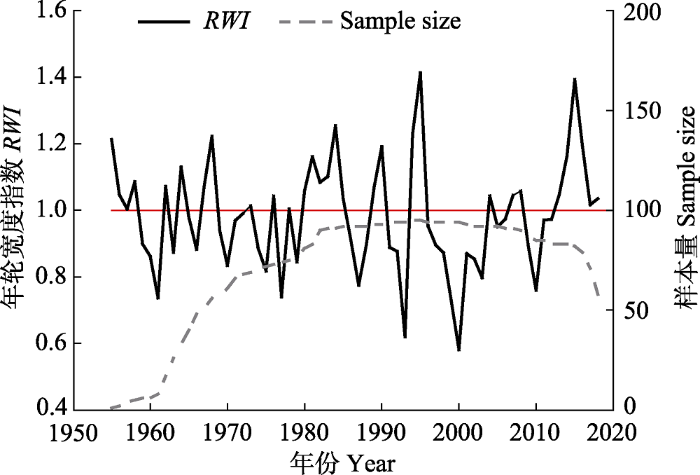 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT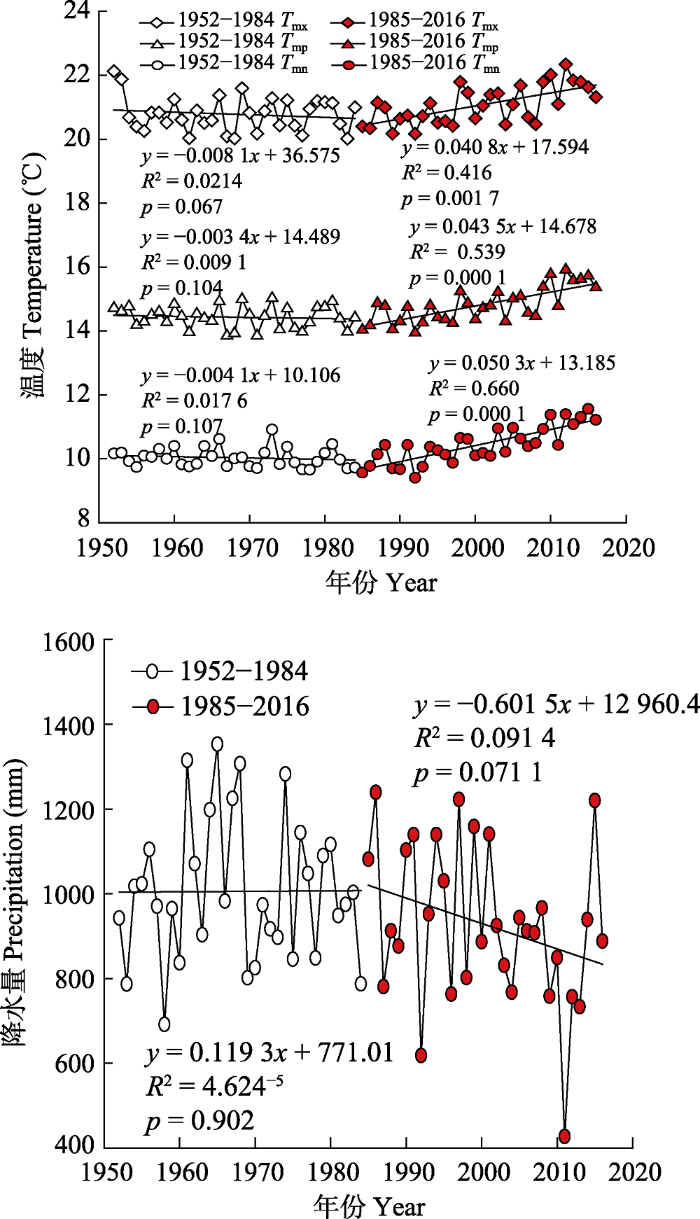 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT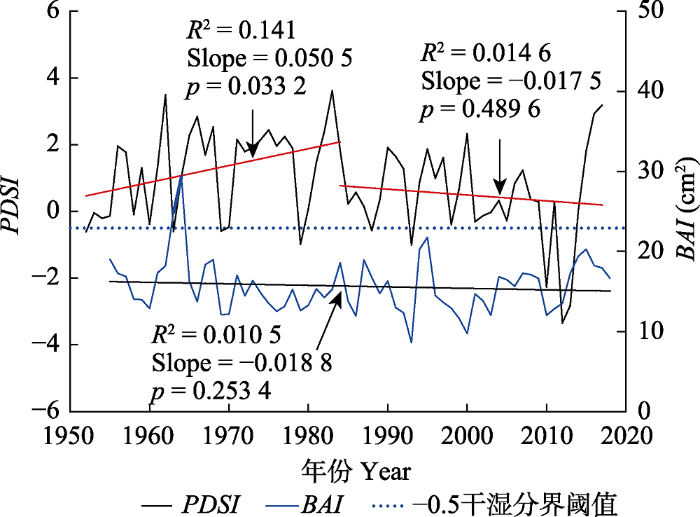 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT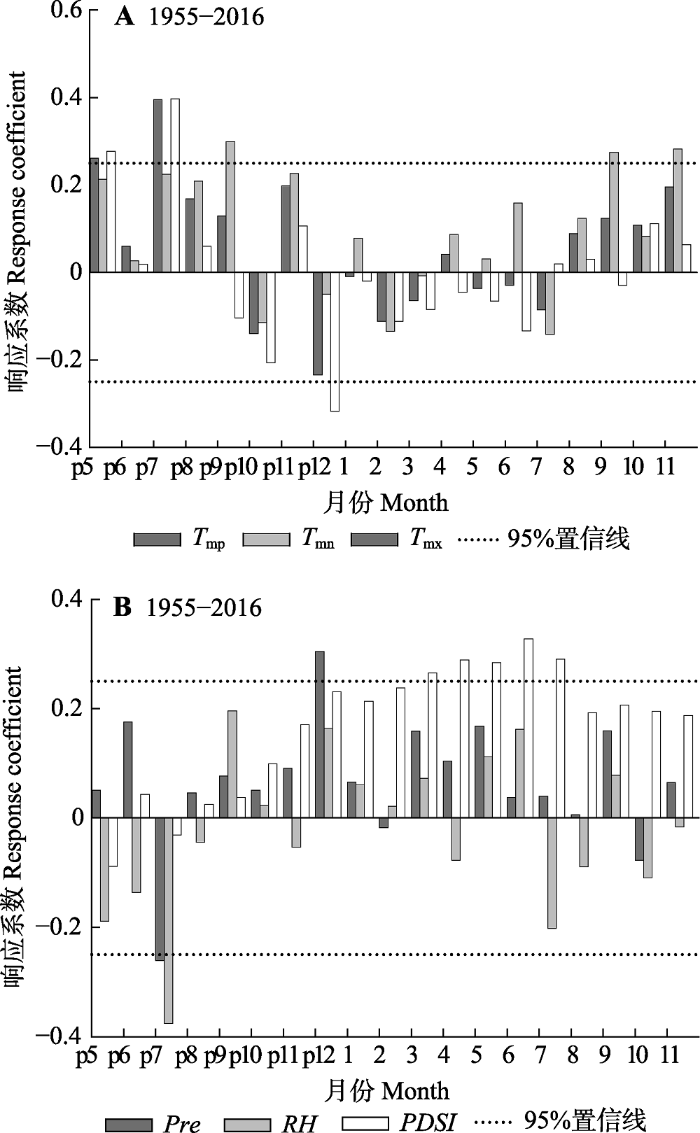 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT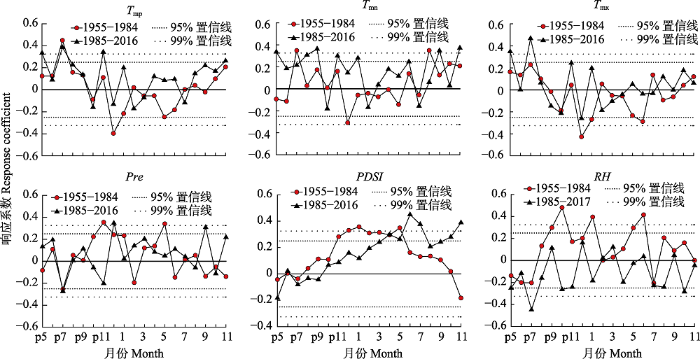 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT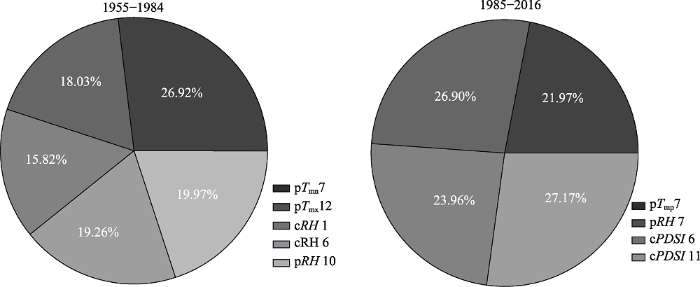 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT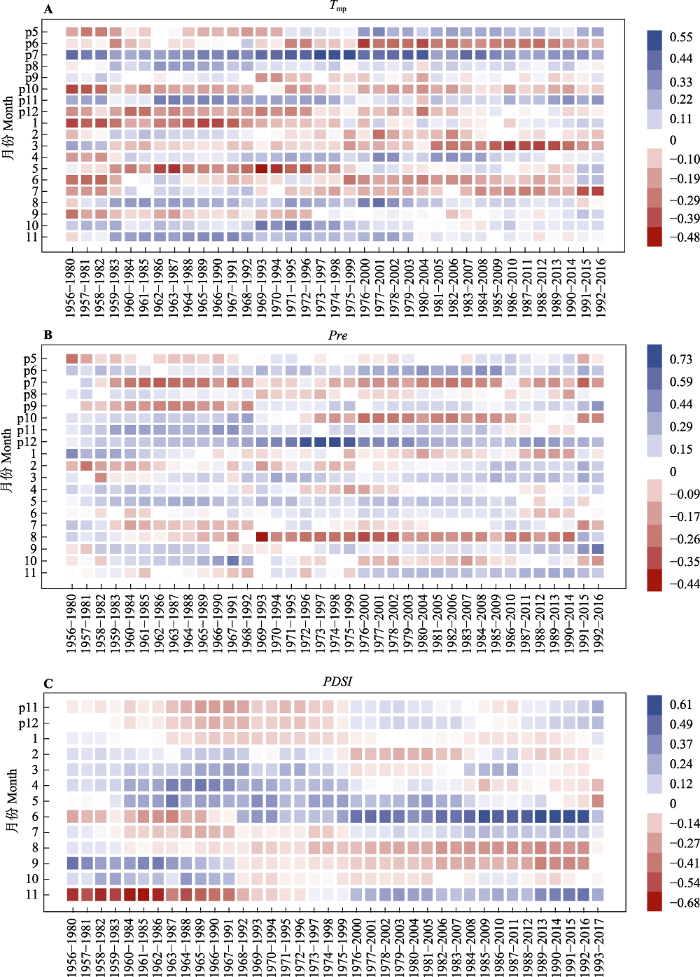 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}