张雪皎
1 , 高贤明
2 , 吉成均
1 , 康慕谊
3 , 4 , 王仁卿
5 , 岳明
6 , 张峰
7 , 唐志尧
, , 1 , * 1 北京大学城市与环境学院, 北京大学生态研究中心, 地表过程分析与模拟教育部重点实验室, 北京 100871 2 中国科学院植物研究所植被与环境变化国家重点实验室, 北京 100093 3 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 4 北京师范大学地理科学学部自然资源学院, 北京 100875 5 山东大学生命科学学院, 济南 250100 6 西北大学西部资源生物与现代生物技术教育部重点实验室, 西安 710069 7 山西大学黄土高原研究所, 太原 030006 Response of abundance distribution of five species of Quercus to climate change in northern China Xue-Jiao ZHANG
1 , Xian-Ming GAO
2 , Cheng-Jun JI
1 , Mu-Yi KANG
3 , 4 , Ren-Qing WANG
5 , Ming YUE
6 , Feng ZHANG
7 , Zhi-Yao TANG
, , 1 , * 1 Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 2 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 3 State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China 4 College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 5 School of Life Sciences, Shandong University, Jinan 250100, China 6 Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, Northwest University, Xi’an 710069, China 7 Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 通讯作者: * zytang@urban.pku.edu.cn 编委: 王襄平
责任编辑: 李敏, 赵航(实习)
收稿日期: 2018-10-11
接受日期: 2019-01-30
网络出版日期: 2019-09-20
基金资助: 国家科技基础性工作专项 (2011FY110300 )国家科技基础性工作专项 (2015FY210200 )
Corresponding authors: * zytang@urban.pku.edu.cn Received: 2018-10-11
Accepted: 2019-01-30
Online: 2019-09-20
Fund supported: Supported by the National Basic Work of Science and Technology of China (2011FY110300 )Supported by the National Basic Work of Science and Technology of China (2015FY210200 )
摘要 植物分布与气候之间的关系是预估未来气候变化对生态系统影响的实现基础。以往的物种分布模型通常以物种的分布区或者分布点的物种存在数据作为物种分布的响应变量。相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响。该研究通过野外调查获取了华北及周边地区1 045个样方的栎属树木多度, 利用广义线性模型、广义加性模型和随机森林模型模拟栓皮栎(
Quercus variabilis )、麻栎(
Q. acutissima )、槲栎(
Q. aliena )、锐齿槲栎(
Q. aliena var.
acuteserrata )和蒙古栎(
Q. mongolica ) 5个树种多度的地理分布及未来2个不同时期(2050年和2070年)的潜在分布。结果表明: 随机森林模型对5个栎属树种的多度的拟合结果要优于广义线性模型和广义加性模型; 典型浓度路径(RCP) 8.5下的5个栎属树种在未来两个时期的多度变化幅度都要大于RCP 2.6下的变化, 在超过一半面积的区域中麻栎、槲栎、锐齿槲栎和蒙古栎的多度减少, 其中内蒙古东北部和黑龙江北部地区是5种栎属植物多度减少的集中分布地区。未来气候变化背景下, 需要加强对这几个区域的监测与物种保护。
关键词: 广义线性模型 ;
广义加性模型 ;
随机森林模型 ;
物种分布模型 Abstract Aims To develop a statistically appropriate species distribution model for the abundance of five species from
Quercus in the northern China, and to predict the change of abundance under climate change.
Methods We surveyed abundance data of five
Quercus species from 1 045 plots in the northern China, and then fit the abundance with climatic variables using random forest model (RF). We then predict the abundance of these five
Quercus species in 2050 and 2070 under Representation Concentration Pathways (RCP) 2.6 and 8.5.
Important findings The change magnitudes of abundance for all 5 species under RCP 8.5 were larger than under RCP 2.6. Except for
Quercus variabilis, abundances of other four species declined under climate change to 2050 and 2070 in more than half of the current distribution areas. Moreover, the northeastern part of Nei Mongol and the northern part of Heilongjiang will be the hotspots of decrease of abundance. Therefore, it is necessary to strengthen the monitoring and species protection in the areas mentioned above with the increasing threaten of climate change.
Keywords: generalized linear model ;
generalized additive model ;
random forest model ;
species distribution model PDF (12114KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 张雪皎, 高贤明, 吉成均, 康慕谊, 王仁卿, 岳明, 张峰, 唐志尧. 中国北方5种栎属树木多度分布及其对未来气候变化的响应. 植物生态学报, 2019, 43(9): 774-782. DOI:
10.17521/cjpe.2018.0249 ZHANG Xue-Jiao, GAO Xian-Ming, JI Cheng-Jun, KANG Mu-Yi, WANG Ren-Qing, YUE Ming, ZHANG Feng, TANG Zhi-Yao.
Response of abundance distribution of five species of Quercus to climate change in northern China .
Chinese Journal of Plant Ecology , 2019, 43(9): 774-782. DOI:
10.17521/cjpe.2018.0249 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 )。物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 )。传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 )。然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 )。相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 )。
近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 )。栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 )。栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种。在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复。本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布。
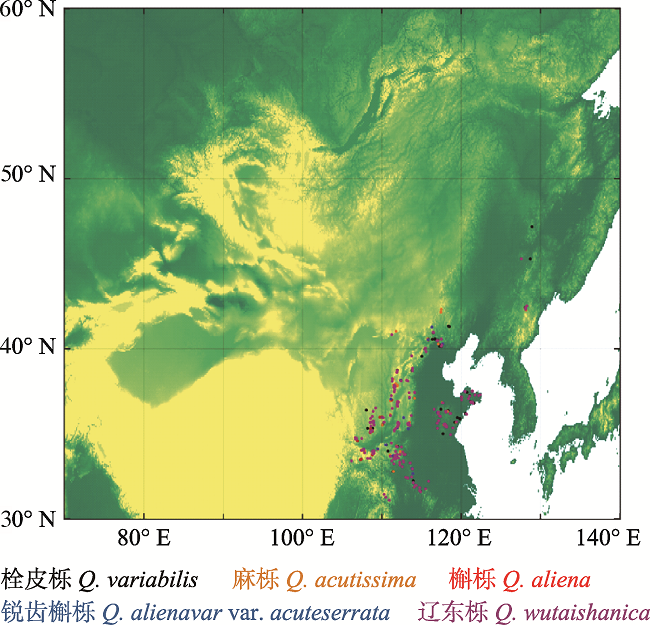
1 材料和方法 1.1 数据收集 研究以野外样方调查所获取的栎属植物多度作为响应变量。在华北及周边地区共调查1 045个栎林的样方(
图1 ), 每个样方面积为600-1 000 m
2 。在每个样方, 调查乔木层所有胸径≥3 cm的树木, 记录其胸径、树高, 共记录到栎属树木5种: 栓皮栎(
Q. variabilis )、槲栎(
Q. aliena )、锐齿槲栎(
Q. aliena var.
acuteserrata )、蒙古栎(
Q. mongolica )、麻栎(
Q. acutissima )。为统一单位和方便计算, 将样方中的各种栎属植物的密度统一折算为1 000 m
2 中的个体数量。这些样方的乔木层平均物种数为 7种·1 000 m
-2 , 平均树木密度为95棵·1 000 m
-2 (
表1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1栎属5种植物在中国北部的样方分布图。 Fig. 1Plots distribution of five species of Quercus in northern China. Table 1 表1 表1 中国北方5种常见栎属树木样方调查统计
Table 1
Survey statistics of plots of five species in Quercus in the northern China 物种 样方数 平均密度-2 ) 样方的气温范围 样方的降水范围 栓皮栎 Q. variabilis 304 89 ± 101 6.8-15.9 529-1 431 麻栎 Q. acutissima 195 39 ± 55 4.1-15.8 451-1 614 槲栎 Q. aliena 188 22 ± 43 3.6-16.0 335-1 775 锐齿槲栎 Q. aliena 147 37 ± 34 1.4-11.6 337-1 502 蒙古栎 Q. mongolica 492 55 ± 68 0.1-14.4 335-1 006
新窗口打开 |
下载CSV 研究使用的气候数据从世界气候数据库(http://www.worldclim.org/,
Hijmans et al ., 2005 )提取, 该数据库的Version 1.4收集了1960-2000年世界各地气象站记录的气象信息, 插值形成分辨率为30ʺ × 30ʺ (约1 km × 1 km)的全球气候栅格数据, 并基于每月气温和每月降水记录, 计算得到19个生物气候因子, 分别为年平均气温、气温日较差、等温性、气温季节性、最热月最高气温、最冷月最低气温、气温年较差、最湿季平均气温、最干季平均气温、最热季平均气温、最冷季平均气温、年降水量、最湿月降水量、最干月降水量、降水季节性、最湿季降水量、最干季降水量、最热季降水量和最冷季降水量。
未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5。本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果。RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 )。以上气候数据按照研究区裁剪。在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, 。
1.2 模型选择 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系。这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 )。
GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 )。RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 )。
GLM和GAM中, 增加自变量数目可以提高模型解释因变量的能力, 但具有多重共线性的多个自变量应用会导致模型过度拟合(
Sainani, 2013 )。为了避免过度拟合风险, 本研究通过计算19个气候因子的Pearson相关系数, 用单个因子对物种分别进行一次预模拟。基于模型的赤池信息量准则(AIC)值和各因子之间的相关性, 选取拟合度高且共线性小的因子(|
r | < 0.7)(
Yan et al ., 2017 ), 同时检测所选因子二次项是否显著。最终GLM包含所选因子的一次项和检验显著的二次项, GAM包含与线性模型相同的一次项因子。对于最终建立的GLM和GAM, 栓皮栎的预测模型包含了等温性、最热月最高气温、最冷月最低气温、降水季节性和最湿季降水量5个因子; 麻栎的预测模型包含了等温性、最冷月最低气温、气温年较差、最湿季平均气温、最干月降水量和最热季降水量6个变量; 槲栎的预测模型包含了等温性、最湿季平均气温、最干季平均气温、年降水量和最湿月降水量5个变量; 锐齿槲栎的预测模型包含等温性、最冷月最低气温、最湿季平均气温、最干月降水量和降水季节性等5个变量; 蒙古栎的预测模型包含了年平均气温、等温性、气温季节性、最湿季平均气温、年降水量、最湿月降水量和最热季降水量等7个变量。由于RF中不存在共线性问题, 使用19个气候因子作为自变量, 二叉树变量和森林数目选择使模型残差最小的值。
模型准确度用模型预测值与实际观测值之间的Pearson相关系数来表征(
Young & Carr, 2015 )。为验证模型的模拟质量, 用
k 折交叉验证来校验模型, 将数据集等分为
k 份, 每个子集均做一次测试集, 其余的作为训练集, 交叉验证重复
k 次。每次选择一个子集作为测试集, 并将
k 次的交叉验证平均值作为准确率, 这样, 所有的样本都可以被作为训练集和测试集, 每个样本都被验证一次, 可以充分利用已有数据集对算法效果进行测试, 本研究采用10折交叉验证, 最终准确度为100次10折交叉验证结果(
Picard & Denniscook, 1984 )。在5种栎属树木中, GLM对栓皮栎的拟合结果最佳(
r = 0.42); RF对栓皮栎、锐齿槲栎、麻栎的拟合结果较好(
r 值分别为0.61、0.63、0.53), 对槲栎的拟合结果较差(
r = 0.36)。总体而言, RF对5种树木的拟合结果都要优于GAM和GLM (
图2 ), 因此在后续结果中只采用随机森林进行基准气候与未来气候情景下物种多度的预测。预测结果用100次10折交叉验证建立起来的模型预测的平均值来表示。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2现有气候条件下随机森林模型(RF)、广义加性模型(GAM)和广义线性模型(GLM)对预测的栎属5种植物的潜在分布多度与实际观测值的关系及其准确度。A, 栓皮栎(Qva)。B, 麻栎(Qac)。C, 槲栎(Qal)。D, 锐齿槲栎(Qaa)。E, 蒙古栎(Qm)。F, 三个模型的准确度比较, 不同小写字母表示模型间差异显著(p < 0.05)。 Fig. 2Relationship between observed and predicted species abundances for five species in Quercus with random forest model (RF), generalized additive model (GAM), generalized linear model (GLM) and comparison of the accuracy of five species based on general linear model and random forest. A, Q. variabilis (Qva). B, Q. acutissima (Qac). C, Q. aliena (Qal). D, Q. aliena var. acuteserrata (Qaa). E, Q. mongolica (Qm). F, Model fitness of the three models, differed lowercase letters indicated significant differences between the models (p < 0.05). 2 结果 2.1 5种栎属树木的潜在多度分布 现代气候条件下, 随机森林对不同物种多度的模拟结果表明, 栓皮栎在陕西省中部、山西省北部、河北省东北部、北京市北部和黑龙江省北部多度较大; 麻栎多度较大的地区大致处于陕西省中部、山西省北部地区; 槲栎在陕西省中部、山西省北部分布数量较多; 锐齿槲栎在黑龙江省东部、吉林省西南部、辽宁省东部分布数量较多; 蒙古栎在内蒙古自治区东北部、黑龙江省北部及两省区交界处及陕西省北部分布较多(
图3 )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3中国北方5种树种在基准气候(1960-1990年)的多度(棵·1000 m-2 )潜在分布。A, 栓皮栎。B, 麻栎。C, 槲栎。D, 锐齿槲栎。E, 蒙古栎。 Fig. 3Species abundance (number·1 000 m-2 ) distribution maps of the North China for five species produced by random forest model, based on bioclimatic variables at year 1960-1990. A, Quercus variabilis . B, Q. acutissima. C, Q. aliena. D, Q. aliena var. acuteserrata. E, Q. mongolica. 2.2 未来气候变化背景下5种栎属树木的多度变化 与现代气候条件下的多度分布相比, 2050年RCP 2.6下, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度变化分别为平均1.17%、-6.55%、-4.84%、-8.78%和-0.77%, 除栓皮栎外, 其他4个物种的平均多度都减小。其中, 栓皮栎、槲栎和锐齿槲栎的变化不显著, 而麻栎和蒙古栎的变化显著。对栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎来说, 多度减少区域的面积分别占到总分布区域的47.1%、65.6%、59.8%、66.3%和55.2%, 也就是说, 除了栓皮栎外, 其他4个种在该气候情景下有超过一半区域的多度是减少的。RCP 8.5下, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度变化分别为平均2.9%、-2.81%、-2.19%、-16.23%和6.69%, 栓皮栎和蒙古栎的多度增加, 麻栎、槲栎和锐齿槲栎的多度减少, 其中, 麻栎多度减少显著, 其他4个种的变化都不显著。对于栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎来说, 多度减少区域的面积分别占到总分布区域的45.7%、60.4%、59.4%、77.3%和43.9%, 麻栎、槲栎和锐齿槲栎有超过一半区域的多度是减少的, 锐齿槲栎多度减少面积的比例达到70%以上(
图4 )。2050年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度在RCP 2.6和RCP 8.5下多度都集中减少的区域包括陕西省中部、内蒙古自治区东北部和黑龙江省北部区域。此外, 锐齿槲栎的多度在这两种气候情景下都在吉林省东南部、辽宁省东部和陕西省南部减少。栓皮栎、槲栎和蒙古栎在RCP 2.6和RCP 8.5下多度都增多的地区有黑龙江省中部和吉林省东部, 此外, 麻栎的多度在吉林省东部增加。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4中国北方5种树种在2050年时期RCP 2.6 (A-E)和RCP 8.5 (F-J)下预测多度值与现在基准(1960-1990年)的差值变化率分布。A, F, 栓皮栎。B, G, 麻栎。C, H, 槲栎。D, I, 锐齿槲栎。E, J, 蒙古栎。 Fig. 4Future distribution maps of rate of abundance change in the North China for five species produced by random forest model, based on bioclimatic variables under RCP 2.6 dispersal scenario (A-E) and RCP 8.5 dispersal scenario (F-J) in year 2050. A, F, Quercus variabilis . B, G, Q. acutissima. C, H, Q. aliena. D, I, Q. aliena var. acuteserrata. E, J, Q. mongolica. 与现代气候条件下的多度分布相比, 2070年RCP 2.6下, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度变化分别为平均1.46%、0.07%、-0.48%、-8.45%和2.05%, 栓皮栎、麻栎和蒙古栎的多度是增加的, 槲栎和锐齿槲栎的多度是减少的, 5个物种的增加(减少)都不显著。对于这5个物种, 多度减少区域的面积分别占到总分布区域的48.6%、54.4%、56.4%、64.4%和52.4%, 麻栎、槲栎、锐齿槲栎和蒙古栎都有超过一半区域的多度是减少的。RCP 8.5下, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度变化分别为平均-5.45%、-7.99%、-4.2%、-12.87%和-2.05%, 5个物种的多度都是减少的, 其中锐齿槲栎多度的减少不显著, 而其他4个种多度的减少都是显著的。对于这5个物种, 多度减少区域的面积分别占到总分布区域的61.6%、62.0%、61.3%、74.4%和52.4%, 也就是说5个物种都有一半以上区域的多度是减少的, 锐齿槲栎多度减少面积的比例达到70%以上(
图5 )。2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度在RCP 2.6和RCP 8.5下多度都集中减少的区域有黑龙江省东部、内蒙古自治区东北部, 多度都减少的面积相对于2050年下降0.67%。栓皮栎、麻栎、槲栎和蒙古栎的多度都在吉林省东南部、辽宁省东部和湖北省西部增加, 而锐齿槲栎的多度在湖北省西部增加。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5中国北方5种树种在2070年时期RCP 2.6 (A-E)和RCP 8.5 (F-J)下预测多度值与现在基准(1960-1990年)的差值变化率分布。A, F, 栓皮栎。B, G, 麻栎。C, H, 槲栎。D, I, 锐齿槲栎。E, J, 蒙古栎。 Fig. 5Future distribution maps of rate of abundance change in the North China for five species produced by Random Forest models, based on bioclimatic variables under RCP 2.6 dispersal scenario (A-E) and RCP 8.5 dispersal scenario (F-J) in year 2070. A, F, Quercus variabilis . B, G, Q. acutissima. C, H, Q. aliena. D, I, Q. aliena var. acuteserrata. E, J, Q. mongolica. 2.3 5种栎属树木多度变化的海拔分异 栓皮栎、麻栎和槲栎的平均多度在低海拔段(<200 m)的平均增加率要大于高海拔段(>2 000 m), 其中槲栎的平均多度在海拔>200 m区间减少。锐齿槲栎在海拔>2 000 m地区多度增加, 在其他海拔区间减少, RCP 8.5下增加(减少)的幅度大于RCP 2.6。蒙古栎的多度在海拔<100 m和1 000-2 000 m地区都是增加的, 且在海拔1 000-2 000 m区间, RCP 8.5下增加幅度大于RCP 2.6下(
图6 )。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6中国北方不同时期及排放情景下5种栎属植物在各海拔区间的多度平均变化率。A, 栓皮栎。B, 麻栎。C, 槲栎。D, 锐齿槲栎。E, 蒙古栎。 Fig. 6Elevation distribution of abundance change ratio of different periods under different scenario in North China. A, Quercus variabilis . B, Q. acutissima. C, Q. aliena. D, Q. aliena var. acuteserrata. E, Q. mongolica. 3 讨论 在未来不同的气候情景下, 研究区域平均升温1.9 ℃到4.4 ℃, 升温幅度最大的地方在研究区域东北部, 尤其是黑龙江省北部和内蒙古自治区东部, 升温幅度从1.9 ℃到5 ℃。在降水变化方面, 大部分地区未来向着更湿润的方向发展, 降水增幅最大出现在内蒙古自治区西部, 最暖季降水的增幅平均小于年降水量的增幅。RCP 8.5下的升温幅度与降水增幅都要高于RCP 2.6下。在2050年两个情景的预测中, 研究区域RCP 8.5下的栓皮栎、麻栎、槲栎和蒙古栎多度平均增加值都要大于RCP 2.6, 而锐齿槲栎的多度在RCP 8.5下减少值大于RCP 2.6下。到2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎在RCP 8.5下多度增加值都小于RCP 2.6下的值。也就是说, 5个物种在未来两个时期中RCP 8.5下多度增加(减少)的值都要大于RCP 2.6下的值。栓皮栎、麻栎和槲栎的多度在低海拔地区增加值高于高海拔地区, 而 锐齿槲栎和蒙古栎的多度则是在高海拔地区增加 较多。
在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 )。在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 )。对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 )。麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态。2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少。
本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度。但本研究也存在以下不足。气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 )。关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 )。此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子。除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 )。本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 )。此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度。
研究基于RF和基准气候、两个未来时期气候, 得到5种栎属植物多度的潜在分布和未来时期的多度变化。内蒙古自治区东北部和黑龙江省北部地区是本研究中5种栎属树木未来预测的多度集中减少的地区, 该地区也是在2050年和2070年升温幅度最大的区域。此外, 陕西省中部也是在2050年预测下, 5种栎属植物多度集中减小的地区。麻栎、槲栎、锐齿槲栎和蒙古栎在未来预测中, 都有一半以上区域的多度是减少的, 锐齿槲栎多度减少面积的比例达到70%以上。所以在未来气候变化下, 要加强对这几个区域的监测与物种保护。
附录 附录? 中国北方5种树种在2050年RCP 2.6和RCP 8.5下多度预测值 Supplement I Future distribution maps of abundance in the North China for five species under RCP 2.6 and RCP 8.5 scenarios in year 2050
http://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2018.0249-S1.pdf 附录II 中国北方5种树种在2070年RCP 2.6和RCP 8.5下多度预测值 Supplement II Future distribution maps of abundance in the North China for five species under RCP 2.6 and RCP 8.5 scenarios in year 2070
http://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2018.0249-S2.pdf [1] Austin MP Meyers JA 1996 ). Current approaches to modelling the environmental niche of eucalypts: Implication for management of forest biodiversityForest Ecology and Management ,95 -106 . [本文引用: 1] [2] Bellard C Bertelsmeier C Leadley P Thuiller W Courchamp F 2012 ). Impacts of climate change on the future of biodiversityEcology Letters ,365 -377 . [本文引用: 1] [3] Brito C Crespo EG Paulo OS 1999 ). Modelling wildlife distributions: Logistic multiple regression vs overlap analysisEcography ,251 -260 . [本文引用: 1] [4] Cao MC Zhou GS Weng ES 2005 ). Application and comparison of generalized models and classification and regression tree in simulating tree species distributionActa Ecologica Sinica ,2031 -2040 . [本文引用: 1] [ 曹铭昌 , 周广胜 , 翁恩生 ( 2005 ). 广义模型及分类回归树在物种分布模拟中的应用与比较生态学报 , 25, 2031 -2040 .] [本文引用: 1] [5] Editorial Board of the Chinese Vegetation Map, Chinese Academy of Sciences ( 2007 ). Vegetation Map of the People’s Republic of China (1: 1 000 000) . Geological Publishing House, Beijing. [本文引用: 1] [ 中国科学院中国植被图编辑委员会 ( 2007 ). 中华人民共和国植被图(1: 100万) . 地质出版社, 北京.] [本文引用: 1] [6] Chen IC Hill JK Ohlemuller R Roy DB Thomas CD 2011 ). Rapid range shifts of species associated with high levels of climate warmingScience ,1024 -1026 . [本文引用: 1] [7] Cutler DR Edwards Jr TC Beard KH Cutler A Hess KT Gibson J Lawler JJ 2007 ). Random forests for classification in ecologyEcology ,2783 -2792 . [本文引用: 1] [8] Cutler K Breiman L 2004 ). Random ForestsMachine Learning ,157 -176 . [本文引用: 2] [9] Domonkos P 2001 ). Temporal accumulations of extreme daily mean temperature anomaliesTheoretical and Applied Climatology ,17 -32 . [本文引用: 1] [10] Dubuis A Pottier J Rion V Pellissier L Theurillat JP Guisan A 2011 ). Predicting spatial patterns of plant species richness: A comparison of direct macroecological and species stacking modelling approachesDiversity and Distributions ,1122 -1131 . [本文引用: 1] [11] Ehrlén J Morris WF 2015 ). Predicting changes in the distribution and abundance of species under environmental changeEcology Letters ,303 -314 . [本文引用: 1] [12] Franklin J Franklin JF Franklin JD 2009 ). Mapping species distributions: Spatial inference and predictionQuarterly Review of Biology ,818 . [本文引用: 1] [13] Hao ZQ Dai LM He HS Malandnoff DJ Shao GF 2001 ). Potential response of major tree species to climate warming in Changbai Mountain, Northeast ChinaChinese Journal of Applied Ecology ,653 -658 . [本文引用: 1] [ 郝占庆 , 代力民 , 贺红士 , Malandnoff DJ 邵国凡 ( 2001 ). 气候变暖对长白山主要树种的潜在影响应用生态学报 , 12, 653 -658 .] [本文引用: 1] [14] Hastie T Tibshirani R 1986 ). Generalized additive modelsStatistical Science ,297 -310 . [本文引用: 1] [15] Hijmans RJ Cameron SE Parra JL Jones PG Jarvis A 2005 ). Very high resolution interpolated climate surfaces for global land areasInternational Journal of Climatology ,1965 -1978 . [本文引用: 1] [16] Iverson LR Prasad AM Matthews SN Peters M 2008 ). Estimating potential habitat for 134 eastern US tree species under six climate scenariosForest Ecology and Management ,390 -406 . [本文引用: 1] [17] Kaspari M 2001 ). Taxonomic level, trophic biology and the regulation of local abundanceGlobal Ecology and Biogeography ,229 -244 . [本文引用: 1] [18] Kearney M Porter WP 2004 ). Mapping the fundamental niche: Physiology, climate, and the distribution of a nocturnal lizardEcology ,3119 -3131 . [本文引用: 1] [19] Kharin VV Zwiers FW Zhang X Wehner M 2013 ). Changes in temperature and precipitation extremes in the CMIP5 ensembleClimatic Change ,345 -357 . [本文引用: 2] [20] Li GQ Liu CC Liu YG Yang J Zhang XS Guo K 2013 ). Advances in theoretical issues of species distribution modelsActa Ecologica Sinica ,4827 -4835 . [本文引用: 1] [ 李国庆 , 刘长成 , 刘玉国 , 杨军 , 张新时 , 郭柯 ( 2013 ). 物种分布模型的理论进展生态学报 , 33, 4827 -4835 .] [本文引用: 1] [21] Loehle C LeBlanc D 1996 ). Model-based assessments of climate change effects on forests—A critical reviewEcological Modelling ,1 -31 . [本文引用: 1] [22] Nelder JA Wedderburn RWM 1972 ). Generalized linear modelsJournal of the Royal Statistical Society A ,370 -384 . [本文引用: 1] [23] Parmesan C 2006 ). Ecological and evolutionary responses to recent climate changeAnnual Review of Ecology, Evolution, and Systematics ,637 -669 . [本文引用: 1] [24] Peng YS Chen L Li JQ 2007 ). Study on numerical taxonomy of Quercus L.( Fagaceae) in ChinaJournal of Wuhan Botanical Research, 149 -157 . [本文引用: 1] [ 彭焱松 , 陈丽 , 李建强 ( 2007 ). 中国栎属植物的数量分类研究武汉植物学研究 , 25, 149 -157 .] [本文引用: 1] [25] Picard R Cook RD 1984 ). Cross-validation of regression modelsJournal of the American Statistical Association ,575 -583 . [本文引用: 1] [26] Riahi K Rao S Krey V Cho C Chirkov V Fischer G Kindermann G Nakicenovic N Rafaj P 2011 ). RCP 8.5—A scenario of comparatively high greenhouse gas emissionsClimatic Change ,33 -57 . [本文引用: 1] [27] Rong YS Tu QP 2001 ). Characteristics of evolution of potential evaporation in Tianjin region of ChinaJournal of Hydrodynamics ,3 ), 16 -21 . [本文引用: 1] [28] Sainani KL 2013 ). Multivariate regression: The pitfalls of automated variable selectionPM&R , 5, 791 -794 . [本文引用: 1] [29] Spano D Cesaraccio C Duce P Snyder RL 1999 ). Phenological stages of natural species and their use as climate indicatorsInternational Journal of Biometeorology ,124 -133 . [本文引用: 1] [30] Stockwell D Peters D 1999 ). The GARP modelling system: Problems and solutions to automated spatial predictionInternational Journal of Geographical Information Science ,143 -158 . [本文引用: 1] [31] Sturm M Racine C Tape K 2001 ). Increasing shrub abundance in the ArcticNature ,546 -547 . [本文引用: 1] [32] Syphard AD Franklin J 2009 ). Differences in spatial predictions among species distribution modeling methods vary with species traits and environmental predictorsEcography ,907 -918 . [本文引用: 1] [33] Tessarolo G Rangel TF Araújo MB Hortal J 2014 ). Uncertainty associated with survey design in species distribution modelsDiversity and Distributions ,1258 -1269 . [本文引用: 1] [34] Thuiller W 2003 ). Biomod—Optimizing predictions of species distributions and projecting potential future shifts under global changeGlobal Change Biology ,1353 -1362 . [本文引用: 1] [35] Thuiller W Araújo MB Lavorel S 2003 ). Generalized models vs. classification tree analysis: Predicting spatial distributions of plant species at different scalesJournal of Vegetation Science ,669 -680 . [本文引用: 1] [36] van Vuuren DP Stehfest E den Elzen MGJ Kram T van Vliet J Deetman S Isaac M Klein Goldewijk K Hof A Mendoza Beltran A Oostenrijk R van Ruijven B 2011 ). RCP2.6: Exploring the possibility to keep global mean temperature increase below 2 °CClimatic Change ,95 -116 . [本文引用: 1] [37] Wang A Goslee SC Miller DA Sanderson MA Gonet JM 2017 ). Topographic variables improve climatic models of forage species abundance in the northeastern United StatesApplied Vegetation Science ,84 -93 . [本文引用: 3] [38] Wang ZW Ye JL Yao TD 1998 ). Construction of mean annual temperature series for the last one hundred years in ChinaQuarterly Journal of Applied Meteorology ,392 -401 . [本文引用: 1] [ 王昭武 , 叶瑾琳 , 姚檀栋 ( 1998 ). 近百年中国年气温序列的建立应用气象学报 , 9, 392 -401 .] [本文引用: 1] [39] Xu CH Xu Y 2012 ). The projection of temperature and precipitation over China under RCP scenarios using a CMIP5 Multi-Model ensembleAtmospheric & Oceanic Science Letters ,527 -533 . [本文引用: 1] [40] Yan YJ Li Y Wang WJ He JS Yang RH Wu HJ Wang XL Jiao L Tang ZY Yao YJ 2017 ). Range shifts in response to climate change of Ophiocordyceps sinensis, a fungus endemic to the Tibetan PlateauBiological Conservation, 143 -150 . [本文引用: 1] [41] Young M Carr MH 2015 ). Application of species distribution models to explain and predict the distribution, abundance and assemblage structure of nearshore temperate reef fishesDiversity and Distributions ,1428 -1440 . [本文引用: 1] [42] Zhou GS Zhang XS 1996 ). Study on Chinese climate-?vegetation relationshipActa Phytoecologica Sinica ,113 -119 . [本文引用: 1] [ 周广胜 , 张新时 ( 1996 ). 中国气候-植被关系初探植物生态学报 , 20, 113 -119 .] [本文引用: 1] [43] Zhou ZK 1992 ). Origin, phylogeny and dispersal of Quercus from ChinaActa Botanica Yunnanica, 227 -236 . [本文引用: 1] [ 周浙昆 ( 1992 ). 中国栎属的起源演化及其扩散云南植物研究 , 14, 227 -236 .] [本文引用: 1] Current approaches to modelling the environmental niche of eucalypts: Implication for management of forest biodiversity
1
1996
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
Impacts of climate change on the future of biodiversity
1
2012
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Modelling wildlife distributions: Logistic multiple regression vs overlap analysis
1
1999
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
广义模型及分类回归树在物种分布模拟中的应用与比较
1
2005
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
广义模型及分类回归树在物种分布模拟中的应用与比较
1
2005
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
1
2007
... 未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5.本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果.RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 ).以上气候数据按照研究区裁剪.在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, . ...
1
2007
... 未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5.本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果.RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 ).以上气候数据按照研究区裁剪.在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, . ...
Rapid range shifts of species associated with high levels of climate warming
1
2011
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Random forests for classification in ecology
1
2007
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
Random Forests
2
2004
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
Temporal accumulations of extreme daily mean temperature anomalies
1
2001
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
Predicting spatial patterns of plant species richness: A comparison of direct macroecological and species stacking modelling approaches
1
2011
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
Predicting changes in the distribution and abundance of species under environmental change
1
2015
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Mapping species distributions: Spatial inference and prediction
1
2009
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
气候变暖对长白山主要树种的潜在影响
1
2001
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
气候变暖对长白山主要树种的潜在影响
1
2001
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
Generalized additive models
1
1986
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
Very high resolution interpolated climate surfaces for global land areas
1
2005
... 研究使用的气候数据从世界气候数据库(http://www.worldclim.org/,
Hijmans et al ., 2005 )提取, 该数据库的Version 1.4收集了1960-2000年世界各地气象站记录的气象信息, 插值形成分辨率为30ʺ × 30ʺ (约1 km × 1 km)的全球气候栅格数据, 并基于每月气温和每月降水记录, 计算得到19个生物气候因子, 分别为年平均气温、气温日较差、等温性、气温季节性、最热月最高气温、最冷月最低气温、气温年较差、最湿季平均气温、最干季平均气温、最热季平均气温、最冷季平均气温、年降水量、最湿月降水量、最干月降水量、降水季节性、最湿季降水量、最干季降水量、最热季降水量和最冷季降水量. ...
Estimating potential habitat for 134 eastern US tree species under six climate scenarios
1
2008
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
Taxonomic level, trophic biology and the regulation of local abundance
1
2001
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Mapping the fundamental niche: Physiology, climate, and the distribution of a nocturnal lizard
1
2004
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
Changes in temperature and precipitation extremes in the CMIP5 ensemble
2
2013
... 未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5.本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果.RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 ).以上气候数据按照研究区裁剪.在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, . ...
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
物种分布模型的理论进展
1
2013
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
物种分布模型的理论进展
1
2013
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Model-based assessments of climate change effects on forests—A critical review
1
1996
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
Generalized linear models
1
1972
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
Ecological and evolutionary responses to recent climate change
1
2006
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
中国栎属植物的数量分类研究
1
2007
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
中国栎属植物的数量分类研究
1
2007
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
Cross-validation of regression models
1
1984
... 模型准确度用模型预测值与实际观测值之间的Pearson相关系数来表征(
Young & Carr, 2015 ).为验证模型的模拟质量, 用
k 折交叉验证来校验模型, 将数据集等分为
k 份, 每个子集均做一次测试集, 其余的作为训练集, 交叉验证重复
k 次.每次选择一个子集作为测试集, 并将
k 次的交叉验证平均值作为准确率, 这样, 所有的样本都可以被作为训练集和测试集, 每个样本都被验证一次, 可以充分利用已有数据集对算法效果进行测试, 本研究采用10折交叉验证, 最终准确度为100次10折交叉验证结果(
Picard & Denniscook, 1984 ).在5种栎属树木中, GLM对栓皮栎的拟合结果最佳(
r = 0.42); RF对栓皮栎、锐齿槲栎、麻栎的拟合结果较好(
r 值分别为0.61、0.63、0.53), 对槲栎的拟合结果较差(
r = 0.36).总体而言, RF对5种树木的拟合结果都要优于GAM和GLM (
图2 ), 因此在后续结果中只采用随机森林进行基准气候与未来气候情景下物种多度的预测.预测结果用100次10折交叉验证建立起来的模型预测的平均值来表示. ...
RCP 8.5—A scenario of comparatively high greenhouse gas emissions
1
2011
... 未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5.本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果.RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 ).以上气候数据按照研究区裁剪.在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, . ...
Characteristics of evolution of potential evaporation in Tianjin region of China
1
2001
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
Multivariate regression: The pitfalls of automated variable selection
1
2013
... GLM和GAM中, 增加自变量数目可以提高模型解释因变量的能力, 但具有多重共线性的多个自变量应用会导致模型过度拟合(
Sainani, 2013 ).为了避免过度拟合风险, 本研究通过计算19个气候因子的Pearson相关系数, 用单个因子对物种分别进行一次预模拟.基于模型的赤池信息量准则(AIC)值和各因子之间的相关性, 选取拟合度高且共线性小的因子(|
r | < 0.7)(
Yan et al ., 2017 ), 同时检测所选因子二次项是否显著.最终GLM包含所选因子的一次项和检验显著的二次项, GAM包含与线性模型相同的一次项因子.对于最终建立的GLM和GAM, 栓皮栎的预测模型包含了等温性、最热月最高气温、最冷月最低气温、降水季节性和最湿季降水量5个因子; 麻栎的预测模型包含了等温性、最冷月最低气温、气温年较差、最湿季平均气温、最干月降水量和最热季降水量6个变量; 槲栎的预测模型包含了等温性、最湿季平均气温、最干季平均气温、年降水量和最湿月降水量5个变量; 锐齿槲栎的预测模型包含等温性、最冷月最低气温、最湿季平均气温、最干月降水量和降水季节性等5个变量; 蒙古栎的预测模型包含了年平均气温、等温性、气温季节性、最湿季平均气温、年降水量、最湿月降水量和最热季降水量等7个变量.由于RF中不存在共线性问题, 使用19个气候因子作为自变量, 二叉树变量和森林数目选择使模型残差最小的值. ...
Phenological stages of natural species and their use as climate indicators
1
1999
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
The GARP modelling system: Problems and solutions to automated spatial prediction
1
1999
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
Increasing shrub abundance in the Arctic
1
2001
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Differences in spatial predictions among species distribution modeling methods vary with species traits and environmental predictors
1
2009
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
Uncertainty associated with survey design in species distribution models
1
2014
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
Biomod—Optimizing predictions of species distributions and projecting potential future shifts under global change
1
2003
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
Generalized models vs. classification tree analysis: Predicting spatial distributions of plant species at different scales
1
2003
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
RCP2.6: Exploring the possibility to keep global mean temperature increase below 2 °C
1
2011
... 未来气候变化情景选取中国气象局北京气候中心(BCC-CSM 1.1)提供的大气环流模型结果, 该结果预测了4种典型浓度路径(RCP)以表征不同温室气体排放情境, 即RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5.本研究中采用了该气候模型的两个极端排放情景, 即RCP 2.6和RCP 8.5的气候模拟结果.RCP 2.6情景设定温室气体的综合排放水平在未来先上升后下降, 辐射强度为4种情景中最低; RCP 8.5情景设定没有减排措施, 温室气体的排放水平在未来持续增加, 辐射强度为4种情景中最高, 气候变化状况最显著(
Riahi et al ., 2011 ;
van Vuuren et al ., 2011 ;
Kharin et al ., 2013 ).以上气候数据按照研究区裁剪.在预测时用1:100万植被分布图(
中国科学院中国植被图编辑委员会, 2007 )对气候图层进行掩膜裁剪, 提取其中森林和灌丛植被类型, . ...
Topographic variables improve climatic models of forage species abundance in the northeastern United States
3
2017
... 本文选取了广义线性模型(GLM;
Nelder & Wedderburn, 1972 )、广义加性模型(GAM;
Hastie & Tibshirani, 1986 )和随机森林模型(RF;
Cutler & Breiman, 2004 )来模拟物种多度与环境的关系.这几种模型已被成功用于树木和灌丛的分布模型(
Thuiller, 2003 ;
Thuiller et al ., 2003 ;
Dubuis et al. , 2011 ;
Wang et al ., 2017 ). ...
... GLM具有较强的预测物种分布的能力(
Austin & Meyers, 1996 ;
Brito et al ., 1999 ); GAM适用于响应变量与解释变量之间的非线性关系, 本研究中的样方并未覆盖物种完整的生态位, 在事先没有关于响应变量与解释变量关系的明确假设时, GAM可以很好地探索这种关系(
Wang et al ., 2017 ).RF是基于决策树的集成算法, 不用对函数形式进行事先假定, 避免了假设误差, 不需要顾虑一般回归分析面临的多元共线性的问题, 不用做变量选择(
Cutler & Breiman, 2004 ), 被誉为当前最好的算法之一(
Iverson et al ., 2008 ), 在物种分布模拟中得到了广泛应用(
Cutler et al ., 2007 ). ...
... 本研究以物种的绝对多度作为因变量, 多度相较于物种存在数据, 反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响在区域和洲际尺度.但本研究也存在以下不足.气候因素是控制物种分布的主要环境因子, 随着空间尺度的减小, 其他的一些因素, 如地形、土壤等将会对物种分布产生重大影响(
曹铭昌等, 2005 ).关于草场草本植物的研究已经发现, 相对于只有气候因子的多度解释模型, 地形因子的加入可以提升至少10%的模型解释率(
Wang et al ., 2017 ).此外, 物种的扩散还有可能受到地形阻碍, 本研究也没有区分个体的大小及其他个体差异, 研究设定多度与气候的关系在基准气候与未来气候中保持不变, 在探究物种多度对于未来气候变化的响应时, 也没有考虑物种之间的相互作用及其他影响物种分布的环境因子.除气候因素外, 以上这些因素都会影响物种的分布(
Tessarolo et al ., 2014 ).本研究对于当前5种树木多度的模拟是根据当前的实际多度所建立起来的模型, 已经包含了捕食、竞争以及人类活动的影响, 因此并不属于“基础生态位”的模拟, 但在未来气候条件下预测时, 只考虑了气候的变化而没有考虑捕食、竞争等种间关系的变化以及人类活动的干扰, 其结果则更接近于基础生态位的预测(
Stockwell & Peters, 1999 ;
Kearney & Porter, 2004 ).此外, 虽然本文研究的5种树木大部分分布区均处于中国境内, 境外分布区较小, 但这些数据的缺失也限制了SDM模型的精度. ...
近百年中国年气温序列的建立
1
1998
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
近百年中国年气温序列的建立
1
1998
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
The projection of temperature and precipitation over China under RCP scenarios using a CMIP5 Multi-Model ensemble
1
2012
... 在2050年完全没有减排措施的RCP 8.5下, 栓皮栎、麻栎、槲栎和蒙古栎的多度平均增加值都要大于RCP 2.6下, 可能跟该情境下温度的季节性变异和极端低温事件大大减少有关(
Xu & Xu, 2012 ;
Kharin et al ., 2013 ).在2070年比较切合实际的低浓度排放情景下, 栓皮栎、麻栎、蒙古栎的多度平均值也是增加的, 可能与气候变暖导致区域最高气温和最低气温的升高, 使生长季延长有关(
Loehle & LeBlanc, 1996 ).对于许多北方阔叶树, 气温上升5 ℃依然在其适应生长的范围之内(
郝占庆等, 2001 ), 而在增温背景下, 潜在蒸散会增加, 对水资源的消耗将更加显著(
Rong & Tu, 2001 ).麻栎、槲栎、锐齿槲栎和蒙古栎的多度在未来预测中, 有超过一半面积的区域都处于减少状态.2050年和2070年, 栓皮栎、麻栎、槲栎、锐齿槲栎和蒙古栎的多度都在内蒙古自治区东北部、黑龙江省北部地区减少, 可能是由于内蒙古自治区东北部、黑龙江省北部在未来气候情景中降水量增幅较小, 5个物种的多度在该地区集中减少可能是由于降水量减少或者降水量增加不足以平衡气温升高和潜在蒸散的加大(
Domonkos, 2001 ), 此外, 5个物种的多度在2050年都在陕西省中部地区减少, 可能是因为陕西省中部地区在2050年RCP 2.6下出现年降水量减少. ...
Range shifts in response to climate change of
Ophiocordyceps sinensis, a fungus endemic to the Tibetan Plateau
1
2017
... GLM和GAM中, 增加自变量数目可以提高模型解释因变量的能力, 但具有多重共线性的多个自变量应用会导致模型过度拟合(
Sainani, 2013 ).为了避免过度拟合风险, 本研究通过计算19个气候因子的Pearson相关系数, 用单个因子对物种分别进行一次预模拟.基于模型的赤池信息量准则(AIC)值和各因子之间的相关性, 选取拟合度高且共线性小的因子(|
r | < 0.7)(
Yan et al ., 2017 ), 同时检测所选因子二次项是否显著.最终GLM包含所选因子的一次项和检验显著的二次项, GAM包含与线性模型相同的一次项因子.对于最终建立的GLM和GAM, 栓皮栎的预测模型包含了等温性、最热月最高气温、最冷月最低气温、降水季节性和最湿季降水量5个因子; 麻栎的预测模型包含了等温性、最冷月最低气温、气温年较差、最湿季平均气温、最干月降水量和最热季降水量6个变量; 槲栎的预测模型包含了等温性、最湿季平均气温、最干季平均气温、年降水量和最湿月降水量5个变量; 锐齿槲栎的预测模型包含等温性、最冷月最低气温、最湿季平均气温、最干月降水量和降水季节性等5个变量; 蒙古栎的预测模型包含了年平均气温、等温性、气温季节性、最湿季平均气温、年降水量、最湿月降水量和最热季降水量等7个变量.由于RF中不存在共线性问题, 使用19个气候因子作为自变量, 二叉树变量和森林数目选择使模型残差最小的值. ...
Application of species distribution models to explain and predict the distribution, abundance and assemblage structure of nearshore temperate reef fishes
1
2015
... 模型准确度用模型预测值与实际观测值之间的Pearson相关系数来表征(
Young & Carr, 2015 ).为验证模型的模拟质量, 用
k 折交叉验证来校验模型, 将数据集等分为
k 份, 每个子集均做一次测试集, 其余的作为训练集, 交叉验证重复
k 次.每次选择一个子集作为测试集, 并将
k 次的交叉验证平均值作为准确率, 这样, 所有的样本都可以被作为训练集和测试集, 每个样本都被验证一次, 可以充分利用已有数据集对算法效果进行测试, 本研究采用10折交叉验证, 最终准确度为100次10折交叉验证结果(
Picard & Denniscook, 1984 ).在5种栎属树木中, GLM对栓皮栎的拟合结果最佳(
r = 0.42); RF对栓皮栎、锐齿槲栎、麻栎的拟合结果较好(
r 值分别为0.61、0.63、0.53), 对槲栎的拟合结果较差(
r = 0.36).总体而言, RF对5种树木的拟合结果都要优于GAM和GLM (
图2 ), 因此在后续结果中只采用随机森林进行基准气候与未来气候情景下物种多度的预测.预测结果用100次10折交叉验证建立起来的模型预测的平均值来表示. ...
中国气候-植被关系初探
1
1996
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
中国气候-植被关系初探
1
1996
... 植物分布-气候关系是生态学和地理学研究的热点问题, 也是预估未来气候变化对生态系统影响的实现基础(
周广胜和张新时, 1996 ;
Chen et al ., 2011 ;
Bellard et al ., 2012 ).物种分布模型或生态位模型通过统计方法描述物种对气候的响应关系, 判断物种的潜在分布区, 并预测气候变化情景下物种的未来分布区(
Franklin et al ., 2009 ).传统的物种分布模型通常以物种的分布区或者分布点的物种存在数据(即出现与否)作为物种分布的响应变量(
李国庆等, 2013 ).然而, 植物有多种方式响应气候变化, 除了改变其分布区以外, 还会改变其生长物候、生理活动强度, 或者是多度(
Spano et al ., 1999 ;
Sturm et al ., 2001 ;
Parmesan, 2006 ).相较于物种存在数据, 多度反映了一个物种占用资源并把资源分配给个体的能力, 更能衡量物种对区域生态系统的影响(
Kaspari, 2001 ;
Syphard & Franklin, 2009 ;
Ehrlén & Morris, 2015 ). ...
中国栎属的起源演化及其扩散
1
1992
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
中国栎属的起源演化及其扩散
1
1992
... 近100年以来, 华北及周边地区经历了高于全国平均水平的升温趋势, 年平均气温增温速率达到1.1 ℃·100 a
-1 (
王昭武等, 1998 ).栎属(
Quercus )是壳斗科中最大的属, 分布极为广泛, 在欧亚从印度尼西亚的爪哇岛直到北欧62° N的斯堪迪纳亚半岛和52° N的鄂霍茨克海域附近(
周浙昆, 1992 ).栎属树种是温带落叶阔叶林的优势成分, 分布于全国各地(
彭焱松等, 2007 ), 也是我国华北及周边地区常见的森林优势种.在气候变化背景下, 准确预测栎属植物的未来分布, 将促进华北地区的植被动态、植被管理以及脆弱生态系统的修复.本文通过大量野外调查, 获取了华北地区常见栎属树种的多度, 利用物种分布模型拟合其多度分布并预测其在未来气候变化下的多度分布. ...
 ,
, ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT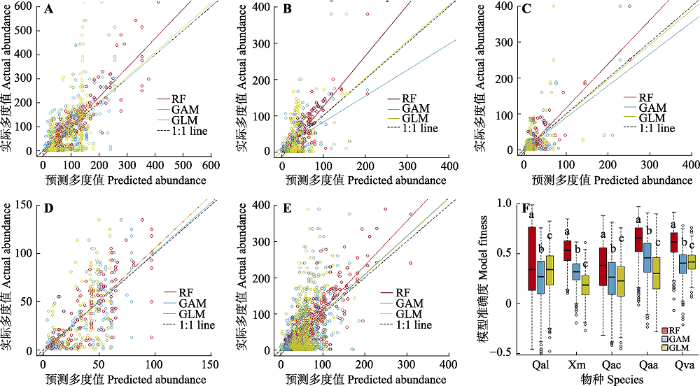 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT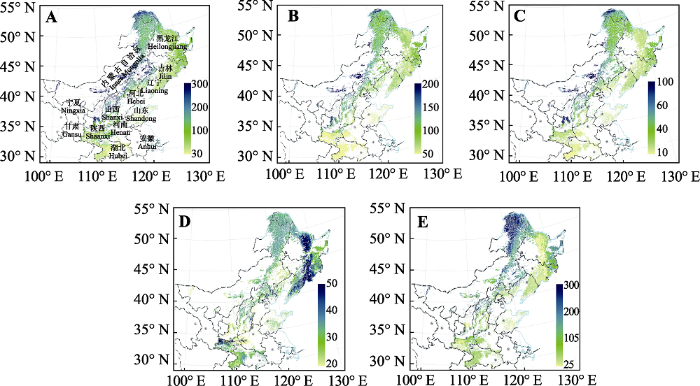 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT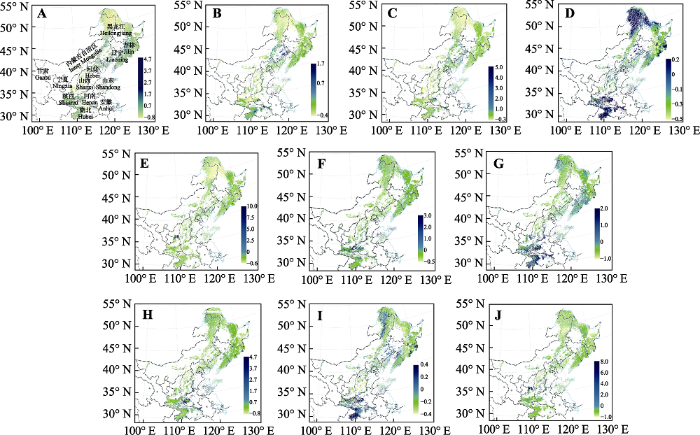 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT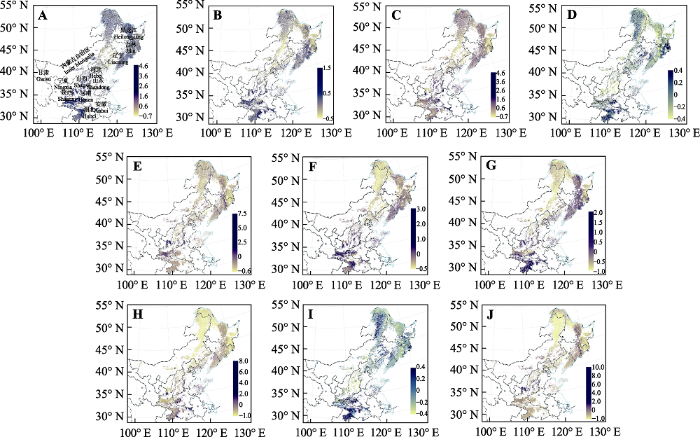 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT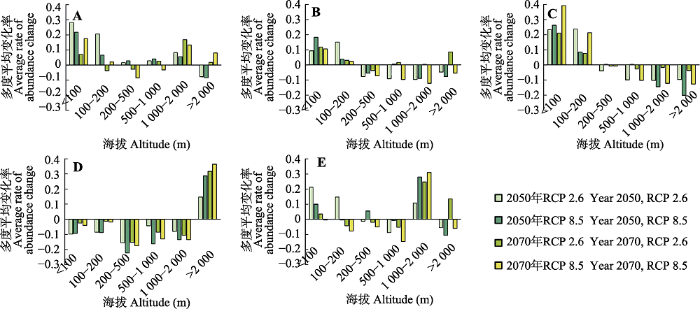 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}