Community assembly of herbaceous layer of the planted forests in the central Loess Plateau, China
Jing-Jing SHI1,2, Ming-Fei ZHAO1,2, Yu-Hang WANG1,2, Feng XUE1,2, Mu-Yi KANG3,1, Yuan JIANG,1,2,3,*1College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 2Beijing Key Laboratory of Chinese Medicine Resource for Protection and Utilization, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 3State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China
Supported by the National Basic Work of Science and Technology of China(2011FY110300) the National Natural Science Foundation of China(41630750) the National Natural Science Foundation of China(41271059)
Abstract Aims Many competing theories seek to identify the mechanisms behind the community assembly. Niche theory and neutral theory are among the two most competing mechanisms. Exploring factors controlling the patterns of β-diversity is an approach to reveal the underlying mechanisms of community assembly. In this paper, we aim to reveal the β-diversity pattern of understory in planted forests of the central Loess Plateau and to evaluate the relative influences of deterministic (environmental filtering) and stochastic (dispersal limitation) processes on the community assembly. Methods We sampled in total 107 sites of planted forests spatially spreading over the central Loess Plateau. In each site, three plots (10 m × 10 m) were randomly designed each at a distance of about 50 m. The floristic composition, community structure, and environmental conditions were investigated in each plot. We calculated the dissimilarity (βsim) of species composition, geographic distance and environmental divergence for each of the 5 671 site pairs. We used random forest (RF) to quantify the influence of the selected factors on species composition, and distance-based multivariate regression method (MRM) to fit the relationship between dissimilarity, geographic distance and environmental divergence. To quantify how and in what extent the environmental filtering and the dispersal limitation influence the species composition among sites, we performed the principle coordinates of neighbor matrices (PCNM) and distance-based RDA (db-RDA) to select the pronounced independent variables. We then partitioned the variance into three parts, namely, 1) independent contributions of geographic distance; 2) independent contributions of environmental divergence; and 3) the joint contribution of both. Important findings The community similarity declined with increasing geographical distance and environmental divergence. Geographical distance and environmental divergence together explained 47.8% of variance in the dissimilarity in species composition. The spatial variables accounted for 14.1%, while the environmental variables accounted for another 9.8% of the variance in dissimilarity of species composition. Annual precipitation and basal area of trees are the two most important environmental factors influencing the pattern of dissimilarity of species composition. These results implied that both environmental filtering and the dispersal limitation play important roles in shaping the community assembly of the herbaceous layers of planted forests in the central Loess Plateau. Keywords:community assembly;niche theory;neutral theory;environmental filtering;dispersal limitation;β diversity;understory community
PDF (1911KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 施晶晶, 赵鸣飞, 王宇航, 薛峰, 康慕谊, 江源. 黄土高原腹地人工林下草本层群落构建机制. 植物生态学报, 2019, 43(9): 834-842. DOI: 10.17521/cjpe.2018.0153 SHI Jing-Jing, ZHAO Ming-Fei, WANG Yu-Hang, XUE Feng, KANG Mu-Yi, JIANG Yuan. Community assembly of herbaceous layer of the planted forests in the central Loess Plateau, China. Chinese Journal of Plant Ecology, 2019, 43(9): 834-842. DOI: 10.17521/cjpe.2018.0153
Fig. 2Results of random forest (RF) model and multiple regression method (MRM) based on distance matrices. The importance of the ecological factors and the test results of significance are shown in A1 and A2, respectively; B and C respectively shows the relationship between dissimilarity of species composition with increasing geographical and environmental distance. AP, annual precipitation; Asp, aspect; BA, basal area; Den, density; DeltaH, delta height; Ele, elevation; Geo, geographical distance; HC, herbaceous coverage; MAT, mean annual temperature; Slo, slope; TC, tree coverage; TH, tree height. Dashed line represents Lowess line, solid for linear fitted regression line.
Table 1 表1 表1黄土高原腹地人工林下草本群落物种组成变差分解结果(%) Table 1Results of variation partitioning of understory species composition of the planted forests in the central Loess Plateau (%)
分解部分 Partitioned parts
分解结果 Value of each part
纯环境因子解释部分 Pure interpretation of environmental variables
9.8
纯空间因子解释部分 Pure interpretation of geographic variables
本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017)。研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016)。反之, 则随机过程的作用更为重要。如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用。而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位。与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异。因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014)。与Bohlman等(2018)在亚马孙地区的研究得出的结论相似。
AdlerPB, HilleRisLambersJ, LevineJM ( 2007). A niche for neutrality Ecology Letters, 10, 95-104. [本文引用: 1]
AndersonMJ, CristTO, ChaseJM, VellendM, InouyeBD, FreestoneAL, SandersNJ, CornellHV, ComitaLS, DaviesKF, HarrisonSP, KraftNJB, StegenJC, SwensonNG ( 2011). Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist Ecology Letters, 14, 19-28. [本文引用: 2]
BaselgaA ( 2010). Partitioning the turnover and nestedness components of beta diversity Global Ecology and Biogeography, 19, 134-143. [本文引用: 1]
BohlmanSA, LauranceWF, LauranceSG, NascimentoHEM, FearnsidePM, AndradeA ( 2008). Importance of soils, topography and geographic distance in structuring central Amazonian tree communities Journal of Vegetation Science, 19, 863-874. [本文引用: 1]
BorcardD, LegendreP ( 2002). All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices Ecological Modelling, 153, 51-68. [本文引用: 1]
BurrascanoS, SabatiniFM, BlasiC ( 2011). Testing indicators of sustainable forest management on understorey composition and diversity in southern Italy through variation partitioning Plant Ecology, 212, 829-841. [本文引用: 1]
ChaseJM ( 2014). Spatial scale resolves the niche versus neutral theory debate Journal of Vegetation Science, 25, 319-322. [本文引用: 3]
ChaseJM, MyersJA ( 2011). Disentangling the importance of ecological niches from stochastic processes across scales Philosophical Transactions of the Royal Society of London, 366, 2351-2363. [本文引用: 2]
ChustG, ChaveJ, ConditR, AguilarS, LaoS, PérezR ( 2006). Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama Journal of Vegetation Science, 17, 83-92. [本文引用: 1]
ConditR, HubbellSP ( 2002). Βeta-diversity in tropical forest trees Science, 295, 666-669. [本文引用: 2]
DixonP ( 2003). VEGAN, a package of R functions for community ecology Journal of Vegetation Science, 14, 927-930. [本文引用: 1]
DonoghueMJ ( 2008). A phylogenetic perspective on the distribution of plant diversity Proceedings of the National Academy of Sciences of the United States of America, 105, 11549-11555. [本文引用: 1]
EhrlingerJ ( 2015). ggRandomForests: Visually Exploring a Random Forests
FuBJ, LiuY, LüY, HeCS, ZengY, WuBF ( 2011). Assessing the soil erosion control service of ecosystems change in the Loess Plateau of China Ecological Complexity, 8, 284-293. [本文引用: 1]
GeorgeLO, BazzazFA ( 2014). The herbaceous layer as a filter determining spatial pattern in forest tree regeneration In: Gilliam FS, Roberts MR eds. The Herbaceous Layer in Forests of Eastern North America. Oxford University Press, New York. [本文引用: 1]
GibsonN, ProberS, MeissnerR, Van LeeuwenS ( 2017). Implications of high species turnover on the south-western Australian sandplains PLOS ONE, 12, e0172977. DOI: 10.1371/journal.pone.0172977. [本文引用: 3]
GilliamFS ( 2007). The ecological significance of the herbaceous layer in temperate forest ecosystems Bioscience, 57, 845-858. [本文引用: 1]
GilliamFS, HockenberryAW, AdamsMB ( 2006). Effects of atmospheric nitrogen deposition on the herbaceous layer of a Central Appalachian hardwood forest Journal of the Torrey Botanical Society, 133, 240-254. [本文引用: 1]
GosleeSC, UrbanDL, LeeuwJD, ZeileisA ( 2007). The ecodist package for dissimilarity-based analysis of ecological data Journal of Statistical Software, 22, 1-19. [本文引用: 1]
GravelD, CanhamCD, BeaudetM, MessierC ( 2006). Reconciling niche and neutrality: The continuum hypothesis Ecology Letters, 9, 399-409. [本文引用: 1]
GrimeJP ( 1977). Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory The American Naturalist, 111, 1169-1194. [本文引用: 1]
HubbellSP ( 2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton, USA. [本文引用: 1]
IhaddadenA, VelázquezE, Rey-BenayasJM, Kadi-HanifiH ( 2013). Climate and vegetation structure determine plant diversity in Quercus ilex woodlands along an aridity and human-use gradient in Northern Algeria Flora, 208, 268-284. [本文引用: 1]
JamoneauA, PassySI, SoininenJ, LeboucherT, Tison- RoseberyJ ( 2018). Βeta diversity of diatom species and ecological guilds: Response to environmental and spatial mechanisms along the stream watercourse Freshwater Biology, 63, 62-73. [本文引用: 1]
JonesMM, GibsonN, YatesC, FerrierS, MokanyK, WilliamsKJ, ManionG, SvenningJC ( 2016). Underestimated effects of climate on plant species turnover in the Southwest Australian Floristic Region Journal of Biogeography, 43, 289-300. [本文引用: 1]
JonesMM, TuomistoH, BorcardD, LegendreP, ClarkDB, OlivasPC ( 2008). Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality Oecologia, 155, 593-604. [本文引用: 1]
KraftNJB, ComitaLS, ChaseJM, SandersNJ, SwensonNG, CristTO, StegenJC, VellendM, BoyleB, AndersonMJ, CornellHV, DaviesKF, FreestoneAL, InouyeBD, HarrisonSP, MyersJA ( 2011). Disentangling the drivers of β diversity along latitudinal and elevational gradients Science, 333, 1755-1758. [本文引用: 1]
LegendreP, de CáceresM ( 2013). Βeta diversity as the variance of community data: Dissimilarity coefficients and partitioning Ecology Letters, 16, 951-963. [本文引用: 1]
LegendreP, LapointeFJ, CasgrainP ( 1994). Modeling brain evolution from behavior: A permutational regression approach Evolution, 48, 1487-1499. [本文引用: 1]
LegendreP, MiXC, RenHB, MaKP, YuMJ, SunIF, HeFL ( 2009). Partitioning beta diversity in a subtropical broad- leaved forest of China Ecology, 90, 663-674.
LeiboldMA, McPeekMA ( 2006). Coexistence of the niche and neutral perspectives in community ecology Ecology, 87, 1399-1410. [本文引用: 1]
LiuHY, YinY, WangQY, HeSY ( 2015). Climatic effects on plant species distribution within the forest-steppe ecotone in northern China Applied Vegetation Science, 18, 43-49. [本文引用: 1]
LiuQF, KangMY, LiuQR ( 2006). Quantitative classification and environmental interpretation of forest tree species in Hungou, Zhongtiao Mountain Journal of Plant Ecology (Chinese Version), 30, 383-391. [本文引用: 1]
MárialigetiS, TinyaF, BidlóA, ódorP ( 2016). Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary Plant Ecology, 217, 549-563. [本文引用: 1]
MarmionM, LuotoM, HeikkinenRK, ThuillerW ( 2009). The performance of state-of-the-art modelling techniques depends on geographical distribution of species Ecological Modelling, 220, 3512-3520. [本文引用: 1]
MayF, HuthA, WiegandT ( 2015). Moving beyond abundance distributions: Neutral theory and spatial patterns in a tropical forest Proceedings of the Royal Society B Biological Sciences, 282, 20141657. DOI: 10.1098/rspb.2014.1657. [本文引用: 1]
MouquetN, LoreauM ( 2003). Community patterns in source- sink metacommunities The American Naturalist, 162, 544-557. [本文引用: 1]
MyersJA, ChaseJM, JiménezI, J?rgensenPM, Araujo-?MurakamiA, Paniagua-ZambranaN, SeidelR ( 2013). Βeta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly Ecology Letters, 16, 151-157. [本文引用: 1]
PageNV, ShankerK ( 2018). Environment and dispersal influence changes in species composition at different scales in woody plants of the Western Ghats, India Journal of Vegetation Science, 29, 74-83. [本文引用: 4]
PetermannJS, KratinaP, MarinoNAC, MacDonaldAAM, SrivastavaDS ( 2015). Resources alter the structure and increase stochasticity in bromeliad microfauna communities PLOS ONE, 10, e0118952. DOI: 10.1371/journal. pone.0118952. [本文引用: 1]
QianH ( 2009). Βeta diversity in relation to dispersal ability for vascular plants in North America Global Ecology and Biogeography, 18, 327-332. [本文引用: 1]
Réjou-MéchainM, PélissierR, Gourlet-FleuryS, CouteronP, NasiR, ThompsonJD ( 2008). Regional Variation in tropical forest tree species composition in the Central African Republic: An assessment based on inventories by forest companies Journal of Tropical Ecology, 24, 663-674. [本文引用: 1]
SabatiniF, Jiménez-AlfaroB, BurrascanoS, BlasiC ( 2014). Drivers of herb-layer species diversity in two unmanaged temperate forests in northern Spain Community Ecology, 15, 147-157. [本文引用: 1]
SauraS, Pascual-HortalL ( 2007). A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study Landscape and Urban Planning, 83, 91-103. [本文引用: 1]
SeidlerTG, PlotkinJB ( 2006). Seed dispersal and spatial pattern in tropical trees PLOS Biology, 4, e344. DOI: 10.1371/journal.pbio.0040344. [本文引用: 1]
SiefertA, RavenscroftC, WeiserMD, SwensonNG ( 2013). Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees Global Ecology and Biogeography, 22, 682-691. [本文引用: 1]
SmithTW, LundholmJT ( 2010). Variation partitioning as a tool to distinguish between niche and neutral processes Ecography, 33, 648-655. [本文引用: 1]
SunL, ZhangGH, LuanLL, LiZW, GengR ( 2016). Distribution of soil organic carbon in surface soil along a precipitation gradient in loess hilly Chinese Journal of Applied Ecology, 27, 532-538. [本文引用: 1]
TanLZ, FanCY, ZhangCY, von GadowK, FanXH ( 2017). How beta diversity and the underlying causes vary with sampling scales in the Changbai mountain forests Ecology and Evolution, 7, 10116-10123. [本文引用: 1]
TangZY, FangJY, ChiXL, YangYH, MaWH, MohhamotA, GuoZD, LiuYN, GastonKJ ( 2012). Geography, environment, and spatial turnover of species in China’s grasslands Ecography, 35, 1103-1109. [本文引用: 2]
ToledoM, PoorterL, Pe?a-ClarosM, AlarcónA, BalcázarJ, Chuvi?aJ, Lea?oC, LiconaJC, ter SteegeH, BongersF ( 2011). Patterns and determinants of floristic variation across lowland forests of Bolivia Biotropica, 43, 405-413. [本文引用: 1]
TuomistoH, RuokolainenK, VormistoJ, DuqueA, SánchezM, ParedesVV, L?hteenojaO ( 2017). Effect of sampling grain on patterns of species richness and turnover in Amazonian forests Ecography, 40, 840-852. [本文引用: 1]
van OijenD, FeijenM, HommelP, den OudenJ, de WaalR ( 2005). Effects of tree species composition on within-?forest distribution of understorey species Applied Vegetation Science, 8, 155-166. [本文引用: 1]
VianaDS, FiguerolaJ, SchwenkK, MancaM, Hob?kA, MjeldeM, PrestonCD, GornallRJ, CroftJM, KingRA, GreenAJ, SantamaríaL ( 2016). Assembly mechanisms determining high species turnover in aquatic communities over regional and continental scales Ecography, 39, 281-288. [本文引用: 1]
WangD, WangXA, GuoH, WangSX, ZhenWN, LiuSL ( 2013). Effect of species dispersal and environmental factors on species assemblages in grassland communities Acta Ecologica Sinica, 33, 4409-4415. [本文引用: 4]
WangSX, ZhaoL, LiN, GuoH, WangXA, DuanRY ( 2016). Community heterogeneity of undergrowth vegetation in Pinus tabuliformis forest on the Loess Plateau of Northwest China Chinese Journal of Ecology, 35, 1197-1203. [本文引用: 1]
WrightDH, CurrieDJ, MaurerBA ( 1993). Energy supply and patterns of species richness on local and regional scales In: Ricklef RE, Schluter D eds. Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago. 66-74. [本文引用: 1]
XiaoJF ( 2014). Satellite evidence for significant biophysical consequences of the “Grain for Green” Program on the Loess Plateau in China Journal of Geophysical Research Biogeosciences, 119, 2261-2275. [本文引用: 1]
YouYM, XuJY, CaiDX, LiuSR, ZhuHG, WenYG ( 2016). Environmental factors affecting plant species diversity of understory plant communities in a Castanopsis hystrix plantation chronosequence in Pingxiang, Guangxi, China Acta Ecologica Sinica, 36, 164-172. [本文引用: 2]
YuM, ZhouZY, KangFF, OuyangS, MiXC, SunJX ( 2013). Gradient analysis and environmental interpretation of understory herb-layer communities in Xiaoshegou of Lingkong Mountain, Shanxi, China Chinese Journal of Plant Ecology, 37, 373-383. [本文引用: 1]
ZhaoMF, WangGY, XingKX, WangYH, XueF, KangMY, LuoK ( 2017). Patterns and determinants of species similarity decay of forest communities in the western Qinling Mountains Biodiversity Science, 25, 3-10. [本文引用: 1]
Importance of soils, topography and geographic distance in structuring central Amazonian tree communities 1 2008
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices 1 2002
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
... 对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
Disentangling the importance of ecological niches from stochastic processes across scales 2 2011
... 揭示群落物种空间分布格局和动态变化机制即群落构建机制是群落生态学的基本目标(Smith & Lundholm, 2010; May et al., 2015; Gibson et al., 2017).生态位理论(确定过程)和中性理论(随机过程)是阐明该机制的两种主要理论(Page & Shanker, 2018), 但这两个理论对群落构建过程的相对重要性目前仍未达成广泛共识(Leibold & McPeek, 2006).生态位理论认为物种组成差异格局是环境异质性的产物, 共存物种间的生态位分化、资源竞争等确定性因素是主要决定因子(Chase & Myers, 2011).然而, 中性理论则认为群落中相同营养级所有个体在生态上等价, 差异主要由扩散限制和生态漂变等随机作用主导(Hubbell, 2001).近年来的研究则倾向于整合两个理论的观点, 认为确定过程和随机过程均在群落构建过程中发挥着重要作用(Gravel et al., 2006; Adler et al., 2007; Chase & Myers, 2011). ...
... ; Chase & Myers, 2011). ...
Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama 1 2006
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
Βeta-diversity in tropical forest trees 2 2002
... β多样性格局反映群落间物种组成的变化情况(Anderson et al., 2011; Legendre & de Cáceres, 2013), 揭示β多样性格局与空间距离和环境异质性之间的关系可衡量生态位理论和中性理论对群落构建过程的相对重要性(Condit et al., 2002; Kraft et al., 2011).若β多样性格局主要受环境过滤影响, 则环境变量对样地间物种组成变化的解释率将最高(Petermann et al., 2015; Jamoneau et al., 2018).若中性理论占主导地位, 则扩散限制是β多样性格局的主要影响因素, 植物种子的拓殖速率与地理距离之间将呈负相关关系.换言之, 地理距离或者空间属性(如采样粒度、研究范围等)将在群落构建过程中扮演更为重要的角色(Seidler & Plotkin, 2006; Page & Shanker, 2018). ...
... 量化确定性和随机过程对群落构建的相对重要性, 可综合考虑空间因子、环境条件以及物种本身的功能属性差异等因素(Condit & Hubbell, 2002), 通过变差分解法分解群落间β多样性的差异进行分析(Legendre et al., 2009; Burrascano et al., 2011; Myers et al., 2013).Gibson等(2017)在澳大利亚西南部沙地平原的研究中指出, 模型所选变量(气候、土壤和空间变量)共可解释群落物种组成变化的87%, 变差分解的结果表明空间和气候因子解释了绝大部分变化.除了变差分解, 相似性随距离的衰减斜率也是研究群落构建机制的常用方法(Anderson et al., 2011; Siefert et al., 2013).Tang等(2013)在中国北方主要草原群落的研究表明, 控制环境“空间自相关”的影响后, 群落物种组成相似性随地理距离和环境差异的增大而减小. ...
Βeta- diversity in tropical forest trees 2002
VEGAN, a package of R functions for community ecology 1 2003
... 揭示群落物种空间分布格局和动态变化机制即群落构建机制是群落生态学的基本目标(Smith & Lundholm, 2010; May et al., 2015; Gibson et al., 2017).生态位理论(确定过程)和中性理论(随机过程)是阐明该机制的两种主要理论(Page & Shanker, 2018), 但这两个理论对群落构建过程的相对重要性目前仍未达成广泛共识(Leibold & McPeek, 2006).生态位理论认为物种组成差异格局是环境异质性的产物, 共存物种间的生态位分化、资源竞争等确定性因素是主要决定因子(Chase & Myers, 2011).然而, 中性理论则认为群落中相同营养级所有个体在生态上等价, 差异主要由扩散限制和生态漂变等随机作用主导(Hubbell, 2001).近年来的研究则倾向于整合两个理论的观点, 认为确定过程和随机过程均在群落构建过程中发挥着重要作用(Gravel et al., 2006; Adler et al., 2007; Chase & Myers, 2011). ...
Climate and vegetation structure determine plant diversity in Quercus ilex woodlands along an aridity and human-use gradient in Northern Algeria 1 2013
... 纯环境变量的独立解释率为9.8%, 通过RF和MRM分析, 进一步发现环境因子中年降水量与胸高断面积和对物种组成变化的影响最大.首先, 研究选取的人工林下草本群落位于半湿润-半干旱区的黄土高原, 对于该区的草本物种而言, 水分是其一大胁迫因子, 因此该区物种多样性通常与降水显著相关(Wright et al., 1993).其次, 研究区各样地间年降水量差异较大.而由降水梯度驱动形成的物种组成变化格局也已被证实, Toledo等(2011)在新热带区(Neoptropics)的研究以及Réjou-Méchain等(2008)在非洲热带森林的研究均表明群落物种组成受降水梯度的影响.但不同于先前的研究, 气候因子中的年平均气温在本区域中发挥的作用相对较小, 研究对象不同可能是造成这一差异的主要原因.上述研究多关注于木本植物, 而本研究则主要着眼于林下草本植物.上层植被可以发挥缓冲作用, 如吸收一定的太阳辐射和削弱风对林下草本层的影响(Liu et al., 2015), 从而减少温度变化对林下物种组成的作用.第三, 胸高断面积变量在一定程度上能够反映林分的冠层结构特征(尤业明等, 2016), 而冠层结构能够影响林分内土壤水分、光照环境以及温湿度等微生境条件的变化(van Oijen et al., 2005), 这进一步影响了林下草本物种组成.Ihaddaden等(2013)在阿尔及利亚北部干旱区以及余敏等(2013)在山西小蛇沟的研究也表明冠层结构对林下草本群落物种组成具有重要影响. ...
Βeta diversity of diatom species and ecological guilds: Response to environmental and spatial mechanisms along the stream watercourse 1 2018
... β多样性格局反映群落间物种组成的变化情况(Anderson et al., 2011; Legendre & de Cáceres, 2013), 揭示β多样性格局与空间距离和环境异质性之间的关系可衡量生态位理论和中性理论对群落构建过程的相对重要性(Condit et al., 2002; Kraft et al., 2011).若β多样性格局主要受环境过滤影响, 则环境变量对样地间物种组成变化的解释率将最高(Petermann et al., 2015; Jamoneau et al., 2018).若中性理论占主导地位, 则扩散限制是β多样性格局的主要影响因素, 植物种子的拓殖速率与地理距离之间将呈负相关关系.换言之, 地理距离或者空间属性(如采样粒度、研究范围等)将在群落构建过程中扮演更为重要的角色(Seidler & Plotkin, 2006; Page & Shanker, 2018). ...
Underestimated effects of climate on plant species turnover in the Southwest Australian Floristic Region 1 2016
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality 1 2008
Regional Variation in tropical forest tree species composition in the Central African Republic: An assessment based on inventories by forest companies 1 2008
... 纯环境变量的独立解释率为9.8%, 通过RF和MRM分析, 进一步发现环境因子中年降水量与胸高断面积和对物种组成变化的影响最大.首先, 研究选取的人工林下草本群落位于半湿润-半干旱区的黄土高原, 对于该区的草本物种而言, 水分是其一大胁迫因子, 因此该区物种多样性通常与降水显著相关(Wright et al., 1993).其次, 研究区各样地间年降水量差异较大.而由降水梯度驱动形成的物种组成变化格局也已被证实, Toledo等(2011)在新热带区(Neoptropics)的研究以及Réjou-Méchain等(2008)在非洲热带森林的研究均表明群落物种组成受降水梯度的影响.但不同于先前的研究, 气候因子中的年平均气温在本区域中发挥的作用相对较小, 研究对象不同可能是造成这一差异的主要原因.上述研究多关注于木本植物, 而本研究则主要着眼于林下草本植物.上层植被可以发挥缓冲作用, 如吸收一定的太阳辐射和削弱风对林下草本层的影响(Liu et al., 2015), 从而减少温度变化对林下物种组成的作用.第三, 胸高断面积变量在一定程度上能够反映林分的冠层结构特征(尤业明等, 2016), 而冠层结构能够影响林分内土壤水分、光照环境以及温湿度等微生境条件的变化(van Oijen et al., 2005), 这进一步影响了林下草本物种组成.Ihaddaden等(2013)在阿尔及利亚北部干旱区以及余敏等(2013)在山西小蛇沟的研究也表明冠层结构对林下草本群落物种组成具有重要影响. ...
Drivers of herb-layer species diversity in two unmanaged temperate forests in northern Spain 1 2014
... 林下草本层是森林生态系统的重要组成部分(Gilliam, 2007), 在森林生态系统中往往占有最高比例的物种多样性(占据森林物种丰富度的90%以上) (Gilliam et al., 2006), 其不仅能促进森林生态系统养分循环, 稳定林分生产力(Sabatini et al., 2014; Márialigeti et al., 2016), 还能通过与乔木幼苗之间的资源竞争影响乔木种苗的存活、发育与生长(George & Bazzaz, 2014).然而, 相比于乔木层物种多样性, 草本层物种多样性尚未引起广泛关注(尤业明等, 2016).鉴于黄土高原的特殊性以及林下草本层的重要性, 本研究拟探究以下两个科学问题: (1)黄土高原地区人工林下草本群落β多样性呈怎样的格局?(2)环境过滤和扩散限制对人工林下草本群落构建的相对重要性如何? ...
A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study 1 2007
Seed dispersal and spatial pattern in tropical trees 1 2006
... β多样性格局反映群落间物种组成的变化情况(Anderson et al., 2011; Legendre & de Cáceres, 2013), 揭示β多样性格局与空间距离和环境异质性之间的关系可衡量生态位理论和中性理论对群落构建过程的相对重要性(Condit et al., 2002; Kraft et al., 2011).若β多样性格局主要受环境过滤影响, 则环境变量对样地间物种组成变化的解释率将最高(Petermann et al., 2015; Jamoneau et al., 2018).若中性理论占主导地位, 则扩散限制是β多样性格局的主要影响因素, 植物种子的拓殖速率与地理距离之间将呈负相关关系.换言之, 地理距离或者空间属性(如采样粒度、研究范围等)将在群落构建过程中扮演更为重要的角色(Seidler & Plotkin, 2006; Page & Shanker, 2018). ...
Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees 1 2013
... 量化确定性和随机过程对群落构建的相对重要性, 可综合考虑空间因子、环境条件以及物种本身的功能属性差异等因素(Condit & Hubbell, 2002), 通过变差分解法分解群落间β多样性的差异进行分析(Legendre et al., 2009; Burrascano et al., 2011; Myers et al., 2013).Gibson等(2017)在澳大利亚西南部沙地平原的研究中指出, 模型所选变量(气候、土壤和空间变量)共可解释群落物种组成变化的87%, 变差分解的结果表明空间和气候因子解释了绝大部分变化.除了变差分解, 相似性随距离的衰减斜率也是研究群落构建机制的常用方法(Anderson et al., 2011; Siefert et al., 2013).Tang等(2013)在中国北方主要草原群落的研究表明, 控制环境“空间自相关”的影响后, 群落物种组成相似性随地理距离和环境差异的增大而减小. ...
Variation partitioning as a tool to distinguish between niche and neutral processes 1 2010
... 揭示群落物种空间分布格局和动态变化机制即群落构建机制是群落生态学的基本目标(Smith & Lundholm, 2010; May et al., 2015; Gibson et al., 2017).生态位理论(确定过程)和中性理论(随机过程)是阐明该机制的两种主要理论(Page & Shanker, 2018), 但这两个理论对群落构建过程的相对重要性目前仍未达成广泛共识(Leibold & McPeek, 2006).生态位理论认为物种组成差异格局是环境异质性的产物, 共存物种间的生态位分化、资源竞争等确定性因素是主要决定因子(Chase & Myers, 2011).然而, 中性理论则认为群落中相同营养级所有个体在生态上等价, 差异主要由扩散限制和生态漂变等随机作用主导(Hubbell, 2001).近年来的研究则倾向于整合两个理论的观点, 认为确定过程和随机过程均在群落构建过程中发挥着重要作用(Gravel et al., 2006; Adler et al., 2007; Chase & Myers, 2011). ...
黄土丘陵区表层土壤有机碳沿降水梯度的分布 1 2016
... 地处黄土高原腹地的研究区(34.47°-39.32° N, 107.57°-111.53° E), 面积约1.0 × 105 km2, 最高海拔约2 000 m, 最低海拔约800 m.该区气候具有典型的温带大陆性季风气候特征, 年平均气温3.6- 14.3 ℃, 年降水量150-750 mm, 且降水主要发生在6-9月.降水最丰富的东南部地区年降水量为600- 750 mm, 而西部和西北部仅150-250 mm, 呈现出从东南向西北显著降低的水分梯度.研究区由暖温带向温带过渡, 植被带跨越了落叶阔叶林带、森林草原带和草原带等(孙龙等, 2016). ...
黄土丘陵区表层土壤有机碳沿降水梯度的分布 1 2016
... 地处黄土高原腹地的研究区(34.47°-39.32° N, 107.57°-111.53° E), 面积约1.0 × 105 km2, 最高海拔约2 000 m, 最低海拔约800 m.该区气候具有典型的温带大陆性季风气候特征, 年平均气温3.6- 14.3 ℃, 年降水量150-750 mm, 且降水主要发生在6-9月.降水最丰富的东南部地区年降水量为600- 750 mm, 而西部和西北部仅150-250 mm, 呈现出从东南向西北显著降低的水分梯度.研究区由暖温带向温带过渡, 植被带跨越了落叶阔叶林带、森林草原带和草原带等(孙龙等, 2016). ...
How beta diversity and the underlying causes vary with sampling scales in the Changbai mountain forests 1 2017
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
Geography, environment, and spatial turnover of species in China’s grasslands 2 2012
... 量化确定性和随机过程对群落构建的相对重要性, 可综合考虑空间因子、环境条件以及物种本身的功能属性差异等因素(Condit & Hubbell, 2002), 通过变差分解法分解群落间β多样性的差异进行分析(Legendre et al., 2009; Burrascano et al., 2011; Myers et al., 2013).Gibson等(2017)在澳大利亚西南部沙地平原的研究中指出, 模型所选变量(气候、土壤和空间变量)共可解释群落物种组成变化的87%, 变差分解的结果表明空间和气候因子解释了绝大部分变化.除了变差分解, 相似性随距离的衰减斜率也是研究群落构建机制的常用方法(Anderson et al., 2011; Siefert et al., 2013).Tang等(2013)在中国北方主要草原群落的研究表明, 控制环境“空间自相关”的影响后, 群落物种组成相似性随地理距离和环境差异的增大而减小. ...
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
Patterns and determinants of floristic variation across lowland forests of Bolivia 1 2011
... 纯环境变量的独立解释率为9.8%, 通过RF和MRM分析, 进一步发现环境因子中年降水量与胸高断面积和对物种组成变化的影响最大.首先, 研究选取的人工林下草本群落位于半湿润-半干旱区的黄土高原, 对于该区的草本物种而言, 水分是其一大胁迫因子, 因此该区物种多样性通常与降水显著相关(Wright et al., 1993).其次, 研究区各样地间年降水量差异较大.而由降水梯度驱动形成的物种组成变化格局也已被证实, Toledo等(2011)在新热带区(Neoptropics)的研究以及Réjou-Méchain等(2008)在非洲热带森林的研究均表明群落物种组成受降水梯度的影响.但不同于先前的研究, 气候因子中的年平均气温在本区域中发挥的作用相对较小, 研究对象不同可能是造成这一差异的主要原因.上述研究多关注于木本植物, 而本研究则主要着眼于林下草本植物.上层植被可以发挥缓冲作用, 如吸收一定的太阳辐射和削弱风对林下草本层的影响(Liu et al., 2015), 从而减少温度变化对林下物种组成的作用.第三, 胸高断面积变量在一定程度上能够反映林分的冠层结构特征(尤业明等, 2016), 而冠层结构能够影响林分内土壤水分、光照环境以及温湿度等微生境条件的变化(van Oijen et al., 2005), 这进一步影响了林下草本物种组成.Ihaddaden等(2013)在阿尔及利亚北部干旱区以及余敏等(2013)在山西小蛇沟的研究也表明冠层结构对林下草本群落物种组成具有重要影响. ...
Effect of sampling grain on patterns of species richness and turnover in Amazonian forests 1 2017
Assembly mechanisms determining high species turnover in aquatic communities over regional and continental scales 1 2016
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
... 本研究结果显示, 相比于纯环境因子, 群落间物种组成相似性变化更多地被空间因子所解释(表1), 这可能是因为随机过程和确定过程的相对作用大小具有尺度效应(Chase, 2014; Tan et al., 2017).研究尺度越大, 环境异质性越高, 环境因子对群落物种组成的过滤作用越明显(Viana et al., 2016).反之, 则随机过程的作用更为重要.如Jones等(2016)对澳大利亚西南部植物区系、Tang等(2013)对中国三大主要草原群落物种周转格局的大尺度研究结果均表明环境过滤发挥了更为重要的作用.而Chust等(2006)对巴拿马热带森林乔木β多样性格局以及王丹等(2013)对子午岭草地群落的小尺度研究则表明地理距离在群落构建过程中占主导地位.与上述小尺度研究相比, 虽然本研究区域南北跨度约达509 km, 东西距离约为200 km, 但可能由于黄土高原区相对均质的气候和土壤等环境条件使得研究区仍然没有包含足够的环境差异.因而即使研究区资源相对贫乏, 水热条件较为严苛, 样地间的相似生境依然减弱了环境过滤对物种周转的作用(Chase, 2014).与Bohlman等(2018)在亚马孙地区的研究得出的结论相似. ...
 ,1,2,3,*
,1,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT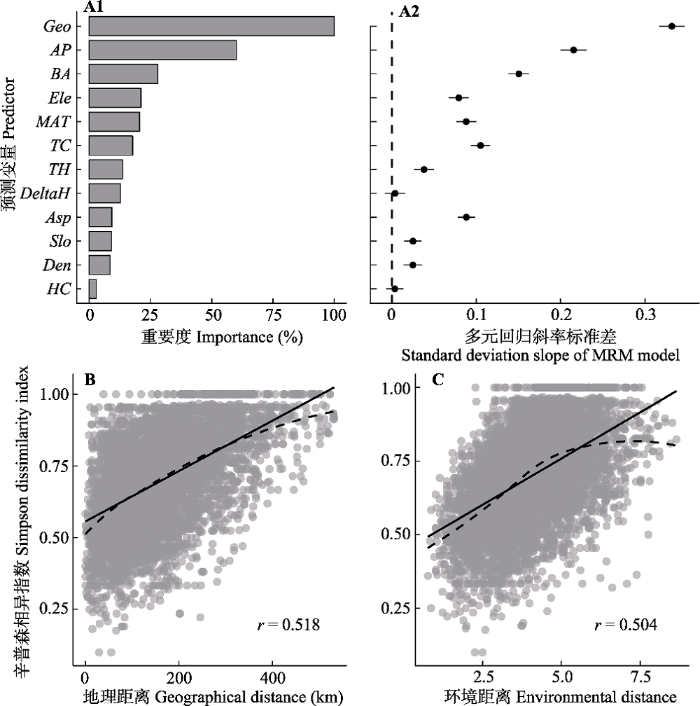 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}