李全弟, 刘旻霞
, * , 夏素娟, 南笑宁, 蒋晓轩西北师范大学地理与环境科学学院, 兰州 730070
Changes in species-abundance relationships of plant communities with slopes in alpine meadows of Gannan, China Quan-Di LI, Min-Xia LIU
, * , Su-Juan XIA, Xiao-Ning NAN, Xiao-Xuan JIANGCollege of Geography and Environmental Sciences, Northwest Normal University, Lanzhou 730070, China
通讯作者: *(
xiaminl@163.com )
编委: 李镇清
责任编辑: 李敏, 实习编辑: 赵航
收稿日期: 2018-12-18
接受日期: 2019-05-12
网络出版日期: 2019-05-20
基金资助: 国家自然科学基金 (31760135 )国家自然科学基金 (31360114 )
Editorial board: Li Zhen-Qing Editor: LI Min, ZHAO Hang Received: 2018-12-18
Accepted: 2019-05-12
Online: 2019-05-20
Fund supported: Supported by the National Natural Science Foundation of China (31760135 )Supported by the National Natural Science Foundation of China (31360114 )
摘要 物种-多度格局研究是揭示群落组织结构和物种区域分布规律的重要手段。该研究以青藏高原东北部的甘南高寒草甸为研究对象, 基于野外调查和室内分析, 研究了不同坡向的环境因子、植物群落分布, 并利用RAD软件程序包对其进行了拟合分析。结果显示: 在南坡-北坡上, 土壤含水量从南坡(0.18 g·g
-1 )到北坡(0.31 g·g
-1 )呈现递增的趋势, 土壤温度从南坡(22.33 ℃)到北坡(18.13 ℃)以及光照强度从南坡(744.15 lx)到北坡(681.93 lx)均呈逐渐减小的趋势。物种-多度分布曲线的斜率从南坡向北坡依次减小。随着坡向由南向北转变, 物种-多度和物种多样性都呈递增的趋势。通过6个模型对坡向梯度的物种-多度分布进行拟合发现, 甘南高寒草甸区的物种-多度分布主要是以生态位模型为主, 其次是随机分布模型。青藏高原高寒草甸微生境梯度上的物种在总体上的资源分配模式是以固定分配模式为主, 稀有种的资源分配模式是以随机性模型为主, 常见种的资源分配模式则是以确定性模型为主。
关键词: 甘南高寒草甸 ;
坡向 ;
物种-多度 ;
常见种 ;
稀有种 Abstract Aims The study of species diversity patterns is useful in understanding community structure and resource allocation patterns in the succession process in alpine meadows. The present study aims to explore: 1) the changes in community species-abundance relationships in alpine meadows of the Qinghai-Xizang Plateau with mountain slopes; and 2) the difference between the abundances of common and rare species, and their roles in the ecological mechanisms.
Methods Field investigation was carried out in alpine meadows of the Qinghai-Xizang Plateau followed by lab analyses in Lanzhou. RAD was used to analyze the difference in environmental factors and plant communities among different slope aspects.
Important findings Soil water content increased gradually from the south to north slopes (from 0.18 g·g
-1 on the south slope to 0.31 g·g
-1 on the north slope), while soil temperature and light intensity showed the opposite trends (from 22.33 ℃ and 744.15 lx on the south slope to 18.13 ℃ and 681.93 lx on the north slope, respectively). The species abundance and species diversity increased, while the slope of species abundance curves decreased from the south to north slopes. Among the six species abundance fitting models, the ecological niche model best explained the species abundance curves followed by the random distribution model. Further analysis showed that the overall resource allocation pattern of species was mainly fixed distribution. In addition, the resource allocation pattern of the rare species was mainly random, while that of the common species was mainly deterministic.
Keywords: Gannan alpine meadow ;
slope gradient ;
species-abundance ;
common species ;
rare species PDF (1251KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 李全弟, 刘旻霞, 夏素娟, 南笑宁, 蒋晓轩. 甘南高寒草甸群落的物种-多度关系沿坡向的变化. 植物生态学报, 2019, 43(5): 418-426. DOI:
10.17521/cjpe.2018.0317 LI Quan-Di, LIU Min-Xia, XIA Su-Juan, NAN Xiao-Ning, JIANG Xiao-Xuan.
Changes in species-abundance relationships of plant communities with slopes in alpine meadows of Gannan, China .
Chinese Journal of Plant Ecology , 2019, 43(5): 418-426. DOI:
10.17521/cjpe.2018.0317 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力。同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 )。物种-多度模式是指群落中物种-多度组成的比例关系。将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 )。物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据。分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据。一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高。在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 )。最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型。
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落。模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型。这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 )。就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 )。生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述。其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主。
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果。目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少。
青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 )。由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减。因此保护青藏高原高寒草甸区的生态系统刻不容缓。坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化。因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义。
本研究采用RAD软件程序包对各个坡向的物种-多度进行模型拟合, 拟通过坡向这一微生境梯度的变化来研究高寒草甸物种-多度的分布格局, 以期指导对群落发展和群落构建过程中的资源分配研究, 并建立拟合模型来分析解决以下两个问题: 1)随着坡向从南到北的变化中, 青藏高原高寒草甸区群落物种-多度分布格局是如何变化的?2)常见种和稀有种多度格局有何不同?在生态学机制(即维持物种多样性及生态平衡)中起到什么重要作用?
1 材料和方法 1.1 试验区概况 本试验是在青藏高原甘南高寒草甸区进行调查采样(
表1 ), 该地区主要的气候类型为寒冷湿润的高原气候, 降水集中在6-8月, 平均年降水量为531 mm; 极端最高气温28 ℃, 极端最低气温-23 ℃, 年平均气温为2.4 ℃。土壤为高寒草甸土。其植被类型以多年生草本植物为主, 主要物种有矮生嵩草(
Kobresia humilis ), 火绒草(
Leontopodium leontopodioides ), 三芒草(
Aristida adscensionis ), 珠芽蓼(
Polygonum viviparum ), 山莴苣(
Lagedium sibiricum ), 米口袋(
Gueldenstaedtia verna ), 莓叶委陵菜(
Potentilla fragarioides ), 金露梅(
Potentilla fruticosa ), 长毛风毛菊(
Saussurea hieracioides ), 甘肃马先蒿(
Pedicularis kansuensis )等。
Table 1 表1 表1 甘南高寒草甸实验样地区域概况(平均值±标准误差)
Table 1
Alpine meadows of Gannan overview of the studied area (mean ± SE ) 坡向 经度 纬度 坡向定位 坡度 海拔 南坡 South 34.93 102.9 0° 30.7 ± 1.5a 3 009 南偏东 Southeast 34.93 102.9 45° 31.0 ± 2.3a 3 001 东坡 East 34.93 102.9 90° 22.8 ± 2.8b 3 000 北偏东 Northeast 34.93 102.9 135° 14.9 ± 2.4c 3 001 北坡 North 34.93 102.9 180° 16.5 ± 3.5c 2 900
Different lowercase letters mean significant differences (p < 0.05). 不同小写字母表示差异显著(p < 0.05)。 新窗口打开 |
下载CSV 1.2 试验方法 2017年7月16号, 通过电子罗盘定位选取正南(0°)、南偏东(45°)、正东(90°)、北偏东(135°)、正北(180°) 5个坡向, 每个坡向顺着山体垂直方向设置4 个样方。每个样方的大小为50 cm × 50 cm, 调查并记录每个样方中物种的盖度、高度以及每个物种的多度。并在每个样方内取0-20 cm土壤测土壤含水量, 光照强度和土壤温度分别采用照度计和土温计从8:00-18:00每隔1 h测量一次, 每组5个重复。
1.3 模型介绍 常见模型有随机分配模型(Rapo)(
Hubbell, 2001 )、随机分类模型(Rane)(
Tokeshi, 1990 )、随机ZM模型(Zipf_Mandelbrot)、断棍模型(bro)(
Tokeshi, 1993 )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 )。
1.3.1 随机模型
Motomura (1932) 在研究植物群落演替的过程中提出了生态位优先占领模型(geo), 从而在几何级数模型的基础上进一步完善该模型。几何级数模型是按物种-多度值从小到大的顺序进行排列, 计算公式如下式:
(1) ${{A}_{i}}=E[p{{(1-p)}^{i-1}}]$ 其中,
Ai 为第
i 个物种的物种-多度值,
E 为总的资源量,
p 为最重要的物种占有资源比例。该模型体现的是生态位假说, 即第一优势种优先占领有限资源的一部分, 第二优势种又占领余下资源的一部分, 顺推而下, 直到所剩的资源不够一个物种生存为止。
MacArthur (1957) 提出了最为经典的断棍模型(bro)。模型假设的是一个群落中所有的资源在一条棍子上, 在棍上随机选取(
s - 1)个点, 把棍分成
s 份, 则代表有
s 个物种占有该群落的所有资源, 每个物种的竞争力是相似的。以
J/S (
J 表示该群落中所有物种-多度之和,
S 是指群落中物种总和)表示该群落里平均相对多度, 即该模型的第
i 个种的多度
Ai 可以表示为:
(2) ${{A}_{j}}=J/S\underset{X=\ i}{\overset{s}{\mathop \sum }}\,\frac{1}{x}\ \ (i=1,2,3\cdots )$ 该模型假设各个物种的多度是彼此独立的, 每个物种的多度假设是随机放在线段上的两点间的长度, 因为每段距离都有重叠的部分, 所以叫生态位重叠模型。
1.3.2 随机分配模型
假定群落中各个物种-多度之间是没有联系的, 环境及外界环境的变化等随机因素导致物种-多度大小与生态位分化无关, 一般情况下物种不能全部占用其生态位, 群落内部的生态位是不饱和的。当群落中有
z 个物种的时候,
Ai 为该物种的多度,
ran 则为不饱和状态下的参数值,
z 为物种数。第
i 个物种的相对多度为:
(3) ${{A}_{i}}={{A}_{\text{min}(i-1)}}ra{{n}^{z}}$ 1.3.3 统计模型
Fisher等(1943) 首次提出对数级数模型(Lser), 该模型属于经验模型, 用来拟合能够满足下面等式的物种-多度数据:
(4) $F(n)=\alpha {{X}^{n}}/n$ 式中,
n = 1, 2, …,
X 是与样本有关的参数,
α 是不受样本大小影响的参数, 反映了群落的内在性质, 且
α > 0。因
F (
n )正比于对数的幂级数展开项, 故称为对数级数或对数序列分布。
1.4 数据分析及优势度的检验 本文通过对不同坡向重要值的计算来确定常见种和稀有种。通过对重要值(
M )的计算和实际试验样地的综合考虑, 将重要值大于0.01的物种作为常见种进行研究, 小于0.01的物种则作为稀有种进行研究(
王育松和上官铁梁, 2010 )。
$M=\frac{相对多度+相对盖度+相对频度+相对高度}{4}$
采用RAD拟合程序包, 以fit411.txt文件的处理方法进行拟合。RAD是一款独立多批次处理数据的编程软件, 通过前期固定编程, 在读取数据(多度数据应按txt文本进行整理)时根据不同命令进行读取, 再根据软件自带参数拟合包进行数据批量处理, 最后输出一个名为Assemblage.txt的文本, 该文本里包含了
r 值(
r 是用模型的
n 个副本的平均密度进行计算, 采用最小二乘法中加入密度最大差异和物种数量的校正因子, 从而增强了模型拟合优度的区分能力)、
Oc 值(经过Ulrich校正过的Preston倍程分组后的χ
2 检验法)、
CL (置信区间)、相对多度、最大密度差、相对丰富度的标准差、多样性指数、均匀度指数等, 最后通过Excel进行分栏并根据相关参数(
r 值、
Oc 值、
CL )进行模型的选取。fit411.txt是指RAD软件在后期读取数的一个匹配文本, 里面的相关参数与相应的模型相互匹配, 这些参数包含了物种数、流量数、所匹配的模型名称、软件运行的迭代器、迭代次数、密度、置信区间及校正模式等, 而所有参数值都采用程序包中的默认值。在优势度的检验上, 采取
r 值、
Oc 值、
CL 值3个指标。通过多次迭代最后得到使
r 值最小的理论密度值即为所求值。因为
r 值受物种最大密度差和群落物种数目影响。所以RAD对物种数目
S 和最大密度差
D 进行了修正。
r 小于或接近10说明拟合效果较好(算法在倍频次运算时会无限接近于10), 大于100表明拟合效果较差(
Ulrich, 2002 )。RAD同样输出Preston倍程分组后的拟合变量
Oc 值, 该值的计算方法与
r 值相似, 将同一多度物种的理论与实测密度之间的欧氏距离换成同一倍程组别的。同样当
Oc 值小于或接近10 (算法在倍频次运算时会无限接近于10)的时候拟合效果较好而当其大于100的时候拟合效果较差。同时RAD还给出在
x %置信区间内的物种频率(
CL , 一般默认为是大于或等于95%的置信区间), 尽管该拟合优度检验方法不如
r 值、
Oc 值灵敏, 但对于随机模型的拟合优度检验非常重要(
肖美玲, 2014 )。
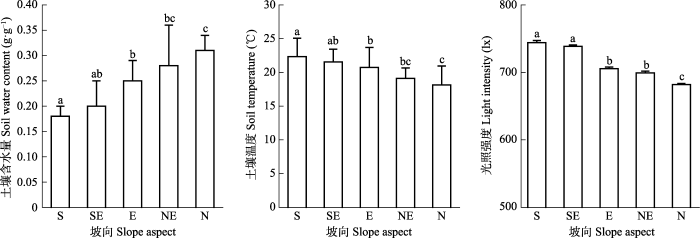
2 结果 2.1 坡向梯度上环境因子的变化特征 如
图1 所示, 在南-北坡梯度上, 土壤含水量从南坡(0.18 g·g
-1 )到北坡(0.31 g·g
-1 )呈现递增的趋势, 南坡、东坡和北坡有显著差异(
p < 0.05); 土壤温度从南坡(22.33 ℃)到北坡(18.13 ℃)呈现递减的趋势, 南坡、东坡和北坡之间存在显著差异(
p < 0.05); 光照强度从南坡(744.15 lx)到北坡(681.93 lx)呈现递减的趋势, 南坡和东南坡之间无显著性差异(
p > 0.05), 东坡和东北坡之间无显著差异(
p > 0.05), 南坡、东坡和北坡之间存在显著差异(
p < 0.05)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1甘南高寒草甸不同坡向的环境因子(平均值+标准误差)。不同小写字母表示差异显著(p < 0.05)。S, 南坡; SE, 东南坡; E, 东坡; NE, 东北坡; N, 北坡。 Fig. 1Environmental factors in different slopes in alpine meadows of Gannan (mean + SE ). Different lowercase letters mean significant differences (p < 0.05). S, South; SE, Southeast; E, East; NE, Northeast; N, North. 2.2 坡向梯度物种-多度的拟合模型分析 从南坡拟合效果(
表2 )来看, 6个模型都可以拟合, 但拟合效果
r 值均在25左右,
Oc 值接近于10, 置信区间
CL 是1。就东南坡拟合效果来看, Norm和ZM、geo模型拟合得较好。从东坡拟合效果来看, Rapo、bro、Norm、geo 4个模型的拟合效果相似, Rane模型在置信区间(0.95)上相比其他几个模型稍有不足。在东北坡上能拟合的模型有Rapo、Rane、Norm、geo 4个模型, 其中拟合效果最好的是Norm模型, 它的
r 值和
Oc 值都小于100并且接近于10, 其次它的置信区间
CL 为1。geo模型北坡的拟合
r 值为15.77,
Oc 值为16.30, 均接近于10,
CL 值为1, 则表明geo模型拟合效果最好。Rapo和geo模型对5个坡向都有较好的拟合效果。
Table 2 表2 表2 甘南高寒草甸不同坡向物种-多度分布模型拟合
Table 2
Species abundance models fitted in different slopes of Alpine meadows of Gannan 坡向 Slope aspect 模型 Model r Oc CL 南坡 Rapo 33.91 5.20 1.00 Rane 33.75 5.20 1.00 ZM 21.88 23.75 1.00 bro 27.22 5.18 1.00 Norm 19.75 3.24 1.00 geo 33.00 3.12 1.00 东南坡 Rapo 89.11 3.47 1.00 Rane 114.40 3.47 1.00 ZM 4.22 9.75 1.00 Bro 11.72 0.76 0.96 Norm 4.21 1.54 1.00 geo 11.72 4.63 1.00 东坡 Rapo 42.06 8.83 1.00 Rane 40.27 8.82 0.95 ZM 24.71 8.27 0.68 bro 77.89 8.036 0.89 Norm 10.77 8.04 1.00 geo 40.14 9.71 1.00 东北坡 Rapo 27.12 16.80 0.95 Rane 25.80 22.64 0.93 ZM 35.36 100.49 1.00 bro 29.67 25.03 1.00 Norm 24.29 10.21 1.00 geo 28.38 21.90 1.00 北坡 Rapo 22.76 22.30 0.96 Rane 22.08 79.73 1.00 ZM 32.34 48.52 0.95 bro 79.72 22.50 1.00 Norm 579.43 143.45 0.00 geo 15.77 16.30 1.00
r, a correction factor of maximum density difference and species number is added in the least square method; Oc, Preston's χ2 test after doubling grouping; CL, confidence interval. bro, broken stick; geo, geometric series; Norm, log normal; Rane, random assortment; Rapo, random fraction; ZM, Zipf_Mandelbrot.r, 采用最小二乘法中加入密度最大差异和物种数量的校正因子; Oc, Preston倍程分组后的χ2 检验法; CL, 置信区间。bro, 断棍模型; geo, 几何级数模型; Norm, 正态分布模型; Rane, 随机分类模型; Rapo, 随机分配模型; ZM, 随机ZM模型。新窗口打开 |
下载CSV 同一模型对不同坡向的拟合结果显示, geo模型对5个坡向的拟合效果整体而言还是比较理想的。该模型在5个坡向的拟合效果的优度排序为北坡(180°) >东北坡(135°) >东南坡(45°) >东坡(90°) >南坡(0°)。本研究所选的6个模型中Rapo模型、Norm模型和bro模型都能拟合5个坡向的物种-多度分布。但随着坡向从南向北的变化过程中, 物种-多度的分布明显不同, 拟合的匹配模型也是不同的。物种-多度格局模型导致不同物种在不同坡向上的资源分配模式也发生变化, 同时也导致群落构建的生态学过程发生变化。
2.3 不同坡向物种-多度曲线 由
图2 可以看出, 坡向在从南坡到北坡的变化过程中, 物种-多度曲线的斜率变小, 物种数由南坡的19种到北坡的26种, 呈现一个递增的趋势, 南坡物种的平均相对多度在0.014左右, 北坡物种的相对多度在0.076左右。且在南坡-北坡的变化中, 常见种明显增多, 同时优势种也发生了变化, 南坡是以三芒草(0.051)和山莴苣(0.047)为主, 东南坡以三芒草(0.271)和火绒草(0.280)为主, 东坡以矮生嵩草(0.252)和火绒草(0.083)为主, 东北坡和北坡则是以矮生嵩草(0.237)和珠芽蓼(0.161)为主。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2甘南高寒草甸各坡向物种-多度分布曲线。S, 南坡; SE, 东南坡; E, 东坡; NE, 东北坡; N, 北坡。 Fig. 2Species abundance curves in different slopes of Gannan alpine meadow. S, South; SE, Southeast; E, East; NE, Northeast; N, North. 2.4 拟合值和实测值的比较 图3 显示, 拟合值和实测值的变化趋势相似。Rapo模型在南坡、东南坡和东北坡上相对多度为0.001左右, 则说明拟合过程中Rapo所拟合的数据只有统计学意义, 在资源的随机分配上并没有实际的意义。而geo模型的拟合对5个坡向而言, 它的物种对资源的分配模式是固定的, 则对5个坡向的共有种和优势种的筛选及拟合都有实际的指导意义,也奠定了常见种和优势种在环境竞争中的地位。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3甘南高寒草甸不同坡向实测值和拟合值的对比。bro, 断棍模型; geo, 几何级数模型; Norm, 正态分布模型; Rane, 随机分类模型; Rapo, 随机分配模型; ZM, 随机ZM模型。 Fig. 3Comparison of observed and fitted values in different slopes of Gannan alpine meadow. bro, broken stick; geo, geometric series; Norm, log normal; Rane, random assortment; Rapo, random fraction; ZM, Zipf_Mandelbrot. 2.5 常见种和稀有种的拟合比较 表3 表明, geo模型的拟合效果比Rane模型更好。在南坡和东北坡, Rane模型的置信区间小于95%。东南坡、东坡、北坡的资源分配模式主要是由随机分配和固定分配两种模式共同作用的结果。
Table 3 表3 表3 甘南高寒草甸常见种和稀有种的模型拟合
Table 3
Model fitting of common and rare species of Gannan alpine meadow 坡向 物种 Species 模型 常见种 Frequent species 稀有种 Rare species r Oc CL r Oc CL 南坡 South Rane 32.55 6.94 1.00 71.91 2.10 0.65 geo 23.70 1.73 1.00 20.16 3.54 1.00 东南坡 Southeast (45°) Rane 14.41 4.93 1.00 32.86 1.53 1.00 geo 17.18 1.64 1.00 31.88 1.53 1.00 东坡 East Rane 120.26 14.80 1.00 62.66 4.52 1.00 geo 14.07 14.80 1.00 44.78 1.50 1.00 东北坡 Northeast (135°) Rane 27.65 2.57 1.00 78.33 6.88 0.66 geo 22.13 2.57 1.00 82.75 26.99 1.00 北坡 North Rane 35.55 4.46 1.00 69.83 7.52 1.00 geo 32.50 4.46 1.00 59.07 6.02 1.00
r, a correction factor of maximum density difference and species number is added in the least square method; Oc, Preston’s χ2 test after doubling grouping; CL, confidence interval. geo, geometric series; Rane, random assortment.r, 采用最小二乘法中加入密度最大差异和物种数量的校正因子; Oc, Preston倍程分组后的χ2 检验法; CL, 置信区间。geo, 几何级数模型; Rane, 随机分类模型。新窗口打开 |
下载CSV 图4A 表明, 常见种的实测值和拟合的geo模型的趋势一致。它们在生态环境中对资源的获取方式基本上是以固定分配的模式进行演变。由
图4B 可知, 稀有种在4个坡向的实测值和拟合值不同, 但趋势均是减小的, 这进一步说明稀有种在各个坡向的 资源分配模式是随机分配模式, 但geo模型并不起 主导作用, 这是随机分配和固定分配共同作用的 结果。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4甘南高寒草甸各坡向常见种、稀有种实测值和几何级数模型(geo)拟合值。 Fig. 4Each aspect of alpine meadow in Gannan common species, rare species observed value and Geometric series (geo) fitting values. 3 讨论 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 )。本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致。表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局。南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主。从南到北, 形成了一个非常明显的植被分布梯度。
在南坡-北坡变化中, 全部物种-多度分布曲线斜率依次减小(
图2 ), 但物种丰富度逐渐增加。
孙小妹等(2014) 通过添加营养元素, 研究了青藏高原高寒草甸植物物种-多度分布格局, 结果表明从物种-多度分布曲线的斜率变化可以判断物种的丰富度。本研究所涉及的优势种和稀有种, 其数量都随着坡向变化从南到北依次增多。北坡常见种的相对多度在0.076左右, 这表明在青藏高原高寒草甸区, 由于土壤含水量从南到北呈现递增的趋势, 而光照强度和土壤温度逐渐减少, 加之北坡、东北坡坡度平缓(平均20°左右), 南坡、东南坡坡度较大(平均33°左右), 这样导致南坡径流增大, 土壤水分、养分流失严重, 南坡相对比较贫瘠, 适宜生存的物种较少; 而北坡水分和养分相对较高, 其生境相对优越, 能容纳更多的物种生存, 因此北坡、东北坡物种多样性最高, 且常见种的丰富度也大。这与
刘旻霞和王刚(2013) 对高寒草甸的阴阳坡物种丰富度的研究结果是一致的, 即物种的丰富度从南坡到北坡是递增的。另外, 在养分、水分相对匮乏的南坡, 由于资源的随机性分配, 优势种的生长、盖度、多度和稀有种相比, 其在资源的获取上更有优势。
bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 )。本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 )。在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主。从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素。
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致。从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 )。北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争。
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致。本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主。相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程。在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 )。常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响。在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高。随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的。本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主。常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致。稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况。同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加。
综上所述, 在青藏高原高寒草甸区, 对物种-多度拟合并且通过对模型分析得出, 青藏高原高寒草甸区物种多度模型主要是以生态位模型为主, 其次是随机分布模型。这也使得青藏高原高寒草甸区的资源分配模式出现了确定性和随机性共存的一个过程, 常见种对维持生态系统的稳定性起着非常重要的作用, 在不同的生态环境中, 常见种在生长过程中从资源的分配获取上可以抑制优势种的快速繁殖, 为稀有种的生长争取一定的随机性分配资源。就常见种来看, 其在生长繁殖的过程中具有优先占领的特权, 但实际上物种-多度分布不止遵循一种规律。我们可以推断出常见种的资源分配模式符合资源划分和占有的过程。稀有种则可能是在迁移率、人为活动等因素共同干扰的条件下形成的。稀有种增加了各坡向的物种丰富度, 同时也是导致高寒草甸物 种-多度分布格局差异性的重要原因。就坡向的拟合来看, 生态位模型和随机模型两者共存下的拟合效果会更好, 这对群落发展和群落构建过程中资源分配的研究具有指导意义。
[1] Che YD Liu MX Li LR Jiao J Xiao W 2017 ). Exploring the community assembly of subalpine meadow communities based on functional traits and community phylogenyChinese Journal of Plant Ecology, 41, 1157 -1167 . [本文引用: 3] [ 车应弟 , 刘旻霞 , 李俐蓉 , 焦骄 , 肖卫 (2017 ). 基于功能性状及系统发育的亚高寒草甸群落构建植物生态学报 , 41, 1157 -1167 .] [本文引用: 3] [2] Duan HL Zhao A Yao Z 2017 ). Modeling plant species- abundance distribution patterns with elevation in typical marshland communities of the Poyang Lake regionPlant Science Journal, 35, 335 -343 . [ 段后浪 , 赵安 , 姚忠 (2017 ). 不同海拔高程梯度下鄱阳湖典型草洲植物群落物种-多度分布格局的模型拟合植物科学学报 , 35, 335 -343 .] [3] Fang JY Shen ZH Cui HT 2004 ). Ecological characteristics of mountains and research issues of mountain ecologyBiodiversity Science, 12, 10 -19 . [本文引用: 1] [ 方精云 , 沈泽昊 , 崔海亭 (2004 ). 试论山地的生态特征及山地生态学的研究内容生物多样性 , 12, 10 -19 .] [本文引用: 1] [4] Fisher RA Corbet AS Williams CB 1943 ). The relationship between the number of species and the number of indi- viduals in a random sample from an animal populationJournal of Animal Ecology , 12, 42 -58 . [本文引用: 2] [5] Fujita Y Robroek BJ De Ruiter PC 2010 ). Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activityOikos , 119, 1665 -1673 . [本文引用: 1] [6] Gao LX Bi RC Yan M 2011 ). Species abundance distribution patterns of Pinus tabulaeformis forest in Huoshan Mountain of Shanxi Province, ChinaChinese Journal of Plant Ecology, 1256 -1270 . [本文引用: 1] [ 高利霞 , 毕润成 , 闫明 (2011 ). 山西霍山油松林的物种-多度分布格局植物生态学报 , 35, 1256 -1270 .] [本文引用: 1] [7] Gong X Brueck K Giese KM 2008 ). Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia ChinaJournal of Arid Environments , 72, 483 -493 . [本文引用: 1] [8] Hubbell SP 2001 ) Unified Neutral Theory of Biodiversity and Biogeography . Princeton University Press, Princeton. [本文引用: 1] [9] Li R Liu MX Zhang C Zhao RD Shao P 2017 ). Distribution characteristics of soil microbial communities along different slope direction of Gannan sub-alpine meadowsEcology and Environmental Sciences, 26, 1884 -1891 . [本文引用: 2] [ 李瑞 , 刘旻霞 , 张灿 , 赵瑞东 , 邵鹏 (2017 ). 甘南亚高寒草甸不同坡向土壤微生物群落分布特征生态环境学报 , 26, 1884 -1891 .] [本文引用: 2] [10] Liu MX Che YD Li LR Jiao J Xiao W 2017a ). Redundancy analysis of leaf traits and environmental factors of alpine meadow in Southern Gansu ProvinceChinese Journal of Ecology, 36, 2473 -2480 . [ 刘旻霞 , 车应弟 , 李俐蓉 , 焦娇 , 肖卫 (2017a ). 甘南高寒草甸微地形上植物叶片特征与环境因子的冗余分析生态学杂志 , 36, 2473 -2480 .] [11] Liu MX Liu JJ Du XG Zheng XG 2010 ). Fitting different models to species abundance distribution patterns in three plant communities in sub-alpine meadowActa Ecologica Sinica, 30, 6935 -6942 . [本文引用: 1] [ 刘梦雪 , 刘佳佳 , 杜晓光 , 郑小刚 (2010 ). 亚高寒草甸不同生境植物群落物种-多度分布格局的拟合生态学报 , 30, 6935 -6942 .] [本文引用: 1] [12] Liu MX Wang G 2013 ). Responses of plant community diversity and soil factors to slope aspect in alpine meadowChinese Journal of Ecology , 32, 259 -265 . [本文引用: 1] [ 刘旻霞 , 王刚 (2013 ). 高寒草甸植物群落多样性及土壤因子对坡向的响应生态学杂志 , 32, 259 -265 .] [本文引用: 1] [13] Liu MX Wang G Sheng HM 2013 ). Environmental factors characteristic of sunny and shady slope gradients in relation to above-ground biomass and species richness in an alpine meadowJournal of Lanzhou University (Natural Sciences )76 -81 . [本文引用: 2] [ 刘旻霞 , 王刚 , 盛红梅 (2013 ). 高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系兰州大学学报(自然科学版) , 49, 76 -81 .] [本文引用: 2] [14] Liu MX Zhao RD Zhang C Li R Shao P 2017b ). Responses of physiological parameters in plants on sub-alpine meadow to slope aspectsChinese Journal of Applied Ecology, 28, 2863 -2869 . [ 刘旻霞 , 赵瑞东 , 张灿 , 李瑞 , 邵鹏 (2017b ). 亚高寒草甸植物叶片生理指标对坡向的响应应用生态学报 , 28, 2863 -2869 .] [15] Liu MX Zhu KJ 2013 ). Characteristics of nitrogen and phosphorus stoichiometry of plants in different functional groups on alpine meadow in the eastern edge of Tibetan PlateauChinese Journal of Grassland, 35(2 ), 52 -58 . [本文引用: 1] [ 刘旻霞 , 朱柯嘉 (2013 ). 青藏高原东缘高寒草甸不同功能群植物氮磷化学计量特征研究中国草地学报 , 35(2 ), 52 -58 .] [本文引用: 1] [16] Ma KM 2003 ). Advances of the study on species abundance patternActa Phytoecologica Sinica, 27, 412 -426 . [本文引用: 1] [ 马克明 (2003 ). 物种-多度格局研究进展植物生态学报 , 27, 412 -426 .] [本文引用: 1] [17] Ma KM Zu YG 2000 ). Fractal properties of vegetation patternActa Phytoecologica Sinica, 46, 111 -117 . [本文引用: 1] [ 马克明 , 祖元刚 (2000 ). 植被格局的分形特征植物生态学报 , 46, 111 -117 .] [本文引用: 1] [18] MacArthur RH 1957 ). On the relative abundance of bird speciesProceedings of the National Academy of Sciences of the United States of America , 43, 293 -295 . [本文引用: 2] [19] Magurran AE 1988 ). Ecological Diversity and Its Measurement Croom Helm, London. [本文引用: 1] [20] Motomura I 1932 ). On the statistical treatment of communitiesZool Mag , 44, 379 -383 . [本文引用: 2] [21] Ren P Wang XA Guo H 2009 ). Species abundance distribution pattern of forest communities on Loess PlateauChinese Journal of Ecology, 29, 1449 -1455 . [本文引用: 1] [ 任萍 , 王孝安 , 郭华 (2009 ). 黄土高原森林群落物种-多度的分布格局生态学杂志 , 29, 1449 -1455 .] [本文引用: 1] [22] Shi JM Fan CF Liu Y Yang QP Fang K Fan FL Yang GY 2015 ). Species-abundance distribution patterns along succession series of Phyllostachys glauca forest in a limestone mountainChinese Journal of Applied Ecology, 3595 -3601 . [本文引用: 1] [ 施建敏 , 范承芳 , 刘扬 , 杨清培 , 方楷 , 范方礼 , 杨光耀 (2015 ). 石灰岩山地淡竹林演替序列的群落物种-多度分布格局应用生态学报 , 26, 3595 -3601 .] [本文引用: 1] [23] Sun XM Xiao ML Shi RL Han F Wang G 2014 ). Effects of nutrient additions on species abundance distribution in an alpine meadow in the Qinghai-Tibetan Plateau, ChinaJournal of Lanzhou University (Natural Sciences )853 -859 . [本文引用: 4] [ 孙小妹 , 肖美玲 , 师瑞玲 , 韩非 , 王刚 (2014 ). 营养元素添加对青藏高原亚高寒草甸物种-多度分布格局的影响兰州大学学报(自然科学版) , 50, 853 -859 .] [本文引用: 4] [24] Tokeshi M 1990 ). Niche apportionment or random assortment: Species abundance patterns revisitedJournal of Animal Ecology , 59, 1129 -1146 . [本文引用: 2] [25] Tokeshi M 1993 ). Species abundance patterns and community structureAdvances in Ecological Research , 24, 112 -186 . [本文引用: 1] [26] Ulrich W 2001 ). Models of relative abundance distributions Ⅰ: Model fitting by stochastic modelsPolish Journal of Ecology , 49, 145 -147 . [本文引用: 1] [27] Ulrich W 2002 ). RAD-A FORTRAN program for the study of relative abundance distributions. . Cited: 2018-12-18. URL [本文引用: 1] [28] Wang SX Zhao L Li N Guo H Wang XA Duan RY 2016 ). The relative contributions of rare and common species to the patterns of species richness in plant communitiesBiodiversity Science, 24, 658 -664 . [本文引用: 1] [ 王世雄 , 赵亮 , 李娜 , 郭华 , 王孝安 , 段仁燕 (2016 ). 稀有种和常见种对植物群落物种丰富度格局的相对贡献生物多样性 , 24, 658 -664 .] [本文引用: 1] [29] Wang YS Shangguan TL 2010 ). Discussion on calculating method of important valuesJournal of Shanxi University, 33, 312 -316 . [ 王育松 , 上官铁梁 (2010 ). 关于重要值计算方法的若干问题山西大学学报(自然科学版) , 33, 312 -316 .] [30] Xiao ML 2014 ). Species Abundance Distribution of Zoker Mound Plant Community Along Restoration Progress and Its Relationship with Mound AreaMaster degree dissertation, Lanzhou University, Lanzhou . [本文引用: 1] [ 肖美玲 (2014 ). 鼢鼠土丘植物群落恢复演替过程中的物种-多度分布及其与鼠丘面积的关系硕士学位论文, 兰州大学, 兰州 .] [本文引用: 1] [31] Zhang JT 1999 ). Species abundance patterns in some broad- leaved deciduous forests in New York, USAActa Phytoecologica Sinica, 23, 481 -489 . [ 张金屯 (1999 ). 美国纽约州阔叶林物种-多度格局的研究植物生态学报 , 23, 481 -489 .] 基于功能性状及系统发育的亚高寒草甸群落构建
3
2017
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
基于功能性状及系统发育的亚高寒草甸群落构建
3
2017
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
不同海拔高程梯度下鄱阳湖典型草洲植物群落物种-多度分布格局的模型拟合
2017
不同海拔高程梯度下鄱阳湖典型草洲植物群落物种-多度分布格局的模型拟合
2017
试论山地的生态特征及山地生态学的研究内容
1
2004
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
试论山地的生态特征及山地生态学的研究内容
1
2004
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
The relationship between the number of species and the number of indi- viduals in a random sample from an animal population
2
1943
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
...
Fisher等(1943) 首次提出对数级数模型(Lser), 该模型属于经验模型, 用来拟合能够满足下面等式的物种-多度数据: ...
Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity
1
2010
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
山西霍山油松林的物种-多度分布格局
1
2011
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
山西霍山油松林的物种-多度分布格局
1
2011
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia China
1
2008
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
1
2001
... 常见模型有随机分配模型(Rapo)(
Hubbell, 2001 )、随机分类模型(Rane)(
Tokeshi, 1990 )、随机ZM模型(Zipf_Mandelbrot)、断棍模型(bro)(
Tokeshi, 1993 )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 ). ...
甘南亚高寒草甸不同坡向土壤微生物群落分布特征
2
2017
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
甘南亚高寒草甸不同坡向土壤微生物群落分布特征
2
2017
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... 坡向是山地的主要地形因子, 虽然各个坡向之间的距离比较短, 但环境因子(光照、土壤含水量、土壤养分)的变化是非常剧烈的(
Gong et al ., 2008 ), 同时还影响不同坡向的植被分布格局(
方精云等, 2004 ).本研究结果表明坡向从南坡向北坡的变化过程中, 土壤含水量逐渐增加, 土壤温度和光照强度则逐渐减小(
图1 ), 这与
刘旻霞等(2017a ,
2017b )的研究结果一致.表明坡向变化导致环境因子的变化, 进而影响不同坡向植被的分布格局.南坡和东南坡主要是以禾草类(三芒草、矮生嵩草)及杂草(山莴苣等)为主; 东坡以豆科(米口袋、甘肃棘豆(
Oxytropis kansuensis )等)和杂类草(火绒草等)为主, 东北坡和北坡则是以灌木(金露梅)和杂类草(珠芽蓼、中华老鹳草(
Geranium sinense )等)为主.从南到北, 形成了一个非常明显的植被分布梯度. ...
甘南高寒草甸微地形上植物叶片特征与环境因子的冗余分析
2017a
甘南高寒草甸微地形上植物叶片特征与环境因子的冗余分析
2017a
亚高寒草甸不同生境植物群落物种-多度分布格局的拟合
1
2010
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
亚高寒草甸不同生境植物群落物种-多度分布格局的拟合
1
2010
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
高寒草甸植物群落多样性及土壤因子对坡向的响应
1
2013
... 在南坡-北坡变化中, 全部物种-多度分布曲线斜率依次减小(
图2 ), 但物种丰富度逐渐增加.
孙小妹等(2014) 通过添加营养元素, 研究了青藏高原高寒草甸植物物种-多度分布格局, 结果表明从物种-多度分布曲线的斜率变化可以判断物种的丰富度.本研究所涉及的优势种和稀有种, 其数量都随着坡向变化从南到北依次增多.北坡常见种的相对多度在0.076左右, 这表明在青藏高原高寒草甸区, 由于土壤含水量从南到北呈现递增的趋势, 而光照强度和土壤温度逐渐减少, 加之北坡、东北坡坡度平缓(平均20°左右), 南坡、东南坡坡度较大(平均33°左右), 这样导致南坡径流增大, 土壤水分、养分流失严重, 南坡相对比较贫瘠, 适宜生存的物种较少; 而北坡水分和养分相对较高, 其生境相对优越, 能容纳更多的物种生存, 因此北坡、东北坡物种多样性最高, 且常见种的丰富度也大.这与
刘旻霞和王刚(2013) 对高寒草甸的阴阳坡物种丰富度的研究结果是一致的, 即物种的丰富度从南坡到北坡是递增的.另外, 在养分、水分相对匮乏的南坡, 由于资源的随机性分配, 优势种的生长、盖度、多度和稀有种相比, 其在资源的获取上更有优势. ...
高寒草甸植物群落多样性及土壤因子对坡向的响应
1
2013
... 在南坡-北坡变化中, 全部物种-多度分布曲线斜率依次减小(
图2 ), 但物种丰富度逐渐增加.
孙小妹等(2014) 通过添加营养元素, 研究了青藏高原高寒草甸植物物种-多度分布格局, 结果表明从物种-多度分布曲线的斜率变化可以判断物种的丰富度.本研究所涉及的优势种和稀有种, 其数量都随着坡向变化从南到北依次增多.北坡常见种的相对多度在0.076左右, 这表明在青藏高原高寒草甸区, 由于土壤含水量从南到北呈现递增的趋势, 而光照强度和土壤温度逐渐减少, 加之北坡、东北坡坡度平缓(平均20°左右), 南坡、东南坡坡度较大(平均33°左右), 这样导致南坡径流增大, 土壤水分、养分流失严重, 南坡相对比较贫瘠, 适宜生存的物种较少; 而北坡水分和养分相对较高, 其生境相对优越, 能容纳更多的物种生存, 因此北坡、东北坡物种多样性最高, 且常见种的丰富度也大.这与
刘旻霞和王刚(2013) 对高寒草甸的阴阳坡物种丰富度的研究结果是一致的, 即物种的丰富度从南坡到北坡是递增的.另外, 在养分、水分相对匮乏的南坡, 由于资源的随机性分配, 优势种的生长、盖度、多度和稀有种相比, 其在资源的获取上更有优势. ...
高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系
2
2013
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系
2
2013
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
亚高寒草甸植物叶片生理指标对坡向的响应
2017b
亚高寒草甸植物叶片生理指标对坡向的响应
2017b
青藏高原东缘高寒草甸不同功能群植物氮磷化学计量特征研究
1
2013
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
青藏高原东缘高寒草甸不同功能群植物氮磷化学计量特征研究
1
2013
... 青藏高原甘南高寒草甸位于青藏高原的东部, 生物资源和物种多样性相对丰富, 也是当地的牧草主产区(
车应弟等, 2017 ).由于自然环境的严酷和人类活动太过频繁, 高寒草甸草场不断退化, 生物多样性锐减.因此保护青藏高原高寒草甸区的生态系统刻不容缓.坡向是高寒草甸的主要地形因子, 随着坡向的变化, 光照强度、土壤含水量(
刘旻霞和王刚, 2013 )、土壤养分(
刘旻霞和朱柯嘉, 2013 )、土壤微生物(
李瑞等, 2017 )以及土壤温度等都会发生明显变化.因此基于坡向梯度研究物种-多度对认识高寒草甸区物种群落的构建和物种演替过程中的资源分配模式有重要的意义. ...
物种-多度格局研究进展
1
2003
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
物种-多度格局研究进展
1
2003
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
植被格局的分形特征
1
2000
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
植被格局的分形特征
1
2000
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
On the relative abundance of bird species
2
1957
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
...
MacArthur (1957) 提出了最为经典的断棍模型(bro).模型假设的是一个群落中所有的资源在一条棍子上, 在棍上随机选取(
s - 1)个点, 把棍分成
s 份, 则代表有
s 个物种占有该群落的所有资源, 每个物种的竞争力是相似的.以
J/S (
J 表示该群落中所有物种-多度之和,
S 是指群落中物种总和)表示该群落里平均相对多度, 即该模型的第
i 个种的多度
Ai 可以表示为: ...
Ecological Diversity and Its Measurement 1
1988
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
On the statistical treatment of communities
2
1932
... 常见模型有随机分配模型(Rapo)(
Hubbell, 2001 )、随机分类模型(Rane)(
Tokeshi, 1990 )、随机ZM模型(Zipf_Mandelbrot)、断棍模型(bro)(
Tokeshi, 1993 )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 ). ...
...
Motomura (1932) 在研究植物群落演替的过程中提出了生态位优先占领模型(geo), 从而在几何级数模型的基础上进一步完善该模型.几何级数模型是按物种-多度值从小到大的顺序进行排列, 计算公式如下式: ...
黄土高原森林群落物种-多度的分布格局
1
2009
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
黄土高原森林群落物种-多度的分布格局
1
2009
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
石灰岩山地淡竹林演替序列的群落物种-多度分布格局
1
2015
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
石灰岩山地淡竹林演替序列的群落物种-多度分布格局
1
2015
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
营养元素添加对青藏高原亚高寒草甸物种-多度分布格局的影响
4
2014
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
... 采用RAD拟合程序包, 以fit411.txt文件的处理方法进行拟合.RAD是一款独立多批次处理数据的编程软件, 通过前期固定编程, 在读取数据(多度数据应按txt文本进行整理)时根据不同命令进行读取, 再根据软件自带参数拟合包进行数据批量处理, 最后输出一个名为Assemblage.txt的文本, 该文本里包含了
r 值(
r 是用模型的
n 个副本的平均密度进行计算, 采用最小二乘法中加入密度最大差异和物种数量的校正因子, 从而增强了模型拟合优度的区分能力)、
Oc 值(经过Ulrich校正过的Preston倍程分组后的χ
2 检验法)、
CL (置信区间)、相对多度、最大密度差、相对丰富度的标准差、多样性指数、均匀度指数等, 最后通过Excel进行分栏并根据相关参数(
r 值、
Oc 值、
CL )进行模型的选取.fit411.txt是指RAD软件在后期读取数的一个匹配文本, 里面的相关参数与相应的模型相互匹配, 这些参数包含了物种数、流量数、所匹配的模型名称、软件运行的迭代器、迭代次数、密度、置信区间及校正模式等, 而所有参数值都采用程序包中的默认值.在优势度的检验上, 采取
r 值、
Oc 值、
CL 值3个指标.通过多次迭代最后得到使
r 值最小的理论密度值即为所求值.因为
r 值受物种最大密度差和群落物种数目影响.所以RAD对物种数目
S 和最大密度差
D 进行了修正.
r 小于或接近10说明拟合效果较好(算法在倍频次运算时会无限接近于10), 大于100表明拟合效果较差(
Ulrich, 2002 ).RAD同样输出Preston倍程分组后的拟合变量
Oc 值, 该值的计算方法与
r 值相似, 将同一多度物种的理论与实测密度之间的欧氏距离换成同一倍程组别的.同样当
Oc 值小于或接近10 (算法在倍频次运算时会无限接近于10)的时候拟合效果较好而当其大于100的时候拟合效果较差.同时RAD还给出在
x %置信区间内的物种频率(
CL , 一般默认为是大于或等于95%的置信区间), 尽管该拟合优度检验方法不如
r 值、
Oc 值灵敏, 但对于随机模型的拟合优度检验非常重要(
肖美玲, 2014 ). ...
... 在南坡-北坡变化中, 全部物种-多度分布曲线斜率依次减小(
图2 ), 但物种丰富度逐渐增加.
孙小妹等(2014) 通过添加营养元素, 研究了青藏高原高寒草甸植物物种-多度分布格局, 结果表明从物种-多度分布曲线的斜率变化可以判断物种的丰富度.本研究所涉及的优势种和稀有种, 其数量都随着坡向变化从南到北依次增多.北坡常见种的相对多度在0.076左右, 这表明在青藏高原高寒草甸区, 由于土壤含水量从南到北呈现递增的趋势, 而光照强度和土壤温度逐渐减少, 加之北坡、东北坡坡度平缓(平均20°左右), 南坡、东南坡坡度较大(平均33°左右), 这样导致南坡径流增大, 土壤水分、养分流失严重, 南坡相对比较贫瘠, 适宜生存的物种较少; 而北坡水分和养分相对较高, 其生境相对优越, 能容纳更多的物种生存, 因此北坡、东北坡物种多样性最高, 且常见种的丰富度也大.这与
刘旻霞和王刚(2013) 对高寒草甸的阴阳坡物种丰富度的研究结果是一致的, 即物种的丰富度从南坡到北坡是递增的.另外, 在养分、水分相对匮乏的南坡, 由于资源的随机性分配, 优势种的生长、盖度、多度和稀有种相比, 其在资源的获取上更有优势. ...
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
营养元素添加对青藏高原亚高寒草甸物种-多度分布格局的影响
4
2014
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
... 采用RAD拟合程序包, 以fit411.txt文件的处理方法进行拟合.RAD是一款独立多批次处理数据的编程软件, 通过前期固定编程, 在读取数据(多度数据应按txt文本进行整理)时根据不同命令进行读取, 再根据软件自带参数拟合包进行数据批量处理, 最后输出一个名为Assemblage.txt的文本, 该文本里包含了
r 值(
r 是用模型的
n 个副本的平均密度进行计算, 采用最小二乘法中加入密度最大差异和物种数量的校正因子, 从而增强了模型拟合优度的区分能力)、
Oc 值(经过Ulrich校正过的Preston倍程分组后的χ
2 检验法)、
CL (置信区间)、相对多度、最大密度差、相对丰富度的标准差、多样性指数、均匀度指数等, 最后通过Excel进行分栏并根据相关参数(
r 值、
Oc 值、
CL )进行模型的选取.fit411.txt是指RAD软件在后期读取数的一个匹配文本, 里面的相关参数与相应的模型相互匹配, 这些参数包含了物种数、流量数、所匹配的模型名称、软件运行的迭代器、迭代次数、密度、置信区间及校正模式等, 而所有参数值都采用程序包中的默认值.在优势度的检验上, 采取
r 值、
Oc 值、
CL 值3个指标.通过多次迭代最后得到使
r 值最小的理论密度值即为所求值.因为
r 值受物种最大密度差和群落物种数目影响.所以RAD对物种数目
S 和最大密度差
D 进行了修正.
r 小于或接近10说明拟合效果较好(算法在倍频次运算时会无限接近于10), 大于100表明拟合效果较差(
Ulrich, 2002 ).RAD同样输出Preston倍程分组后的拟合变量
Oc 值, 该值的计算方法与
r 值相似, 将同一多度物种的理论与实测密度之间的欧氏距离换成同一倍程组别的.同样当
Oc 值小于或接近10 (算法在倍频次运算时会无限接近于10)的时候拟合效果较好而当其大于100的时候拟合效果较差.同时RAD还给出在
x %置信区间内的物种频率(
CL , 一般默认为是大于或等于95%的置信区间), 尽管该拟合优度检验方法不如
r 值、
Oc 值灵敏, 但对于随机模型的拟合优度检验非常重要(
肖美玲, 2014 ). ...
... 在南坡-北坡变化中, 全部物种-多度分布曲线斜率依次减小(
图2 ), 但物种丰富度逐渐增加.
孙小妹等(2014) 通过添加营养元素, 研究了青藏高原高寒草甸植物物种-多度分布格局, 结果表明从物种-多度分布曲线的斜率变化可以判断物种的丰富度.本研究所涉及的优势种和稀有种, 其数量都随着坡向变化从南到北依次增多.北坡常见种的相对多度在0.076左右, 这表明在青藏高原高寒草甸区, 由于土壤含水量从南到北呈现递增的趋势, 而光照强度和土壤温度逐渐减少, 加之北坡、东北坡坡度平缓(平均20°左右), 南坡、东南坡坡度较大(平均33°左右), 这样导致南坡径流增大, 土壤水分、养分流失严重, 南坡相对比较贫瘠, 适宜生存的物种较少; 而北坡水分和养分相对较高, 其生境相对优越, 能容纳更多的物种生存, 因此北坡、东北坡物种多样性最高, 且常见种的丰富度也大.这与
刘旻霞和王刚(2013) 对高寒草甸的阴阳坡物种丰富度的研究结果是一致的, 即物种的丰富度从南坡到北坡是递增的.另外, 在养分、水分相对匮乏的南坡, 由于资源的随机性分配, 优势种的生长、盖度、多度和稀有种相比, 其在资源的获取上更有优势. ...
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
Niche apportionment or random assortment: Species abundance patterns revisited
2
1990
... 常见模型有随机分配模型(Rapo)(
Hubbell, 2001 )、随机分类模型(Rane)(
Tokeshi, 1990 )、随机ZM模型(Zipf_Mandelbrot)、断棍模型(bro)(
Tokeshi, 1993 )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 ). ...
... )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 ). ...
Species abundance patterns and community structure
1
1993
... 常见模型有随机分配模型(Rapo)(
Hubbell, 2001 )、随机分类模型(Rane)(
Tokeshi, 1990 )、随机ZM模型(Zipf_Mandelbrot)、断棍模型(bro)(
Tokeshi, 1993 )、正态分布模型(Norm)(
Tokeshi, 1990 )和几何级数模型(geo)(
Motomura, 1932 ). ...
Models of relative abundance distributions Ⅰ: Model fitting by stochastic models
1
2001
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
1
2002
... bro模型和geo模型代表的是在群落发展过程中它的资源分配具有确定性, 而Rapo模型和Norm模型是假定群落中的资源分配是随机的(
Ulrich, 2001 ).本研究结果表明, 在高寒草甸区各个坡向上所有物种均满足统计模型中的Norm模型, 以及生态位模型中的geo模型、bro模型、ZM模型(
图3 ).在拟合过程中南坡的资源分配模式是以随机分配为主; 北坡的资源分配模式是以确定性分配为主.从模型的拟合角度来分析, 影响青藏高原高寒草甸区的物种-多度分布模型主要是以生态位拟合模型为主, 它也是导致物种资源分配格局趋于确定性的主要因素.
Magurran (1988) 研究发现物种群落的资源分配问题可以通过对常见种和稀有种的研究来确定, 并提出随机分配模型和固定分配模型在资源分配模式上是一体存在、共同作用的结果, 我们的研究结果与之一致.从南坡向北坡的生境变化过程中, 生长资源的竞争是从水资源到光资源的转变(
刘梦雪等, 2010 ).北坡优势种金露梅的植株较高, 对其他草本植物而言, 植株较高的植物侧枝密度大, 这会严重影响这些草本植物的采光, 进而资源竞争会最终转化为光竞争.
刘旻霞等(2013) 对高寒草甸植物功能性状的研究也表明, 随坡向变化, 生境梯度上的水分和养分变化均会导致植株性状发生改变, 本研究结果与之一致.本研究发现, 在坡向发生变化的同时, 各个坡向拟合效果最好的是geo模型, 植被在资源的获取模式上是以固定分配模式为主.相关研究发现生态位模型拟合对森林系统中的草本群落(
Fujita et al ., 2010 )和森林生态系统(
高利霞等, 2011 )的拟合都比较理想, 表明生态位模型的拟合能很好地阐述生物群落的构建过程.在物种丰富度的研究上, 常见种和稀有种有重要的意义(
王世雄等, 2016 ).常见种在物种丰富度和群落构建上的主导作用并不受稀有种和常见种的相对比例的影响.在青藏高原高寒草甸区, 一些常见种随坡向从南坡到北坡的变化过程中演变为共有种, 如矮生嵩草的相对多度在各个坡向都比较高.随着南坡-北坡的变化, 火绒草的相对多度也在减少, 反映出火绒草在生态环境中得到的资源分配也在减少; 珠芽蓼的相对多度随着南坡-北坡的变化是递增的, 这也体现出随坡向的不同, 物种的资源分配模式会从随机分配转变到固定分配, 并不是一成不变的, 同时, 不同坡向的资源分配类型也是有区别的.本研究中, 从南坡到北坡, 稀有种均能占有相对较大的比例(
图4 ), 有时在不同坡向也会同时出现, 如莓叶委陵菜的相对多度在各个坡向均较低, 但几乎每个坡都会出现, 这说明莓叶委陵菜在生存演替过程中, 在不同生境下获取资源的模式还是以随机分配模式为主.常见种的资源分配模式以固定分配模式为主, 这与在高寒草甸营养元素添加条件下的稀有种和常见种的资源分配模式上的研究结果(
孙小妹等, 2014 )一致.稀有种和常见种的生长均有不同的策略, 南坡的物种由于生境恶劣, 而导致生态位分化程度增大, 以此来获得更多的资源, 北坡物种则是通过减小生态位分化程度来适应多物种相互竞争下资源比例的划分情况.同时, 在物种多样性较高的生态环境中, 物种之间的生态位重叠更加明显, 稀有种较高的迁移率抑制了物种多样性的增加. ...
稀有种和常见种对植物群落物种丰富度格局的相对贡献
1
2016
... 本文通过对不同坡向重要值的计算来确定常见种和稀有种.通过对重要值(
M )的计算和实际试验样地的综合考虑, 将重要值大于0.01的物种作为常见种进行研究, 小于0.01的物种则作为稀有种进行研究(
王育松和上官铁梁, 2010 ). ...
稀有种和常见种对植物群落物种丰富度格局的相对贡献
1
2016
... 本文通过对不同坡向重要值的计算来确定常见种和稀有种.通过对重要值(
M )的计算和实际试验样地的综合考虑, 将重要值大于0.01的物种作为常见种进行研究, 小于0.01的物种则作为稀有种进行研究(
王育松和上官铁梁, 2010 ). ...
关于重要值计算方法的若干问题
2010
关于重要值计算方法的若干问题
2010
鼢鼠土丘植物群落恢复演替过程中的物种-多度分布及其与鼠丘面积的关系
1
2014
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
鼢鼠土丘植物群落恢复演替过程中的物种-多度分布及其与鼠丘面积的关系
1
2014
... 物种-多度是指在一定区域内不同群落中不同物种的分布数量, 是描述植物群落生物多样性的一个指标, 反映的是该物种占有资源的能力.同时也体现不同物种之间相互作用及相互影响的结果, 反映的是物种间的相互作用机制(
施建敏等, 2015 ).物种-多度模式是指群落中物种-多度组成的比例关系.将群落中分别有1、2……
n 个个体的物种依次列出, 排成了群落的多度谱, 以个体数目为横轴, 物种数目为纵轴作图即可得到基本的物种-多度分布图(
肖美玲, 2014 ).物种-多度分布曲线能显示出常见种和稀有种的比例, 因此也可以用作判断濒危物种的依据.分布模型揭示了群落的区域分布特征, 同时也为物种保护等级的确定提供了相关依据.一般情况下, 群落区域内大多数物种-多度都是较低的, 只有少量的常见种的物种-多度偏高.在物种-多度的研究上经常使用数学和生态学相结合的方法, 从而建立一定的数学模型来拟合多度格局的时空变化, 同时多度格局的时空变化是生态学研究的重要内容, 在物种-多度模式的发展过程中, 有大量的模型被提出(
马克明, 2003 ).最初用来研究物种-多度分布格局采用的模型为几何级数模型, 也被称为生态位优先占领模型.
Fisher等(1943) 最先采用对数级数模型来描述了物种组成数目较多的群落.模型拟合类型按照生态学意义可以大致分为三大类(
马克明和祖元刚, 2000 ), 统计学模型、生态位模型和中性模型.这三类模型中, 有****曾对其中生态学意义不明显的统计学模型提出质疑, 认为统计学模型在对实际的样方数据解释上存在着很大的风险(
MacArthur, 1957 ).就目前研究而言, 基于生态位理论提出的一些数学模型, 如断棍模型、重叠生态位模型和生态位单元模型对多度的格局描述比较全面(
张金屯, 1999 ).生态位模型能够比较明确地对物种-多度的生态过程有一个全面的描述.其中在物种-多度的研究上, 对生态位模型的研究设计最为广泛, 并以解释生态学宏观格局为主.
段后浪等(2017) 对湖泊草洲植物的研究主要采用断棍模型和生态位优先模型, 研究表明灰化薹草(
Carex cinerascens )处于演替中后期, 盖度较大, 其大量的地下根茎为种群的繁殖和资源的获取提供了有利条件;
任萍等(2009) 利用生态位模型对不同演替阶段的群落物种-多度格局进行了研究, 结果表明, 断棍模型和生态位优先占领模型在演替中前期拟合效果较好, 生态位重叠模型和生态位优先占领模型在演替后期有不错的拟合效果.目前对物种-多度格局的研究, 主要针对动物群落和森林生态系统, 对草地物种-多度分布格局的研究相对较少. ...
美国纽约州阔叶林物种-多度格局的研究
1999
美国纽约州阔叶林物种-多度格局的研究
1999
 ,*, 夏素娟, 南笑宁, 蒋晓轩西北师范大学地理与环境科学学院, 兰州 730070
,*, 夏素娟, 南笑宁, 蒋晓轩西北师范大学地理与环境科学学院, 兰州 730070
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT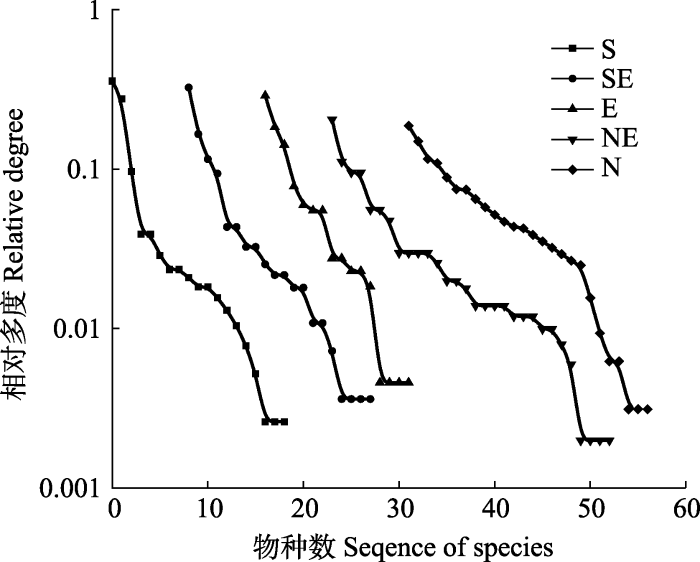 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT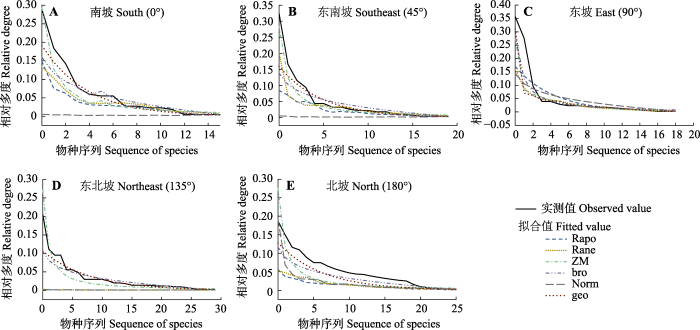 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT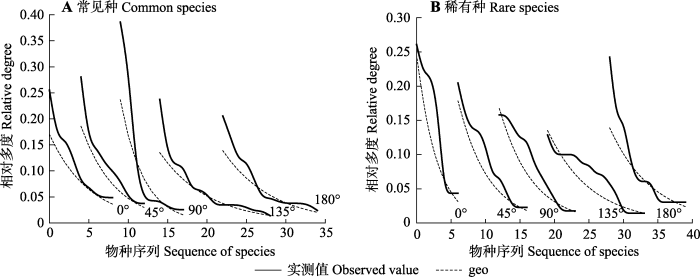 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}