Abstract AimsEpichlo? infection can affect the intraspecific and interspecific competitive ability of the host plants. However, few studies have been reported on the effects of endophyte infection on the diversity and productivity of host communities. Methods We used endophyte-infected (E+) and uninfected (E-) Achnatherum sibiricum, plus five other common species in its native community to construct a microcosm plant community to investigate the community-level consequences of endophyte interactions under different nutrient and mowing treatments. Important findings Community-level consequences of endophyte infection depend on the mowing treatment. Endophyte infection did not alter plant community diversity under non-mowing conditions. Under mowing conditions, however, endophyte infection significantly increased the diversity by increasing the abundance of lower abundance species such as Agropyron cristatum and Stipa grandis, while decreasing the abundance of dominant species such as Leymus chinensis. No significant endophyte effect on the productivity of plant community was found. With respect to nutrient treatments, we found that nutrient addition had no significant effect either on the diversity or productivity of the plant community. The positive effect of endophyte on A. sibiricum occurred only under nutrient addition treatment in the host species level. Keywords:fungal endophyte;Achnatherum sibiricum;community diversity;productivity;abundance
PDF (1171KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 吴曼, 李娟娟, 刘金铭, 任安芝, 高玉葆. 刈割干扰和养分添加条件下Epichlo?内生真菌感染对羽茅所在群落多样性和生产力的影响. 植物生态学报, 2019, 43(2): 85-93. DOI: 10.17521/cjpe.2018.0242 WU Man, LI Juan-Juan, LIU Jin-Ming, REN An-Zhi, GAO Yu-Bao. Effects of Epichlo? infection on the diversity and productivity of Achnatherum sibiricum community under various nutrient and mowing conditions. Chinese Journal of Plant Ecology, 2019, 43(2): 85-93. DOI: 10.17521/cjpe.2018.0242
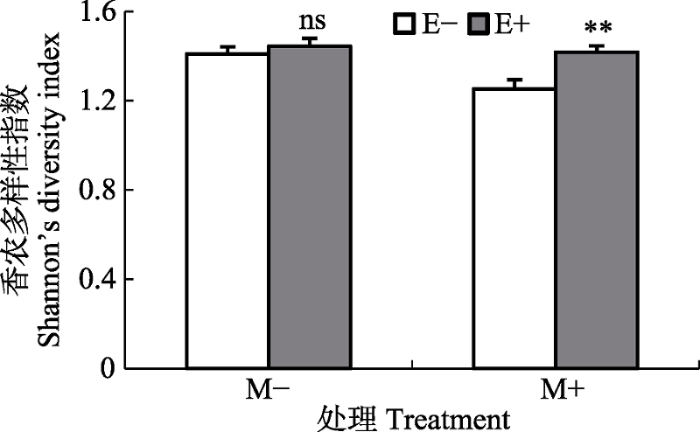
新窗口打开|下载原图ZIP|生成PPT 图1在无刈割处理(M-)和刈割处理(M+)条件下内生真菌感染对宿主群落多样性的影响(平均值+标准误差, n = 5)。E+, 内生真菌感染; E-, 内生真菌不感染。**, p < 0.01; ns, p > 0.05。
Fig. 1Effects of endophyte infection on Shannon’s diversity index of host plant communities under no-mowing (M-) and mowing (M+) conditions (mean + SE, n = 5). E+, endophyte- infected; E-, endophyte-uninfected. **, p < 0.01; ns, p > 0.05.
Fig. 2Effects of endophyte infection and nitrogen addition on the aboveground shoot biomass of host plant communities (mean + SE, n = 5). E+, endophyte-infected; E-, endophyte- uninfected; N-, no-nitrogen addition; N+, nitrogen addition. Different lowercase letters mean significant difference between E+ and E- plants community (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图3在无刈割处理(M-)和刈割处理(M+)条件下内生真菌感染对物种多度的影响(平均值+标准误差, n = 5)。E+, 内生真菌感染; E-, 无内生真菌感染。AC, 冰草; AF, 冷蒿; AS, 羽茅; CS, 糙隐子草; LC, 羊草; SG, 大针茅。星号表示E+、E-植物群落间差异显著, *, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 3Effects of endophyte infection on the species abundance under no-mowing (M-) and mowing (M+) conditions (mean + SE, n = 5). E+, endophyte-infected; E-, endophyte- uninfected. AC, Agropyron cristatum; AF, Artemisia frigida; AS, Achnatherum sibiricum; CS, Cleistogenes squarrosa; LC, Leymus chinensis; SG, Stipa grandis. The asterisk denotes significant difference between E+ and E- plants, *, p < 0.05; **, p < 0.01; ***, p < 0.001.
相比于养分添加, 物种多度受内生真菌感染的影响较大, 无刈割处理时, 内生真菌感染显著降低了羊草的多度(p = 0.028)。刈割处理后, 内生真菌不仅显著降低了羊草的多度(p = 0.007), 而且显著增加了群落中羽茅、冰草和大针茅的多度(p ? 0.001, p = 0.002, p ? 0.001)。无论是否刈割, 内生真菌对C4植物糙隐子草和非固氮植物冷蒿的多度均无显著影响(图3; 附录III)。
新窗口打开|下载原图ZIP|生成PPT 图5磷添加对土壤(A)和优势种(B-D)磷浓度的影响。B, 羽茅。C, 羊草。D, 冷蒿(平均值+标准误差, n = 5)。E-, 无内生真菌感染; E+, 内生真菌感染; P-, 无磷添加; P+, 磷添加。***, p < 0.001。
Fig. 5Effects of phosphorus addition on P concentration of the soil (A) and dominant plant species (B-D). B, Achnatherum sibiricum. C, Leymus chinensis. D, Artemisia frigida (mean + SE, n = 5). E-, endophyte-uninfected; E+, endophyte-infected; P-, no-phosphorus addition; P+, phosphorus addition. ***, p < 0.001.
Supplement Ⅰ Three-way ANOVA of the effects of endophyte infection, nitrogen and phosphorus addition on community diversity under different mowing levels
Supplement V Multiple ANOVA of effects of endophyte infection, nutrients addition and mowing on P concentration of the soil, Achnatherum sibiricum, Leymus chinensis and Artemisia frigida
AfkhamiME, StraussSY ( 2016). Native fungal endophytes suppress an exotic dominant and increase plant diversity over small and large spatial scales Ecology, 97, 1159-1169. DOI:10.1890/15-1166.1URL [本文引用: 8]
ArachevaletaM, BaconCW, HovelandCS, RadcliffeDE ( 1989). Effect of the tall fescue endophyte on plant response to environmental stress Agronomy Journal, 81, 83-90. DOI:10.2134/agronj1989.00021962008100010015xURL [本文引用: 1]
BisselsS, H?lzelN, OtteA ( 2004). Population structure of the threatened perennial Serratula tinctoria in relation to vegetation and management Applied Vegetation Science, 7, 267-274. [本文引用: 1]
BremD, LeuchtmannA ( 2002). Intraspecific competition of endophyte infected vs uninfected plants of two woodland grass species Oikos, 96, 281-290. DOI:10.1034/j.1600-0706.2002.960210.xURL [本文引用: 1]
CaoJ, HoldenNM, LüXT, DuG ( 2011). The effect of grazing management on plant species richness on the Qinghai-?Tibetan Plateau Grass and Forage Science, 66, 333-336. DOI:10.1111/gfs.2011.66.issue-3URL [本文引用: 1]
CheplickGP, ClayK, MarksS ( 1989). Interactions between infection by endophytic fungi and nutrient limitation in the grasses Lolium perenne and Festuca arundinacea New Phytologist, 111, 89-97. [本文引用: 3]
ChristensenMJ ( 1996). Antifungal activity in grasses infected with Acremonium and Epichlo? endophytes Australasian Plant Pathology, 25, 186-191. [本文引用: 1]
ClayK ( 1987). Effects of fungal endophytes on the seed and seedling biology of Lolium perenne and Festuca arundinacea Oecologia, 73, 358-362. DOI:10.1007/BF00385251URL [本文引用: 1]
JonesTA, GardnerDR, ChattertonNJ ( 2000). Cattle prefer endophyte-free robust needle grass Journal of Range Management, 53, 427-431. DOI:10.2307/4003755URL [本文引用: 1]
KnappAK, HooverDL, BlairJM, BuisG, BurkepileDE, ChamberlainA, CollinsSL, FynnRWS, KirkmanKP, SmithMD, BlakeD, GovenderN, O’NealP, SchreckT, ZinnA ( 2012). A test of two mechanisms proposed to optimize grassland aboveground primary productivity in response to grazing Journal of Plant Ecology, 5, 357-365. DOI:10.1093/jpe/rts020URL [本文引用: 1]
LanZC, BaiYF ( 2012). Testing mechanisms of N-enrichment- induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long- term ecosystem responses Philosophical Transactions of the Royal Society B, 367, 3125-3134. DOI:10.1098/rstb.2011.0352URL [本文引用: 1]
LatchGCM, ChristensenM ( 1985). Artificial infection of grasses with endophytes Annals of Applied Biology, 107, 17-24. DOI:10.1111/aab.1985.107.issue-1URL
LeuchtmannA, BaconCW, SchardlCL ( 2014). Nomenclatural realignment of Neotyphodium species with genus Epichlo? Mycologia, 106, 202-215. [本文引用: 1]
LewisGC ( 2004). Effects of biotic and abiotic stress on the growth of three genotypes of Lolium perenne with and without infection by the fungal endophyte Neotyphodium lolii Journal of Chemical Ecology, 144, 53-63. [本文引用: 2]
LewisGC, BakkenAK, MacDuffJH, RaistrickN ( 1996). Effect of infection by the endophytic fungus Acremonium lolii on growth and nitrogen uptake by perennial ryegrass(Lolium perenne) in flowing solution culture Annals of Applied Biology, 129, 451-460. [本文引用: 1]
LiCJ, NanZB, PaulVH, DapprichPD, LiuY ( 2004). A new Neotyphodium species symbiotic with drunken horse grass(Achnatherum inebrians) in China Mycotaxon, 90, 141-147. [本文引用: 1]
LiX, HanR, RenAZ, GaoYB ( 2010). Using high-temperature treatment to construct endophyte-free Achnatherum sibiricum. Microbiology China, 37, 1395-1400.
LiXS, ZhangXZ, GuX, ZhengY ( 1998). The relationship between the toxic metabolin and the endophytic fungi of drunken horse grass (Achnatherum inebrians). Grass- Feeding Livestock, 124, 44-46. [本文引用: 1]
LouisMS, FaethSH ( 1997). The effect of endophytic fungi on the fitness of Arizona fescue (Festuca arizonica) under varying nitrogen levels Bulletin of the Ecological Society of America, 78, 317-327. [本文引用: 1]
MalinowskiDP, AlloushGA, BeleskyDP ( 2000). Leaf endophyte Neotyphodium coenophialum modifies mineral uptake in tall fescue Plant and Soil, 227, 115-126.
MalinowskiDP, BeleskyDP ( 2006). Ecological importance of Neotyphodium spp. grass endophytes in agroecosystems Grassland Science, 52, 1-14. [本文引用: 1]
MalinowskiDP, BeleskyDP, HillNS, BaligarVC, FeddersJM ( 1998). Influence of phosphorus on the growth and ergot alkaloid content of Neotyphodium coenophialum-infected tall fescue(Festuca arundinacea Schreb.) Plant and Soil, 198, 53-61. [本文引用: 1]
MalinowskiDP, BrauerDK, BeleskyDP ( 1999). The endophyte Neotyphodium coenophialum affects root morphology of tall fescue grown under phosphorus deficiency Journal of Agronomy and Crop Science, 183, 53-60. [本文引用: 3]
MalinowskiDP, LeuchtmannA, SchmidtD, NosbergerJ ( 1997). Symbiosis with Neotyphodium uncinatum endophyte may increase the competitive ability of meadow fescue Agronomy Journal, 89, 833-839. [本文引用: 1]
MarksS, ClayK ( 2007). Low resource availability differentially affects the growth of host grasses infected by fungal endophytes International Journal of Plant Sciences, 168, 1269-1277. DOI:10.1086/521834URL [本文引用: 2]
MarksS, ClayK, CheplickGP ( 1991). Effects of fungal endophytes on interspecific and intraspecific competition in the grasses Festuca arundinacea and Lolium perenne Journal of Applied Ecology, 28, 194-204. [本文引用: 3]
MoraesPVD, WittWW, PhillipsTD, RossiP, PanozzoLE ( 2014). Relative competitiveness of Nimblewill ( Muhlenbergia schreberi) with tall fescues and kentucky bluegrass American Journal of Plant Sciences, 5, 3777-3787. [本文引用: 1]
NewmanJA, AbnerML, DadoRG, GibsonDJ, BrookingsA, ParsonAJ ( 2003). Effects of elevated CO2, nitrogen and fungal endophyte-infection on tall fescue: Growth, photosynthesis, chemical composition and digestibility Global Change Biology, 9, 425-437. DOI:10.1046/j.1365-2486.2003.00601.xURL [本文引用: 1]
OberhoferM, GusewellS, LeuchtmannA ( 2014). Effects of natural hybrid and non-hybrid Epichlo? endophytes on the response of Hordelymus europaeus to drought stress New Phytologist, 201, 242-253. [本文引用: 1]
QinJH, GaoY, LiuH, ZhouY, RenAZ, GaoYB ( 2016). Effect of Endophyte infection and clipping treatment on resistance and tolerance of Achnatherum sibiricum Frontiers in Microbiology, 7, 1988. DOI: 10.3389/fmicb.2016.01988. [本文引用: 2]
RavelC, CourtyC, CoudretA, CharmetG ( 1997). Beneficial effects of Neotyphodium lolii on the growth and the water status in perennial ryegrass cultivated under nitrogen deficiency or drought stress Agronomie, 17, 173-181. [本文引用: 1]
RenAZ, GaoYB, WangW, WangJL, ZhaoNX ( 2009). Influence of nitrogen fertilizer and endophyte infection on ecophysiological parameters and mineral element content of perennial ryegrass Journal of Integrative Plant Biology, 51, 75-83. DOI:10.1111/jipb.2008.51.issue-1URL [本文引用: 2]
RenAZ, GaoYB, ZhouF ( 2007). Response of Neotyphodium lolii-infected perennial ryegrass to phosphorus deficiency Plant Soil and Environment, 53, 113-119. [本文引用: 1]
RudgersJA, OrrS ( 2009). Non-native grass alters growth of native tree species via leaf and soil microbes Journal of Ecology, 97, 247-255. DOI:10.1111/jec.2009.97.issue-2URL [本文引用: 2]
SaikkonenK, LehtonenP, HelanderM, KorichevaJ, FaethSH ( 2006). Model systems in ecology: Dissecting the endophyte?-???grass literature Trends in Plant Science, 11, 428-433. DOI:10.1016/j.tplants.2006.07.001URL [本文引用: 1]
SchulthessFM, FaethSH ( 1998). Distribution, abundances, and associations of the endophytic fungal community of Arizona fescue (Festuca arizonica) Mycologia, 90, 569-578. [本文引用: 1]
SuJQ, LiXR, LiXJ, LiF ( 2013). Effects of additional N on herbaceous species of desertified steppe in arid regions of China: A four-year field study Ecological Research, 28, 21-28. DOI:10.1007/s11284-012-0994-9URL [本文引用: 1]
SullivanTJ, RodstromJ, VandopJ, LibrizziJ, GrahamC, SchardlCL, BultmanTL ( 2007). Symbiont-mediated changes in Lolium arundinaceum inducible defenses: Evidence from changes in gene expression and leaf composition New Phytologist, 176, 673-679. [本文引用: 1]
SutherlandBL, HumeDE, TapperBA ( 1999). Allelopathic effects of endophyte-infected perennial ryegrass extracts on white clover seedlings New Zealand Journal of Agricultural Research, 42, 19-26. DOI:10.1080/00288233.1999.9513349URL [本文引用: 1]
Vázquez-de-AldanaBR, RomoM, García-CiudadA, PetiscoC, García-CriadoB ( 2011). Infection with the fungal endophyte Epichlo? festucae may alter the allelopathic potential of red fescue Annals of Applied Biology, 159, 281-290. [本文引用: 1]
Vázquez-de-AldanaBR, ZabalgogeazcoaI, García-CiudadA, García- CriadoB ( 2013). An Epichlo? endophyte affects the competitive ability of Festuca rubra against other grassland species Plant & Soil, 362, 201-213. [本文引用: 1]
WeiYK, GaoYB, XuH, SuD, ZhangX, WangYH, LinF, ChenL, NieLY, RenAZ ( 2006). Occurrence of endophytes in grasses native to northern China Grass and Forage Science, 61, 422-429. DOI:10.1111/gfs.2006.61.issue-4URL [本文引用: 1]
WhiteJF, ReddyPV ( 1998). Examination of structure and molecular phylogenetic relationships of some graminicolous symbionts in genera Epichlo? and ParEpichlo? Mycologia, 90, 226-234. [本文引用: 1]
XiaC, ZhangXX, ChristensenMJ, NanZB, LiCJ ( 2015). Epichlo? endophyte affects the ability of powdery mildew (Blumeria graminis) to colonise drunken horse grass(Achnatherum inebrians) Fungal Ecology, 16, 26-33. [本文引用: 1]
YangGW, LiuN, LuWJ, WangS, KanHM, ZhangYJ, XuL, ChenYL ( 2014). The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability Journal of Ecology, 102, 1072-1082. DOI:10.1111/1365-2745.12249URL [本文引用: 1]
YangGW, YangX, ZhangWJ, WeiYQ, GeG, LuWJ, SunJJ, LiuN, KanHM, ShenY, ZhangYJ ( 2015). Arbuscular mycorrhizal fungi affect plant community structure under various nutrient conditions and stabilize the community productivity Oikos, 125, 576-585. [本文引用: 2]
ZhangX, RenAZ, WeiYK, LinF, LiC, LiuZJ, GaoYB ( 2009). Taxonomy, diversity and origins of symbiotic endophytes of Achnatherum sibiricum in the Inner Mongolia Steppe of China FEMS Microbiology Letters, 301, 12-20. [本文引用: 1]
ZhouF, GaoYB, MaWJ ( 2003). Effects of phosphorus deficiency on growth of perennial ryegrass-fungal endophyte symbiont and phenolic content in root Plant Physiology Communications , 39, 321-324. [本文引用: 2]
Testing mechanisms of N-enrichment- induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long- term ecosystem responses 1 2012
... 氮、磷添加量参考已有的研究报道(Lan & Bai, 2012; Yang et al., 2014, 2015), 对于供氮组, 每周施加39.2 mmol·L-1 NH4NO3一次, 每次10 mL, 相当于10 g·m-2·a-1; 对于供磷组, 每周施加35.4 mmol·L-1 KH2PO4一次, 每次10 mL, 相当于10 g·m-2·a-1.对照组添加等量的水.刈割处理在幼苗定植后的第8周和第16周分别进行, 留茬高度为5 cm. ...
Artificial infection of grasses with endophytes 1985
Nomenclatural realignment of Neotyphodium species with genus Epichlo? 1 2014
... 内生真菌(fungal endophyte)是指在植物中度过大部分或全部的生命周期但不使宿主植物表现明显病症的一类真菌(Arnold et al., 2000).目前已知的感染禾本科植物的内生真菌共有7个属, 分别是Atkinsonella、Balansia、Balansiopsis、Echinodothis、Epichlo?、Myriogenospora和ParEpichlo? (White & Reddy, 1998), 其中研究较为深入的是与冷季型禾草共生的子囊菌纲(Ascomycetes)麦角菌科(Clavicipitaceae)的Epichlo?内生真菌(其无性型曾被命名为Neotyphodium).宿主植物研究最多的是人工禾草黑麦草(Lolium perenne)和苇状羊茅(Festuca arundinacea), 二者分别与内生真菌Epichlo? festucae var. lolii和Epichlo? coenophiala构成共生关系(Cheplick, 2004; Malinouski & Belesky, 2006; Leuchtmann et al., 2014).已有研究表明: 内生真菌不仅可以促进宿主植物的生长发育(Clay, 1987), 而且能够提高宿主植物对干旱(Elmi & West, 1995; Oberhofer et al., 2014)、食草动物取食(Rudgers & Clay, 2008; Schardl et al., 2012)、病原菌危害(Christensen, 1996; Xia et al., 2015)等非生物和生物胁迫的抗性. ...
Effects of biotic and abiotic stress on the growth of three genotypes of Lolium perenne with and without infection by the fungal endophyte Neotyphodium lolii 2 2004
... 越来越多的研究表明, 内生真菌对宿主植物的影响与所在环境的养分供应有关.在氮供应方面, 目前的研究大多认为当宿主植株处于低氮生长环境时, 内生真菌会因为对养分的消耗而对宿主产生不利影响(Marks & Clay, 2007), 但也有少数研究认为内生真菌在低氮生长条件下对宿主有利(Arachavaleta et al., 1989; Cheplick et al., 1989; Lewis et al., 1996; Ravel et al., 1997; Newman et al., 2003; Lewis et al., 2004; Ren et al., 2009).在磷供应方面, 以人工禾草苇状羊茅和黑麦草为对象的研究表明, 在低磷处理下, 内生真菌能够提高禾草宿主对土壤磷胁迫的抵御能力, 而在正常供磷条件下, 内生真菌对宿主的有益贡献消失(Malinowski et al., 1998, 1999, 2000; Ren et al., 2007).本研究表明, 内生真菌对宿主羽茅的促进作用出现在供氮条件下, 与已有的多数报道相一致, 但内生真菌-禾草共生体对磷供应的响应与已有的在人工禾草中的研究结果不同. 已有研究中, 只有在低磷条件下内生真菌感染才会促进宿主植物的生长(Malinowski et al., 1999; 周芳等, 2003), 本研究中磷添加显著降低E-羽茅的多度, 但对E+羽茅无显著影响,导致在磷添加条件下内生真菌感染显著提高了宿主植物的多度, 这可能与本实验所采用的土壤本底中的养分含量有关.本实验所用土壤是植物原生境土壤, 即使在无磷添加的处理下, 土壤的全磷浓度仍达到0.03%, 没有出现已报道文献中的低磷处理.进一步对优势种磷浓度的分析发现, E-羽茅所在群落中, 羽茅的磷浓度未随磷添加而增加, 而羊草和冷蒿的磷浓度均有随磷添加而增加的趋势(分别增加了22%和21%); 对于E+羽茅群落, 群落中优势种的磷浓度均有随磷添加而增加的趋势, 只是增加幅度不同, 所以我们推测: E-羽茅群落中磷添加导致E-羽茅多度下降可能与磷添加促进羊草和冷蒿等的生长而间接抑制了羽茅的生长有关; 而对于E+羽茅群落, 磷添加对生长影响的差异不明显, 因此对E+羽茅的多度影响不显著. ...
Effect of infection by the endophytic fungus Acremonium lolii on growth and nitrogen uptake by perennial ryegrass(Lolium perenne) in flowing solution culture 1 1996
... 越来越多的研究表明, 内生真菌对宿主植物的影响与所在环境的养分供应有关.在氮供应方面, 目前的研究大多认为当宿主植株处于低氮生长环境时, 内生真菌会因为对养分的消耗而对宿主产生不利影响(Marks & Clay, 2007), 但也有少数研究认为内生真菌在低氮生长条件下对宿主有利(Arachavaleta et al., 1989; Cheplick et al., 1989; Lewis et al., 1996; Ravel et al., 1997; Newman et al., 2003; Lewis et al., 2004; Ren et al., 2009).在磷供应方面, 以人工禾草苇状羊茅和黑麦草为对象的研究表明, 在低磷处理下, 内生真菌能够提高禾草宿主对土壤磷胁迫的抵御能力, 而在正常供磷条件下, 内生真菌对宿主的有益贡献消失(Malinowski et al., 1998, 1999, 2000; Ren et al., 2007).本研究表明, 内生真菌对宿主羽茅的促进作用出现在供氮条件下, 与已有的多数报道相一致, 但内生真菌-禾草共生体对磷供应的响应与已有的在人工禾草中的研究结果不同. 已有研究中, 只有在低磷条件下内生真菌感染才会促进宿主植物的生长(Malinowski et al., 1999; 周芳等, 2003), 本研究中磷添加显著降低E-羽茅的多度, 但对E+羽茅无显著影响,导致在磷添加条件下内生真菌感染显著提高了宿主植物的多度, 这可能与本实验所采用的土壤本底中的养分含量有关.本实验所用土壤是植物原生境土壤, 即使在无磷添加的处理下, 土壤的全磷浓度仍达到0.03%, 没有出现已报道文献中的低磷处理.进一步对优势种磷浓度的分析发现, E-羽茅所在群落中, 羽茅的磷浓度未随磷添加而增加, 而羊草和冷蒿的磷浓度均有随磷添加而增加的趋势(分别增加了22%和21%); 对于E+羽茅群落, 群落中优势种的磷浓度均有随磷添加而增加的趋势, 只是增加幅度不同, 所以我们推测: E-羽茅群落中磷添加导致E-羽茅多度下降可能与磷添加促进羊草和冷蒿等的生长而间接抑制了羽茅的生长有关; 而对于E+羽茅群落, 磷添加对生长影响的差异不明显, 因此对E+羽茅的多度影响不显著. ...
A new Neotyphodium species symbiotic with drunken horse grass(Achnatherum inebrians) in China 1 2004
... 羽茅(Achnatherum sibiricum)作为一种疏丛型禾草, 是在羊草(Leymus chinensis)群落和大针茅群落中常见的伴生种.与羽茅同为芨芨草属(Achnatherum)的另外两个种, 即分布于我国天然草原的醉马草(Achnatherum inebrians)和分布于美洲大陆的Achnatherum robustum均感染内生真菌, 且均能够产生使草食家畜中毒的生物碱(李学森等, 1998; Jones et al., 2000; Li et al., 2004), 而羽茅则不同, Zhang等(2009)发现, 在正常生长条件下, 羽茅在同一地理种群感染有两种不同种类的优势菌株(Epichlo? gansuensis和Epichlo? sibirica), 且均没有检出生物碱, 刈割处理后, 少数染菌羽茅样品中检出波胺, 但含量极低, 不足以对食草动物产生毒性(Qin et al., 2016), 因此羽茅是研究内生真菌和天然禾草共生关系的理想材料.考虑到刈割是内蒙古草地一种最普遍的利用方式, 本研究通过构建包含内生真菌感染(E+)和不感染(E-)羽茅的人工群落, 探究在不同养分水平和刈割干扰处理下, 内生真菌感染对宿主所在群落的影响, 我们提出以下3个科学问题: (1)内生真菌感染是否会影响宿主所在群落的物种多样性?(2)内生真菌感染是否会影响宿主所在群落的生产力?(3)内生真菌对宿主群落的影响是否与养分处理和刈割干扰有关? ...
高温处理构建不感染内生真菌羽茅种群的方法探讨 2010
高温处理构建不感染内生真菌羽茅种群的方法探讨 2010
醉马草有毒物质与其内生真菌的关系 1 1998
... 羽茅(Achnatherum sibiricum)作为一种疏丛型禾草, 是在羊草(Leymus chinensis)群落和大针茅群落中常见的伴生种.与羽茅同为芨芨草属(Achnatherum)的另外两个种, 即分布于我国天然草原的醉马草(Achnatherum inebrians)和分布于美洲大陆的Achnatherum robustum均感染内生真菌, 且均能够产生使草食家畜中毒的生物碱(李学森等, 1998; Jones et al., 2000; Li et al., 2004), 而羽茅则不同, Zhang等(2009)发现, 在正常生长条件下, 羽茅在同一地理种群感染有两种不同种类的优势菌株(Epichlo? gansuensis和Epichlo? sibirica), 且均没有检出生物碱, 刈割处理后, 少数染菌羽茅样品中检出波胺, 但含量极低, 不足以对食草动物产生毒性(Qin et al., 2016), 因此羽茅是研究内生真菌和天然禾草共生关系的理想材料.考虑到刈割是内蒙古草地一种最普遍的利用方式, 本研究通过构建包含内生真菌感染(E+)和不感染(E-)羽茅的人工群落, 探究在不同养分水平和刈割干扰处理下, 内生真菌感染对宿主所在群落的影响, 我们提出以下3个科学问题: (1)内生真菌感染是否会影响宿主所在群落的物种多样性?(2)内生真菌感染是否会影响宿主所在群落的生产力?(3)内生真菌对宿主群落的影响是否与养分处理和刈割干扰有关? ...
醉马草有毒物质与其内生真菌的关系 1 1998
... 羽茅(Achnatherum sibiricum)作为一种疏丛型禾草, 是在羊草(Leymus chinensis)群落和大针茅群落中常见的伴生种.与羽茅同为芨芨草属(Achnatherum)的另外两个种, 即分布于我国天然草原的醉马草(Achnatherum inebrians)和分布于美洲大陆的Achnatherum robustum均感染内生真菌, 且均能够产生使草食家畜中毒的生物碱(李学森等, 1998; Jones et al., 2000; Li et al., 2004), 而羽茅则不同, Zhang等(2009)发现, 在正常生长条件下, 羽茅在同一地理种群感染有两种不同种类的优势菌株(Epichlo? gansuensis和Epichlo? sibirica), 且均没有检出生物碱, 刈割处理后, 少数染菌羽茅样品中检出波胺, 但含量极低, 不足以对食草动物产生毒性(Qin et al., 2016), 因此羽茅是研究内生真菌和天然禾草共生关系的理想材料.考虑到刈割是内蒙古草地一种最普遍的利用方式, 本研究通过构建包含内生真菌感染(E+)和不感染(E-)羽茅的人工群落, 探究在不同养分水平和刈割干扰处理下, 内生真菌感染对宿主所在群落的影响, 我们提出以下3个科学问题: (1)内生真菌感染是否会影响宿主所在群落的物种多样性?(2)内生真菌感染是否会影响宿主所在群落的生产力?(3)内生真菌对宿主群落的影响是否与养分处理和刈割干扰有关? ...
The effect of endophytic fungi on the fitness of Arizona fescue (Festuca arizonica) under varying nitrogen levels 1 1997
Beneficial effects of Neotyphodium lolii on the growth and the water status in perennial ryegrass cultivated under nitrogen deficiency or drought stress 1 1997
... 越来越多的研究表明, 内生真菌对宿主植物的影响与所在环境的养分供应有关.在氮供应方面, 目前的研究大多认为当宿主植株处于低氮生长环境时, 内生真菌会因为对养分的消耗而对宿主产生不利影响(Marks & Clay, 2007), 但也有少数研究认为内生真菌在低氮生长条件下对宿主有利(Arachavaleta et al., 1989; Cheplick et al., 1989; Lewis et al., 1996; Ravel et al., 1997; Newman et al., 2003; Lewis et al., 2004; Ren et al., 2009).在磷供应方面, 以人工禾草苇状羊茅和黑麦草为对象的研究表明, 在低磷处理下, 内生真菌能够提高禾草宿主对土壤磷胁迫的抵御能力, 而在正常供磷条件下, 内生真菌对宿主的有益贡献消失(Malinowski et al., 1998, 1999, 2000; Ren et al., 2007).本研究表明, 内生真菌对宿主羽茅的促进作用出现在供氮条件下, 与已有的多数报道相一致, 但内生真菌-禾草共生体对磷供应的响应与已有的在人工禾草中的研究结果不同. 已有研究中, 只有在低磷条件下内生真菌感染才会促进宿主植物的生长(Malinowski et al., 1999; 周芳等, 2003), 本研究中磷添加显著降低E-羽茅的多度, 但对E+羽茅无显著影响,导致在磷添加条件下内生真菌感染显著提高了宿主植物的多度, 这可能与本实验所采用的土壤本底中的养分含量有关.本实验所用土壤是植物原生境土壤, 即使在无磷添加的处理下, 土壤的全磷浓度仍达到0.03%, 没有出现已报道文献中的低磷处理.进一步对优势种磷浓度的分析发现, E-羽茅所在群落中, 羽茅的磷浓度未随磷添加而增加, 而羊草和冷蒿的磷浓度均有随磷添加而增加的趋势(分别增加了22%和21%); 对于E+羽茅群落, 群落中优势种的磷浓度均有随磷添加而增加的趋势, 只是增加幅度不同, 所以我们推测: E-羽茅群落中磷添加导致E-羽茅多度下降可能与磷添加促进羊草和冷蒿等的生长而间接抑制了羽茅的生长有关; 而对于E+羽茅群落, 磷添加对生长影响的差异不明显, 因此对E+羽茅的多度影响不显著. ...
Influence of nitrogen fertilizer and endophyte infection on ecophysiological parameters and mineral element content of perennial ryegrass 2 2009
Examination of structure and molecular phylogenetic relationships of some graminicolous symbionts in genera Epichlo? and ParEpichlo? 1 1998
... 内生真菌(fungal endophyte)是指在植物中度过大部分或全部的生命周期但不使宿主植物表现明显病症的一类真菌(Arnold et al., 2000).目前已知的感染禾本科植物的内生真菌共有7个属, 分别是Atkinsonella、Balansia、Balansiopsis、Echinodothis、Epichlo?、Myriogenospora和ParEpichlo? (White & Reddy, 1998), 其中研究较为深入的是与冷季型禾草共生的子囊菌纲(Ascomycetes)麦角菌科(Clavicipitaceae)的Epichlo?内生真菌(其无性型曾被命名为Neotyphodium).宿主植物研究最多的是人工禾草黑麦草(Lolium perenne)和苇状羊茅(Festuca arundinacea), 二者分别与内生真菌Epichlo? festucae var. lolii和Epichlo? coenophiala构成共生关系(Cheplick, 2004; Malinouski & Belesky, 2006; Leuchtmann et al., 2014).已有研究表明: 内生真菌不仅可以促进宿主植物的生长发育(Clay, 1987), 而且能够提高宿主植物对干旱(Elmi & West, 1995; Oberhofer et al., 2014)、食草动物取食(Rudgers & Clay, 2008; Schardl et al., 2012)、病原菌危害(Christensen, 1996; Xia et al., 2015)等非生物和生物胁迫的抗性. ...
Epichlo? endophyte affects the ability of powdery mildew (Blumeria graminis) to colonise drunken horse grass(Achnatherum inebrians) 1 2015
... 内生真菌(fungal endophyte)是指在植物中度过大部分或全部的生命周期但不使宿主植物表现明显病症的一类真菌(Arnold et al., 2000).目前已知的感染禾本科植物的内生真菌共有7个属, 分别是Atkinsonella、Balansia、Balansiopsis、Echinodothis、Epichlo?、Myriogenospora和ParEpichlo? (White & Reddy, 1998), 其中研究较为深入的是与冷季型禾草共生的子囊菌纲(Ascomycetes)麦角菌科(Clavicipitaceae)的Epichlo?内生真菌(其无性型曾被命名为Neotyphodium).宿主植物研究最多的是人工禾草黑麦草(Lolium perenne)和苇状羊茅(Festuca arundinacea), 二者分别与内生真菌Epichlo? festucae var. lolii和Epichlo? coenophiala构成共生关系(Cheplick, 2004; Malinouski & Belesky, 2006; Leuchtmann et al., 2014).已有研究表明: 内生真菌不仅可以促进宿主植物的生长发育(Clay, 1987), 而且能够提高宿主植物对干旱(Elmi & West, 1995; Oberhofer et al., 2014)、食草动物取食(Rudgers & Clay, 2008; Schardl et al., 2012)、病原菌危害(Christensen, 1996; Xia et al., 2015)等非生物和生物胁迫的抗性. ...
The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability 1 2014
... 氮、磷添加量参考已有的研究报道(Lan & Bai, 2012; Yang et al., 2014, 2015), 对于供氮组, 每周施加39.2 mmol·L-1 NH4NO3一次, 每次10 mL, 相当于10 g·m-2·a-1; 对于供磷组, 每周施加35.4 mmol·L-1 KH2PO4一次, 每次10 mL, 相当于10 g·m-2·a-1.对照组添加等量的水.刈割处理在幼苗定植后的第8周和第16周分别进行, 留茬高度为5 cm. ...
Arbuscular mycorrhizal fungi affect plant community structure under various nutrient conditions and stabilize the community productivity 2 2015
... 氮、磷添加量参考已有的研究报道(Lan & Bai, 2012; Yang et al., 2014, 2015), 对于供氮组, 每周施加39.2 mmol·L-1 NH4NO3一次, 每次10 mL, 相当于10 g·m-2·a-1; 对于供磷组, 每周施加35.4 mmol·L-1 KH2PO4一次, 每次10 mL, 相当于10 g·m-2·a-1.对照组添加等量的水.刈割处理在幼苗定植后的第8周和第16周分别进行, 留茬高度为5 cm. ...
... 式中, N为群落中物种的总数, bi为第i种物种生物量; 物种多度是物种i的生物量占总生物量的百分比(Yang et al., 2015). ...
Taxonomy, diversity and origins of symbiotic endophytes of Achnatherum sibiricum in the Inner Mongolia Steppe of China 1 2009
... 羽茅(Achnatherum sibiricum)作为一种疏丛型禾草, 是在羊草(Leymus chinensis)群落和大针茅群落中常见的伴生种.与羽茅同为芨芨草属(Achnatherum)的另外两个种, 即分布于我国天然草原的醉马草(Achnatherum inebrians)和分布于美洲大陆的Achnatherum robustum均感染内生真菌, 且均能够产生使草食家畜中毒的生物碱(李学森等, 1998; Jones et al., 2000; Li et al., 2004), 而羽茅则不同, Zhang等(2009)发现, 在正常生长条件下, 羽茅在同一地理种群感染有两种不同种类的优势菌株(Epichlo? gansuensis和Epichlo? sibirica), 且均没有检出生物碱, 刈割处理后, 少数染菌羽茅样品中检出波胺, 但含量极低, 不足以对食草动物产生毒性(Qin et al., 2016), 因此羽茅是研究内生真菌和天然禾草共生关系的理想材料.考虑到刈割是内蒙古草地一种最普遍的利用方式, 本研究通过构建包含内生真菌感染(E+)和不感染(E-)羽茅的人工群落, 探究在不同养分水平和刈割干扰处理下, 内生真菌感染对宿主所在群落的影响, 我们提出以下3个科学问题: (1)内生真菌感染是否会影响宿主所在群落的物种多样性?(2)内生真菌感染是否会影响宿主所在群落的生产力?(3)内生真菌对宿主群落的影响是否与养分处理和刈割干扰有关? ...
 ,*, 高玉葆南开大学生命科学学院, 天津 300071
,*, 高玉葆南开大学生命科学学院, 天津 300071
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT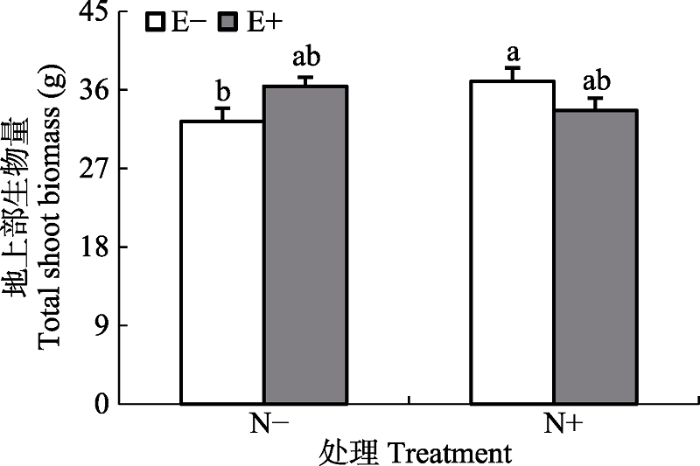 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT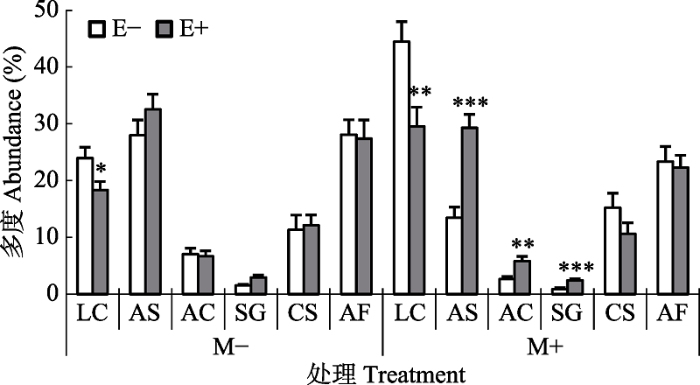 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT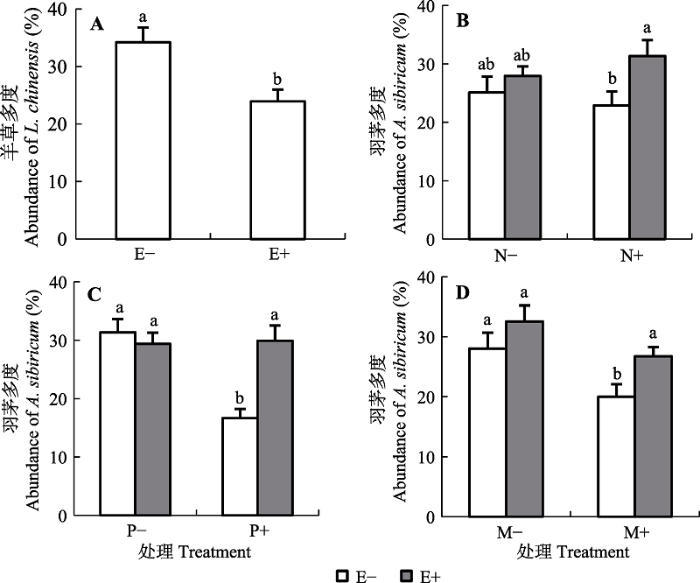 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT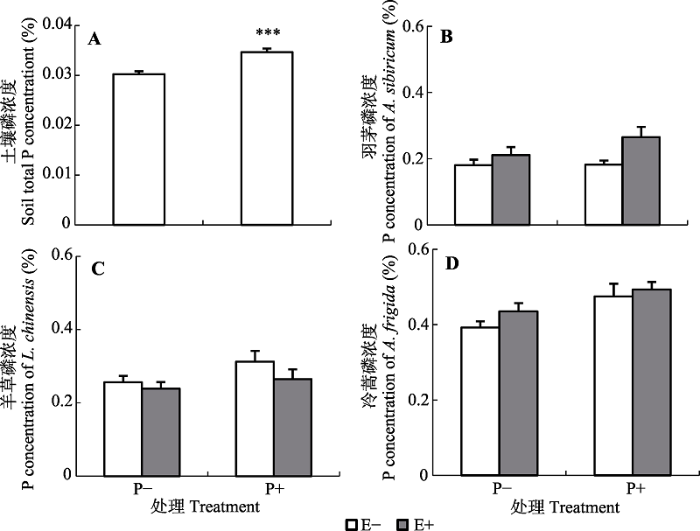 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}