高文童
1 , 张春艳
1 , 2 , 董廷发
1 , 2 , 胥晓
, 1 , 2 , * 1 西华师范大学生命科学学院, 四川南充 637009 2 西华师范大学西南野生动植物资源保护教育部重点实验室, 四川南充 637009 Effects of arbuscular mycorrhizal fungi on the root growth of male and female Populus cathayana individuals grown under different sexual combination patterns GAO Wen-Tong
1 , ZHANG Chun-Yan
1 , 2 , DONG Ting-Fa
1 , 2 , XU Xiao
, 1 , 2 , * 1 College of Life Sciences, China West Normal University, Nanchong, Sichuan 637009, China 2 Key Laboratory of Southwest China Wildlife Resources Conservation (China West Normal University), Ministry of Education, Nanchong, Sichuan 637009, China 通讯作者: * xuxiao_cwnu@163.com 编委: 陈保冬
责任编辑: 李敏
收稿日期: 2018-10-29
接受日期: 2019-01-09
网络出版日期: 2019-01-20
Received: 2018-10-29
Accepted: 2019-01-09
Online: 2019-01-20
Fund supported: Supported by the National Natural Science Foundation of China .31370596
摘要 丛枝菌根真菌(AMF)对雌雄异株植物根系生长, 尤其是对邻近生长的不同雌雄个体的影响还鲜有研究。该研究以泥土:河沙:蛭石体积比为1:1:1的混合物为培养基质, 分别在雄-雄、雌-雌和雄-雌3种组合栽培模式下对青杨(Populus cathayana )雌雄幼苗进行接种和不接种摩西球囊霉(Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系在侵染率、生物量、形态、碳、氮含量等方面的差异来分析AMF对青杨雌雄幼苗根系生长发育的影响。结果发现: 与对照组相比, 接种AMF对3种栽培模式下青杨雌雄植株的侵染率、根干质量、根系形态(除分枝强度、比表面积)和碳、氮含量影响显著。此外, 不同性别组合模式对青杨雌雄植株的根干质量、根系形态和碳、氮含量影响显著。接种AMF后, 与雌-雌合栽模式下的雌株相比, 雄-雌合栽模式下雌株的根干质量、氮含量都有不同程度的提高, 根系形态发生改变; 而与雄-雄合栽模式下的雄株相比, 雄-雌合栽模式下雄株的相应指标出现降低或轻微增加。该研究表明AMF对不同性别组合模式下青杨植株根系生长具有显著促进作用, 尤其是雄-雌合栽模式下AMF接种最有利于雌株根系的生长发育。 关键词: 丛枝菌根真菌 ;
青杨 ;
雌雄异株 ;
栽培模式 ;
根系生长 Abstract Aims Methods Populus cathayana saplings uninoculated and inoculated with AMF under three sex combination patterns (male vs. male, MM; female vs. female, FF; male vs. female, MF) were defined as control (CK) and AMF treatment group, respectively. Subsequently, we compared the differences in colonization rate, root dry mass, root morphology, carbon (C) content and nitrogen (N) content between CK and AMF treatments under different sexual combination patterns.Important findings P . cathayana . After inoculation with AMF, root dry mass, root morphology and N content of female individuals were increased whereas these parameters of males were decreased or slightly increased in inter-sexual groups compared with the respective intra-sexual groups. Collectively, our data demonstrate the growth-promoting effects of AMF on the roots of P. cathayana individuals grown under different sexual combination patterns, and such beneficial effects are most pronounced in females grown under inter-sexual combination patterns.Keywords: arbuscular mycorrhizal fungi ;
Populus cathayana ;
dioecious ;
cultivation pattern ;
root growth PDF (2235KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 本文引用格式 高文童, 张春艳, 董廷发, 胥晓. 丛枝菌根真菌对不同性别组合模式下青杨雌雄植株根系生长的影响.
植物生态学报 [J], 2019, 43(1): 37-45 doi:10.17521/cjpe.2018.0261
GAO Wen-Tong.
Effects of arbuscular mycorrhizal fungi on the root growth of male and female Populus cathayana individuals grown under different sexual combination patterns .
Chinese Journal of Plant Ecology [J], 2019, 43(1): 37-45 doi:10.17521/cjpe.2018.0261
丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 )。它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等。随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视。大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 )。然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物。
相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 )。如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 )。此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等。虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因。由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 )。因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一。然而, 相关的研究并未见报道。
青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物。该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 )。目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道。因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响。研究结果可为青杨的科学栽培提供理论参考。
1 材料和方法 1.1 实验材料和生长环境 实验地点位于四川省南充市西华师范大学试验地(106.07° E, 30.80° N, 海拔276 m)。该地区属亚热带季风性湿润气候, 年平均气温、降水量、日照时间分别为16.8 ℃、1 065 mm、1 980 h (
罗培和周申立, 2007 )。2017年3月从试验地的青杨成树(引种自青海大通)上采集一年生枝条进行扦插(雌雄各50根扦插条)。扦插所用的基质为泥土、河沙、蛭石体积比1:1:1的混合物, 于100 ℃蒸汽灭菌2 h两次以消除AMF的繁殖体和其他微生物。在扦插之前将枝条用70%酒精消毒, 并用蒸馏水漂洗, 最后放入生根粉溶液中浸泡约15 s后进行扦插。待扦插条发芽生长60天后, 选取长短、粗细、健康状况一致的84株幼苗(雌雄各42株)移栽到长宽高分别为40 cm × 20 cm × 30 cm的玻璃缸中(分别按雄-雄, 雌-雌, 雄-雌组合栽培)。实验中的玻璃缸内部装置与
Johansen和Jensen (1996) 的类似。在玻璃缸中间放置两块厚0.25 cm的多孔有机玻璃板和一张单个孔径为37 μm的尼龙网(尼龙网放置在两个玻璃板之间)将容器分割为两个室。该结构的目的是防止根系穿过, 但允许菌丝相连(参考
He et al ., 2004 )(
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1不同性别合栽模式示意图, 每盒含两株幼苗。 Fig. 1Schematic diagram of mixed-sexes plantation designs, each with two seedlings. 1.2 实验设计 实验采用双因素完全随机设计。2种接种处理: 接种或不接种摩西球囊霉, 3种性别组合: 雄-雄、雌-雌、雄-雌。在接种处理组中每一株幼苗接种10 g菌剂(接种剂来自北京市农林科学院植物营养与资源研究所, 包括纯培养物的基质、根系、孢子以及外生菌丝), 未接种处理组中每一株幼苗接种进行蒸汽灭菌后的菌剂10 g, 并施加10 mL不含AMF的接种剂滤液。以没有接种菌剂的3种性别组合为对照组, 以接种菌剂的3种性别组合为接种组, 总共6种组合, 每种组合7个重复。
1.3 测定指标和方法 1.3.1 根系形态、生物量、碳、氮含量的测定
实验处理约180天后(一个生长季), 于2017年9月对不同处理组的青杨雌雄幼苗进行采收, 从根系 上选取2-3个具有代表性的侧枝用根系分析系统(WinRHIZO, Regents Instruments, Quebec, Canada)进行扫描, 并测量各株幼苗的总根长、根尖数, 计算分枝强度、比根长和比表面积。最后将收集的根系放在恒温干燥箱中70 ℃, 48 h烘干至恒质量, 随后用电子天平称量根系总干质量。最后使用碳氮分析仪(Vario TOC, Elementar Company, Hanau, German)测量根系的碳、氮含量。
1.3.2 根系侵染率的测定
将每种组合下的植株(7个重复)根系分别洗净后保存在FAA固定液中, 固定完成后每个植物根系样品选择至少100段直径小于1 mm, 长度约1 cm的新鲜根段放在10%的KOH溶液中90 ℃水浴加热1 h, 待加热结束后放入碱性双氧水中进行脱色, 而后取出根系用墨水和醋(95%醋和5%墨水)进行染色, 随后用酸化的水冲洗根段数次(
Vierheilig et al ., 2010 ), 最后随机选取70个根段整齐排列在载玻片上用显微镜进行观测。根据McGonigle等(1990)的方法用以下公式计算菌根侵染率:
菌根侵染率(%) = 菌根根段/观察的总根段数× 100%
1.4 数据分析和处理 采用SPSS 19.0进行数据分析。分析前对数据进行方差齐性检验, 不符合方差齐性时对数据进行对数转换。组合、接种及其交互作用的评价采用双因素方差分析, 组间平均值的比较采用Duncan’s多重比较。显著性水平设定为
α = 0. 05。
2 结果和分析 2.1 青杨雌雄植株侵染率的差异 由
图2A 可见, 接种AMF显著提高了3种栽培模式下青杨雌雄植株的菌根侵染率(
p < 0.001), 尤其是雄-雌合栽模式下的雌株菌根侵染率与对照相比增加了14.3倍。不同栽培模式对青杨雌雄植株的菌根侵染率无显著影响, 但接种AMF后导致雄-雌合栽模式下的雌株菌根侵染率显著高于雄株。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2接种丛枝菌根真菌(AMF)对不同栽培模式下青杨雌雄植株菌根侵染率和根干质量的影响(平均值±标准误差, n = 7)。柱形上方不同小写字母表示相应处理之间差异显著(采用Duncan’s多重比较, p < 0.05)。C, 组合效应; I, 接种效应; C × I, 组合与接种交互效应。ns, 没有显著差异; ***, p < 0.001。MM/M, 性别内组合下的雄株; FF/F, 性别内组合下的雌株; MF/M, 性别间组合下的雄株; MF/F, 性别间组合下的雌株。 Fig. 2Effects of arbuscular mycorrhizal fungi (AMF) on the colonization rate and root dry mass of male and female Populus cathayana individuals grown under different sexual combination patterns (mean ± SE , n = 7). Different lowercase letters above the columns represent significant differences between corresponding treatments according to Duncan’s multiple range test (p < 0.05). C, combination pattern effect; I, inoculation effect; C × I, combination pattern and inoculation interaction effect. ns, not significant; ***, p < 0.001. MM/M, males in the intra-sexual combination; FF/F, females in the intra-sexual combination; MF/M, males in the inter-sexual combination; MF/F, females in the inter-sexual combination. 2.2 青杨雌雄植株根干质量的差异 由
图2B 可见, 与对照组相比, 接种AMF显著增加了不同性别组合栽培模式下青杨雌雄植株的根干质量(
p < 0.001), 其中雄-雄合栽模式下雄株根干质量增加最多, 是未接种的1.7倍; 不同栽培模式对青杨植株的根干质量也有显著影响(
p < 0.001)。未接种AMF时, 雄-雌合栽模式下的雌株根干质量高于雌-雌合栽中的雌株, 但雄株无显著差异; 接种AMF处理下, 不同合栽模式下相应性别植株根干质量均无显著差异。此外, 在3种栽培模式中雌株的根干质量均显著高于雄株。
2.3 青杨雌雄植株根系形态的差异 由
表1 知, 与对照组相比, 接种AMF对不同性别组合栽培模式下青杨植株根系的比根长、总根长和根尖数影响显著(
p < 0.05), 其中雄-雌合栽模式下雄株的总根长以及雌株的比根长和根尖数增加最多, 而雄-雄合栽模式下青杨雄株的比根长显著下降; 不同栽培模式对青杨植株的分枝强度、比表面积、比根长、总根长和根尖数均有显著影响(
p < 0.05)。在对照组中雄-雄合栽模式下雄株的比表面积和比根长显著大于雄-雌合栽模式下的雄株, 而分枝强度则相反。雌-雌合栽模式下雌株的分枝强度显著小于雄-雌合栽模式下的雌株。在接种AMF后, 雄-雄合栽模式下雄株的分枝强度显著小于雄-雌合栽下的雄株, 而雌-雌合栽模式下雌株的分枝强度、比根长、总根长和根尖数均显著小于雄-雌合栽模式下的雌株。此外, 由
图3 可见, 与对照相比, 接种AMF的青杨植株, 尤其是雄-雌合栽模式下的雌株的根系构型明显优于未接种的植株。
Table 1 表1 表1 接种丛枝菌根真菌(AMF)对不同栽培模式下青杨雌雄植株根系形态的影响(平均值±标准误差,
n = 7)
Table 1
Effects of arbuscular mycorrhizal fungi (AMF) on the root morphology of male and female Populus cathayana individuals grown under different sexual combination patterns (mean ± SE, n = 7) 组别 组合 分枝强度2 ) 比表面积2 ) 比根长3 ) 总根长3 ) 根尖数3 ) 对照 雄雄/雄 MM/M 5.01 ± 0.34b 3.49 ± 0.49ab 5.97 ± 0.49b 25.58 ± 2.23ef 20.97 ± 1.69cd 雌雌/雌 FF/F 4.26 ± 0.37b 2.99 ± 0.24bc 4.58 ± 0.34cd 33.08 ± 2.39de 19.09 ± 1.84d 雄雌/雄 MF/M 7.81 ± 0.52a 1.75 ± 0.27d 3.80 ± 0.28d 14.81 ± 1.25f 17.71 ± 1.37d 雄雌/雌 MF/F 7.83 ± 0.29a 2.70 ± 0.44c 5.04 ± 0.54bcd 42.46 ± 4.38cd 18.33 ± 1.50d AMF 雄雄/雄 MM/M 4.46 ± 0.30b 2.14 ± 0.20cd 4.09 ± 0.18d 58.47 ± 4.25c 25.66 ± 1.27c 雌雌/雌 FF/F 4.83 ± 0.34b 3.28 ± 0.16ab 5.67 ± 0.48bc 105.30 ± 9.82b 38.47 ± 1.75b 雄雌/雄 MF/M 6.70 ± 0.53a 2.89 ± 0.20bc 4.99 ± 0.43bcd 55.88 ± 5.66c 25.95 ± 1.51c 雄雌/雌 MF/F 6.88 ± 0.31a 3.99 ± 0.31a 7.29 ± 0.52a 137.96 ± 8.17a 51.88 ± 3.42a p > F I 0.069ns 0.126ns 0.033* <0.001*** <0.001*** p > F C <0.001*** 0.011* 0.002** <0.001*** <0.001*** p > F C×I 0.142ns <0.001*** <0.001*** <0.001*** <0.001***
Different lowercase letters following the means in the same column represent significant differences between corresponding treatments according to Duncan’s multiple range test (p < 0.05). C, combination pattern effect; I, inoculation effect; C × I, combination pattern and inoculation interaction effect. ns, not significant; ***, p < 0.001. MM/M, males in the intra-sexual combination; FF/F, females in the intra-sexual combination; MF/M, males in the inter-sexual combination; MF/F, females in the inter-sexual combination. 同列均值后不同小写字母表示相应处理之间差异显著(采用Duncan’s多重比较, p < 0.05)。C, 组合效应; I, 接种效应; C × I, 组合与接种交互效应。ns, 没有显著差异; ***, p < 0.001。MM/M, 性别内组合下的雄株; FF/F, 性别内组合下的雌株; MF/M, 性别间组合下的雄株; MF/F, 性别间组合下的雌株。 新窗口打开 |
下载CSV 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3接种丛枝菌根真菌(AMF)对不同栽培模式下青杨雌雄植株根系构型的影响。MM/M, 性别内组合下的雄株; FF/F, 性别内组合下的雌株; MF/M, 性别间组合下的雄株; MF/F, 性别间组合下的雌株。 Fig. 3Effects of arbuscular mycorrhizal fungi (AMF) on the root system architecture of male and female Populus cathayana individuals grown under different sexual combination patterns. MM/M, males in the intra-sexual combination; FF/F, females in the intra-sexual combination; MF/M, males in the inter-sexual combination; MF/F, females in the inter-sexual combination. 2.4 青杨雌雄植株根系碳、氮含量的差异 由
图4 可见, 与对照组相比, 接种AMF处理对3种性别栽培模式下青杨雌雄植株的碳、氮含量影响显著(
p < 0.001和
p < 0.01), 接种AMF后雌-雌合栽模式下雌株的碳、氮含量分别增加了6%和49%; 不同栽培模式对青杨雌雄植株碳、氮含量也有显著影响(
p < 0.05和
p < 0.001)。在对照组中雌-雌合栽模式下的雌株氮含量显著低于雄-雌合栽模式下的雌株, 而在接种处理组中雄-雄合栽模式下雄株的氮含量显著高于雄-雌合栽模式下的雄株。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4接种丛枝菌根真菌(AMF)对不同栽培模式下青杨雌雄植株根系碳和氮的影响(平均值±标准误差, n = 7)。柱形上方不同小写字母表示相应处理之间差异显著(采用Duncan’s多重比较, p < 0.05)。C, 组合效应; I, 接种效应; C × I, 组合与接种交互效应。ns, 没有显著差异; ***, p < 0.001。MM/M, 性别内组合下的雄株; FF/F, 性别内组合下的雌株; MF/M, 性别间组合下的雄株; MF/F, 性别间组合下的雌株。 Fig. 4Effects of arbuscular mycorrhizal fungi (AMF) on the C content and N content in roots of male and female Populus cathayana individuals grown under different sexual combination patterns (mean ± SE, n = 7). Different lowercase letters above the columns represent significant differences between corresponding treatments according to Duncan’s multiple range test (p < 0.05). C, combination pattern effect; I, inoculation effect; C × I, combination pattern and inoculation interaction effect. ns, not significant; ***, p < 0.001. MM/M, males in the intra-sexual combination; FF/F, females in the intra-sexual combination; MF/M, males in the inter-sexual combination; MF/F, females in the inter-sexual combination. 3 讨论 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题。尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%)。由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上。根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度。因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系。
此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育。AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 )。与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 )。与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用。然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证。
另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响。如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好。其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 )。二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 )。三是不同性别的植株因对资源的需求不同而可能存在互助作用。AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续。本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反。这虽然验证了我们的假设, 但仍需开展进一步的研究。
4 结论 综上所述, 我们以青杨为模式物种, 研究了接种AMF对不同栽培模式下青杨雌雄植株根系生长发育的影响。结果表明AMF、不同性别栽培模式以及两者的交互作用都对青杨雌雄植株的根系生长发育有不同程度的影响; 接种AMF后, 雄-雌合栽最有利于青杨雌株根系的生长发育, 这为杨树的优质培育提供了新的思路。另外, 我们还发现AMF对青杨雌雄植株间的相互作用有潜在的影响, 这对种群的稳定性和延续可能具有重要的意义。
[1] Bonfante P Genre A 2010 ). Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosisNature Communications, 1, 48 . DOI: 10.1038/ncomms1046 . [本文引用: 1] [2] Buee M Rossignol M Jauneau A Ranjeva R Bécard G 2000 ). The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudatesMolecular Plant-Microbe Interactions, 13, 693 -698 . DOI:
10.1094/MPMI.2000.13.6.693 URL [本文引用: 1] [3] Chen J Dong TF Duan BL Korpelainen H Niinemets ü Li CY 2015 a). Sexual competition and N supply interactively affect the dimorphism and competiveness of opposite sexes in Populus cathayana . Plant, Cell & Environment, 1285 -1298 . [本文引用: 2] [4] Chen J Duan BL Wang M Korpelainen H Li CY 2014 ). Intra- and inter-sexual competition of Populus cathayana under different watering regimes. Functional Ecology, 124 -136 . [本文引用: 1] [5] Chen J Han QQ Duan BL Korpelainen H Li CY 2017 ). Sex-specific competition differently regulates ecophysiological responses and phytoremediation of Populus cathayana under Pb stress. Plant and Soil, 203 -218 . [本文引用: 1] [6] Chen LH Hu XW Yang WQ Xu ZF Zhang DJ Gao S 2015 b). The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in Populus cathayana . Ecotoxicology and Environmental Safety, 460 -468 . [本文引用: 2] [7] Chen ZC Shi ZY Tian CY Feng G 2008 ). Effects of arbuscular mycorrhizal fungi inoculation on growth and nutrient uptake of two ephemeral plantsJournal of Plant Ecology (Chinese Version )648 -653 . [本文引用: 1] [ 陈志超 , 石兆勇 , 田长彦 , 冯固 ( 2008 ). 接种AM真菌对短命植物生长发育及矿质养分吸收的影响植物生态学报 , 32, 648 -653 .] [本文引用: 1] [8] Dawson TE Ehleringer JR 1993 ). Gender-specific physiology, carbon isotope discrimination, and habitat distribution in boxelder,Acer negundo. Ecology, 74, 798 -815 . DOI:
10.2307/1940807 URL [本文引用: 1] [9] Dong T Li J Liao Y Liu J Chen B Xu X 2017 ). Root-?mediated sex recognition in a dioecious treeScientific Reports, 7, 801 . DOI: 10.1038/s41598-017-00894-2 . URL [本文引用: 1] [10] Giovannetti M Sbrana C Avio L Strani P 2004 ). Patterns of below-ground plant interconnections established by means of arbuscular mycorrhizal networksNew Phytologist, 164, 175 -181 . DOI:
10.1111/j.1469-8137.2004.01145.x URL [本文引用: 1] [11] Graham JH Leonard RT Menge JA 1981 ). Membrane-?mediated decrease in root exudation responsible for phorphorus inhibition of vesicular-arbuscular mycorrhiza formationPlant Physiology, 68, 548 -552 . DOI:
10.1104/pp.68.3.548 URL [本文引用: 1] [12] Guo HL Liu SJ Xu J Song SR Tang JJ Chen X 2017 ). Effects of arbuscular mycorrhizal fungi on growth and root traits of dicotyledons plants: A meta analysisChinese Journal of Ecology, 36, 1855 -1864 . [本文引用: 1] [ 郭晗铃 , 刘世俊 , 徐静 , 宋书锐 , 唐建军 , 陈欣 ( 2017 ). 丛枝菌根真菌对双子叶植物生长和根系特征的影响: 整合分析生态学杂志 , 36, 1855 -1864 .] [本文引用: 1] [13] Hawkins TS Schiff NM Leininger TD Gardiner ES Devall MS Hamel PB Wilson AD Connor KF 2009 ). Growth and intraspecific competitive abilities of the dioecious Lindera melissifolia(Lauraceae) in varied flooding regimes. The Journal of the Torrey Botanical Society, 91 -101 . [本文引用: 1] [14] He XH Critchley C Ng H Bledsoe CS 2004 ). Reciprocal N ( 15 NH4 + or 15 NO3 -) transfer between nonN2 -fixing Eucalyptus maculata and N2 -fixing Casuarina cunninghamiana linked by the ectomycorrhizal fungus Pisolithus sp. New Phytologist, 629 -640 . [本文引用: 2] [15] Hodge A Campbell CD Fitter AH 2001 ). An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic materialNature, 413, 297 . DOI:
10.1038/35095041 [本文引用: 1] [16] Hu XW 2016 ). Effect of Arbuscular Mycorrhizal Fungi on Sex-specific Responses of Physiological and Biochemical Characteristics of Populus cathayana under Pb Pollution. Master degree dissertation, Sichuan Agricultural University, Chengdu. [本文引用: 1] [ 胡相伟 ( 2016 ). 丛枝菌根对受铅胁迫的雌雄青杨(Populus cathayana )生理生化特性的影响硕士学位论文, 四川农业大学, 成都 .] [本文引用: 1] [17] Huang KC Xu X Li XF He JD Yang YX Huan HH 2014 ). Gender-specific characteristics of tree-ring growth and differential responses to climate change in the dioecious tree Populus cathayana in Xiaowutai Mountains, China. Chinese Journal of Plant Ecology, 270 -280 . [本文引用: 1] [ 黄科朝 , 胥晓 , 李霄峰 , 贺俊东 , 杨延霞 , 郇慧慧 ( 2014 ). 小五台山青杨雌雄植株树轮生长特性及其对气候变化的响应差异植物生态学报 , 38, 270 -280 .] [本文引用: 1] [18] Ji CL Tian MM Ma JF Jin HR 2010 ). Advances in the researches on the effects of arbuscular mycorrhizal fungi on plant nutrition metabolism and growth effectsJournal of Zhejiang Normal University (Natural Sciences )303 -309 . [本文引用: 1] [ 吉春龙 , 田萌萌 , 马继芳 , 金海如 ( 2010 ). 丛枝菌根真菌对植物营养代谢与生长影响的研究进展浙江师范大学学报(自然科学版) , 33, 303 -309 .] [本文引用: 1] [19] Johansen A Jensen ES 1996 ). Transfer of N and P from intact or decomposing roots of pea to barley interconnected by an arbuscular mycorrhizal fungusSoil Biology & Biochemistry, 28, 73 -81 . [本文引用: 1] [20] Kong PP 2011 ). Effects of Arbuscular Mycorrhizal Fungi on Growth and Temperature Tolerance of Rosa hybrid and Chrysanthemum morifolium. Master degree dissertation, Academy of Agricultural Sciences, Beijing. [本文引用: 1] [ 孔佩佩 ( 2011 ). 丛枝菌根真菌对切花月季和切花菊生长及温度胁迫耐受性的影响硕士学位论文, 中国农业科学院, 北京 .] [本文引用: 1] [21] Krishna H Singh SK Sharma RR Khawale RN Grover M Patel VB 2005 ). Biochemical changes in micropropagated grape (Vitis vinifera L.) plantlets due to arbuscular-mycorrhizal fungi (AMF) inoculation during ex vitro acclimatization. Scientia Horticulturae, 554 -567 . [本文引用: 1] [22] Lehmann A Rillig MC 2015 ). Arbuscular mycorrhizal contribution to copper, manganese and iron nutrient concentrations in crops—A meta-analysisSoil Biology & Biochemistry, 81, 147 -158 . [本文引用: 1] [23] Leigh J Hodge A Fitter AH 2009 ). Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic materialNew Phytologist, 181, 199 -207 . DOI:
10.1111/j.1469-8137.2008.02630.x URL [本文引用: 1] [24] Li Z 2017 ). Influence of Arbuscular Mycorrhiza Fungi (AMF) Inoculation on Drought Tolerance of Populus cathayana Rehder Males and Females. PhD dissertation, Northwest A&F University, Yangling, Shaanxi. [本文引用: 1] [ 李朕 ( 2017 ). 丛枝菌根真菌(AMF)提高青杨雌雄株抗旱性的研究博士学位论文, 西北农林科技大学, 陕西杨凌 .] [本文引用: 1] [25] Liu T Tang M 2014 ). Effects of arbuscular mycorrhizal fungi on growth and anatomical properties of stomata andxylem in poplarsChinese Journal of Plant Ecology, 38, 1001 -1007 . [本文引用: 1] [ 刘婷 , 唐明 ( 2014 ). 丛枝菌根真菌对杨树生长、气孔和木质部微观结构的影响植物生态学报 , 38, 1001 -1007 .] [本文引用: 1] [26] Liu XJ Zeng M Du JB Ren Z 2006 ). Effects of AMF on the growth and mineral nutrition of grape cuttingsJournal of Southwest Agricultural University (Natural Science )286 -289 . [本文引用: 1] [ 刘晓捷 , 曾明 , 杜建斌 , 任争 ( 2006 ). AMF对葡萄扦插苗矿质营养及生长的影响西南大学学报(自然科学版) , 28, 286 -289 .] [本文引用: 1] [27] Luo P Zhou SL 2007 ). Effect of land use on ecological benefit of farm belt in suburbsJournal of Ecology and Rural Environment, 23(4 ), 6 -10 . Magsci [本文引用: 1] 借鉴生态效益综合评价的有关理论和数学模型,以四川省南充市高坪区为例,选取生物多样性指数、植被覆盖指数、水土保持效益指数、土壤改良指数、涵养水源指数、拦截径流指数和湿度指数7个指标作为评价因子,建立生态效益评价体系,对该区域1996和2005年2个时期综合生态效益进行评估和比较。结果表明,该区域综合生态效益值分别为7.85和7.95,2005年比1996年略为升高;基于G IS的图层叠置、分割等分析功能,就区域土地利用变化对其内部生态效益造成的变化进行了分析,该区域可被划分为生态效益显著增强区、增强区、保持区、减弱区和显著减弱区5个等级。
[ 罗培 , 周申立 ( 2007 ). 土地利用变化对城郊农业区生态效益的影响——以四川省南充市高坪区为例生态与农村环境学报 , 23(4 ), 6 -10 .] Magsci [本文引用: 1] 借鉴生态效益综合评价的有关理论和数学模型,以四川省南充市高坪区为例,选取生物多样性指数、植被覆盖指数、水土保持效益指数、土壤改良指数、涵养水源指数、拦截径流指数和湿度指数7个指标作为评价因子,建立生态效益评价体系,对该区域1996和2005年2个时期综合生态效益进行评估和比较。结果表明,该区域综合生态效益值分别为7.85和7.95,2005年比1996年略为升高;基于G IS的图层叠置、分割等分析功能,就区域土地利用变化对其内部生态效益造成的变化进行了分析,该区域可被划分为生态效益显著增强区、增强区、保持区、减弱区和显著减弱区5个等级。
[28] McGonigle TP Miller MH Evans DG Fairchild GL Swan JA 1990 ). A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungiNew Phytologist, 115, 495 -501 . DOI:
10.1111/nph.1990.115.issue-3 URL [29] Renner SS 2014 ). The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online databaseAmerican Journal of Botany, 101, 1588 -1596 . DOI:
10.3732/ajb.1400196 URL [本文引用: 2] [30] Rogers SR Eppley SM 2012 ). Testing the interaction between inter-sexual competition and phosphorus availability in a dioecious grassBotany, 90, 704 -710 . DOI:
10.1139/b2012-042 URL [本文引用: 1] [31] Sharif M Claassen N 2011 ). Action mechanisms of arbuscular mycorrhizal fungi in phosphorus uptake by Capsicum annuum L. Pedosphere, 502 -511 . [本文引用: 1] [32] Smith SE Read DJ 2010 ). Mycorrhizal Symbiosis . Academic Press, London. [本文引用: 1] [33] Smith SE Smith FA Jakobsen I 2003 ). Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responsesPlant Physiology, 133, 16 -20 . DOI:
10.1104/pp.103.024380 URL [本文引用: 1] [34] Song FQ 2002 ). Ecological and Physiological Research on VA Mycorrhizas of Populus ussuriensis PhD dissertation, Northeast Forestry University, Harbin. [本文引用: 2] [ 宋福强 ( 2002 ). 大青杨VA菌根生理生态学研究博士学位论文, 东北林业大学, 哈尔滨 .] [本文引用: 2] [35] van der Heijden MGA Boller T Wiemken A Sanders IR 1998 ). Different arbuscular mycorrhizal fungal species are potential determinants of plant community structureEcology, 79, 2082 -2091 . DOI:
10.1890/0012-9658(1998)079[2082:DAMFSA]2.0.CO;2 URL [本文引用: 1] [36] Vierheilig H Schweiger P Brundrett M 2010 ). An overview of methods for the detection and observation of arbuscular mycorrhizal fungi in rootsPhysiologia Plantarum, 125, 393 -404 . [本文引用: 1] [37] Wang ZF Xu X Li XF Yang P Yuan XL 2011 ). The distribution of male and female Populus cathayana populations along an altitudinal gradient. Acta Ecologica Sinica, 7067 -7074 . [本文引用: 1] [ 王志峰 , 胥晓 , 李霄峰 , 杨鹏 , 袁新利 ( 2011 ). 青杨雌雄群体沿海拔梯度的分布特征生态学报 , 31, 7067 -7074 .] [本文引用: 1] [38] Wu N Li Z Liu H Tang M 2015 ). Influence of arbuscular mycorrhiza on photosynthesis and water status of Populus cathayana Rehder males and females under salt stress. Acta Physiologiae Plantarum, 183 . DOI: 10.1007/?s11738-015-1932-6 . [本文引用: 1] [39] Wu N Li Z Wu F Tang M 2016 ). Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Scientific Reports, 37663 . DOI: 10.1038/srep37663 . [本文引用: 1] [40] Wu QP Tang Y Dong TF Liao YM Li DD He XH Xu X 2018 ). Additional AM fungi inoculation increase Populus cathayana intersexual competition. Frontiers in Plant Science, 607 . DOI: 10.3389/fpls.2018.00607 . [本文引用: 1] [41] Wu QS Yuan FY Fei YJ Li L Huang YM Liu CY 2014 ). Effects of arbuscular mycorrhizal fungi on root system architecture and sugar contents of white cloverActa Prataculturae Sinica, 23, 199 -204 . [本文引用: 1] [ 吴强盛 , 袁芳英 , 费永俊 , 李莉 , 黄咏明 , 刘春艳 ( 2014 ). 丛枝菌根真菌对白三叶根系构型和糖含量的影响草业学报 , 23, 199 -204 .] [本文引用: 1] [42] Xu X 2016 ). Research advances in root recognition in plantsJournal of China West Normal University (Natural Sciences )365 -369 . [本文引用: 2] [ 胥晓 ( 2016 ). 植物根系识别研究西华师范大学学报(自然科学版) , 37, 365 -369 .] [本文引用: 2] [43] Xu X Peng G Wu C Korpelainen H Li CY 2008 ). Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana . Tree Physiology 1751 -1759 . [本文引用: 1] [44] Yang P Xu X 2012 ). Effects of waterlogging stress on the growth and physiological characteristics of male and female Populus cathayana seedlings. Chinese Journal of Plant Ecology, 81 -87 . [本文引用: 1] [ 杨鹏 , 胥晓 ( 2012 ). 淹水胁迫对青杨雌雄幼苗生理特性和生长的影响植物生态学报 , 36, 81 -87 .] [本文引用: 1] [45] Zeng Z 2016 ). Effect of Opposite Sexual Neighbor on the Root Growth, Component of Root Exudates of Female and Male Populus cathayana Rehd. Seedlings. Master degree dissertation, China West Normal University, Nanchong, Sichuan. [本文引用: 2] [ 曾贞 ( 2016 ). 异性邻株对青杨雌雄植株根的生长及其分泌物成分的影响硕士学位论文, 西华师范大学, 四川南充 .] [本文引用: 2] [46] Zhang YM Ma KM Qu LY 2017 ). Inoculation with arbuscular mycorrhizal fungi enhances the root system of Bauhinia faberi var. microphylla seedlings under drought stress conditions. Acta Ecologica Sinica, 2611 -2619 . [本文引用: 1] [ 张亚敏 , 马克明 , 曲来叶 ( 2017 ). 干旱条件下接种AM真菌对小马鞍羊蹄甲幼苗根系的影响生态学报 , 37, 2611 -2619 .] [本文引用: 1] [47] Zhao QH Sun LT Wang Y Ding ZT Li M 2014 ). Effects of arbuscular mycorrhizal fungi and nitrogen regimes on plant growth, nutrient uptake and tea quality in Camellia sinensis(L.) O. Kuntze. Plant Physiology Journal, 164 -170 . [本文引用: 1] [ 赵青华 , 孙立涛 , 王玉 , 丁兆堂 , 李敏 ( 2014 ). 丛枝菌根真菌和施氮量对茶树生长、矿质元素吸收与茶叶品质的影响植物生理学报 , 50, 164 -170 .] [本文引用: 1] [48] Zhao X Yan XF 2006 ). Effects of arbuscular mycorrhizal fungi on the growth and absorption of nitrogen and phosphorus in Camptotheca acuminata seedlings. Journal of Plant Ecology (Chinese Version )947 -953 . [本文引用: 1] [ 赵昕 , 阎秀峰 ( 2006 ). 丛枝菌根对喜树幼苗生长和氮、磷吸收的影响植物生态学报 , 30, 947 -953 .] [本文引用: 1] [49] Zhu J Liu G Xiao J Zhu SH Zeng Z Xu X Dong TF 2016 ). Biomass of mulberry under planting modes of different gender combinationsChinese Journal of Ecology, 35, 2336 -2340 . [本文引用: 2] [ 朱娟 , 刘刚 , 肖娟 , 竺诗慧 , 曾贞 , 胥晓 , 董廷发 ( 2016 ). 桑树不同性别组合种植模式下的生物量生态学杂志 , 35, 2336 -2340 .] [本文引用: 2] [50] Zhu SH Dong TF Liu G Xiao J Zhu J Zeng Z Chen DF Xu X 2016 ). Effects of root exudates on the growth and development of male and female Morus alba seedlings. Plant Physiology Journal, 134 -140 . [本文引用: 3] [ 竺诗慧 , 董廷发 , 刘刚 , 肖娟 , 朱娟 , 曾贞 , 陈德甫 , 胥晓 ( 2016 ). 桑树(Morus alba )幼苗根系分泌物对雌雄植株生长发育的影响植物生理学报 , 52, 134 -140 .] [本文引用: 3] [51] Zou YN Wu QS Li Y Huang YM 2014 ). Effects of arbuscular mycorrhizal fungi on root system morphology and sucrose and glucose contents of Poncirus trifoliata . Chinese Journal of Applied Ecology, 1125 -1129 . [本文引用: 1] [ 邹英宁 , 吴强盛 , 李艳 , 黄咏明 ( 2014 ). 丛枝菌根真菌对枳根系形态和蔗糖、葡萄糖含量的影响应用生态学报 , 25, 1125 -1129 .] [本文引用: 1] Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis
1
2010
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudates
1
2000
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
Sexual competition and N supply interactively affect the dimorphism and competiveness of opposite sexes in
Populus cathayana .
2
2015
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
Intra- and inter-sexual competition of
Populus cathayana under different watering regimes.
1
2014
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
Sex-specific competition differently regulates ecophysiological responses and phytoremediation of
Populus cathayana under Pb stress.
1
2017
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in
Populus cathayana .
2
2015
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
接种AM真菌对短命植物生长发育及矿质养分吸收的影响
1
2008
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
接种AM真菌对短命植物生长发育及矿质养分吸收的影响
1
2008
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
Gender-specific physiology, carbon isotope discrimination, and habitat distribution in boxelder,Acer negundo.
1
1993
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
Root-?mediated sex recognition in a dioecious tree
1
2017
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
Patterns of below-ground plant interconnections established by means of arbuscular mycorrhizal networks
1
2004
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
Membrane-?mediated decrease in root exudation responsible for phorphorus inhibition of vesicular-arbuscular mycorrhiza formation
1
1981
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
丛枝菌根真菌对双子叶植物生长和根系特征的影响: 整合分析
1
2017
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
丛枝菌根真菌对双子叶植物生长和根系特征的影响: 整合分析
1
2017
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
Growth and intraspecific competitive abilities of the dioecious Lindera melissifolia(Lauraceae) in varied flooding regimes.
1
2009
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
Reciprocal N (
15 NH
4 + or
15 NO
3 -) transfer between nonN
2 -fixing
Eucalyptus maculata and N
2 -fixing
Casuarina cunninghamiana linked by the ectomycorrhizal fungus
Pisolithus sp.
2
2004
... 实验地点位于四川省南充市西华师范大学试验地(106.07° E, 30.80° N, 海拔276 m).该地区属亚热带季风性湿润气候, 年平均气温、降水量、日照时间分别为16.8 ℃、1 065 mm、1 980 h (
罗培和周申立, 2007 ).2017年3月从试验地的青杨成树(引种自青海大通)上采集一年生枝条进行扦插(雌雄各50根扦插条).扦插所用的基质为泥土、河沙、蛭石体积比1:1:1的混合物, 于100 ℃蒸汽灭菌2 h两次以消除AMF的繁殖体和其他微生物.在扦插之前将枝条用70%酒精消毒, 并用蒸馏水漂洗, 最后放入生根粉溶液中浸泡约15 s后进行扦插.待扦插条发芽生长60天后, 选取长短、粗细、健康状况一致的84株幼苗(雌雄各42株)移栽到长宽高分别为40 cm × 20 cm × 30 cm的玻璃缸中(分别按雄-雄, 雌-雌, 雄-雌组合栽培).实验中的玻璃缸内部装置与
Johansen和Jensen (1996) 的类似.在玻璃缸中间放置两块厚0.25 cm的多孔有机玻璃板和一张单个孔径为37 μm的尼龙网(尼龙网放置在两个玻璃板之间)将容器分割为两个室.该结构的目的是防止根系穿过, 但允许菌丝相连(参考
He et al ., 2004 )(
图1 ). ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material
1
2001
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
丛枝菌根对受铅胁迫的雌雄青杨(
Populus cathayana )生理生化特性的影响
1
2016
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
丛枝菌根对受铅胁迫的雌雄青杨(
Populus cathayana )生理生化特性的影响
1
2016
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
小五台山青杨雌雄植株树轮生长特性及其对气候变化的响应差异
1
2014
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
小五台山青杨雌雄植株树轮生长特性及其对气候变化的响应差异
1
2014
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
丛枝菌根真菌对植物营养代谢与生长影响的研究进展
1
2010
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
丛枝菌根真菌对植物营养代谢与生长影响的研究进展
1
2010
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
Transfer of N and P from intact or decomposing roots of pea to barley interconnected by an arbuscular mycorrhizal fungus
1
1996
... 实验地点位于四川省南充市西华师范大学试验地(106.07° E, 30.80° N, 海拔276 m).该地区属亚热带季风性湿润气候, 年平均气温、降水量、日照时间分别为16.8 ℃、1 065 mm、1 980 h (
罗培和周申立, 2007 ).2017年3月从试验地的青杨成树(引种自青海大通)上采集一年生枝条进行扦插(雌雄各50根扦插条).扦插所用的基质为泥土、河沙、蛭石体积比1:1:1的混合物, 于100 ℃蒸汽灭菌2 h两次以消除AMF的繁殖体和其他微生物.在扦插之前将枝条用70%酒精消毒, 并用蒸馏水漂洗, 最后放入生根粉溶液中浸泡约15 s后进行扦插.待扦插条发芽生长60天后, 选取长短、粗细、健康状况一致的84株幼苗(雌雄各42株)移栽到长宽高分别为40 cm × 20 cm × 30 cm的玻璃缸中(分别按雄-雄, 雌-雌, 雄-雌组合栽培).实验中的玻璃缸内部装置与
Johansen和Jensen (1996) 的类似.在玻璃缸中间放置两块厚0.25 cm的多孔有机玻璃板和一张单个孔径为37 μm的尼龙网(尼龙网放置在两个玻璃板之间)将容器分割为两个室.该结构的目的是防止根系穿过, 但允许菌丝相连(参考
He et al ., 2004 )(
图1 ). ...
丛枝菌根真菌对切花月季和切花菊生长及温度胁迫耐受性的影响
1
2011
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌对切花月季和切花菊生长及温度胁迫耐受性的影响
1
2011
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
Biochemical changes in micropropagated grape (
Vitis vinifera L.) plantlets due to arbuscular-mycorrhizal fungi (AMF) inoculation during ex vitro acclimatization.
1
2005
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
Arbuscular mycorrhizal contribution to copper, manganese and iron nutrient concentrations in crops—A meta-analysis
1
2015
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material
1
2009
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
丛枝菌根真菌(AMF)提高青杨雌雄株抗旱性的研究
1
2017
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
丛枝菌根真菌(AMF)提高青杨雌雄株抗旱性的研究
1
2017
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
丛枝菌根真菌对杨树生长、气孔和木质部微观结构的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌对杨树生长、气孔和木质部微观结构的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
AMF对葡萄扦插苗矿质营养及生长的影响
1
2006
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
AMF对葡萄扦插苗矿质营养及生长的影响
1
2006
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
土地利用变化对城郊农业区生态效益的影响——以四川省南充市高坪区为例
1
2007
... 实验地点位于四川省南充市西华师范大学试验地(106.07° E, 30.80° N, 海拔276 m).该地区属亚热带季风性湿润气候, 年平均气温、降水量、日照时间分别为16.8 ℃、1 065 mm、1 980 h (
罗培和周申立, 2007 ).2017年3月从试验地的青杨成树(引种自青海大通)上采集一年生枝条进行扦插(雌雄各50根扦插条).扦插所用的基质为泥土、河沙、蛭石体积比1:1:1的混合物, 于100 ℃蒸汽灭菌2 h两次以消除AMF的繁殖体和其他微生物.在扦插之前将枝条用70%酒精消毒, 并用蒸馏水漂洗, 最后放入生根粉溶液中浸泡约15 s后进行扦插.待扦插条发芽生长60天后, 选取长短、粗细、健康状况一致的84株幼苗(雌雄各42株)移栽到长宽高分别为40 cm × 20 cm × 30 cm的玻璃缸中(分别按雄-雄, 雌-雌, 雄-雌组合栽培).实验中的玻璃缸内部装置与
Johansen和Jensen (1996) 的类似.在玻璃缸中间放置两块厚0.25 cm的多孔有机玻璃板和一张单个孔径为37 μm的尼龙网(尼龙网放置在两个玻璃板之间)将容器分割为两个室.该结构的目的是防止根系穿过, 但允许菌丝相连(参考
He et al ., 2004 )(
图1 ). ...
土地利用变化对城郊农业区生态效益的影响——以四川省南充市高坪区为例
1
2007
... 实验地点位于四川省南充市西华师范大学试验地(106.07° E, 30.80° N, 海拔276 m).该地区属亚热带季风性湿润气候, 年平均气温、降水量、日照时间分别为16.8 ℃、1 065 mm、1 980 h (
罗培和周申立, 2007 ).2017年3月从试验地的青杨成树(引种自青海大通)上采集一年生枝条进行扦插(雌雄各50根扦插条).扦插所用的基质为泥土、河沙、蛭石体积比1:1:1的混合物, 于100 ℃蒸汽灭菌2 h两次以消除AMF的繁殖体和其他微生物.在扦插之前将枝条用70%酒精消毒, 并用蒸馏水漂洗, 最后放入生根粉溶液中浸泡约15 s后进行扦插.待扦插条发芽生长60天后, 选取长短、粗细、健康状况一致的84株幼苗(雌雄各42株)移栽到长宽高分别为40 cm × 20 cm × 30 cm的玻璃缸中(分别按雄-雄, 雌-雌, 雄-雌组合栽培).实验中的玻璃缸内部装置与
Johansen和Jensen (1996) 的类似.在玻璃缸中间放置两块厚0.25 cm的多孔有机玻璃板和一张单个孔径为37 μm的尼龙网(尼龙网放置在两个玻璃板之间)将容器分割为两个室.该结构的目的是防止根系穿过, 但允许菌丝相连(参考
He et al ., 2004 )(
图1 ). ...
A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi
0
1990
The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database
2
2014
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
Testing the interaction between inter-sexual competition and phosphorus availability in a dioecious grass
1
2012
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
Action mechanisms of arbuscular mycorrhizal fungi in phosphorus uptake by
Capsicum annuum L.
1
2011
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
1
2010
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses
1
2003
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
大青杨VA菌根生理生态学研究
2
2002
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
大青杨VA菌根生理生态学研究
2
2002
... 青杨属于杨柳科杨属(
Populus ), 是我国常见的雌雄异株植物.该树种由于具有生长快、存活率高、适应性强等特点, 是近年来雌雄异株植物研究的常用材料, 也是AMF营养型树种(
宋福强, 2002 ).目前, 关于接种AMF对青杨的影响主要集中在抗旱性(
李朕, 2017 )和对重金属的吸附能力(
胡相伟, 2016 )方面, 而关于接种AMF对不同性别组合栽培模式下雌雄植株根系影响的研究未见报道.因此, 本研究以青杨扦插苗为试验材料, 分别在雄-雄、雌-雌和雄-雌3种组合的栽培模式下对青杨雌雄幼苗进行接种和不接种摩西球囊霉(
Funneliformis mosseae )处理, 通过比较接种AMF与否雌雄植株根系的侵染率、生物量、形态、碳氮含量等在不同性别组合栽模式下的差异, 揭示了AMF对不同性别组合栽培模式下的青杨根系发育的影响.研究结果可为青杨的科学栽培提供理论参考. ...
... 此外, 本研究还发现接种AMF后青杨雌雄植株根系的生物量和总根长显著增加, 这与
宋福强(2002) 对大青杨(
P .
ussuriensis )的研究结果吻合, 二者都证实了接种AMF可以促进杨属植物根系的生长发育.AMF促进植物根系生长的原因主要是增加了寄主植物对磷的吸收(
郭晗铃等, 2017 ), 但近年也发现AMF可以从土壤有机物(如叶凋落物)中获取氮并转移到寄主植物根中(
Hodge et al ., 2001 ;
Leigh et al ., 2009 ), 提高植物利用氮的能力并促进光合作用, 从而有利于植物的生长(
Krishna et al ., 2005 ).与此同时, 植株光合速率与光合能力的提高可以使根系间接获得更多的可溶性糖或淀粉等碳水化合物, 从而导致根系中碳含量的增加(
吉春龙等, 2010 ).与上述结果相一致, 我们也发现了接种AMF对3种栽培模式下青杨雌雄植株根系的碳、氮含量具有一定的促进作用.然而, 由于实验设计的缺陷, 我们未考虑接种AMF前后雌雄植株根系中磷含量的变化, 故无法在磷吸收方面提供更多的证据, 还需要在下一步研究工作中予以验证. ...
Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure
1
1998
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
An overview of methods for the detection and observation of arbuscular mycorrhizal fungi in roots
1
2010
... 将每种组合下的植株(7个重复)根系分别洗净后保存在FAA固定液中, 固定完成后每个植物根系样品选择至少100段直径小于1 mm, 长度约1 cm的新鲜根段放在10%的KOH溶液中90 ℃水浴加热1 h, 待加热结束后放入碱性双氧水中进行脱色, 而后取出根系用墨水和醋(95%醋和5%墨水)进行染色, 随后用酸化的水冲洗根段数次(
Vierheilig et al ., 2010 ), 最后随机选取70个根段整齐排列在载玻片上用显微镜进行观测.根据McGonigle等(1990)的方法用以下公式计算菌根侵染率: ...
青杨雌雄群体沿海拔梯度的分布特征
1
2011
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
青杨雌雄群体沿海拔梯度的分布特征
1
2011
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
Influence of arbuscular mycorrhiza on photosynthesis and water status of
Populus cathayana Rehder males and females under salt stress.
1
2015
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
Comparative photochemistry activity and antioxidant responses in male and female
Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt.
1
2016
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
Additional AM fungi inoculation increase
Populus cathayana intersexual competition.
1
2018
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
丛枝菌根真菌对白三叶根系构型和糖含量的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌对白三叶根系构型和糖含量的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
植物根系识别研究
2
2016
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
植物根系识别研究
2
2016
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
Drought inhibits photosynthetic capacity more in females than in males of
Populus cathayana .
1
2008
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
淹水胁迫对青杨雌雄幼苗生理特性和生长的影响
1
2012
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
淹水胁迫对青杨雌雄幼苗生理特性和生长的影响
1
2012
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
异性邻株对青杨雌雄植株根的生长及其分泌物成分的影响
2
2016
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
异性邻株对青杨雌雄植株根的生长及其分泌物成分的影响
2
2016
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
干旱条件下接种AM真菌对小马鞍羊蹄甲幼苗根系的影响
1
2017
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
干旱条件下接种AM真菌对小马鞍羊蹄甲幼苗根系的影响
1
2017
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌和施氮量对茶树生长、矿质元素吸收与茶叶品质的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌和施氮量对茶树生长、矿质元素吸收与茶叶品质的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根对喜树幼苗生长和氮、磷吸收的影响
1
2006
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根对喜树幼苗生长和氮、磷吸收的影响
1
2006
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
桑树不同性别组合种植模式下的生物量
2
2016
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
桑树不同性别组合种植模式下的生物量
2
2016
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
桑树(
Morus alba )幼苗根系分泌物对雌雄植株生长发育的影响
3
2016
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
桑树(
Morus alba )幼苗根系分泌物对雌雄植株生长发育的影响
3
2016
... 相关研究表明, 在30多万种被子植物中还存在约6%的雌雄异株植物(
Renner, 2014 ), 由于该类植物对自然界长期适应性进化以及雌雄个体对资源的需求和分配的不同, 从而导致雌雄植株对环境的适应以及个体间的竞争和合作能力出现差异(
Xu et al ., 2008 ;
杨鹏和胥晓, 2012 ;
黄科朝等, 2014 ).如
Lindera melissifolia 在雄-雌合栽时具有相同的竞争能力(
Hawkins et al ., 2009 ),
Distichlis spicata 在同性合栽时地下竞争能力更弱, 而桑(
Morus alba )在雄-雄合栽时的竞争能力更强(
Rogers & Eppley, 2012 ;
朱娟等, 2016 ), 青杨(
Populus cathayana )雌株在水分充足时竞争能力比雄株强而在干旱胁迫下则相反(
Chen et al ., 2014 ).此外, 低氮环境下青杨在雄-雌合栽时的雌株比同性合栽下的雌株具有更高的根干质量(
Chen et al ., 2015a ), 桑在雄-雌合栽模式下相互促进并具有较高的总生物量(
朱娟等, 2016 ;
竺诗慧等, 2016 )等.虽然上述研究证实了雌雄异株植物个体在不同雌雄组合模式下表现出了不一致的竞争或互利现象, 但均未深入揭示其引起差异的原因.由于自然界中的雌雄异株植物种群的性别比例总体趋近于1:1 (
王志锋等, 2011 ), 雌雄个体可以通过根系间的AMF相互作用(
Bonfante & Genre, 2010 ;
Wu et al ., 2018 ).因此, 我们推测AMF可能是引起不同性别组合模式下雌雄植株生长出现差异的原因之一.然而, 相关的研究并未见报道. ...
... 前人的研究证实了接种AMF会提高植株根系的菌根侵染率, 但很少有人关注接种AMF后雌雄植株的侵染率差异问题.尽管由于接种剂的类型或剂量的原因, 本研究中青杨雌雄植株的侵染率与
Chen等(2015b) 和
Wu等(2016) 的研究结果相比偏低, 但研究结果表明接种AMF不仅显著增加了青杨雌雄植株根系的侵染率, 而且在雄-雌合栽模式下青杨雌株的侵染率显著高于雄株(雌株的侵染率为43%-46%, 雄株的侵染率为39%-42%).由于性别二态性原因, 雌雄植株在资源需求、生理过程(如气体交换、水分利用和元素吸收等)、形态发育、生物量积累、抗逆能力以及分布格局等方面明显不同(
Dawson & Ehleringer, 1993 ;
Renner, 2014 ;
Wu et al ., 2015 ), 这种差异甚至体现在不同性别合栽模式下雌雄植株对邻株根系的性别识别以及引起根系分泌物的变化(
胥晓, 2016 ;
曾贞, 2016 ;
竺诗慧等, 2016 )上.根据
Graham等(1981) 和
Buee等(2000) 的研究, 根系分泌物的成分和含量差异会影响AMF对寄主植物的侵染程度.因此, 雄-雌合栽模式下青杨雌雄植株的侵染率出现差异极可能与雌雄互作后引起根系分泌物的差异有很大的关系. ...
... 另一方面, 与
Chen等(2015) 的研究结果类似, 我们也发现青杨雌雄植株的根质量、根形态以及氮含量与栽培模式关系密切, 而且还受到AMF的影响.如异性合栽模式下雌株的根干质量、比表面积、比根长、总根长和氮含量均显著大于雄株, 反映了雌株在异性合栽模式下生长更好.其原因可能在于: 一是植株的根系在生长过程中具有自我识别和性别识别能力, 它们能通过调节自身根系的生长发育去适应周围其他植株的根系(
胥晓, 2016 ), 青杨植株在同性组合时会改变根系的生长, 但在异性组合时却无该现象(
Dong et al ., 2017 ).二是植株的根系分泌物与植株的根系发育密切相关(
曾贞, 2016 ), 异性植株根系分泌物对雌株的株高、基径、叶面积和根、茎、叶生物量等具有显著的促进作用(
竺诗慧等, 2016 ).三是不同性别的植株因对资源的需求不同而可能存在互助作用.AMF寄主特异性很低, 根外菌丝可以成为植株间物质交换的桥梁(
Giovannetti et al. , 2004 ;
He et al ., 2004 ), 在雄-雌合栽模式下的雄株极有可能将自身的氮素通过菌丝转移给资源需求更高的雌株, 使得雌株和雄株合栽后更有利于雌株根系的发育, 以维持种群的延续.本实验中根系的氮含量与
Chen等(2017) 的研究结果一致, 都表明在雄-雌合栽模式下的雌株氮含量高于雌-雌合栽模式, 雄株的氮含量则相反.这虽然验证了我们的假设, 但仍需开展进一步的研究. ...
丛枝菌根真菌对枳根系形态和蔗糖、葡萄糖含量的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
丛枝菌根真菌对枳根系形态和蔗糖、葡萄糖含量的影响
1
2014
... 丛枝菌根真菌(AMF)是自然生态系统中广泛存在且能与绝大多数高等植物的根系形成互惠共生体的土壤真菌(
van der Heijden et al ., 1998 ).它可以扩大宿主植物根系在土壤中的有效吸收范围, 促进宿主植物对碳、氮、磷、硫、铁、锌、铜、锰、钾等元素的吸收(
赵昕和阎秀峰, 2006 ;
Sharif & Claassen, 2011 ;
Lehmann & Rillig, 2015 ), 提高水分利用率(
Smith & Read, 2010 ), 提高植株光合效率以及调节叶片气孔开闭(
刘婷和唐明, 2014 )等, 促进植物生长, 增加作物产量(
刘晓捷等, 2006 ), 改善作物产品品质(
赵青华等, 2014 ), 增强植物抗病能力(
Smith et al ., 2003 )和抗逆性(
陈志超等, 2008 ;
Chen et al. , 2015b )等.随着研究的不断深入, AMF在根系生长发育中所扮演的角色逐渐引起人们的重视.大量的文献表明接种AMF能够促进宿主植物根系的生长发育, 如接种AMF增加了枳(
Poncirus trifoliata )的根系投影面积、表面积、体积和总长度(
邹英宁等, 2014 ); 干旱胁迫下接种AMF显著增加了小鞍叶羊蹄甲(
Bauhinia faberi var.
microphylla )幼苗的总根长、根表面积、根分枝数和根尖数(
张亚敏等, 2017 ); 常温条件下接种AMF能增加丰水月季(
Rosa hybrida )和菊花(
Chrysanthemum morifolium )的根系长度(
孔佩佩, 2011 );白车轴草(
Trifolium repens )在接种沾屑多样孢囊霉(
Diversispora spurca )、地表球囊霉(
Glomus versiforme )和隐类球囊霉(
Paraglomus occultum )后植株的根干质量、长度、投影面积、表面积、体积、根尖数、分枝数和交叉数均得到提高(
吴强盛等, 2014 ).然而, 上述研究多集中在雌雄同株植物中而很少涉及雌雄异株植物. ...
 ,1,2,*
,1,2,*
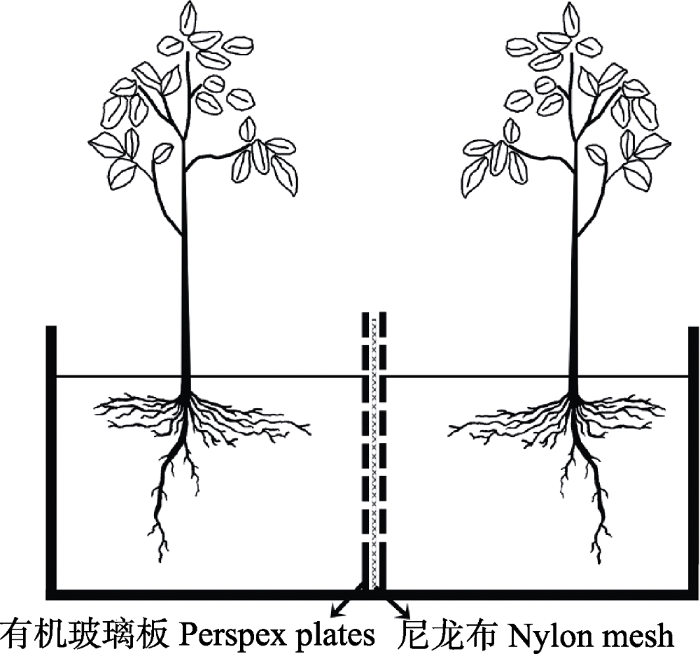 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT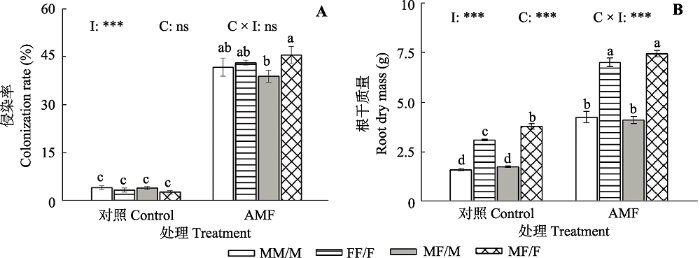 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT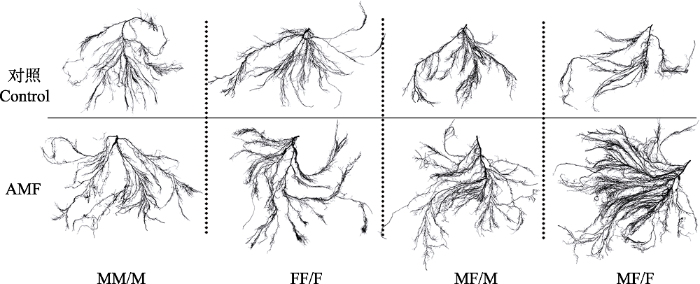 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT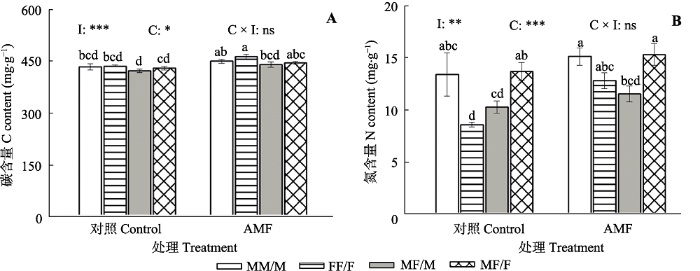 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}