 ,1, 段世华2, 安婷1, 康华靖,
,1, 段世华2, 安婷1, 康华靖, ,3,*
,3,*Construction of CO2-response model of electron transport rate in C4 crop and its application
YE Zi-Piao,1, DUAN Shi-Hua2, AN Ting1, KANG Hua-Jing,,3,*通讯作者: *康华靖 ORCID: 0000-0003-3808-3115kanghuajing@126.com; 叶子飘ORCID: 0000-0002-7598-1822
编委: 杜盛
责任编辑: 李敏
收稿日期:2018-05-29网络出版日期:2018-10-20
| 基金资助: |
Received:2018-05-29Online:2018-10-20
| Fund supported: |

摘要
准确估算光合电子流对CO2响应的变化趋势对深入了解光合过程具有重要意义。该研究在植物光合作用对CO2响应新模型(模型I)的基础上构建了电子传递速率(J)对CO2的响应模型(模型II), 并对用LI-6400-40便携式光合仪测量的玉米(Zea mays)和千穗谷(Amaranthus hypochondriacus)的数据进行了拟合。结果表明, 模型II可以很好地拟合玉米和千穗谷叶片J对CO2浓度的响应曲线(J-Ca曲线), 得到玉米和千穗谷的最大电子传递速率分别为262.41和393.07 mmol·m -2·s -1, 与估算值相符合。在此基础上, 对光合电子流分配到其他路径进行了探讨。结果显示, 380 mmol·mol -1 CO2浓度下玉米和千穗谷碳同化所需的电子流为247.92和285.16 mmol·m -2·s -1, 分配到其他途径的光合电子流为14.49和107.91 mmol·m -2·s -1(考虑植物CO2的回收利用)。比较两种植物的其他途径光合电子流分配值发现, 两者相差6倍之多。分析认为这与千穗谷和玉米的催化脱羧反应酶种类以及脱羧反应发生的部位不同密切相关。该发现为人们研究C4植物中烟酰胺腺嘌呤二核苷磷酸苹果酸酶型和烟酰胺腺嘌呤二核苷酸苹果酸酶型两种亚型之间的差异提供了一个新的视角。此外, 构建的电子传递速率对CO2的响应模型为人们研究C4植物的光合电子流的变化规律提供了一个可供选择的数学工具。
关键词:
Abstract
Aims Accurate estimation of variation tendency of photosynthetic electron flow response to CO2 is of great significance to understand the photosynthetic processes.
Methods A model of electron transport rate (J) response to CO2 (model II) was developed based on a new model of photosynthesis response to CO2 (model I). The data of maize (Zea mays) and grain amaranth (Amaranthus hypochondriacus) that were measured by LI-6400-40 portable photosynthetic apparatus were fitted by the two models, respectively.
Important findings The results indicated that the model II could well characterize and fit the CO2-response curves of electron transport rate (J-Ca curve) for maize and grain amaranth, and the maximum electron transport rates of maize and grain amaranth were 262.41 and 393.07 mmol·m -2·s -1, which were in very close agreement with the estimated values (p > 0.05), respectively. Based on these results, the allocation to other pathways of photosynthetic electronic flow were discussed. At 380 mmol·mol -1 CO2, the photosynthetic electron flows for carbon assimilation of maize and grain amaranth carbon were 247.92 and 285.16 mmol·m -2·s -1, respectively, when the CO2 for recovery of mitochondrial respiration was considered, and the photosynthetic electron flows for other pathways were 14.49 and 107.91 mmol·m -2·s -1, respectively. The photosynthetic electron flows for other pathways in grain amaranth were more six times than that in maize. The analysis shows that this difference is closely related to the types of catalytic decarboxylase and the locations of decarboxylation reactions. This finding provides a new perspective for investigating the differences between the two subtypes of nicotinamide adenine dinucleotide phosphate malic acid enzyme type and nicotinamide adenine dinucleotide malic acid enzyme type in C4 species. In addition, the CO2-response model of electron transport rate offers us an alternative mathematical tool for investigating the photosynthetic electron flow of C4 crop.
Keywords:
PDF (1041KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
叶子飘, 段世华, 安婷, 康华靖. C4作物电子传递速率对CO2响应模型的构建及应用. 植物生态学报[J], 2018, 42(10): 1000-1008 DOI:10.17521/cjpe.2018.0129
YE Zi-Piao, DUAN Shi-Hua, AN Ting, KANG Hua-Jing.
光合作用是植物(包括光合细菌)利用光能, 将CO2或H2S合成有机物并释放O2的过程。当光照射植物叶片时, 光能被捕光色素分子吸收后由基态跃迁到激发态, 处于激发态上的激子有3条退激发途径(Baker, 2008), 其中大部分激子是经共振方式传递到光系统II (PSII), 由此产生电荷分离, 形成光合电子流。这些光合电子流传给叶绿体中的电子受体脱镁叶绿素a分子(Phe-), 然后依次经过细胞色素b6f蛋白复合体(Cytb6f)和光系统I (PSI), 最后传递给烟酰胺腺嘌呤二核苷磷酸(NADP)形成NADPH。同时, 在电子的传递过程中产生的跨膜质子梯度驱动三磷酸腺苷(ATP)合酶, 形成ATP (薛娴等, 2017)。由此可知, 定量分析电子传递速率(J)对光和对CO2的响应是研究植物的光合能力大小和光合特性的基础。
当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型)。该模型经过30多年的发展, 已经成为被广泛应用的模型(Farquhar & Busch, 2017; 梁星云和刘世荣, 2017; Silva-Pérez et al., 2017; 唐星林等, 2017a, 2017b; Walker et al., 2017)。用该模型拟合植物的Ac-C曲线可以获得描述植物光合作用的几个重要生化参数, 如最大羧化速率(Vcmax)、最大电子传递速率(Jmax)、磷酸丙糖利用速率(VTPU)等。然而, 该模型主要针对C3植物构建。C4植物是从C3植物进化来的一种高光效种类。与C3植物相比, C4植物能通过C4途径的酶系统(如磷酸烯醇式丙酮酸酶、NADP-苹果酸脱氢酶、NADP-苹果酸酶和丙酮酸磷酸二激酶等)保持较高的光合效率, 如C4植物叶肉细胞叶绿体含有磷酸烯醇式丙酮酸酶, 磷酸烯醇式丙酮酸的再生和草酰乙酸的还原均发生在叶肉细胞内; 而维管束鞘细胞含有脱羧酶和Rubisco, 苹果酸的脱羧和CO2通过Calvin循环的再固定则在鞘细胞中进行。利用空间上分隔的叶肉细胞和维管束鞘细胞(构成Kranz结构)可有效地同化外界的CO2 (Hatch, 1987; Ku et al., 1999)。由此可见, C4植物的光合过程与C3植物有较大差异。所以, FvCB生化模型不适合C4植物光合过程的描述和拟合。针对C4植物与C3植物光合过程存在较大的差异, 1978年Berry和Farquhar首次构建了C4植物的光合作用模型(Berry & Farquhar, 1978), 而后由Collatz等(1992)、von Caemmerer和Furbank (1999)进行了修改和补充。此后, von Caemmerer (2013)进一步完善了C4模型。该模型主要以NADPH和ATP的产生和消耗作为研究对象, 并不涉及植物的J对CO2的响应问题。
目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道。而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义。此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(康华靖等, 2014; 叶子飘等, 2017), 且不等于0 μmol·m-2·s-1。然而, 在这种条件下C4植物的光合电子流的作用和分配尚不清楚, 还有待于进一步的研究。为此, 本文首先在植物光合作用对CO2响应新模型(叶子飘, 2010)的基础上构建J对CO2的响应模型; 其次以玉米(Zea mays)和千穗谷(Amaranthus hypochondriacus)两种C4植物为研究对象, 利用便携式光合作用-荧光测量系统获取这2种植物的J对CO2浓度的响应曲线(J-Ca, Ca为环境CO2浓度; J-Ci曲线, Ci为胞间CO2浓度); 再次, 用所构建的J对CO2的响应模型分别拟合测量曲线, 得到Jmax和对应的饱和CO2浓度, 并与观测值进行比较从而确定所构建的模型是否合理; 最后定量分析当环境CO2浓度等于0 μmol·mol-1时玉米和千穗谷的光合电子流的大小及其分配。
1 材料和方法
1.1 模型构建
1.1.1 光合作用对CO2的响应模型本文拟构建的J对CO2的响应模型, 是建立在光合作用对CO2响应的新模型(叶子飘, 2010)(简称模型I)基础之上。模型I的具体表达式为:
式中, Ac为净光合速率, αc为光合作用对CO2响应曲线的初始斜率, βc和γc为两个与CO2浓度无关的系数, C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci), Rl为光下呼吸速率, 它与光强存在非线性响应关系(叶子飘等, 2017), 不同植物光下呼吸速率的差异性影响着系数βc和γc的值。为了简化, 本文假设Rl近似为常数。利用模型I可得到植物叶片的饱和CO2浓度(Csat, 既可以表示为大气饱和CO2浓度, 用Ca-sat表示; 也可以表示为饱和胞间CO2浓度, 用Ci-sat表示)和最大净光合速率(Ac-max): 它们分别由${{C}_{\text{sat}}}=\frac{\sqrt{{\text{(}{{\beta }_{\text{c}}}+{{\gamma }_{\text{c}}}\text{)}}/{{{\beta }_{\text{c}}}}\;}-\text{1}}{{{\gamma }_{\text{c}}}}$和${{A}_{\text{c-max}}}={{\alpha }_{\text{c}}}{{\left( \frac{\sqrt{{{\beta }_{\text{c}}}+{{\gamma }_{\text{c}}}}-\sqrt{{{\beta }_{\text{c}}}}}{{{\gamma }_{\text{c}}}} \right)}^{2}}-{{R}_{\text{l}}}$计算获得(叶子飘, 2010)。
1.1.2 电子传递速率对CO2的响应模型
由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异。其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(Eichelmann et al., 2011; Heber, 2002; Miyake et al., 2002, 2010; Peltier et al., 2010), 其光合电子流不等于0 μmol·m-2·s-1。基于以上的分析, 构建的植物叶片J对CO2响应模型(简称模型II)的具体表达式为:
式中, J为植物叶片的总J, αce为J对CO2响应曲线的初始斜率, βce和γce为两个与CO2浓度无关的系数。同样, 模型II中C可以表示为Ca或Ci。J0为叶室CO2浓度等于0 μmol·mol-1 (由于光呼吸和暗呼吸的正常进行, Ci不可能为0)时的J, 简称为零点J。类似地, 假设J0为常数, 则利用模型II可得到植物叶片的饱和CO2浓度(Cc-sat)和Jmax分别为${{C}_{\text{c-sat}}}=\frac{\sqrt{{\text{(}{{\beta }_{\text{ce}}}+{{\gamma }_{\text{ce}}}\text{)}}/{{{\beta }_{\text{ce}}}}\;}-\text{1}}{{{\gamma }_{\text{ce}}}}$和${{J}_{\text{max}}}={{\alpha }_{\text{ce}}}{{\left( \frac{\sqrt{{{\beta }_{\text{ce}}}+{{\gamma }_{\text{ce}}}}-\sqrt{{{\beta }_{\text{ce}}}}}{{{\gamma }_{\text{ce}}}} \right)}^{2}}+{{J}_{0}}$。
1.2 材料培养及叶片选择
以玉米和千穗谷为供试材料。玉米和千穗谷分别于2012年6月上旬和6月中旬播种, 大田常规管理。数据测量于2012年7月25和7月29日的晴天进行。玉米和千穗谷分别处于喇叭口期和营养生长期。随机选取长势较为一致的健壮植株。玉米以其果穗叶为测量对象, 千穗谷则以自上而下第6片完全展开叶为测量对象。此时所选叶片已完全展开, 叶色深绿, 叶形饱满。每株植物选取1片叶子。每种植物测量5-7片叶子。1.3 光合数据测量
利用便携式光合仪(LI-6400-40, LI-COR, Lincoln, USA)于晴天的9:00-12:00进行数据测量。数据测量前进行开机检查及预热。数据测量时, 在自然光下诱导1.5-2.0 h后, 采用开放式气路, 设定叶室温度(Tblock)为35 ℃, 流速为400 μmol·s-1, 空气相对湿度控制在50%-75%, 使用CO2注入系统提供不同的外界CO2梯度(Ca): 1 400、1 200、1 000、800、600、400、380、200、150、100、80、50和0 μmol·mol-1。CO2浓度控制则是通过使用北京力高泰科技有限公司提供的CO2小钢瓶提供CO2来源, 利用LI-6400注入系统, 经过仪器校对后进行控制。由前期预实验的结果可知, 玉米和千穗谷的饱和光强约为2 000 μmol·m-2·s-1, 因此本试验用LI-6400-40荧光探头提供2 000 μmol·m-2·s-1的光合有效辐射(PAR)(红蓝光源, 红光比例为90%, 蓝光比例为10%)。采用自动测量程序(Flr A-Ci curve)同时记录叶片的气体交换和叶绿素荧光等参数。利用这种方法测量饱和光强下玉米和千穗谷叶片的Ac, 即为光合作用对CO2的响应曲线(Ac-Ca); 同时也获得了玉米和千穗谷叶片的J, 即为J对CO2的响应曲线(J-Ca曲线)。另外, 为了探讨暗呼吸CO2的回收利用部分(Re), 试验也测量了CO2浓度分别为380和0 μmol·mol-1时玉米和千穗谷叶片的暗呼吸速率(Rn)。每个CO2浓度记录最小等待时间为2 min, 最大等待时间为3 min, 数据记录之前仪器均自动进行参比室和样品室之间的匹配。
1.4 数据处理
C3植物参与碳还原的光合电子流(Jc)(Valentini et al., 1995)公式为:式中, Rp为光呼吸速率, Rd为光下暗呼吸速率。由于C4植物的光呼吸过程可以忽略不计, 因此, C4植物Jc的公式为:
饱和光强下, 其他途径的电子传递流(Ja)计算如下:
饱和光强下, 暗呼吸CO2的回收利用部分(Re)计算如下:
利用模型I拟合光合作用对CO2的响应曲线(Ac-Ca曲线)得到最大净光合速率(Ac-max), 对应的饱和CO2浓度, CO2补偿点等参数。同时用模型II拟合这些Jc-Ca曲线获得Jmax、对应的饱和CO2浓度(Ca-sat)和零点J (J0)等参数。用确定系数(R2)表示模型拟合与观测点的符合程度。以上拟合采用“光合计算软件” (http://photosynthetic.sinaapp.com/calc.html, 井冈山大学数理学院)对实验数据进行分析; 采用DPS 7.5软件对玉米和千穗谷的相关参数进行方差分析(ANOVA分析)。作图用Origin 8.5软件完成, 然后用Adobe Illustrator CS5进行图形处理。结果取5次测量的平均值。
2 结果
2.1 光合作用对CO2的响应
图1为2种植物的光合作用对大气CO2的响应曲线(Ac-Ca曲线)。由图1可知, 在较低的CO2浓度(< 400 mmol·mol-1)下玉米和千穗谷的响应曲线上升较快, 在设定的最高CO2浓度(1 400 mmol·mol-1)下均达到了饱和。此外, 由图1还可知, 用模型I可以很好地拟合2种C4植物的Ac-Ca曲线, R2均高于0.99。图1
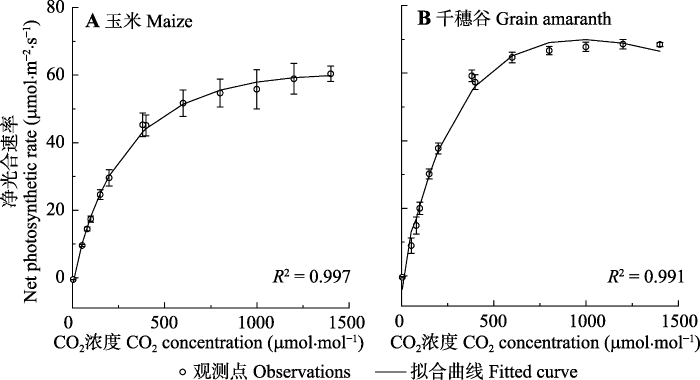 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1玉米和千穗谷光合作用对CO2的响应曲线(平均值±标准误差, n = 5)。
Fig. 1CO2-response curves of photosynthesis in maize and grain amaranth (mean ± SE, n = 5).
由表1的数据可知, 千穗谷的初始斜率(αc)、最大净光合速率(Acmax)显著高于玉米(p < 0.05); 而玉米的饱和CO2浓度(约1 400 mmol·mol-1)则显著高于千穗谷(约1 000 mmol·mol-1)(p < 0.05)。玉米和千穗谷的CO2补偿点均较低, 分别约为4.25和13.41 mmol·mol-1。这与C4植物对CO2利用率较高的光合生理特性相一致。另外, 从表1的数据还可知, 除光下呼吸速率参数外, 模型I拟合玉米和千穗谷的Ac-Ca曲线得到的其他光合参数与观测值高度一致。而模型I给出的光下呼吸速率与观测值存在显著差异(p < 0.05), 分析其可能的原因是模型I没有考虑到CO2浓度对光下呼吸速率的影响。研究表明, CO2浓度对光呼吸、暗呼吸以及光下暗呼吸速率均有显著影响(康华靖等, 2014)。
Table 1
表1
表1由模型I拟合玉米和千穗谷的光合作用对CO2的响应曲线(Ac-Ca)得到光合参数以及相应的估算值(平均值±标准误差, n = 5)
Table 1
| 光合参数 Photosynthetic parameter | 玉米 Maize | 千穗谷 Grain amaranth | ||
|---|---|---|---|---|
| 拟合值 Fitted value | 估算值 Estimated value | 拟合值 Fitted value | 估算值 Estimated value | |
| αc | 0.247 ± 0.033b | — | 0.334 ± 0.022a | — |
| Ac-max (mmol·m-2·s-1) | 59.12 ± 0.67b | ? 60.39 | 69.97 ± 0.71a | ? 70.49 |
| Ca-sat (mmol·mol-1) | 1 335.74 ± 196.52a | ? 1 400 | 976.25 ± 12.06b | ? 1 000 |
| Γ (mmol·mol-1) | 4.35 ± 2.08b | ? 4.25b | 12.77 ± 0.53a | ? 13.41 |
| Rl (mmol·m-2·s-1) | 1.13 ± 0.64b | ? 0.24 | 2.44 ± 0.15a | ? 0.28 |
| R2 | 0.997 | — | 0.991 | — |
新窗口打开|下载CSV
2.2 光合电子传递速率对CO2的响应
图2给出了玉米和千穗谷2种植物J对CO2的响应曲线(J-Ca曲线)。由图2可知, 玉米的J随CO2浓度的增加而先快速增加, 然后趋于稳定; 千穗谷在J到达最大值后, 随CO2浓度的增加而略有下降趋势。此外, 由图2还可知, 用模型II可较好地拟合2种植物的Jc-Ca曲线, R2达到了0.978以上。图2
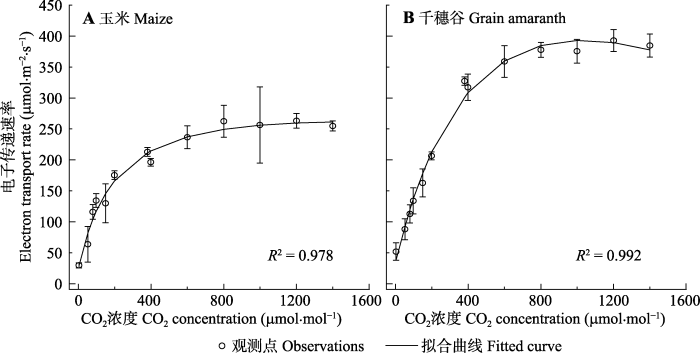 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2玉米和千穗谷的光合电子传递速率对CO2的响应曲线(平均值±标准误差, n = 5)。
Fig. 2CO2-response curves of photosynthetic electron transport rate (J-Ca) in maize and grain amaranth (mean ± SE, n = 5).
由表2的数据可知, 玉米和千穗谷的初始斜率(αce)相近且数值较高(分别为1.215和1.208 mol·m-2·s-1), 表明这两种植物的光能转换为光合电子流的能力较强; J对应的饱和CO2浓度也比较相近(p > 0.05)。而千穗谷的Jmax (约为397 mmol·m-2·s-1)则显著高于玉米(约为265 mmol·m-2·s-1) (p < 0.05)。此外, 从表2的数据还可知, 用模型II拟合玉米和千穗谷的J-Ca曲线得到的光合参数与观测值基本一致。
Table 2
表2
表2由模型II拟合玉米和千穗谷的光合电子传递速率对CO2的响应曲线(J-Ca )得到Jmax和Ca-sat等参数以及相应的估算值(平均值±标准误差, n = 5)
Table 2
| 光合参数 Photosynthetic parameter | 玉米 Maize | 千穗谷 Grain amaranth | ||
|---|---|---|---|---|
| 拟合值 Fitted value | 估算值 Estimated value | 拟合值 Fitted value | 估算值 Estimated value | |
| αce (mol·m-2·s-1) | 1.215 ± 0.543a | — | 1.208 ± 0.357a | — |
| Jmax (mmol·m-2·s-1) | 262.41 ± 1.64b | ? 265.66 | 393.07 ± 37.84a | ? 397.82 |
| Ca-sat (mmol·mol-1) | 1 198.58 ± 342.78a | ? 1 200 | 1 229.10 ± 59.14a | ? 1 200 |
| J0 (mmol·m-2·s-1) | 22.22 ± 8.35a | ? 27.69 | 27.43 ± 4.97a | ? 29.26 |
| R2 | 0.978 | — | 0.992 | — |
新窗口打开|下载CSV
3 讨论和结论
在植物光合作用的原初反应过程中, 由捕光色素分子收集的光能共振激发传递到PSII和PSI后产生电荷分离, 形成光合电子流。这些电子被电子受体接受后依次传递, 最终电子被传递到高能化合物NADPH和ATP中, 完成了光能到化学能的转换并为碳同化和光呼吸提供能量(薛娴等, 2017)。准确估算光合电子流对CO2响应的变化趋势、Ac-max、Jmax以及它们对应的饱和CO2浓度对深入了解光合内部过程具有重要意义。3.1 光合作用和光合电子传递速率对CO2的响应
从本研究来看, 基于模型I构建的J对CO2响应模型(模型II)也可很好地拟合玉米和千穗谷的J-Ca响应曲线(R2 > 0.978), 给出的Jmax等光合参数整体上与测量值高度吻合。因此, 模型II可以定量研究玉米和千穗谷这2种植物的J-Ca响应曲线的变化趋势。对比图1和图2还可以发现, 模型II拟合玉米和千穗谷的J-Ca响应曲线, 其趋势与其相应的Ac-Ca响应曲线基本一致。由表2可知, 千穗谷的Jmax (393.07 mmol·m-2·s-1)则远高于玉米(262.41 mmol·m-2·s-1)。这表明千穗谷比玉米产生ATP和NADPH的能力更强, 因而碳同化能力也更强。表1数据显示, 千穗谷的最大净光合速率(69.97 mmol·m-2·s-1)也显著高于玉米(59.12 mmol·m-2·s-1)(p < 0.05)。然而, 从饱和CO2浓度来看, 虽然玉米和千穗谷之间的Ca-sat (J所对应的饱和CO2浓度)没有差异(p > 0.05), 分别为1 198.58和1 229.10 mmol·mol-1, 但玉米和千穗谷之间的Csat (光合速率所对应的饱和CO2浓度)则存在显著差异(p < 0.05), 分别为1 335.74和976.25 mmol·mol-1。这表明千穗谷的光合速率比玉米更容易达到CO2饱和。千穗谷的J显著高于玉米, 而光合作用反而更容易达到饱和。3.2 暗呼吸CO2回收利用对光合电子流分配的影响
植物的光合作用是光合能量和底物相互偶联的氧化还原反应(林植芳等, 2000)。因此, 可根据碳同化速率计算其J。由于C4植物的光呼吸过程可以忽略不计, 那么从光合生理过程来看, 在常规大气条件下, C4植物光合作用正常进行时的碳代谢只有光合碳同化和暗呼吸。在有光条件下且当外界CO2浓度为0 mmol·mol-1时, C4植物光合碳同化过程被抑制, 而暗呼吸活动则正常进行。因此, 此时的表观光合速率测量值应为光下暗呼吸速率(Rd)(康华靖等, 2014)。由表1的数据可知, 玉米和千穗谷的光下暗呼吸速率分别为0.24和0.28 mmol·m-2·s-1。玉米和千穗谷的暗呼吸速率(Rn)测量值见表3, 分别为3.27和3.17 mmol·m-2·s-1。比较数据可知, 玉米和千穗谷的Rd远小于其对应的Rn。部分研究者认为这是由于暗呼吸速率受光强抑制(Foyer & Noctor, 2000; Yin et al., 2011)。然而, 也有研究认为光呼吸和暗呼吸产生的CO2释放点靠近叶绿体内侧, 且没有边界层、气孔以及细胞壁的限制。因此, 其CO2极易被光合作用重新利用(Loreto et al., 1999)。13C同位素的研究已证实玉米叶片暗呼吸的CO2释放均可被光合作用重新利用(Loreto et al., 2001)。康华靖等(2014)利用气体交换法的研究结果也表明, 植物Rd的降低不是由于光强的抑制, 而是由于光下暗呼吸放出的CO2又重新回收利用所引起; 其回收利用率随着光强的增加而增大。基于该观点来看, 更有利于理解当外界CO2浓度为0 mmol·mol-1时, 植物叶片的J仍保持一定的数值。根据公式(6)计算出玉米和千穗谷的暗呼吸被重新回收利用部分(Re)见表3, 分别为3.02和2.89 mmol·m-2·s-1。Table 3
表3
表32 000 μmol·m-2·s-1光强下玉米和千穗谷光合作用相关参数及电子流分配比较
Table 3
| 光合作用参数 Photosynthetic parameter (mmol·m-2·s-1) | [CO2] = 380 mmol·mol-1 | [CO2] = 0 mmol·mol-1 | ||
|---|---|---|---|---|
| 玉米 Maize | 千穗谷 Grain amaranth | 玉米 Maize | 千穗谷Grain amaranth | |
| Ac-max | 59.12 | 69.97 | - | - |
| Rn | 2.86 | 1.32 | 3.27 | 3.17 |
| Rd | 1.43 | 0.66 | 0.24 | 0.28 |
| Re | 1.43 | 0.66 | 3.02 | 2.89 |
| Jmax | 262.41 | 393.07 | - | - |
| J0 | - | - | 22.22 | 27.43 |
| Jc | 242.20 | 282.52 | 0.96 | 1.12 |
| Ja | 20.21 | 110.55 | 21.26 | 26.31 |
| J°c | 247.92 | 285.16 | 13.08 | 12.68 |
| J°a | 14.49 | 107.91 | 9.14 | 14.75 |
新窗口打开|下载CSV
从光合碳同化电子流分配来看, 当CO2浓度为0 mmol·mol-1时, 根据公式(3)可知, 如不考虑暗呼吸CO2回收利用, 玉米和千穗谷分配到C同化的电子流分别为0.96和1.12 mmol·m-2·s-1 (表3); 如考虑暗呼吸CO2的回收利用, 玉米和千穗谷分配到C同化的电子流则分别为13.08和12.68 mmol·m-2·s-1 (表3)。由此可见, CO2浓度为0 mmol·mol-1时如不考虑暗呼吸CO2回收利用将严重低估光合碳同化电子流分配。此外,由于CO2浓度为380 mmol·mol-1时的光下暗呼吸速率值难以直接测量。传统计算认为Rd是Rn的一半(Fila et al., 2006)。由此根据公式(3)计算出CO2浓度为380 mmol·mol-1时, 如不考虑CO2的回收利用, 玉米和千穗谷碳同化所分配的电子流分别为242.20和282.52 mmol·m-2·s-1 (表3); 如考虑CO2的回收利用, 玉米和千穗谷碳同化所分配的电子流则分别为247.92和285.16 mmol·m-2·s-1 (表3)。由此可见, CO2浓度为380 mmol·mol-1时, 是否考虑暗呼吸CO2的回收利用对其光合碳同化电子流分配的影响较小。
而从光合电子流分配到其他途径来看, 当CO2浓度为0 mmol·mol-1时, 如不考虑暗呼吸CO2的回收利用, 玉米和千穗谷的其他途径所分配电子流分别为21.26和26.31 mmol·m-2·s-1; 如考虑暗呼吸CO2的回收利用, 玉米和千穗谷的其他途径所分配电子流分别为9.14和14.75 mmol·m-2·s-1 (表3)。由此可见, CO2浓度为0 mmol·mol-1时如不考虑暗呼吸CO2的回收利用, 将严重高估其他途径的电子流分配。同样, CO2浓度为380 mmol·mol-1时, 如不考虑暗呼吸CO2的回收利用, 玉米和千穗谷的其他途径所分配电子流分别为20.21和110.55 mmol·m-2·s-1; 如考虑暗呼吸CO2的回收利用, 玉米和千穗谷的其他途径所分配电子流分别为14.49和107.91 mmol·m-2·s-1 (表3)。由此可见, CO2浓度为380 mmol·mol-1时, 是否考虑暗呼吸CO2的回收利用对其他途径电子流分配的影响也相对较小, 尤其在千穗谷上表现得更为不明显。
3.3 玉米和千穗谷的光合电子流分配到非碳同化路径差异显著的原因及可能的解释
当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%。由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(Miyake et al., 2002)、围绕PSI的电子循环(Heber, 2002)、米勒反应等所消耗的电子流均占极小比例。这与目前在计算碳同化和光呼吸所分配的光合电子流时, 往往将其他途径分配的电子流视为常数或者忽略(Epron et al., 1995; Valentini et al., 1995)的做法相一致。比较其他途径电子传递速率值来看, 以CO2浓度为380 mmol·mol-1时的数据为例(考虑暗呼吸CO2的回收利用), 玉米仅为14.49 mmol·m-2·s-1, 而千穗谷高达107.91 mmol·m-2·s-1, 两者相差6倍多。分析其可能的原因是与千穗谷和玉米的催化脱羧反应酶种类以及脱羧反应发生的部位不同有直接的关联(Taylor et al., 2010; 冯瑞云等, 2011; 贺飞燕等, 2017; 李小博等, 2017)。玉米是NADP苹果酸酶(NADP-ME)型; 而千穗谷是NAD苹果酸酶(NAD-ME)型。玉米脱羧后形成的丙酮酸返回到叶肉细胞中, 由丙酮酸磷酸双激酶催化, 形成磷酸烯醇式丙酮酸(PEP)然后重新接受CO2 (Taylor et al., 2010; 冯瑞云等, 2011; 贺飞燕等, 2017); 而在千穗谷中形成的丙氨酸先在叶肉细胞质中转变为丙酮酸, 再生成PEP。由此可知, 在生成PEP方面, 千穗谷比玉米要多一步才能完成。这可能是导致千穗谷比玉米需要消耗更多的光合电子流和更容易达到CO2饱和的主要的和直接的原因。总之, 本文构建的J对CO2响应模型可以很好地拟合玉米和千穗谷的J-Ca响应曲线, 给出的Jmax、Ca-sat和J0等相关参数与观测值高度相符。如果能综合利用模型I和模型II, 则可以为研究植物的光合特性、光合电子流对CO2的响应趋势、零点光合电子流和植物对逆境的响应提供有效的数学工具。同时, 也为人们从光合作用和光合电子流对CO2响应的角度研究C4植物中NADP-ME和NAD-ME两种亚型之间的差异提供一个新的视角。

致谢
温州市重点科技创新团队项目(C20150008)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
In: Hall D, Coombs J, Goodwin T eds.
[本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 1]

Light response (at 300 ppm CO(2) and 10-50 ppm O(2) in N(2)) and CO(2) response curves [at absorbed photon fluence rate (PAD) of 550 μmol m(-2) s(-1)] of O(2) evolution and CO(2) uptake were measured in tobacco (Nicotiana tabacum L.) leaves grown on either NO(3)(-) or NH(4)(+) as N source and in potato (Solanum tuberosum L.), sorghum (Sorghum bicolor L. Moench), and amaranth (Amaranthus cruentus L.) leaves grown on NH(4)NO(3). Photosynthetic O(2) evolution in excess of CO(2) uptake was measured with a stabilized zirconia O(2) electrode and an infrared CO(2) analyser, respectively, and the difference assumed to represent the rate of electron flow to acceptors alternative to CO(2), mainly NO(2)(-), SO(4)(2-), and oxaloacetate. In NO(3)(-)-grown tobacco, as well as in sorghum, amaranth, and young potato, the photosynthetic O(2)-CO(2) flux difference rapidly increased to about 1 μmol m(-2) s(-1) at very low PADs and the process was saturated at 50 μmol quanta m(-2) s(-1). At higher PADs the O(2)-CO(2) flux difference continued to increase proportionally with the photosynthetic rate to a maximum of about 2 μmol m(-2) s(-1). In NH(4)(+)-grown tobacco, as well as in potato during tuber filling, the low-PAD component of surplus O(2) evolution was virtually absent. The low-PAD phase was ascribed to photoreduction of NO(2)(-) which successfully competes with CO(2) reduction and saturates at a rate of about 1 μmol O(2) m(-2) s(-1) (9% of the maximum O(2) evolution rate). The high-PAD component of about 1 μmol O(2) m(-2) s(-1), superimposed on NO(2)(-) reduction, may represent oxaloacetate reduction. The roles of NO(2)(-), oxaloacetate, and O(2) reduction in the regulation of ATP/NADPH balance are discussed.
[本文引用: 1]
DOIURLPMID [本文引用: 1]
Abstract Mitochondrial respiration often appears to be inhibited in the light when compared with measurements in the dark. This inhibition is inferred from the response of the net CO2 assimilation rate (A) to absorbed irradiance (I), changing slope around the light compensation point (Ic). We suggest a model that provides a plausible mechanistic explanation of this ok effect . The model uses the mathematical description of photosynthesis developed by Farquhar, von Caemmerer and Berry; it involves no inhibition of respiration rate in the light. We also describe a fitting technique for quantifying the Kok effect at low I. Changes in the chloroplastic CO2 partial pressure (Cc) can explain curvature of A vs I, its diminution in C4 plants and at low oxygen concentrations or high carbon dioxide concentrations in C3 plants, and effects of dark respiration rate and of temperature. It also explains the apparent inhibition of respiration in the light as inferred by the Laisk approach. While there are probably other sources of curvature in A vs I, variation in Cc can largely explain the curvature at low irradiance, and suggests that interpretation of day respiration compared with dark respiration of leaves on the basis of the Kok effect needs reassessment.
DOIURL [本文引用: 2]
磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase,PEPC)是C4途径的关键酶之一.本研究克隆了双子叶植物籽粒苋(A.hypochondriacus L.) C4型PEPC基因的cDNA和gDNA全长序列.该基因编码区cDNA全长2895bp (GenBank登录号为HQ186302),编码964个氨基酸残基.对应的gDNA全长为6 785 bp,含10个外显子和9个内含子,内含子总长3 890 bp,其中第一内含子最长,为1 662 bp,具GATA-motif、G-box等15个光应答元件,9个激素应答元件,29个CAAT box,72个TATA box.该特征在本研究范围内的植物中,为籽粒苋所独有.比较多种植物表明,不同植物之间的C3/C4型PEPC基因的外显子长度具有较高的一致性,而内含子的长度变化较大.不同植物PEPC基因对核苷酸具有一定的选择性,并且外显子和内含子的GC含量呈正相关.总的趋势是单子叶植物外显子和内含子的GC含量均高于双子叶植物.利用半定量RT-PCR分析表达模式,发现籽粒苋PEPC基因主要在叶片中表达,表达量随光照时间延长而增加.
DOIURL [本文引用: 2]
磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase,PEPC)是C4途径的关键酶之一.本研究克隆了双子叶植物籽粒苋(A.hypochondriacus L.) C4型PEPC基因的cDNA和gDNA全长序列.该基因编码区cDNA全长2895bp (GenBank登录号为HQ186302),编码964个氨基酸残基.对应的gDNA全长为6 785 bp,含10个外显子和9个内含子,内含子总长3 890 bp,其中第一内含子最长,为1 662 bp,具GATA-motif、G-box等15个光应答元件,9个激素应答元件,29个CAAT box,72个TATA box.该特征在本研究范围内的植物中,为籽粒苋所独有.比较多种植物表明,不同植物之间的C3/C4型PEPC基因的外显子长度具有较高的一致性,而内含子的长度变化较大.不同植物PEPC基因对核苷酸具有一定的选择性,并且外显子和内含子的GC含量呈正相关.总的趋势是单子叶植物外显子和内含子的GC含量均高于双子叶植物.利用半定量RT-PCR分析表达模式,发现籽粒苋PEPC基因主要在叶片中表达,表达量随光照时间延长而增加.
DOIURLPMID [本文引用: 1]
Abstract In vitro-cultured plants typically show a low photosynthetic activity, which is considered detrimental to subsequent ex vitro acclimatization. Studies conducted so far have approached this problem by analysing the biochemical and photochemical aspects of photosynthesis, while very little attention has been paid to the role of leaf conductance to CO(2) diffusion, which often represents an important constraint to CO(2) assimilation in naturally grown plants. Mesophyll conductance, in particular, has never been determined in in vitro plants, and no information exists as to whether it represents a limitation to carbon assimilation during in vitro growth and subsequent ex vitro acclimatization. In this study, by means of simultaneous gas exchange and chlorophyll fluorescence measurements, the stomatal and mesophyll conductance to CO(2) diffusion were assessed in in vitro-cultured plants of the grapevine rootstock '41B' (Vitis vinifera 'Chasselas'xVitis berlandieri), prior to and after ex vitro acclimatization. Their impact on electron transport rate partitioning and on limitation of potential net assimilation rate was analysed. In vitro plants had a high stomatal conductance, 155 versus 50 mmol m(-2) s(-1) in acclimatized plants, which ensured a higher CO(2) concentration in the chloroplasts, and a 7% higher electron flow to the carbon reduction pathway. The high stomatal conductance was counterbalanced by a low mesophyll conductance, 43 versus 285 mmol m(-2) s(-1), which accounted for a 14.5% estimated relative limitation to photosynthesis against 2.1% estimated in acclimatized plants. It was concluded that mesophyll conductance represents an important limitation for in vitro plant photosynthesis, and that in acclimatization studies the correct comparison of photosynthetic activity between in vitro and acclimatized plants must take into account the contribution of both stomatal and mesophyll conductance.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Publication » C4 photosynthesis: a unique blend of modified bichemistry, anatomy and ultrastructure.
DOIURL [本文引用: 2]
为了进一步探究籽粒苋丙酮酸磷酸双激酶(AhPPDK)蛋白的作用机制,构建了AhPPDK基因的原核表达载体,通过碱裂解提取质粒,经限制性内切酶酶切,采用1.0%琼脂糖凝胶电泳检测酶切产物,选择正确表达的阳性重组子,将测序正确的重组质粒pEASY-E1-AhPPDK转化菌株Transetta(DE3);利用IPTG诱导蛋白表达.SDS-PAGE凝胶电泳分析表明,重组AhPPDK能在大肠杆菌Transset(DE3)中高效表达,表达的重组蛋白质分子量约为108 kDa,与预期分子量相符,且为可溶性蛋白.利用紫外分光光度法测量结果表明,原核表达的AhPPDK具有酶活性,且上清粗酶液活性高于沉淀粗酶液.研究结果可为进一步探明AhPPDK蛋白的作用机制和转基因利用奠定基础.
DOIURL [本文引用: 2]
为了进一步探究籽粒苋丙酮酸磷酸双激酶(AhPPDK)蛋白的作用机制,构建了AhPPDK基因的原核表达载体,通过碱裂解提取质粒,经限制性内切酶酶切,采用1.0%琼脂糖凝胶电泳检测酶切产物,选择正确表达的阳性重组子,将测序正确的重组质粒pEASY-E1-AhPPDK转化菌株Transetta(DE3);利用IPTG诱导蛋白表达.SDS-PAGE凝胶电泳分析表明,重组AhPPDK能在大肠杆菌Transset(DE3)中高效表达,表达的重组蛋白质分子量约为108 kDa,与预期分子量相符,且为可溶性蛋白.利用紫外分光光度法测量结果表明,原核表达的AhPPDK具有酶活性,且上清粗酶液活性高于沉淀粗酶液.研究结果可为进一步探明AhPPDK蛋白的作用机制和转基因利用奠定基础.
DOIURL [本文引用: 2]
DOIURL [本文引用: 3]
以C3作物(小麦,Triticum aestivum和大豆,Glycine max)和C4作物(玉米,Zeamays和千穗谷,Amaranthus hypochondria-cus)为例,探讨了其光下暗呼吸速率降低的原因.结果表明,2% O2条件下,CO2浓度为0时,叶室CO2浓度维持在0左右,而胞间CO2浓度(Ci)显著高于叶室CO2浓度.分析认为这是由于此时植物的暗呼吸仍在正 常进行.因此,该测量条件下的表观光合速率应为CO2浓度为0时的光下暗呼吸速率(Rd).CO2浓度为0时,不同光强下的Rd均随光强的升高而降低,且 在低光强(50μmol·m-2·s 1)和高光强(2 000 μmol·m-2·s-1)之间存在显著差异,说明光强对Rd具有较大影响.在2% O2条件下,经饱和光强充分活化而断光后,以上4种作物叶片的暗呼吸速率(Rn)均随着时间的推移而下降,说明光强并未抑制暗呼吸速率.试验结果表 明,Rd的降低是由于CO2被重新回收利用所导致,CO2回收利用率随光强的升高而增大,从低光强(50 μmol·m-2·s-1)到高光强(2000μmnol·m-2·s-1),小麦、大豆、玉米和千穗谷的回收利用率范围变动分别为 22.65%-52.91%、22.40%-55.31%、54.24%-87.59%和72.43%-90.07%.
DOIURL [本文引用: 3]
以C3作物(小麦,Triticum aestivum和大豆,Glycine max)和C4作物(玉米,Zeamays和千穗谷,Amaranthus hypochondria-cus)为例,探讨了其光下暗呼吸速率降低的原因.结果表明,2% O2条件下,CO2浓度为0时,叶室CO2浓度维持在0左右,而胞间CO2浓度(Ci)显著高于叶室CO2浓度.分析认为这是由于此时植物的暗呼吸仍在正 常进行.因此,该测量条件下的表观光合速率应为CO2浓度为0时的光下暗呼吸速率(Rd).CO2浓度为0时,不同光强下的Rd均随光强的升高而降低,且 在低光强(50μmol·m-2·s 1)和高光强(2 000 μmol·m-2·s-1)之间存在显著差异,说明光强对Rd具有较大影响.在2% O2条件下,经饱和光强充分活化而断光后,以上4种作物叶片的暗呼吸速率(Rn)均随着时间的推移而下降,说明光强并未抑制暗呼吸速率.试验结果表 明,Rd的降低是由于CO2被重新回收利用所导致,CO2回收利用率随光强的升高而增大,从低光强(50 μmol·m-2·s-1)到高光强(2000μmnol·m-2·s-1),小麦、大豆、玉米和千穗谷的回收利用率范围变动分别为 22.65%-52.91%、22.40%-55.31%、54.24%-87.59%和72.43%-90.07%.
DOIURLPMID [本文引用: 1]
Using an Agrobacterium-mediated transformation system, we have introduced the intact gene of maize phosphoenolpyruvate carboxylase (PEPC), which catalyzes the initial fixation of atmospheric CO2 in C4 plants into the C3 crop rice. Most transgenic rice plants showed high-level expression of the maize gene; the activities of PEPC in leaves of some transgenic plants were two- to threefold higher than those in maize, and the enzyme accounted for up to 12% of the total leaf soluble protein. RNA gel blot and Southern blot analyses showed that the level of expression of the maize PEPC in transgenic rice plants correlated with the amount of transcript and the copy number of the inserted maize gene. Physiologically, the transgenic plants exhibited reduced O2 inhibition of photosynthesis and photosynthetic rates comparable to those of untransformed plants. The results demonstrate a successful strategy for installing the key biochemical component of the C4 pathway of photosynthesis in C3 plants.
URL [本文引用: 1]
为了明确玉米C_4型pepc(磷酸烯醇式丙酮酸羧化酶基因)、ppdk(丙酮酸磷酸双激酶基因)、nadp-me(NADP-苹果酸酶基因)基因对拟南芥光合特性的影响效应,以PC(转Zm pepc基因)、PK(转Zm ppdk基因)、ME(转Zm nadp-me基因)、PCK(转Zm pepc+ppdk基因)、PKM(转Zm ppdk+nadp-me基因)5种转基因拟南芥和野生型WT为材料,鉴定了5种转基因拟南芥株系的外源基因表达量,测定了250μmol·m~(-2)·s~(-1)、800μmol·m~(-2)·s~(-1)、1 400μmol·m~(-2)·s~(-1)3种光照强度处理下转基因拟南芥株系相应的光合酶活性、光合速率。结果表明,5种转基因拟南芥在转录、翻译水平上均高表达了导入的外源基因。250μmol·m~(-2)·s~(-1)光强下,转基因拟南芥材料PC、PK、ME、PCK、PKM的PEPC酶活性较WT分别提高14.4%、9.1%、4.4%、21.6%、3.5%;PPDK酶活性较WT分别提高5.2%、33.2%、14.8%、24.9%、8.1%;NADP-ME酶活性较WT分别提高12.8%、42.0%、126.4%、31.9%、64.6%;净光合速率较WT分别提高26.6%、26.4%、5.4%、43.1%、14.1%,其中PC、PCK的C_4光合酶活性和净光合速率表现最为突出。随着光强胁迫的增加,转基因拟南芥上述测定指标较WT高出的幅度增加,较250μmol·m~(-2)·s~(-1)光强下降低的幅度小于WT,说明转基因拟南芥耐强光胁迫能力更强。转基因拟南芥酶活特性、光合速率存在基因型差异,单基因材料PC>PK>ME,双基因材料PCK>PKM,且PCK为本研究所有基因型中最优的材料。
URL [本文引用: 1]
为了明确玉米C_4型pepc(磷酸烯醇式丙酮酸羧化酶基因)、ppdk(丙酮酸磷酸双激酶基因)、nadp-me(NADP-苹果酸酶基因)基因对拟南芥光合特性的影响效应,以PC(转Zm pepc基因)、PK(转Zm ppdk基因)、ME(转Zm nadp-me基因)、PCK(转Zm pepc+ppdk基因)、PKM(转Zm ppdk+nadp-me基因)5种转基因拟南芥和野生型WT为材料,鉴定了5种转基因拟南芥株系的外源基因表达量,测定了250μmol·m~(-2)·s~(-1)、800μmol·m~(-2)·s~(-1)、1 400μmol·m~(-2)·s~(-1)3种光照强度处理下转基因拟南芥株系相应的光合酶活性、光合速率。结果表明,5种转基因拟南芥在转录、翻译水平上均高表达了导入的外源基因。250μmol·m~(-2)·s~(-1)光强下,转基因拟南芥材料PC、PK、ME、PCK、PKM的PEPC酶活性较WT分别提高14.4%、9.1%、4.4%、21.6%、3.5%;PPDK酶活性较WT分别提高5.2%、33.2%、14.8%、24.9%、8.1%;NADP-ME酶活性较WT分别提高12.8%、42.0%、126.4%、31.9%、64.6%;净光合速率较WT分别提高26.6%、26.4%、5.4%、43.1%、14.1%,其中PC、PCK的C_4光合酶活性和净光合速率表现最为突出。随着光强胁迫的增加,转基因拟南芥上述测定指标较WT高出的幅度增加,较250μmol·m~(-2)·s~(-1)光强下降低的幅度小于WT,说明转基因拟南芥耐强光胁迫能力更强。转基因拟南芥酶活特性、光合速率存在基因型差异,单基因材料PC>PK>ME,双基因材料PCK>PKM,且PCK为本研究所有基因型中最优的材料。
DOIURL [本文引用: 1]
由Farquhar、von Caemmerer和Berry提出的生物化学光合模型(以下简称FvCB模型)是一个基于光合碳反应过程的CO_2响应模型。此模型认为C3植物叶片光合速率(A)由3个生物化学过程速率中的最低者——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)所能支持的羧化速率、电子传递所能支持的核酮糖-1,5-双磷酸(Ru BP)再生速率和磷酸丙糖(TP)利用速率决定。利用改进的FvCB模型对光合速率-胞间CO_2浓度(A-C_i)曲线进行拟合,能有效地估计最大羧化速率、最大电子传递速率、TP利用速率、明呼吸速率、叶肉细胞导度等生化参数,促进我们对植物光合生理及其响应环境变化的理解和预测。该文首先详细地描述了FvCB模型,并分析了此模型分段性和过参数化的特点。然后介绍利用FvCB模型对A-C_i曲线进行拟合,从而估计叶片光合生化参数的研究进展。光合生化参数估计经历了主观分段、分段拟合到客观分段、整体拟合几个阶段,目标函数的最小化方法也从传统的最小二乘法为主转向基于现代计算机技术的迭代算法(如遗传算法、模拟退火算法)。然而,如要进一步提高参数估计的可靠性和精确性,还需加强Rubisco动力学属性和温度依赖性方面的研究。最后,为了获取能更有效地进行参数估计的光合数据,根据目前对FvCB模型拟合的认知,整合并改进了A-C_i曲线的测定方法。
DOIURL [本文引用: 1]
由Farquhar、von Caemmerer和Berry提出的生物化学光合模型(以下简称FvCB模型)是一个基于光合碳反应过程的CO_2响应模型。此模型认为C3植物叶片光合速率(A)由3个生物化学过程速率中的最低者——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)所能支持的羧化速率、电子传递所能支持的核酮糖-1,5-双磷酸(Ru BP)再生速率和磷酸丙糖(TP)利用速率决定。利用改进的FvCB模型对光合速率-胞间CO_2浓度(A-C_i)曲线进行拟合,能有效地估计最大羧化速率、最大电子传递速率、TP利用速率、明呼吸速率、叶肉细胞导度等生化参数,促进我们对植物光合生理及其响应环境变化的理解和预测。该文首先详细地描述了FvCB模型,并分析了此模型分段性和过参数化的特点。然后介绍利用FvCB模型对A-C_i曲线进行拟合,从而估计叶片光合生化参数的研究进展。光合生化参数估计经历了主观分段、分段拟合到客观分段、整体拟合几个阶段,目标函数的最小化方法也从传统的最小二乘法为主转向基于现代计算机技术的迭代算法(如遗传算法、模拟退火算法)。然而,如要进一步提高参数估计的可靠性和精确性,还需加强Rubisco动力学属性和温度依赖性方面的研究。最后,为了获取能更有效地进行参数估计的光合数据,根据目前对FvCB模型拟合的认知,整合并改进了A-C_i曲线的测定方法。
DOIURL [本文引用: 1]
研究夏季生长于自然光和遮阴降低光强至36%和16%下的4种亚 热带自然林植物幼苗的光合特性.除荷树外,黧蒴和林下灌木九节与罗伞在自然光下的光合速率比36%光下低,出现光合驯化(下调)现象.高光强下总的光合电 子传递速率JF及其向光呼吸传递的比率JO/JF明显增大.JO/JF在自然光下的叶片中高达0.5~0.6,与提高的Rubisco氧化速率、乙醇酸氧 化酶活性和光呼吸速率相一致,表明提高光合电子向光呼吸途径的传递比率是高光高温下森林植物的一种保护性调节机制.
DOIURL [本文引用: 1]
研究夏季生长于自然光和遮阴降低光强至36%和16%下的4种亚 热带自然林植物幼苗的光合特性.除荷树外,黧蒴和林下灌木九节与罗伞在自然光下的光合速率比36%光下低,出现光合驯化(下调)现象.高光强下总的光合电 子传递速率JF及其向光呼吸传递的比率JO/JF明显增大.JO/JF在自然光下的叶片中高达0.5~0.6,与提高的Rubisco氧化速率、乙醇酸氧 化酶活性和光呼吸速率相一致,表明提高光合电子向光呼吸途径的传递比率是高光高温下森林植物的一种保护性调节机制.
DOIURL [本文引用: 1]
Abstract Photorespiration rate can be estimated in vivo but the actual amount of photorespiratory CO2 emitted or recycled by the leaf is largely unknown. We exploited the insensitivity of infrared gas analy-zers to 13CO2 to detect the photorespiratory CO2 emission of leaves exposed to air containing only 13CO2. Photorespiratory CO2 emission was calculated by subtracting the 12CO2 emitted under non-photorespiratory (2% O2) conditions from that emitted under photorespiratory (21% O2) conditions. Illuminated leaves of herbaceous and tree species emitted a constant amount of 12CO2 within 30 60 s after switching to 13CO2. Photorespiratory CO2 emission was less than 20% of the photorespiratory rate simultaneously calculated by combining fluorescence and gas exchange. Thus, the leaves recycled more than 80% of photorespiratory CO2. The highest recycling was associated with high rates of photosynthesis in tomato and spinach. In the dark, mitochondrial respiration measured by the emission of 12CO2 in air with 13CO2 was similar to that measured by conventional gas exchange in ambient air, thus confirming the accuracy of our method. In the light, mitochondrial respiration estimated from the emission of 12CO2 after complete labeling of photorespiration was lower than in the dark, suggesting either light-inhibition or recycling of respiratory carbon.
[本文引用: 1]
DOIURLPMID [本文引用: 1]
An electron flow in addition to the major electron sinks in C(3) plants [both photosynthetic carbon reduction (PCR) and photorespiratory carbon oxidation (PCO) cycles] is termed an alternative electron flow (AEF) and functions in the chloroplasts of leaves. The water-water cycle (WWC; Mehler-ascorbate peroxidase pathway) and cyclic electron flow around PSI (CEF-PSI) have been studied as the main AEFs in chloroplasts and are proposed to play a physiologically important role in both the regulation of photosynthesis and the alleviation of photoinhibition. In the present review, I discuss the molecular mechanisms of both AEFs and their functions in vivo. To determine their physiological function, accurate measurement of the electron flux of AEFs in vivo are required. Methods to assay electron flux in CEF-PSI have been developed recently and their problematic points are discussed. The common physiological function of both the WWC and CEF-PSI is the supply of ATP to drive net CO(2) assimilation. The requirement for ATP depends on the activities of both PCR and PCO cycles, and changes in both WWC and CEF-PSI were compared with the data obtained in intact leaves. Furthermore, the fact that CEF-PSI cannot function independently has been demonstrated. I propose a model for the regulation of CEF-PSI by WWC, in which WWC is indispensable as an electron sink for the expression of CEF-PSI activity.
DOIURLPMID [本文引用: 2]
Abstract Using thylakoid membranes, we previously demonstrated that accumulated electrons in the photosynthetic electron transport system induces the electron flow from the acceptor side of PSII to its donor side only in the presence of a pH gradient ((Delta)pH) across the thylakoid membranes. This electron flow has been referred to as cyclic electron flow within PSII (CEF-PSII) [Miyake and Yokota (2001) Plant Cell Physiol. 42: 508]. In the present study, we examined whether CEF-PSII operates in isolated intact chloroplasts from spinach leaves, by correlating the quantum yield of PSII [Phi(PSII)] with the activity of the linear electron flow [V(O(2))]. The addition of the protonophore nigericin to the intact chloroplasts decreased Phi(PSII), but increased V(O(2)), and relative electron flux in PSII [Phi(PSII) x PFD] and V(O(2)) were proportional to one another. Phi(PSII) x PFD at a given V(O(2)) was much higher in the presence of (Delta)pH than that in its absence. These effects of nigericin on the relationship between Phi(PSII) x PFD and V(O(2)) are consistent with those previously observed in thylakoid membranes, indicating the occurrence of CEF-PSII also in intact chloroplasts. In the presence of (Delta)pH, CEF-PSII accounted for the excess electron flux in PSII that could not be attributed to photosynthetic linear electron flow. The activity of CEF-PSII increased with increased light intensity and almost corresponded to that of the water-water cycle (WWC), implying that CEF-PSII can dissipate excess photon energy in cooperation with WWC to protect PSII from photoinhibition under limited photosynthesis conditions.
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 1]
Abstract We examined the effects of leaf temperature on the estimation of maximal Rubisco capacity (Vcmax) from gas exchange measurements of wheat leaves using a C3 photosynthesis model. Cultivars of spring wheat (Triticum aestivum (L)) and triticale (x Triticosecale Wittmack) were grown in a greenhouse or in the field and measured at a range of temperatures under controlled conditions in a growth cabinet (2 and 21 % O2) or in the field using natural diurnal variation in temperature, respectively. Published Rubisco kinetic constants for tobacco did not describe the observed CO2 response curves well as temperature varied. By assuming values for the Rubisco Michaelis-Menten constants for CO2 (Kc) and O2 (Ko) at 25 C derived from tobacco and the activation energies of Vcmax from wheat and respiration in the light, Rd, from tobacco, we derived activation energies for Kc and Ko (93.7 and 33.6 kJ mol-1, respectively) that considerably improved the fit of the model to observed data. We confirmed that temperature dependence of dark respiration for wheat was well described by the activation energy for Rd from tobacco. The new parameters improved the estimation of Vcmax under field conditions, where temperatures increased through the day.
URL [本文引用: 1]
为提高叶片光合速率并更好地理解叶片光合生理对环境因子变化的响应机制,FvCB模型(C_3植物光合生化模型)常用于分析不同环境条件下CO_2响应曲线并预测叶片活体内光合系统的内在变化状况。系统介绍了FvCB模型的建立、发展过程和拟合方法等基本理论,综述了该模型在叶片光合生理对光、CO_2、水、温度和N营养等环境因子变化的响应机制中的应用研究。为进一步完善FvCB模型并更好地理解叶片活体内光合系统对环境因子变化的响应机制,未来拟加强以下研究:1)羧化速率与光合电子传递速率之间的联系;2)叶肉导度的具体组分及其对FvCB模型参数估计的影响;3)叶片气孔导度和叶肉导度对环境因子变化的调控机制。
URL [本文引用: 1]
为提高叶片光合速率并更好地理解叶片光合生理对环境因子变化的响应机制,FvCB模型(C_3植物光合生化模型)常用于分析不同环境条件下CO_2响应曲线并预测叶片活体内光合系统的内在变化状况。系统介绍了FvCB模型的建立、发展过程和拟合方法等基本理论,综述了该模型在叶片光合生理对光、CO_2、水、温度和N营养等环境因子变化的响应机制中的应用研究。为进一步完善FvCB模型并更好地理解叶片活体内光合系统对环境因子变化的响应机制,未来拟加强以下研究:1)羧化速率与光合电子传递速率之间的联系;2)叶肉导度的具体组分及其对FvCB模型参数估计的影响;3)叶片气孔导度和叶肉导度对环境因子变化的调控机制。
DOIURL [本文引用: 1]
为研究不同生活型植物的光合能力及其叶片光合机构,采用直角双曲线修正模型和C3植物FvCB模型对7种木本植物和4种草本植物的CO2响应曲线进行拟合,并对不同木本植物、不同草本植物和2种生活型植物的最大净光合速率(Pn max)、Rubisco酶最大羧化速率(Vc max)、最大电子传递速率(Jmax)、光合暗呼吸速率(Rd)和叶肉阻力(rm)等参数进行比较.结果表明:7种木本植物Pn max大小顺序为乌桕、苎麻〉润楠、海桐〉青冈、苦槠、娜塔栎;乌桕、苎麻、润楠和海桐的Vc max显著大于青冈和苦槠;Jmax)大小顺序为乌桕〉苎麻、海桐〉娜塔栎、苦槠和青冈;润楠和苦槠的rm显著大于乌桕、海桐和苎麻.商陆的Pn max显著大于藿香蓟和土牛膝;4种草本植物的Vc max无显著差异;商陆的Jmax显著大于藿香蓟;龙葵和土牛膝的rm显著大于藿香蓟;商陆的Rd显著大于藿香蓟和土牛膝.木本植物的Pn max、Vc max、Jmax和rm光合参数均显著大于草本植物,但二者的Rd无显著差异.不同物种之间以及2种生活型植物光合能力的差异主要是由叶片内部Rubisco酶羧化能力、电子传递能力和叶肉阻力等差异引起的.
DOIURL [本文引用: 1]
为研究不同生活型植物的光合能力及其叶片光合机构,采用直角双曲线修正模型和C3植物FvCB模型对7种木本植物和4种草本植物的CO2响应曲线进行拟合,并对不同木本植物、不同草本植物和2种生活型植物的最大净光合速率(Pn max)、Rubisco酶最大羧化速率(Vc max)、最大电子传递速率(Jmax)、光合暗呼吸速率(Rd)和叶肉阻力(rm)等参数进行比较.结果表明:7种木本植物Pn max大小顺序为乌桕、苎麻〉润楠、海桐〉青冈、苦槠、娜塔栎;乌桕、苎麻、润楠和海桐的Vc max显著大于青冈和苦槠;Jmax)大小顺序为乌桕〉苎麻、海桐〉娜塔栎、苦槠和青冈;润楠和苦槠的rm显著大于乌桕、海桐和苎麻.商陆的Pn max显著大于藿香蓟和土牛膝;4种草本植物的Vc max无显著差异;商陆的Jmax显著大于藿香蓟;龙葵和土牛膝的rm显著大于藿香蓟;商陆的Rd显著大于藿香蓟和土牛膝.木本植物的Pn max、Vc max、Jmax和rm光合参数均显著大于草本植物,但二者的Rd无显著差异.不同物种之间以及2种生活型植物光合能力的差异主要是由叶片内部Rubisco酶羧化能力、电子传递能力和叶肉阻力等差异引起的.
DOIURLPMID [本文引用: 2]
The protein content of seeds determines their nutritive value, downstream processing properties and market value. Up to 95% of seed protein is derived from amino acids that are exported to the seed after degradation of existing protein in leaves, but the pathways responsible for this nitrogen metabolism are poorly defined. The enzyme pyruvate,orthophosphate dikinase (PPDK) interconverts pyruvate and phosphoenolpyruvate, and is found in both plastids and the cytosol in plants. PPDK plays a cardinal role in C4 photosynthesis, but its role in the leaves of C3 species has remained unclear. We demonstrate that both the cytosolic and chloroplastic isoforms of PPDK are up-regulated in naturally senescing leaves. Cytosolic PPDK accumulates preferentially in the veins, while chloroplastic PPDK also accumulates in mesophyll cells. Analysis of microarrays and labelling patterns after feeding 13C-labelled pyruvate indicated that PPDK functions in a pathway that generates the transport amino acid glutamine, which is then loaded into the phloem. In Arabidopsis thaliana, over-expression of PPDK during senescence can significantly accelerate nitrogen remobilization from leaves, and thereby increase rosette growth rate and the weight and nitrogen content of seeds. This indicates an important role for cytosolic PPDK in the leaves of C3 plants, and allows us to propose a metabolic pathway that is responsible for production of transport amino acids during natural leaf senescence. Given that increased seed size and nitrogen content are desirable agronomic traits, and that efficient remobilization of nitrogen within the plant reduces the demand for fertiliser applications, PPDK and the pathway in which it operates are targets for crop improvement.
DOIURL [本文引用: 2]
Diurnal time courses of net CO 2 assimilation rates, stomatal conductance and light-driven electron fluxes were measured in situ on attached leaves of 30-year-old Turkey oak trees ( Quercus cerris L.) under natural summer conditions in central Italy. Combined measurements of gas exchange and chlorophyll a fluorescence under low O 2 concentrations allowed the demonstration of a linear relationship between the photochemical efficiency of PSII (fluorescence measurements) and the apparent quantum yield of gross photosynthesis (gas exchange). This relationship was used under normal O 2 to compute total light-driven electron fluxes, and to partition them into fractions used for RuBP carboxylation or RuBP oxygenation. This procedure also yielded an indirect estimate of the rate of photorespiration in vivo . The time courses of light-driven electron flow, net CO 2 assimilation and photorespiration paralleled that of photosynthetic photon flux density, with important afternoon deviations as soon as a severe drought stress occurred, whereas photochemical efficiency and maximal fluorescence underwent large but reversible diurnal decreases. The latter observation indicated the occurrence of a large non-photochemical energy dissipation at PSII. We estimated that less than 60% of the total photosynthetic electron flow was used for carbon assimilation at midday, while about 40% was devoted to photorespiration. The rate of carbon loss by photorespiration ( R 1 ) reached mean levels of 56% of net assimilation rates. The potential application of this technique to analysis of the relative contributions of thermal de-excitation at PSII and photorespiratory carbon recycling in the protection of photosynthesis against stress effects is discussed.
DOIURLPMID [本文引用: 1]
In the challenge to increase photosynthetic rate per leaf area mathematical models of photosynthesis can be used to help interpret gas exchange measurements made under different environmental conditions and predict underlying photosynthetic biochemistry. To do this successfully it is important to improve the modelling of temperature dependencies of CO2 assimilation and gain better understanding of internal CO2 diffusion limitations. Despite these shortcomings steady-state models of photosynthesis provide simple easy to use tools for thought experiments to explore photosynthetic pathway changes such as redirecting photorespiratory CO2, inserting bicarbonate pumps into C3 chloroplasts or inserting C4 photosynthesis into rice. Here a number of models derived from the C3 model by Farquhar, von Caemmerer and Berry are discussed and compared.
In: Sage RF, Monson R eds.
[本文引用: 1]
URL [本文引用: 2]
在植物光合作用的光反应中,由叶绿素等色素分子收集的光能通过光系统I(PSI)和光系统II(PSII)驱动了光合电子传递进程。当电子最终被传递到高能化合物NADPH和ATP中,完成光能到化学能的转换并为固碳反应提供能量的过程,被称为线形电子流(LEF)。而电子仅绕着PSI传递,通过形成跨膜质子梯度而驱动ATP酶合成ATP的过程,被称为循环电子流(CEF)。近年来,CEF的催化机制、生理作用和调控机制等吸引了研究者们的关注,特别是反向遗传学的发展,为更深入理解循环电子传递带来契机。本文综述了CEF的研究历史和最新进展,并从植物逆境适应以及植物进化等角度进一步分析了CEF的形成与生理意义,为今后的相关研究提供参考。
URL [本文引用: 2]
在植物光合作用的光反应中,由叶绿素等色素分子收集的光能通过光系统I(PSI)和光系统II(PSII)驱动了光合电子传递进程。当电子最终被传递到高能化合物NADPH和ATP中,完成光能到化学能的转换并为固碳反应提供能量的过程,被称为线形电子流(LEF)。而电子仅绕着PSI传递,通过形成跨膜质子梯度而驱动ATP酶合成ATP的过程,被称为循环电子流(CEF)。近年来,CEF的催化机制、生理作用和调控机制等吸引了研究者们的关注,特别是反向遗传学的发展,为更深入理解循环电子传递带来契机。本文综述了CEF的研究历史和最新进展,并从植物逆境适应以及植物进化等角度进一步分析了CEF的形成与生理意义,为今后的相关研究提供参考。
DOIURL [本文引用: 3]
光合作用对光和CO2响应模型是研究植物生理和植物生态学的重要工具,可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO2响应模型的研究进展和存在的问题,并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO2固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成,任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换,而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应,即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中,可能是今后光合作用对光响应机理模型的发展方向。
DOIURL [本文引用: 3]
光合作用对光和CO2响应模型是研究植物生理和植物生态学的重要工具,可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO2响应模型的研究进展和存在的问题,并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO2固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成,任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换,而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应,即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中,可能是今后光合作用对光响应机理模型的发展方向。
DOIURL [本文引用: 2]
利用低氧法(2% O_2)研究了大豆叶片光呼吸速率(R_p)对光强和CO_2浓度的响应。结果表明:当光合有效辐射强度(PAR)小于600μmol·m~(-2)·s~(-1)时,大豆叶片的R_p随光强的升高而几乎直线增加;当PAR约为1200μmol·m~(-2)·s~(-1)时,R_p达到最大值(12.69·mol CO_2·m~(-2)·s~(-1)),随后R_p随PAR的升高呈下降趋势;构建的光呼吸速率与光强的方程式拟合结果表明,大豆叶片最大光呼吸速率为13.42·mol CO_2·m~(-2)·s~(-1),其对应的光强为1207.74·mol·m~(-2)·s~(-1),该拟合值与实际测量值极为吻合(P0.05);当PAR一定(2000μmol·m~(-2)·s~(-1))时,随着CO_2浓度的增加(0~1200μmol·mol~(-1)),大豆叶片的R_p呈先升高后下降变化,在600μmol·mol~(-1)时达到最大值(9.97·mol CO_2·m~(-2)·s~(-1));构建的光呼吸速率与CO_2浓度的方程式拟合结果表明,大豆叶片最大光呼吸速率为10.21·mol CO_2·m~(-2)·s~(-1),其对应的外界CO_2浓度为625.74·mol·mol~(-1)。该拟合值也与实际测量值极为吻合(P"0.05)。本文所构建的方程式可较好地拟合光呼吸速率对不同光强和不同CO_2浓度的响应,这对定量研究光呼吸提供了强有力的手段。
DOIURL [本文引用: 2]
利用低氧法(2% O_2)研究了大豆叶片光呼吸速率(R_p)对光强和CO_2浓度的响应。结果表明:当光合有效辐射强度(PAR)小于600μmol·m~(-2)·s~(-1)时,大豆叶片的R_p随光强的升高而几乎直线增加;当PAR约为1200μmol·m~(-2)·s~(-1)时,R_p达到最大值(12.69·mol CO_2·m~(-2)·s~(-1)),随后R_p随PAR的升高呈下降趋势;构建的光呼吸速率与光强的方程式拟合结果表明,大豆叶片最大光呼吸速率为13.42·mol CO_2·m~(-2)·s~(-1),其对应的光强为1207.74·mol·m~(-2)·s~(-1),该拟合值与实际测量值极为吻合(P0.05);当PAR一定(2000μmol·m~(-2)·s~(-1))时,随着CO_2浓度的增加(0~1200μmol·mol~(-1)),大豆叶片的R_p呈先升高后下降变化,在600μmol·mol~(-1)时达到最大值(9.97·mol CO_2·m~(-2)·s~(-1));构建的光呼吸速率与CO_2浓度的方程式拟合结果表明,大豆叶片最大光呼吸速率为10.21·mol CO_2·m~(-2)·s~(-1),其对应的外界CO_2浓度为625.74·mol·mol~(-1)。该拟合值也与实际测量值极为吻合(P"0.05)。本文所构建的方程式可较好地拟合光呼吸速率对不同光强和不同CO_2浓度的响应,这对定量研究光呼吸提供了强有力的手段。
DOIURLPMID [本文引用: 1]
Day respiration (Rd) is an important parameter in leaf ecophysiology. It is difficult to measure directly and is indirectly estimated from gas exchange (GE) measurements of the net photosynthetic rate (A), commonly using the Laisk method or the Kok method. Recently a new method was proposed to estimateRdindirectly from combined GE and chlorophyll fluorescence (CF) measurements across a range of low irradiances. Here this method is tested for estimatingRdin five C3and one C4crop species. Values estimated by this new method agreed with those by the Laisk method for the C3species. The Laisk method, however, is only valid for C3species and requires measurements at very low CO2levels. In contrast, the new method can be applied to both C3and C4plants and at any CO2level. TheRdestimates by the new method were consistently somewhat higher than those by the Kok method, because using CF data corrects for errors due to any non-linearity betweenAand irradiance of the used data range. Like the Kok and Laisk methods, the new method is based on the assumption thatRdvaries little with light intensity, which is still subject to debate. Theoretically, the new method, like the Kok method, works best for non-photorespiratory conditions. As CF information is required, data for the new method are usually collected using a small leaf chamber, whereas the Kok and Laisk methods use only GE data, allowing the use of a larger chamber to reduce the noise-to-signal ratio of GE measurements.
Chlorophyll fluorescence: A probe of photosynthesis in vivo
1
2008
... 光合作用是植物(包括光合细菌)利用光能, 将CO2或H2S合成有机物并释放O2的过程.当光照射植物叶片时, 光能被捕光色素分子吸收后由基态跃迁到激发态, 处于激发态上的激子有3条退激发途径(
The CO2 concentrating function of C4 photosynthesis: A biochemical model
1
1978
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
Coupled photosynthesis stomatal model for leaves of C4 plants
1
1992
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
The rate of nitrite reduction in leaves as indicated by O2 and CO2 exchange during photosynthesis
1
2011
... 由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异.其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(
Limitation of net CO2 assimilation rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica L. and Castanea sativa Mill.)
1
1995
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
Changes in the chloroplastic CO2 concentration explain much of the observed Kok effect: A model
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
籽粒苋C4型磷酸烯醇式丙酮酸羧化酶基因的克隆和表达
2
2011
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
... ;
籽粒苋C4型磷酸烯醇式丙酮酸羧化酶基因的克隆和表达
2
2011
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
... ;
Relationships between leaf conductance to CO2 diffusion and photosynthesis in micropropagated grapevine plants, before and after ex vitro acclimatization
1
2006
... 从光合碳同化电子流分配来看, 当CO2浓度为0 mmol·mol-1时, 根据公式(3)可知, 如不考虑暗呼吸CO2回收利用, 玉米和千穗谷分配到C同化的电子流分别为0.96和1.12 mmol·m-2·s-1 (
Oxygen processing in photosynthesis: Regulation and signaling
1
2000
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
C4 photosynthesis: a unique blend of modified biochemistry, anatomy and ultrastructure
1
1987
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
籽粒苋C4关键酶丙酮酸磷酸双激酶基因的原核表达及酶活性测定
2
2017
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
... ;
籽粒苋C4关键酶丙酮酸磷酸双激酶基因的原核表达及酶活性测定
2
2017
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
... ;
Irrungen, Wirrungen? The Mehler reaction in relation to cyclic electron transport in C3 plants
2
2002
... 由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异.其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
四种作物光下暗呼吸速率降低的原因
3
2014
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 由
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
四种作物光下暗呼吸速率降低的原因
3
2014
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 由
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants
1
1999
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
转玉米C4光合途径pepc、ppdk、nadp-me基因拟南芥光合特性对强光胁迫的反应
1
2017
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
转玉米C4光合途径pepc、ppdk、nadp-me基因拟南芥光合特性对强光胁迫的反应
1
2017
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
FvCB生物化学光合模型及A-Ci曲线测定
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
FvCB生物化学光合模型及A-Ci曲线测定
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
光强对4种亚热带森林植物光合电子传递向光呼吸分配的影响
1
2000
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
光强对4种亚热带森林植物光合电子传递向光呼吸分配的影响
1
2000
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
Estimation of photorespiratory carbon dioxide recycling during photosynthesis
1
1999
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
Respiration in the light measured by 12CO2 emission in 13CO2 atmosphere in maize leaves
1
2001
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(
Alternative electron flows (water-water cycle and cyclic electron flow around PSI) in photosynthesis: Molecular mechanisms and physiological functions
1
2010
... 由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异.其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(
Cyclic electron flow within PSII functions in intact chloroplasts from spinach leaves
2
2002
... 由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异.其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
Auxiliary electron transport pathways in chloroplasts of micro algae
1
2010
... 由于植物的光合电子流与碳同化密切相关, 由此可推测J对CO2的响应模型应该与模型I相似, 但也存在较大的差异.其原因是: 在测量植物光合作用和J对CO2响应的数据时, 因一直有测量光强的存在, 所以即使光合仪叶室中的CO2浓度为0 μmol·mol-1, 此时仍然存在假电子循环等其他消耗光合电子流的路径(
Biochemical model of C3 photosynthesis applied to wheat at different temperatures
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
基于FvCB模型的叶片光合生理对环境因子的响应研究进展
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
基于FvCB模型的叶片光合生理对环境因子的响应研究进展
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
基于FvCB 模型的几种草本和木本植物光合生理生化特性
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
基于FvCB 模型的几种草本和木本植物光合生理生化特性
1
2017
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
Cytosolic pyruvate, orthophosphate dikinase functions in nitrogen remobilization during leaf senescence and limits individual seed growth and nitrogen content
2
2010
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
... (
In situ estimation of net CO2 assimilation, photosynthetic electron flow and photorespiration in Tukey oak (Q. cerris L.) leaves: Diurnal cycles under different levels of water supply
2
1995
... C3植物参与碳还原的光合电子流(Jc)(
... 当CO2浓度为380 mmol·mol-1时, 根据考虑暗呼吸CO2的回收利用而计算出的玉米其他途径所分配电子流(14.49 mmol·m-2·s-1)仅占其相应Jmax (262.41 mmol·m-2·s-1)的5.52%.由此可见, 在玉米中除光合碳同化外的其他路径, 如围绕PSII的电子循环(
Steady-state models of photosynthesis
1
2013
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
Modeling of C4 photosynthesis
1
... 当用饱和光照射植物时, 人们主要关注光合作用对CO2浓度的响应(Ac-C曲线)问题, 且已经有了较为成熟的模型用于描述这些过程, 其中最为成熟的模型是由Farquhar、von Caemmerer和Berry提出的生化模型(以下简称FvCB生化模型).该模型经过30多年的发展, 已经成为被广泛应用的模型(
植物光合作用循环电子传递的研究进展
2
2017
... 光合作用是植物(包括光合细菌)利用光能, 将CO2或H2S合成有机物并释放O2的过程.当光照射植物叶片时, 光能被捕光色素分子吸收后由基态跃迁到激发态, 处于激发态上的激子有3条退激发途径(
... 在植物光合作用的原初反应过程中, 由捕光色素分子收集的光能共振激发传递到PSII和PSI后产生电荷分离, 形成光合电子流.这些电子被电子受体接受后依次传递, 最终电子被传递到高能化合物NADPH和ATP中, 完成了光能到化学能的转换并为碳同化和光呼吸提供能量(
植物光合作用循环电子传递的研究进展
2
2017
... 光合作用是植物(包括光合细菌)利用光能, 将CO2或H2S合成有机物并释放O2的过程.当光照射植物叶片时, 光能被捕光色素分子吸收后由基态跃迁到激发态, 处于激发态上的激子有3条退激发途径(
... 在植物光合作用的原初反应过程中, 由捕光色素分子收集的光能共振激发传递到PSII和PSI后产生电荷分离, 形成光合电子流.这些电子被电子受体接受后依次传递, 最终电子被传递到高能化合物NADPH和ATP中, 完成了光能到化学能的转换并为碳同化和光呼吸提供能量(
光合作用对光和CO2响应模型的研究进展
3
2010
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 本文拟构建的J对CO2的响应模型, 是建立在光合作用对CO2响应的新模型(
... 式中, Ac为净光合速率, αc为光合作用对CO2响应曲线的初始斜率, βc和γc为两个与CO2浓度无关的系数, C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci), Rl为光下呼吸速率, 它与光强存在非线性响应关系(
光合作用对光和CO2响应模型的研究进展
3
2010
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 本文拟构建的J对CO2的响应模型, 是建立在光合作用对CO2响应的新模型(
... 式中, Ac为净光合速率, αc为光合作用对CO2响应曲线的初始斜率, βc和γc为两个与CO2浓度无关的系数, C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci), Rl为光下呼吸速率, 它与光强存在非线性响应关系(
大豆叶片光呼吸对光强和CO2浓度的响应
2
2017
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 式中, Ac为净光合速率, αc为光合作用对CO2响应曲线的初始斜率, βc和γc为两个与CO2浓度无关的系数, C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci), Rl为光下呼吸速率, 它与光强存在非线性响应关系(
大豆叶片光呼吸对光强和CO2浓度的响应
2
2017
... 目前, C4植物叶片的J对CO2浓度的响应遵循怎样的变化规律尚未见报道.而该问题的阐明对定量研究C4植物光合电子流的分配具有重要的意义.此外, 已有的研究结果表明, 在测量植物的Ac-C曲线时, 当环境CO2浓度设置为0 μmol·mol-1时, 其J因植物种类的不同而有差异(
... 式中, Ac为净光合速率, αc为光合作用对CO2响应曲线的初始斜率, βc和γc为两个与CO2浓度无关的系数, C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci), Rl为光下呼吸速率, 它与光强存在非线性响应关系(
Evaluating a new method to estimate the rate of leaf respiration in the light by analysis of combined gas exchange and chlorophyll fluorescence measurements
1
2011
... 植物的光合作用是光合能量和底物相互偶联的氧化还原反应(

{kind=link}
{kind=link}
{kind=link}
{kind=link}