Simulation on the light-response curves of electron transport rate of Quercus variabilis and Robinia pseudoacacia leaves in the Xiaolangdi area, China
LI Li-Yuan1, LI Jun2, TONG Xiao-Juan,1,*, MENG Ping3, ZHANG Jin-Song3, ZHANG Jing-Ru11 College of Forestry, Beijing Forestry University, Beijing 100083, China 2 Key Laboratory of Water Cycle and Related Land Surface Processes, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 3 Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration, Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, China
Supported by the National Natural Science Foundation of China.31570617 Supported by the National Natural Science Foundation of China.31100322 the Fundamental Research Funds for the Central Universities.YX2011-19
Abstract Aims The objectives are to compare the merits and demerits of rectangular hyperbola, nonrectangular hyperbola and the electron transport rate light-response Ye model, to investigate the difference of electron transport rate between the shaded and sunlit leaves, and to discuss the influence of bioenvironmental factors on the characteristic parameters of electron transport rate. Methods The light-response (J-I) curves of electron transport rate were measured by the LI-6400XT fluorescence measurement system in Quercus variabilis and Robinia pseudoacacia plantations in north China. The rectangular hyperbola, nonrectangular hyperbola and the Ye model were used to simulate electron transport rate of the light-response curves. Important findings The results showed that the determination coefficient of the J-I curves fitted by three models were more than 0.96. Compared with the rectangular hyperbola and nonrectangular hyperbola, the determination coefficient of the Ye model was the highest (> 0.99). The dynamic downregulation of photosystem II and the saturated light intensity (Isat) cannot be simulated and obtained by the rectangular hyperbola model and the nonrectangular hyperbola model. The maximum electron transport rate (Jmax) obtained by the rectangular hyperbola model was obviously higher than the measured one. The dynamic downregulation of photosystem II was well simulated by the Ye model. The Jmax and Isat values obtained by the Ye model were close to the measured ones. The Jmax values of the shaded leaves of Quercus variabilis and Robinia pseudoacacia were 25.0% and 18.0% lower than the sunlit leaves, respectively. The Isat values of the sunlit leaves of Q. variabilis and R. pseudoacacia were 26.0% and 10.1% higher than those of the shaded leaves. Jmax of Q. variabilis and R. pseudoacacia was correlated with temperature. Isat of R. pseudoacacia was correlated with temperature, soil water content and net photosynthetic rate. The initial slope (α) values of the J-I curves for Q. variabilis and R. pseudoacacia had significant negative relationships with net photosynthetic rate. Keywords:Quercus variabilis; Robinia pseudoacacia;Ye model;shaded leaf;sunlit leaf;electron transport rate
PDF (1217KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 李理渊, 李俊, 同小娟, 孟平, 张劲松, 张静茹. 黄河小浪底栓皮栎、刺槐叶片电子传递速率-光响应的模拟. 植物生态学报[J], 2018, 42(10): 1009-1021 DOI:10.17521/cjpe.2018.0063 LI Li-Yuan, LI Jun, TONG Xiao-Juan, MENG Ping, ZHANG Jin-Song, ZHANG Jing-Ru. Simulation on the light-response curves of electron transport rate of Quercus variabilis and Robinia pseudoacacia leaves in the Xiaolangdi area, China. Chinese Journal of Plant Ecology[J], 2018, 42(10): 1009-1021 DOI:10.17521/cjpe.2018.0063
新窗口打开|下载原图ZIP|生成PPT 图1不同模型拟合的栓皮栎、刺槐叶片电子传递速率-光响应曲线(平均值±标准误差, n = 3)。A-D, 栓皮栎6、7、8和9月。E-H, 刺槐6、7、8和9月。
Fig. 1Electron transport rate light-response curves fitted by different models (rectangular hyperbola, nonrectangular hyperbola and Ye model) for Q. variabilis and R. pseudoacacia (mean ± SE, n = 3). A-D, Q. variabilis in June, July, August and September. E-H, R. pseudoacacia in June, July, August and September.
Table 1 表1 表1栓皮栎、刺槐叶片电子传递速率的特征参数(平均值±标准误差, n = 3) Table 1Characteristic parameters of electron transport rate of Quercus variabilis and Robinia pseudoacacia leaves (mean ± SE, n = 3)
树种 Species
月份 Month
模型 Model
α
Isat (μmol·m-2·s-1)
Jmax (μmol·m-2·s-1)
R2
栓皮栎 Quercus variabilis
6月 June
实测 Measured
-
1 000
65.51
-
直角双曲线模型 Rectangular hyperbola model
0.46 ± 0.06
-
69.94 ± 5.85
0.963
非直角双曲线模型 Nonrectangular hyperbola model
0.27 ± 0.03
-
64.41 ± 4.61
0.985
叶子飘模型 Ye model
0.35 ± 0.03
1 042.28 ± 96.81
65.23 ± 6.45
0.999
7月 July
实测 Measured
-
1200
84.11
-
直角双曲线模型 Rectangular hyperbola model
0.54 ± 0.05
-
89.48 ± 2.60
0.971
非直角双曲线模型 Nonrectangular hyperbola model
0.29 ± 0.04
-
79.41 ± 1.48
0.994
叶子飘模型 Ye model
0.41 ± 0.04
1 072.50 ± 46.54
82.04 ± 2.18
0.996
8月 August
实测 Measured
-
1 200
101.98
-
直角双曲线模型 Rectangular hyperbola model
0.50 ± 0.02
-
117.96 ± 7.03
0.973
非直角双曲线模型 Nonrectangular hyperbola model
0.34 ± 0.08
-
105.33 ± 2.62
0.995
叶子飘模型 Ye model
0.38 ± 0.02
1 184.70 ± 38.30
103.60 ± 5.17
0.996
9月 September
实测 Measured
-
1 000
47.55
-
直角双曲线模型 Rectangular hyperbola model
0.57 ± 0.06
-
49.94 ± 2.81
0.968
非直角双曲线模型 Nonrectangular hyperbola model
0.28 ± 0.02
-
47.02 ± 2.60
0.992
叶子飘模型 Ye model
0.44 ± 0.03
990.56 ± 231.06
48.20 ± 1.19
0.994
刺槐 Robinia pseudoacacia
6月 June
实测 Measured
-
1 000
63.57
-
直角双曲线模型 Rectangular hyperbola model
0.47 ± 0.03
-
68.74 ± 5.31
0.970
非直角双曲线模型 Nonrectangular hyperbola model
0.25 ± 0.01
-
62.43 ± 4.79
0.987
叶子飘模型 Ye model
0.36 ± 0.02
1 043.59 ± 60.56
63.67 ± 4.14
0.997
7月 July
实测 Measured
-
1500
178.89
-
直角双曲线模型 Rectangular hyperbola model
0.50 ± 0.01
-
234.74 ± 2.78
0.978
非直角双曲线模型 Nonrectangular hyperbola model
0.30 ± 0.01
-
183.35 ± 1.76
0.998
叶子飘模型 Ye model
0.37 ± 0.01
1 437.14 ± 24.26
182.83 ± 1.21
0.998
8月 August
实测 Measured
-
1 300
127.78
-
直角双曲线模型 Rectangular hyperbola model
0.46 ± 0.02
-
156.02 ± 8.13
0.976
非直角双曲线模型 Nonrectangular hyperbola model
0.28 ± 0.02
-
129.45 ± 6.91
0.997
叶子飘模型 Ye model
0.36 ± 0.02
1 349.56 ± 28.31
128.97 ± 6.53
0.997
9月 September
实测 Measured
-
1 200
121.21
-
直角双曲线模型 Rectangular hyperbola model
0.50 ± 0.02
-
142.64 ± 5.45
0.975
非直角双曲线模型 Nonrectangular hyperbola model
0.30 ± 0.01
-
118.55 ± 5.08
0.995
叶子飘模型 Ye model
0.38 ± 0.01
1 275.05 ± 24.58
121.47 ± 4.87
0.998
α, initial slope; Isat, saturated irradiance; Jmax, maximu electron transport rate; R2, the determined coefficient. The measured values in the table were obtained from the measured curves. α, 初始斜率; Isat, 饱和光强; Jmax, 最大电子传递速率; R2, 决定系数。表中的实测值是由实测曲线得到。
Table 2 表2 表2阴生叶、阳生叶电子传递速率的特征参数(叶子飘模型拟合)(平均值±标准误差, n = 9) Table 2Characteristic parameters of electron transport rate of shaded and sunlit leaves (fitted by Ye model) (mean ± SE, n = 9)
树种 Species
月份 Month
叶位 Leaf position
拟合/实测 Fitted/measured
α
Isat (μmol·m-2·s-1)
Jmax (μmol·m-2·s-1)
R2
栓皮栎 Quercus variabilis
6月 June
阴生叶 Shaded leaf
拟合 Fitted
0.39 ± 0.07
1 019.31 ± 55.64
55.17 ± 7.31
0.993
实测 Measured
-
1 000
55.49
-
阳生叶 Sunlit leaf
拟合 Fitted
0.32 ± 0.01
1 075.41 ± 139.41
75.66 ± 7.80
0.997
实测 Measured
-
1 000
75.53
-
7月 July
阴生叶 Shaded leaf
拟合 Fitted
0.44 ± 0.03
981.87 ± 179.95
69.97 ± 7.69
0.994
实测 Measured
-
1 000
68.65
-
阳生叶 Sunlit leaf
拟合 Fitted
0.41 ± 0.07
1 172.54 ± 172.76
95.22 ± 4.41
0.996
实测 Measured
-
1 500
94.99
-
8月 August
阴生叶 Shaded leaf
拟合 Fitted
0.40 ± 0.04
1 092.35 ± 97.59
86.36 ± 7.47
0.996
实测 Measured
-
1 000
85.40
-
阳生叶 Sunlit leaf
拟合 Fitted
0.38 ± 0.02
1 246.39 ± 9.25
121.32 ± 5.96
0.997
实测 Measured
-
1 200
120.60
-
9月 September
阴生叶 Shaded leaf
拟合 Fitted
0.48 ± 0.08
736.50 ± 86.71
44.07 ± 0.08
0.995
实测 Measured
-
1 000
43.59
-
阳生叶 Sunlit leaf
拟合 Fitted
0.42 ± 0.06
1 141.58 ± 119.35
53.75 ± 4.93
0.991
实测 Measured
-
1 800
53.54
-
刺槐 Robinia pseudoacacia
6月 June
阴生叶 Shaded leaf
拟合 Fitted
0.37 ± 0.02
1 045.23 ± 74.27
57.01 ± 6.10
0.993
实测 Measured
-
1 000
57.47
-
阳生叶 Sunlit leaf
拟合 Fitted
0.37 ± 0.02
1 047.57 ± 76.42
70.38 ± 4.50
0.997
实测 Measured
-
1 000
69.67
-
7月 July
阴生叶 Shaded leaf
拟合 Fitted
0.38 ± 0.01
1 265.91 ± 33.74
168.49 ± 5.99
0.997
实测 Measured
-
1 000
166.75
-
阳生叶 Sunlit leaf
拟合 Fitted
0.36 ± 0.02
1 654.66 ± 48.58
200.67 ± 8.92
0.999
实测 Measured
-
1 800
199.69
-
8月 August
阴生叶 Shaded leaf
拟合 Fitted
0.36 ± 0.02
1 316.81 ± 39.36
112.22 ± 7.87
0.997
实测 Measured
-
1 200
111.20
-
阳生叶 Sunlit leaf
拟合 Fitted
0.37 ± 0.02
1 372.99 ± 25.97
145.84 ± 9.12
0.996
实测 Measured
-
1 800
144.59
-
9月 September
阴生叶 Shaded leaf
拟合 Fitted
0.40 ± 0.01
1 240.05 ± 50.69
112.46 ± 5.47
0.995
实测 Measured
-
1 000
111.86
-
阳生叶 Sunlit leaf
拟合 Fitted
0.37 ± 0.01
1 303.29 ± 46.66
130.79 ± 5.67
0.998
实测 Measured
-
1 200
133.03
-
α, initial slope; Isat, saturated irradiance; Jmax, maximu electron transport rate; R2, the determined coefficient. The measured values in the table were obtained from the measured curves. α, 初始斜率; Isat, 饱和光强; Jmax, 最大电子传递速率; R2, 决定系数。表中的实测值是由实测曲线得到。
新窗口打开|下载原图ZIP|生成PPT 图2阴生叶、阳生叶电子传递速率-光响应曲线比较(叶子飘模型拟合)(平均值±标准误差, n = 9)。A-D, 栓皮栎6、7、8和9月, E-H, 刺槐6、7、8和9月。
Fig. 2Electron transport rate light-response curves of shaded and sunlit leaves (fitted by Ye model)(mean ± SE, n = 9). A-D, Q. variabilis in June, July, August and September. E-H, R. pseudoacacia in June, July, August and September.
Table 3 表3 表3栓皮栎、刺槐叶片电子传递速率特征参数与环境/生物因子的相关关系 Table 3Correlations of characteristic parameters of electron transport rate and bioenvironmental factors in Quercus variabilis and Robinia pseudoacacia
栓皮栎 Quercus variabilis
刺槐 Robinia pseudoacacia
α
Isat
Jmax
α
Isat
Jmax
Ta
-0.571
0.480
0.902**
-0.477
0.801*
0.806*
SWC
-0.364
0.856
0.953*
-0.344
0.965*
0.912
Pn
-0.800*
0.542
0.745*
-0.719*
0.764*
0.661
Chl
-0.293
0.873
0.961*
-0.435
0.933
0.871
*, p < 0.05;**, p < 0.01. α, Isat, Jmax see Table 2. Ta, air temperature; SWC, soil water content; Pn, net photosynthetic rate; Chl, chlorophyll content. *, p < 0.05;**, p < 0.01。α, Isat, Jmax同表2。Ta, 气温; SWC, 土壤含水量; Pn, 净光合速率; Chl, 叶绿素含量。
AmbrosioND, ArenaC, SantoAV ( 2006). Temperature response of photosynthesis, excitation energy dissipation and alternative electron sinks to carbon assimilation in Beta vulgaris L Environmental and Experimental Botany, 55, 248-257. DOIURL [本文引用: 1] Photosynthesis, excitation energy dissipation and alternative electron sinks to carbon assimilation at different temperatures (5–35 °C range) were studied in plants of Beta vulgaris L. grown outdoors at springtime. Gas exchange and chlorophyll fluorescence measurements were performed in plants exposed to different temperatures for two hours at saturating irradiance (1000 μmol photons m 612 s 611). The photosynthetic rate decreased significantly either by lowering the temperature below 25 °C or by increasing it above 25 °C. At low temperatures (5, 10 and 15 °C) a significant decrease of electron transport rate (ETR) was found and processes alternative to CO 2 fixation, as sinks of electrons, occurred but they were not affected by the temperature. However, at 5 °C a low and statistically significant photorespiratory rate was observed and the non-photochemical quenching (NPQ) was the highest. At higher temperature (30–35 °C) ETR saturated and hence non-assimilative processes, alternative to CO 2 fixation, sustained the photochemical activity. Among processes consuming O 2 photorespiration increased at 30–35 °C, while processes non-consuming O 2 were non-affected. By increasing the temperature up to 30–35 °C NPQ values did not show appreciable differences compared to 25 °C. The obtained results indicate that in B. vulgaris plants the thermal dissipation represents the main dissipative process of the excess excitation energy at low temperatures. On the contrary, at high temperatures (30–35 °C) the main dissipation of the excess excitation energy is due to alternative pathways to CO 2 assimilation.
BakerNR ( 2008). Chlorophyll fluorescence: A probe of photosynthesis in vivo Annual Review of Plant Biology, 59, 89-113. DOIURL [本文引用: 1]
BalyEC ( 1935). The kinetics of photosynthesis Proceedings of the Royal Society of London Series B: Biological Sciences, 117, 218-239. DOIURL [本文引用: 2]
BernacchiCJ, MorganPB, OrtDR, LongSP ( 2005). The growth of soybean under free air [CO2] enrichment (FACE) stimulates photosynthesis while decreasing in vivo Rubisco capacity Planta, 220, 434-446. [本文引用: 1]
BradingP, WarnerME, DaveyP, SmithDJ, AchterbergEP, SuggettDJ ( 2011). Differential effects of ocean acidification on growth and photosynthesis among phylotypes of Symbiodinium (Dinophyceae) Limnology and Oceanography, 56, 927-938. DOIURL [本文引用: 1] We investigated the effect of elevated partial pressure of CO2 (pCO2) on the photosynthesis and growth of four phylotypes (ITS2 types A1, A13, A2, and B1) from the genus Symbiodinium, a diverse dinoflagellate group that is important, both free-living and in symbiosis, for the viability of cnidarians and is thus a potentially important model dinoflagellate group. The response of Symbiodinium to an elevated pCO2 was phylotype-specific. Phylotypes A1 and B1 were largely unaffected by a doubling in pCO2; in contrast, the growth rate of A13 and the photosynthetic capacity of A2 both increased by ~ 60. In no case was there an effect of ocean acidification (OA) upon respiration (dark- or light-dependent) for any of the phylotypes examined. Our observations suggest that OA might preferentially select among free-living populations of Symbiodinium, with implications for future symbioses that rely on algal acquisition from the environment (i.e., horizontal transmission). Furthermore, the carbon environment within the host could differentially affect the physiology of different Symbiodinium phylotypes. The range of responses we observed also highlights that the choice of species is an important consideration in OA research and that further investigation across phylogenetic diversity, for both the direction of effect and the underlying mechanism(s) involved, is warranted. 2011, by the American Society of Limnology and Oceanography, Inc.
ChazdonRL, PearcyRW, LeeDW, FetcherN, MulkeySS ( 1996). Photosynthetic responses of tropical forest plants to contrasting light environments In: Mulkey SS, Chazdon RL, Smith AP eds. Tropical Forest Plant Ecophysiology. Chapman and Hall, New York. 5-55. DOIURL [本文引用: 1] Across the complex matrix of microsites that compose tropical forests, light availability varies more dramatically than any other single plant resource. On a sunny day, instantaneous measurements of photosynthetically active radiation range over 3 orders of magnitude, from less than 10 mol m -2 s -1 in closed-canopy understory of mature forests to well over 1000 mol m -2 s -1 in exposed microsites of gaps and large clearings, or at the top of the forest canopy (Chazdon & Fetcher, 1984b; Figure 1.1). Among the environmental factors that influence plant growth and survival in tropical forests, light availability is likely to be the resource most frequently limiting growth, survival, and reproduction (Chazdon, 1988; Fetcher, Oberbauer & Chazdon, 1994). Photosynthetic utilization of light is therefore a major component of the regeneration responses of forest species within the larger context of forest dynamics and succession.
ChenY, XuDQ ( 2006). Two patterns of leaf photosynthetic responses to irradiance transition from saturating to limiting one in some plant species New Phytologist, 169, 789-798. DOIURLPMID [本文引用: 1] 61 Plants often regulate the amount and size of light-harvesting antenna (LHCII) to maximize photosynthesis at low light and avoid photodamage at high light. 61 Gas exchange, 77 K chlorophyll fluorescence, photosystem II (PSII) electron transport as well as LHCII protein were measured in leaves irradiated at different light intensities. 61 After irradiance transition from saturating to limiting one leaf photosynthetic rate in some species such as soybean and rice declined first to a low level, then increased slowly to a stable value (V pattern), while in other species such as wheat and pumpkin it dropped immediately to a stable value (L pattern). Saturating pre-irradiation led to significant declines of both 77 K fluorescence parameter F685/F735 and light-limited PSII electron transport rate in soybean but not in wheat leaves, indicating that some LHCIIs dissociate from PSII in soybean but not in wheat leaves. 61 The L pattern of LHCII-decreased rice mutant and the V pattern of its wild type demonstrate that the V pattern is linked to dissociation/reassociation of some LHCIIs from/to PSII.
EichelmannH, OjaV, PetersonRB, LaiskA ( 2011). The rate of nitrite reduction in leaves as indicated by O2 and CO2 exchange during photosynthesis Journal of Experimental Botany, 62, 2205-2215. DOIURLPMID [本文引用: 1] Light response (at 300 ppm CO(2) and 10-50 ppm O(2) in N(2)) and CO(2) response curves [at absorbed photon fluence rate (PAD) of 550 μmol m(-2) s(-1)] of O(2) evolution and CO(2) uptake were measured in tobacco (Nicotiana tabacum L.) leaves grown on either NO(3)(-) or NH(4)(+) as N source and in potato (Solanum tuberosum L.), sorghum (Sorghum bicolor L. Moench), and amaranth (Amaranthus cruentus L.) leaves grown on NH(4)NO(3). Photosynthetic O(2) evolution in excess of CO(2) uptake was measured with a stabilized zirconia O(2) electrode and an infrared CO(2) analyser, respectively, and the difference assumed to represent the rate of electron flow to acceptors alternative to CO(2), mainly NO(2)(-), SO(4)(2-), and oxaloacetate. In NO(3)(-)-grown tobacco, as well as in sorghum, amaranth, and young potato, the photosynthetic O(2)-CO(2) flux difference rapidly increased to about 1 μmol m(-2) s(-1) at very low PADs and the process was saturated at 50 μmol quanta m(-2) s(-1). At higher PADs the O(2)-CO(2) flux difference continued to increase proportionally with the photosynthetic rate to a maximum of about 2 μmol m(-2) s(-1). In NH(4)(+)-grown tobacco, as well as in potato during tuber filling, the low-PAD component of surplus O(2) evolution was virtually absent. The low-PAD phase was ascribed to photoreduction of NO(2)(-) which successfully competes with CO(2) reduction and saturates at a rate of about 1 μmol O(2) m(-2) s(-1) (9% of the maximum O(2) evolution rate). The high-PAD component of about 1 μmol O(2) m(-2) s(-1), superimposed on NO(2)(-) reduction, may represent oxaloacetate reduction. The roles of NO(2)(-), oxaloacetate, and O(2) reduction in the regulation of ATP/NADPH balance are discussed.
Ferrier-PagèsC, TambuttéE, ZamoumT, SegondsN, MerlePL, BensoussanN, AllemandD, GarrabouJ, TambuttéS ( 2009). Physiological response of the symbiotic gorgonian Eunicella singularis to a longterm temperature increase Journal of Experimental Biology, 212, 3007-3015. DOIURLPMID [本文引用: 1] http://jeb.biologists.org/cgi/doi/10.1242/jeb.031823
FilaG, BadeckFW, MeyerS, CerovicZ, GhashghaieJ ( 2006). Relationships between leaf conductance to CO2 diffusion and photosynthesis in micropropagated grapevine plants, before and after ex vitro acclimatization Journal of Experimental Botany, 57, 2687-2695. DOIURLPMID [本文引用: 1] Abstract In vitro-cultured plants typically show a low photosynthetic activity, which is considered detrimental to subsequent ex vitro acclimatization. Studies conducted so far have approached this problem by analysing the biochemical and photochemical aspects of photosynthesis, while very little attention has been paid to the role of leaf conductance to CO(2) diffusion, which often represents an important constraint to CO(2) assimilation in naturally grown plants. Mesophyll conductance, in particular, has never been determined in in vitro plants, and no information exists as to whether it represents a limitation to carbon assimilation during in vitro growth and subsequent ex vitro acclimatization. In this study, by means of simultaneous gas exchange and chlorophyll fluorescence measurements, the stomatal and mesophyll conductance to CO(2) diffusion were assessed in in vitro-cultured plants of the grapevine rootstock '41B' (Vitis vinifera 'Chasselas'xVitis berlandieri), prior to and after ex vitro acclimatization. Their impact on electron transport rate partitioning and on limitation of potential net assimilation rate was analysed. In vitro plants had a high stomatal conductance, 155 versus 50 mmol m(-2) s(-1) in acclimatized plants, which ensured a higher CO(2) concentration in the chloroplasts, and a 7% higher electron flow to the carbon reduction pathway. The high stomatal conductance was counterbalanced by a low mesophyll conductance, 43 versus 285 mmol m(-2) s(-1), which accounted for a 14.5% estimated relative limitation to photosynthesis against 2.1% estimated in acclimatized plants. It was concluded that mesophyll conductance represents an important limitation for in vitro plant photosynthesis, and that in acclimatization studies the correct comparison of photosynthetic activity between in vitro and acclimatized plants must take into account the contribution of both stomatal and mesophyll conductance.
FlexasJ, BarónM, BotaJ, DucruetJM, GalléA, GalmésJ, JiménezM, PouA, Ribas-CarbóM, SajnaniC, TomàsM, MedranoH ( 2009). Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri var. rupestris) Journal of Experimental Botany, 60, 2361-2377. DOIURLPMID [本文引用: 1] The hybrid Richter-110 (Vitis berlandieri Vitis rupestris) has the reputation of being a genotype strongly adapted to drought. A study was performed with plants of R-110 subjected to sustained water-withholding to induce acclimation to two different levels of water stress, followed by rewatering to induce recovery. The goal was to analyse how photosynthesis is regulated during acclimation to water stress and recovery. In particular, the regulation of stomatal conductance (g
Govindjee ( 2002). A role for a light-harvesting antenna complex of photosystem II in photoprotection Plant Cell, 14, 1663-1668. DOIURLPMID [本文引用: 1] A role for a light-harvesting antenna complex of photosystem II in photoprotection GOVINDJEE The Plant Cell 14, 1663-1668, 2002
GuoW, ZhanSY, YinH, LiXY, LüX, YangL, WangY ( 2016). Effect of enhanced UV-B radiation on photosynthetic electron transport and light response characteristics of japonica Journal of Nanjing Agricultural University, 39, 603-610. DOIURL [本文引用: 1] [目的]光合作用对UV-B辐射增强的响应对评价粳稻在全球气候变化背景下的适应能力至关重要。本文旨在分析UV-B辐射增强对粳稻剑叶光合作用内在机制的影响。[方法]以北方粳稻品种‘沈农265’为试验材料,采用盆栽形式,试验设对照组CK(采用自然光照射),处理组T1、T2分别在自然光照基础上增加UV-B辐射1.05和2.1 W·m^-2,每个处理重复3或6次。选取粳稻幼苗移栽于无孔试验桶中,紫外灯照射时间为08:00—16:00,阴雨天不进行UV-B处理,直至成熟收获。试验期间对粳稻各生长时期进行剑叶活体测量,测定光合电子流传递分配、光合响应曲线及参数,包括非环式光合电子流中参与碳还原或光呼吸的那一部分电子流Jc、Jo,表观量子效率(AQY),最大净光合速率(P(max))和暗呼吸速率(Rd)的变化特征。[结果]对光合电子传递的影响:UV-B辐射增强在剑叶各个生长期Jc和Jo(非环式电子流中参与光呼吸和碳还原的电子流)均降低,Jo/JF与Jo/Jc(光呼吸所占的比例)均增加,说明UV-B增强降低了光合电子进入碳还原的部分;UV-B处理在剑叶各个生长期也降低了Vo(Rubisco的氧化速率)和Vc(Rubisco的羧化速率),但Vo/Vc呈上升趋势,且与UV-B辐射强度成正比,说明UV-B辐射增强降低了Rubisco(核酮糖-1,5-二磷酸加氧酶)的羧化速率。对光合响应特性的影响:剑叶各生长期在UV-B处理下,CK与T1、T2的光合响应曲线趋势相同,净光合速率(Pn)从大到小的处理为对照组、低剂量UV-B辐射、高剂量辐射;除成熟期,LSP(光饱和点)均降低,在灌浆期降幅最大,除灌浆期,LCP(光补偿点)均上升,在孕穗期升幅最大;AQY总体呈下降趋势;P(max)显著下降,T2对P(max)的抑制程度大于T1;除成熟期,Rd(暗呼吸速率)都呈增加趋势,灌浆期增幅最高。[结论]UV-B辐射增强总体上增加了剑叶光呼吸耗能,降低了光合电子 [ 郭巍, 战莘晔, 殷红, 李雪莹, 吕晓, 杨璐, 王一 ( 2016). UV-B辐射增强对粳稻光合电子传递与光响应特性的影响 南京农业大学学报, 39, 603-610.] DOIURL [本文引用: 1] [目的]光合作用对UV-B辐射增强的响应对评价粳稻在全球气候变化背景下的适应能力至关重要。本文旨在分析UV-B辐射增强对粳稻剑叶光合作用内在机制的影响。[方法]以北方粳稻品种‘沈农265’为试验材料,采用盆栽形式,试验设对照组CK(采用自然光照射),处理组T1、T2分别在自然光照基础上增加UV-B辐射1.05和2.1 W·m^-2,每个处理重复3或6次。选取粳稻幼苗移栽于无孔试验桶中,紫外灯照射时间为08:00—16:00,阴雨天不进行UV-B处理,直至成熟收获。试验期间对粳稻各生长时期进行剑叶活体测量,测定光合电子流传递分配、光合响应曲线及参数,包括非环式光合电子流中参与碳还原或光呼吸的那一部分电子流Jc、Jo,表观量子效率(AQY),最大净光合速率(P(max))和暗呼吸速率(Rd)的变化特征。[结果]对光合电子传递的影响:UV-B辐射增强在剑叶各个生长期Jc和Jo(非环式电子流中参与光呼吸和碳还原的电子流)均降低,Jo/JF与Jo/Jc(光呼吸所占的比例)均增加,说明UV-B增强降低了光合电子进入碳还原的部分;UV-B处理在剑叶各个生长期也降低了Vo(Rubisco的氧化速率)和Vc(Rubisco的羧化速率),但Vo/Vc呈上升趋势,且与UV-B辐射强度成正比,说明UV-B辐射增强降低了Rubisco(核酮糖-1,5-二磷酸加氧酶)的羧化速率。对光合响应特性的影响:剑叶各生长期在UV-B处理下,CK与T1、T2的光合响应曲线趋势相同,净光合速率(Pn)从大到小的处理为对照组、低剂量UV-B辐射、高剂量辐射;除成熟期,LSP(光饱和点)均降低,在灌浆期降幅最大,除灌浆期,LCP(光补偿点)均上升,在孕穗期升幅最大;AQY总体呈下降趋势;P(max)显著下降,T2对P(max)的抑制程度大于T1;除成熟期,Rd(暗呼吸速率)都呈增加趋势,灌浆期增幅最高。[结论]UV-B辐射增强总体上增加了剑叶光呼吸耗能,降低了光合电子
HarleyPC, ThomasRB, ReynoldsJF, StrainetBR ( 1992). Modelling photosynthesis of cotton grown in elevated CO2 Plant, Cell & Environment, 15, 271-282. DOIURL [本文引用: 1] Cotton plants were grown in CO 2 -controlled growth chambers in atmospheres of either 35 or 65 Pa CO 2 . A widely accepted model of C 3 leaf photosynthesis was parameterized for leaves from both CO 2 treatments using non-linear least squares regression techniques, but in order to achieve reasonable fits, it was necessary to include a phosphate limitation resulting from inadequate triose phosphate utilization. Despite the accumulation of large amounts of starch (>50 g m 2 ) in the high CO 2 plants, the photosynthetic characteristics of leaves in both treatments were similar, although the maximum rate of Rubisco activity (Vc max ), estimated from A versus C i response curves measured at 29 C, was 10% lower in leaves from plants grown in high CO 2 . The relationship between key model parameters and total leaf N was linear, the only difference between CO 2 treatments being a slight reduction in the slope of the line relating Vc max to leaf N in plants grown at high CO 2 . Stomatal conductance of leaves of plants grown and measured at 65 Pa CO 2 was approximately 32% lower than that of plants grown and measured at 35 Pa. Because photosynthetic capacity of leaves grown in high CO 2 was only slightly less than that of leaves grown in 35 Pa CO 2 , net photosynthesis measured at the growth CO 2 , light and temperature conditions was approximately 25% greater in leaves of plants grown in high CO 2 , despite the reduction in leaf conductance. Greater assimilation rate was one factor allowing plants grown in high CO 2 to incorporate 30% more biomass during the first 36 d of growth.
HarrisonWG, PlattT ( 1986). Photosynthesis-irradiance relationships in polar and temperate phytoplankton populations Polar Biology, 5, 153-164. DOIURL [本文引用: 1] Analyses of some 700 photosynthesis-irradiance (P-I) experiments made on natural marine phytoplankton assemblages in over six years of field work are summarized. Observed variations in P-I parameters are discussed in relation to geographical location, sample depth and temporal cycles (diel to interannual). Results suggest that P-I characteristics and therefore regional primary production are largely under physical control; temperature and light appear to be the most important environmental covariables.
HuWH, YeZP, YanXH, YangXS ( 2017). PSII function and intrinsic characteristics of light-harvesting pigment molecules for sun- and shading-leaf in Magnolia grandiflora during overwintering Bulletin of Botanical Research, 37, 281-287. URL [本文引用: 1] 捕光色素分子的内禀特性不仅决定了光能的吸收与传递,也将影响到激发能向光化学反应、热耗散和叶绿素荧光的分配。本文采用叶绿素荧光技术和光合电子流对光响应机理模型,研究了越冬期广玉兰(Magnolia grandiflora)阳生叶和阴生叶两种不同光环境下叶片PSⅡ功能及其捕光色素分子内禀特性的差异,以探索广玉兰越冬的光保护策略。结果表明:越冬期低温导致叶片轻微光抑制的发生,全光照加剧了阳生叶光抑制程度,而弱光环境有利于阴生叶光抑制的恢复。阳生叶可通过降低叶绿素含量和捕光色素分子数量以减少对光能的吸收,并且具有较强的光化学和热耗散能力以保护光合机构免受低温强光伤害。而阴生叶虽然其光化学反应能力相对较弱,但具有较强的热耗散能力,可有效地保护其免受短时曝露在强光下的伤害。 [ 胡文海, 叶子飘, 闫小红, 杨旭升 ( 2017). 越冬期广玉兰阳生叶和阴生叶PSII功能及捕光色素分子内禀特性的比较研究 植物研究, 37, 281-287.] URL [本文引用: 1] 捕光色素分子的内禀特性不仅决定了光能的吸收与传递,也将影响到激发能向光化学反应、热耗散和叶绿素荧光的分配。本文采用叶绿素荧光技术和光合电子流对光响应机理模型,研究了越冬期广玉兰(Magnolia grandiflora)阳生叶和阴生叶两种不同光环境下叶片PSⅡ功能及其捕光色素分子内禀特性的差异,以探索广玉兰越冬的光保护策略。结果表明:越冬期低温导致叶片轻微光抑制的发生,全光照加剧了阳生叶光抑制程度,而弱光环境有利于阴生叶光抑制的恢复。阳生叶可通过降低叶绿素含量和捕光色素分子数量以减少对光能的吸收,并且具有较强的光化学和热耗散能力以保护光合机构免受低温强光伤害。而阴生叶虽然其光化学反应能力相对较弱,但具有较强的热耗散能力,可有效地保护其免受短时曝露在强光下的伤害。
JoaoS, RuiS ( 2004). Can chlorophyll fluorescence be used to estimate photosynthetic production in the seagrass Zosteranoltii Marine Biology & Ecology, 307, 207-216. DOIURL [本文引用: 1] Seagrass photosynthesis is usually measured in laboratory experiments, following oxygen evolution in closed chambers. Pulse amplitude modulated (PAM) fluorometry constitutes an alternative and non-intrusive method of measuring photosynthesis in the field. Validation of electron transport rate (ETR) measurements as reliable estimators of actual photosynthetic production requires that a significant linear relationship between oxygen production and ETR is demonstrated, and also that a 0.25 molar ratio between these two measures is verified. In this work, both parameters were measured simultaneously in laboratory experiments, over a range of light intensities, under well-defined and controlled conditions. A linear relationship was observed between the average rates of oxygen production and the electron transport rates for Zostera noltii (Hornemann) obtained at several irradiances. The molar ratio found between oxygen production and ETR was 0.15±0.02, lower than the theoretically expected value of 0.25. The use of PAM fluorescence as a valid proxy for photosynthetic production was validated for the range of 35–490 μmol photons m 612 s 611 (PAR), under the assumption that the electron sinks responsible for the molar ratio deviation remain constant in similar experimental conditions.
KangHJ, LiH, TaoYL, ZhangHL, QuanW, OuyangZ ( 2015). Discussion on simultaneous measurements of leaf gas exchange and chlorophyll fluorescence for estimating photosynthetic electron allocation Acta Ecologica Sinica, 35, 1217-1224. DOIURL [本文引用: 2] 光合电子流分配是植物光合控制的一个重要环节。然而,传统电子流分配的计算方法存在诸多问题尚未引起人们的注意,如:(1)低估了光呼吸每释放一个CO2分子所消耗的电子数;(2)混淆了相对电子传递速率和绝对电子传递速率;(3)忽略了除碳同化和光呼吸外的其他电子流分配途径;(4)难以准确获取光下暗呼吸速率值,从而导致碳同化电子流(Jc)及光呼吸速率(R。)的不准确估算等。以小麦和大豆气体交换与荧光同步测量数据为例,结果表明大豆电子传递速率与碳同化两者对光强的响应一致性较好,同时达到最大值;而小麦的一致性相对较差,说明电子传递速率和碳同化并非完全一致,推测认为有可能与作物对同化产物输出的模式不同有关。通过光呼吸速率换算出的电子流(12×Rp)与实际测量电子流(△J0)之间存在较大的差异;另外,传统方法估算出的光呼吸速率(估算值)与光呼吸测量值之间也存在较大差异,分析认为这主要是由于绝对光合速率与相对电子传递速率之间差异造成。 [ 康华靖, 李红, 陶月良, 张海利, 权伟, 欧阳竹 ( 2015). 气体交换与荧光同步测量估算植物光合电子流的分配 生态学报, 35, 1217-1224.] DOIURL [本文引用: 2] 光合电子流分配是植物光合控制的一个重要环节。然而,传统电子流分配的计算方法存在诸多问题尚未引起人们的注意,如:(1)低估了光呼吸每释放一个CO2分子所消耗的电子数;(2)混淆了相对电子传递速率和绝对电子传递速率;(3)忽略了除碳同化和光呼吸外的其他电子流分配途径;(4)难以准确获取光下暗呼吸速率值,从而导致碳同化电子流(Jc)及光呼吸速率(R。)的不准确估算等。以小麦和大豆气体交换与荧光同步测量数据为例,结果表明大豆电子传递速率与碳同化两者对光强的响应一致性较好,同时达到最大值;而小麦的一致性相对较差,说明电子传递速率和碳同化并非完全一致,推测认为有可能与作物对同化产物输出的模式不同有关。通过光呼吸速率换算出的电子流(12×Rp)与实际测量电子流(△J0)之间存在较大的差异;另外,传统方法估算出的光呼吸速率(估算值)与光呼吸测量值之间也存在较大差异,分析认为这主要是由于绝对光合速率与相对电子传递速率之间差异造成。
LawlorDW, CornicG ( 2002). Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants Plant, Cell & Environment, 25, 275-294. DOIURLPMID [本文引用: 1] Summary Experimental studies on CO 2 assimilation of mesophytic C3 plants in relation to relative water content (RWC) are discussed. Decreasing RWC slows the actual rate of photosynthetic CO 2 assimilation (A) and decreases the potential rate (A pot ). Generally, as RWC falls from c . 100 to c. 75%, the stomatal conductance (g s ) decreases, and with it A. However, there are two general types of relation of A pot to RWC, which are called Type 1 and Type 2. Type 1 has two main phases. As RWC decreases from 100 to c. 75%, A pot is unaffected, but decreasing stomatal conductance (g s ) results in smaller A, and lower CO 2 concentration inside the leaf (C i ) and in the chloroplast (C c ), the latter falling possibly to the compensation point. Down-regulation of electron transport occurs by energy quenching mechanisms, and changes in carbohydrate and nitrogen metabolism are considered acclimatory, caused by low C i and reversible by elevated CO 2 . Below 75% RWC, there is metabolic inhibition of A pot , inhibition of A then being partly (but progressively less) reversible by elevated CO 2 ; g s regulates A progressively less, and C i and CO 2 compensation point, rise. It is suggested that this is the true stress phase, where the decrease in A pot is caused by decreased ATP synthesis and a consequent decreased synthesis of RuBP. In the Type 2 response, A pot decreases progressively at RWC 100 to 75%, with A being progressively less restored to the unstressed value by elevated CO 2 . Decreased g s leads to a lower C i and C c but they probably do not reach compensation point: g s becomes progressively less important and metabolic limitations more important as RWC falls. The primary effect of low RWC on A pot is most probably caused by limited RuBP synthesis, as a result of decreased ATP synthesis, either through inhibition of Coupling Factor activity or amount due to increased ion concentration. Carbohydrate synthesis and accumulation decrease. Type 2 response is considered equivalent to Type 1 at RWC below c. 75%, with A pot inhibited by limited ATP and RuBP synthesis, respiratory metabolism dominates and C i and rise. The importance of inhibited ATP synthesis as a primary cause of decreasing A pot is discussed. Factors determining the Type 1 and Type 2 responses are unknown. Electron transport is maintained (but down-regulated) in Types 1 and 2 over a wide range of RWC, and a large reduced/oxidized adenylate ratio results. Metabolic imbalance results in amino acid accumulation and decreased and altered protein synthesis. These conditions profoundly affect cell functions and ultimately cause cell death. Type 1 and 2 responses may reflect differences in g s and in sensitivity of metabolism to decreasing RWC.
LiYX, ShenSH, LiL, WangXM, ZhangFC, HanXM ( 2012). Effects of soil moisture on leaf gas exchange and chlorophyll fluorescence parameters of winter wheat during its late growth stage Chinese Journal of Ecology, 31, 74-80. URL [本文引用: 1] 通过防雨棚小区栽培,控制土壤供水系数(Kw)分别为0.8、0.6、0.4、0.2,以自然状况下的小区为对照(CK),研究土壤水分条件对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响。结果表明:Kw为0.6处理的冬小麦叶片叶绿素含量与0.8处理接近,且显著高于其他处理(P〈0.05);Kw为0.6处理对冬小麦叶片的气孔导度和蒸腾速率有轻度抑制,但其光合速率却高于0.8处理,而Kw为0.2处理的光合速率、气孔导度及蒸腾速率均为最低;气孔限制值在Kw为0.4处理下最高,其次为0.2处理,0.8处理下最低;冬小麦叶片的表观量子效率在Kw为0.4处理下最高,光补偿点总体上随着土壤水分含量的降低呈下降趋势,而光饱和点及最大光合速率则以Kw为0.6处理最高,其次为0.8处理,0.2处理最低;冬小麦叶片的天线转化效率Fv′/Fm′、电子传递速率ETR、实际量子效率ФPSII及光化学猝灭qP均以Kw为0.6处理最高,其次为0.8处理,0.2处理下最低;在Kw为0.2处理下,冬小麦光合作用主要受非气孔因素限制,而在0.4处理下,则主要受气孔因素限制。 [ 李永秀, 申双和, 李丽, 汪秀敏, 张富存, 韩小梅 ( 2012). 土壤水分对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响 生态学杂志, 31, 74-80.] URL [本文引用: 1] 通过防雨棚小区栽培,控制土壤供水系数(Kw)分别为0.8、0.6、0.4、0.2,以自然状况下的小区为对照(CK),研究土壤水分条件对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响。结果表明:Kw为0.6处理的冬小麦叶片叶绿素含量与0.8处理接近,且显著高于其他处理(P〈0.05);Kw为0.6处理对冬小麦叶片的气孔导度和蒸腾速率有轻度抑制,但其光合速率却高于0.8处理,而Kw为0.2处理的光合速率、气孔导度及蒸腾速率均为最低;气孔限制值在Kw为0.4处理下最高,其次为0.2处理,0.8处理下最低;冬小麦叶片的表观量子效率在Kw为0.4处理下最高,光补偿点总体上随着土壤水分含量的降低呈下降趋势,而光饱和点及最大光合速率则以Kw为0.6处理最高,其次为0.8处理,0.2处理最低;冬小麦叶片的天线转化效率Fv′/Fm′、电子传递速率ETR、实际量子效率ФPSII及光化学猝灭qP均以Kw为0.6处理最高,其次为0.8处理,0.2处理下最低;在Kw为0.2处理下,冬小麦光合作用主要受非气孔因素限制,而在0.4处理下,则主要受气孔因素限制。
LiuJJ, LiJY, ZhangJG ( 2015). Influences of drought stress on photosynthetic characteristics and water use efficiency of 4 tree species under elevated CO2 concentration Forest Research, 28, 339-345. URL [本文引用: 1] 在不同CO2浓度(380、720μmol·mol-1)的密闭式生长箱内,对5年生油松和侧柏苗、3年生元宝枫和刺槐苗进行培养,研究CO2浓度升高与干旱胁迫对4种树苗光合特性和水分利用效率的影响。结果表明:高CO2浓度均能增加正常水分和重度干旱胁迫下4个树苗的光合速率(Pn)、胞间CO2浓度(Ci)和瞬时水分利用效率(WUEi),而降低蒸腾速率(Tr)和气孔导度(Cond);在轻度干旱和重度干旱条件下,Pn、Ci、Tr、Cond和WUEi增加,刺槐的WUEi却减少。CO2浓度增加,4个树种在同一干旱时期的碳稳定同位素比值(δ13C)减少。随着干旱胁迫加剧,不同CO2浓度下4个树种的Pn、Tr和Cond减少,而720μmol·mol-1CO2浓度下4个树种和380μmol·mol-1CO2浓度下刺槐和元宝枫的WUEi和δ13C增加,而380μmol·mol-1CO2浓度下油松和侧柏的WUEi和δ13C先增加,到重度干旱时又下降。CO2浓度增加与干旱胁迫的交互作用减弱了干旱胁迫或者CO2浓度增加中的某一因子对气孔变化的敏感性,使得气孔变化缓慢,延迟了水分胁迫的发生。 [ 刘娟娟, 李吉跃, 张建国 ( 2015). 高CO2浓度和干旱胁迫对4种树苗光合特性的影响 林业科学研究, 28, 339-345.] URL [本文引用: 1] 在不同CO2浓度(380、720μmol·mol-1)的密闭式生长箱内,对5年生油松和侧柏苗、3年生元宝枫和刺槐苗进行培养,研究CO2浓度升高与干旱胁迫对4种树苗光合特性和水分利用效率的影响。结果表明:高CO2浓度均能增加正常水分和重度干旱胁迫下4个树苗的光合速率(Pn)、胞间CO2浓度(Ci)和瞬时水分利用效率(WUEi),而降低蒸腾速率(Tr)和气孔导度(Cond);在轻度干旱和重度干旱条件下,Pn、Ci、Tr、Cond和WUEi增加,刺槐的WUEi却减少。CO2浓度增加,4个树种在同一干旱时期的碳稳定同位素比值(δ13C)减少。随着干旱胁迫加剧,不同CO2浓度下4个树种的Pn、Tr和Cond减少,而720μmol·mol-1CO2浓度下4个树种和380μmol·mol-1CO2浓度下刺槐和元宝枫的WUEi和δ13C增加,而380μmol·mol-1CO2浓度下油松和侧柏的WUEi和δ13C先增加,到重度干旱时又下降。CO2浓度增加与干旱胁迫的交互作用减弱了干旱胁迫或者CO2浓度增加中的某一因子对气孔变化的敏感性,使得气孔变化缓慢,延迟了水分胁迫的发生。
LongSP, BernacchiCJ ( 2003). Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error Journal of Experimental Botany, 54, 2393-2401. DOIURLPMID [本文引用: 1] The principles, equipment and procedures for measuring leaf and canopy gas exchange have been described previously as has chlorophyll fluorescence. Simultaneous measurement of the responses of leaf gas exchange and modulated chlorophyll fluorescence to light and CO2 concentration now provide a means to determine a wide range of key biochemical and biophysical limitations on photosynthesis in vivo. Here the mathematical frameworks and practical procedures for determining these parameters in vivo are consolidated. Leaf CO2 uptake (A) versus intercellular CO2 concentration (Ci) curves may now be routinely obtained from commercial gas exchange systems. The potential pitfalls, and means to avoid these, are examined. Calculation of in vivo maximum rates of ribulose-1,5-bisphosphate (RuBP) carboxylase/oxygenase (Rubisco) carboxylation (VC,max), electron transport driving regeneration of RuBP (Jmax), and triose-phosphate utilization (VTPU) are explained; these three parameters are now widely assumed to represent the major limitations to light-saturated photosynthesis. Precision in determining these in intact leaves is improved by the simultaneous measurement of electron transport via modulated chlorophyll fluorescence. The A/Ci response also provides a simple practical method for quantifying the limitation that stomata impose on CO2 assimilation. Determining the rate of photorespiratory release of oxygen (Rl) has previously only been possible by isotopic methods, now, by combining gas exchange and fluorescence measurements, Rl may be determined simply and routinely in the field. The physical diffusion of CO2 from the intercellular air space to the site of Rubisco in C3 leaves has long been suspected of being a limitation on photosynthesis, but it has commonly been ignored because of the lack of a practical method for its determination. Again combining gas exchange and fluorescence provides a means to determine mesophyll conductance. This method is described and provides insights into the magnitude and basis of this limitation.
MartinsSCV, GalmésJ, MolinsA, DamattaFM ( 2013). Improving the estimation of mesophyll conductance to CO2: On the role of electron transport rate correction and respiration Journal of Experimental Botany, 64, 3285-3298. DOIURL [本文引用: 1]
NelsonN, YocumCF ( 2006). Structure and function of photosystems I and II Annual Review of Plant Biology, 57, 521-565. DOIURLPMID [本文引用: 1] Oxygenic photosynthesis, the principal converter of sunlight into chemical energy on earth, is catalyzed by four multi-subunit membrane-protein complexes: photosystem I (PSI), photosystem II (PSII), the cytochrome b(6)fcomplex, and F-ATPase. PSI generates the most negative redox potential in nature and largely determines the global amount of enthalpy in living systems. PSII generates an oxidant whose redox potential is high enough to enable it to oxidize H(2)O, a substrate so abundant that it assures a practically unlimited electron source for life on earth. During the last century, the sophisticated techniques of spectroscopy, molecular genetics, and biochemistry were used to reveal the structure and function of the two photosystems. The new structures of PSI and PSII from cyanobacteria, algae, and plants has shed light not only on the architecture and mechanism of action of these intricate membrane complexes, but also on the evolutionary forces that shaped oxygenic photosynthesis.
ÖrdögA, WodalaB, RózsavögyiT, TariI, HorváthF ( 2013). Regulation of guard cell photosynthetic electron transport by nitric oxide Journal of Experimental Botany, 64, 1357-1366. DOIURLPMID [本文引用: 1] Abstract Nitric oxide (NO) is one of the key elements in the complex signalling pathway leading to stomatal closure by inducing reversible protein phosphorylation and Ca(2+) release from intracellular stores. As photosynthesis in guard cells also contributes to stomatal function, the aim of this study was to explore the potential role of NO as a photosynthetic regulator. This work provides the first description of the reversible inhibition of the effect of NO on guard cell photosynthetic electron transport. Pulse amplitude modulation (PAM) chlorophyll fluorescence measurements on individual stomata of peeled abaxial epidermal strips indicated that exogenously applied 450nM NO rapidly increases the relative fluorescence yield, followed by a slow and constant decline. It was found that NO instantly decreases photochemical fluorescence quenching coefficients (qP and qL), the operating quantum efficiency of photosystem II ( PSII), and non-photochemical quenching (NPQ) to close to zero with different kinetics. NO caused a decrease in NPQ, which is followed by a slow and continuous rise. The removal of NO from the medium surrounding the epidermal strips using a rapid liquid perfusion system showed that the effect of NO on qP and PSII, and thus on the linear electron transport rate through PSII (ETR), is reversible, and the constant rise in NPQ disappears, resulting in a near steady-state value. The reversible inhibition by NO of the ETR could be restored by bicarbonate, a compound known to compete with NO for one of the two coordination sites of the non-haem iron (II) in the QAFe(2+)QB complex.
OsborneCP, WytheEJ, IbrahimDG, GilbertME, RipleyBS ( 2008). Low temperature effects on leaf physiology and survivorship in the C3 and C4 subspecies of Alloteropsis semialata Journal of Experimental Botany, 59, 1743-1754. [本文引用: 1]
PavlovicA, SlovákováL, PandolfiC, MancusoS ( 2011). On the mechanism underlying photosynthetic limitation upon trigger hair irritation in the carnivorous plant venus flytrap (Dionaea muscipula Ellis) Journal of Experimental Botany, 62, 1991-2000. DOIURLPMID [本文引用: 1] ABSTRACT Mechanical stimulation of trigger hairs on the adaxial surface of the trap of Dionaea muscipula leads to the generation of action potentials and to rapid leaf movement. After rapid closure secures the prey, the struggle against the trigger hairs results in generation of further action potentials which inhibit photosynthesis. A detailed analysis of chlorophyll a fluorescence kinetics and gas exchange measurements in response to generation of action potentials in irritated D. muscipula traps was used to determine the 'site effect' of the electrical signal-induced inhibition of photosynthesis. Irritation of trigger hairs and subsequent generation of action potentials resulted in a decrease in the effective photochemical quantum yield of photosystem II (Φ(PSII)) and the rate of net photosynthesis (A(N)). During the first seconds of irritation, increased excitation pressure in photosystem II (PSII) was the major contributor to the decreased Φ(PSII). Within 65165min, non-photochemical quenching (NPQ) released the excitation pressure at PSII. Measurements of the fast chlorophyll a fluorescence transient (O-J-I-P) revealed a direct impact of action potentials on the charge separation-recombination reactions in PSII, although the effect seems to be small rather than substantial. All the data presented here indicate that the main primary target of the electrical signal-induced inhibition of photosynthesis is the dark reaction, whereas the inhibition of electron transport is only a consequence of reduced carboxylation efficiency. In addition, the study also provides valuable data confirming the hypothesis that chlorophyll a fluorescence is under electrochemical control.
PlattT, GallegosCL, HarrisonWG ( 1980). Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton Journal of Marine Research, 38, 687-701. [本文引用: 1]
QianQ, QuLJ, YuanM, WangXJ, YangWC, WangT, KongHZ, JiangGM, ChongK ( 2013). Research advances on plant science in China in 2012 Chinese Bulletin of Botany, 48, 231-287. DOIURL [本文引用: 1] Plant science in China has been developing rapidly in 2012. Chinese scientists reported a lot of original research findings of high level in various aspects of plant biology, including the research into plant genome, the signaling pathway of plant innate immune resistance, the mechanism of DNA demethylation and DNA double-strand break repair in plants, and so on. This review aims to provide an overall picture of plant research in China and highlights some of the important findings in 2012. [ 钱前, 瞿礼嘉, 袁明, 王小菁, 杨维才, 王台, 孔宏智, 蒋高明, 种康 ( 2013). 2012年中国植物科学若干领域重要研究进展 植物学报, 48, 231-287.] DOIURL [本文引用: 1] Plant science in China has been developing rapidly in 2012. Chinese scientists reported a lot of original research findings of high level in various aspects of plant biology, including the research into plant genome, the signaling pathway of plant innate immune resistance, the mechanism of DNA demethylation and DNA double-strand break repair in plants, and so on. This review aims to provide an overall picture of plant research in China and highlights some of the important findings in 2012.
QiuEF, ChenZM, HongW, ZhengYS, HuangBL, YangZQ ( 2006). Comparison on ecophysiological characteristics between sun and shade leaves in different age Dendrocalamus latiflorus Acta Ecologica Sinica, 26, 3297-3301. DOIURL [本文引用: 2] 麻竹是中国重要的大型经济竹种,其栽培已从过去河滩、四旁零散种植发展到规模化培育,通过山地麻竹发笋期 内不同年龄植株阴阳叶养分和代谢动态的比较研究,结果表明麻竹阳叶氮素、磷素浓度比阴叶高,但钾素浓度阳叶低于阴叶;从发笋初期至末期阴阳叶氮、磷、钾素 浓度都呈逐渐减少的变化趋势,阴阳叶氮、磷、钾素浓度差异逐渐减小;阳叶在净光合速率、暗呼吸速率、CO2补偿点、光补偿点、光饱和点等方面较阴叶高,光 呼吸较低,但不同年龄麻竹之间各指标变化有所不同. [ 邱尔发, 陈卓梅, 洪伟, 郑郁善, 黄宝龙, 杨主泉 ( 2006). 不同年龄麻竹阴阳叶生态生理特性 生态学报, 26, 3297-3301.] DOIURL [本文引用: 2] 麻竹是中国重要的大型经济竹种,其栽培已从过去河滩、四旁零散种植发展到规模化培育,通过山地麻竹发笋期 内不同年龄植株阴阳叶养分和代谢动态的比较研究,结果表明麻竹阳叶氮素、磷素浓度比阴叶高,但钾素浓度阳叶低于阴叶;从发笋初期至末期阴阳叶氮、磷、钾素 浓度都呈逐渐减少的变化趋势,阴阳叶氮、磷、钾素浓度差异逐渐减小;阳叶在净光合速率、暗呼吸速率、CO2补偿点、光补偿点、光饱和点等方面较阴叶高,光 呼吸较低,但不同年龄麻竹之间各指标变化有所不同.
RalphPJ, GademannR ( 2005). Rapid light curves: A powerful tool to assess photosynthetic activity Aquatic Botany, 82, 222-237. DOIURL [本文引用: 1] Rapid light curves provide detailed information on the saturation characteristics of electron transport, as well as the overall photosynthetic performance of a plant. Rapid light curves were collected from samples of Zostera marina grown under low and high-light conditions (50 and 300 μmol photons m 612 s 611) and the distinctive patterns of RLC parameters are discussed, in terms of differential sink capacity and PSII reaction centre closure. Derived cardinal points of a rapid light curve ( α, E k and rETR max) describe the photosynthetic capacity of a seagrass leaf, its light adaptation state and its capacity to tolerate short-term changes in light. The shapes of the corresponding F and F ′ m curves also provide information on the development of the trans-thylakoid proton gradient and thermal energy dissipation. Low-light leaves showed limited photosynthetic capacity and reduced activity of non-photochemical quenching pathways, whereas photosynthesis of high light leaves were not limited and showed an elevated level of non-photochemical quenching, possibly associated with xanthophyll cycle activity. Light-dark kinetics are also discussed in relation to relaxation of non-photochemical quenching and its various components. A curve fitting model is recommended based on the double exponential decay function. In this paper, we explain the fundamental aspects of a RLC, describe how it reflects the response to light exposure of a leaf, how to interpret these curves, and how to quantitatively describe and compare RLCs.
RascherU, LiebigM, LüttgeU ( 2000). Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer onsite in the field Plant, Cell & Environment, 23, 1397-1405. DOIURL [本文引用: 1] ABSTRACT Miniaturized pulse-amplitude modulated photosynthesis yield analysers are primarily designed for measuring effective quantum yield ( F / F m ') of photosystem II under momentary ambient light conditions in the field. Although this provides important ecophysiological information, it is often necessary to learn more about the potential intrinsic capacities of leaves by measuring light-response curves. Thus, instruments provide light-curve programmes, where light intensities are increased in short intervals and instant light-response curves are recorded within a few minutes. This method can be criticized because photosynthesis will most likely not be in steady state. This technical report shows that with the appropriate precautions instant light curves can nevertheless provide reliable information about cardinal points of photosynthesis. First, the geometry of the light source of the instrument in relation to the quantum sensor must be considered and quantum sensor readings must be corrected. Second, the measurements of the light-response curves must be compared with readings of effective quantum yield of photosystem II under ambient light conditions where photosynthesis is in steady state. This may show that in the critical range of the light curves either both measurements perfectly coincide or are offset against each other by a constant value (examples are given here). In the first case results of light curves can be taken at face values, and in the second case a simple correction can be applied. With these precautions and careful interpretations instant light-response curves can be an enormous advantage in ecophysiological field work.
RenB, LiJ, TongXJ, MuYM, ZhangJS, MengP ( 2017). Simulation on photosynthetic light-response of Quercus variabilis and Robinia pseudoacacia in the southern foot of the Taihang Mountain Chinese Journal of Ecology, 36, 2206-2216. DOIURL [本文引用: 1] 植物光合作用-光响应模型研究是评价植物光合响应机制和获得光合参数的主要途径。本文以太行山南麓栓皮栎(Quercus variabilis)和刺槐(Robinia pseudoacacia)人工林为研究对象,采用LI-6400XT光合测定系统测定了这两种人工林叶片光合作用-光响应曲线,利用直角双曲线模型(RH)、非直角双曲线模型(NRH)以及直角双曲线的修正模型-叶子飘模型(YZP)对光合-光响应曲线进行了拟合,对比了3种模型的最大净光合速率P_(max)、初始光能利用率α、暗呼吸速率R_d等光合参数。结果表明,RH模型的P_(max)、α和R_d均较大,并且无法直接得到光饱和点I_s。NRH模型不能很好地模拟光抑制现象,在P_(max)的模拟上明显偏大,但在α的模拟中比较接近真实值。YZP模型则具有更好的适用性,能较好地模拟光抑制现象。对栓皮栎和刺槐阴、阳叶光合作用-光响应曲线研究发现,阳叶的P_(max)大于阴叶(栓皮栎、刺槐的阳叶P_(max)分别高出阴叶的24.41%和18.28%),阴叶的α高于阳叶(栓皮栎、刺槐的阴叶α值分别平均高出阳叶的22.08%、15.41%),阴叶的R_d也高于阳叶,尤其在刺槐上表现明显(刺槐阴叶R_d高出阳叶的7.13%)。栓皮栎P_(max)、R_d和I_s主要受相对湿度和气孔导度的影响,α主要受相对湿度的影响。刺槐的P_(max)和I_s与相对湿度和气孔导度均呈显著正相关关系。 [ 任博, 李俊, 同小娟, 母艳梅, 张劲松, 孟平 ( 2017). 太行山南麓栓皮栎和刺槐叶片光合光响应模拟 生态学杂志, 36, 2206-2216.] DOIURL [本文引用: 1] 植物光合作用-光响应模型研究是评价植物光合响应机制和获得光合参数的主要途径。本文以太行山南麓栓皮栎(Quercus variabilis)和刺槐(Robinia pseudoacacia)人工林为研究对象,采用LI-6400XT光合测定系统测定了这两种人工林叶片光合作用-光响应曲线,利用直角双曲线模型(RH)、非直角双曲线模型(NRH)以及直角双曲线的修正模型-叶子飘模型(YZP)对光合-光响应曲线进行了拟合,对比了3种模型的最大净光合速率P_(max)、初始光能利用率α、暗呼吸速率R_d等光合参数。结果表明,RH模型的P_(max)、α和R_d均较大,并且无法直接得到光饱和点I_s。NRH模型不能很好地模拟光抑制现象,在P_(max)的模拟上明显偏大,但在α的模拟中比较接近真实值。YZP模型则具有更好的适用性,能较好地模拟光抑制现象。对栓皮栎和刺槐阴、阳叶光合作用-光响应曲线研究发现,阳叶的P_(max)大于阴叶(栓皮栎、刺槐的阳叶P_(max)分别高出阴叶的24.41%和18.28%),阴叶的α高于阳叶(栓皮栎、刺槐的阴叶α值分别平均高出阳叶的22.08%、15.41%),阴叶的R_d也高于阳叶,尤其在刺槐上表现明显(刺槐阴叶R_d高出阳叶的7.13%)。栓皮栎P_(max)、R_d和I_s主要受相对湿度和气孔导度的影响,α主要受相对湿度的影响。刺槐的P_(max)和I_s与相对湿度和气孔导度均呈显著正相关关系。
ReynoldsM, FoulkesJ, FurbankR, GriffithsS, KingJ, MurchieE, ParryM, SlaferG ( 2012). Achieving yield gains in wheat Plant, Cell & Environment, 35, 1799-1823. DOIURLPMID [本文引用: 1] Wheat provides 20% of calories and protein consumed by humans. Recent genetic gains are <1% per annum (p.a.), insufficient to meet future demand. The Wheat Yield Consortium brings expertise in photosynthesis, crop adaptation and genetics to a common breeding platform. Theory suggest radiation use efficiency (RUE) of wheat could be increased 50%; strategies include modifying specificity, catalytic rate and regulation of Rubisco, up-regulating Calvin cycle enzymes, introducing chloroplast CO2 concentrating mechanisms, optimizing light and N distribution of canopies while minimizing photoinhibition, and increasing spike photosynthesis. Maximum yield expression will also require dynamic optimization of source: sink so that dry matter partitioning to reproductive structures is not at the cost of the roots, stems and leaves needed to maintain physiological and structural integrity. Crop development should favour spike fertility to maximize harvest index so phenology must be tailored to different photoperiods, and sensitivity to unpredictable weather must be modulated to reduce conservative responses that reduce harvest index. Strategic crossing of complementary physiological traits will be augmented with wide crossing, while genome-wide selection and high throughput phenotyping and genotyping will increase efficiency of progeny screening. To ensure investment in breeding achieves agronomic impact, sustainable crop management must also be promoted through crop improvement networks.
RobakowskiP ( 2005). Susceptibility to low-temperature photoinhibition in three conifers differing in successional status Tree Physiology, 25, 1151-1160. DOIURLPMID [本文引用: 1] Abstract Susceptibility to photoinhibition of the evergreen conifers Abies alba Mill., Picea abies (L.) Karst. and Pinus mugo Turra was investigated in an unheated greenhouse during winter and spring 2003. Photosynthetic performance of the seedlings was assessed by chlorophyll a fluorescence and analyses of chlorophyll and total carotenoid concentrations in needles. During winter months, maximum quantum yield of PSII photochemistry (ratio of variable to maximum fluorescence, Fv/Fm) was significantly greater in A. alba than in P. abies and P. mugo. Abies alba also sustained higher maximum apparent electron transport rate (ETRmax) than P. abies and P. mugo. Total concentrations of chlorophyll and carotenoids in needles decreased during the winter in P. mugo and P. abies, but remained stable in A. alba. For all species, Fv/Fm decreased from December until February and then increased to a maximum in April. Photoinhibition was greatest (Fv/Fm < 0.80) in all seedlings in February, the month with the lowest mean temperature. Saturating photosynthetic photon flux (PPFsat) and ETRmax were positively related to air temperature. All species had lower values of ETRmax and PPFsat in winter than in spring. Non-photochemical quenching of chlorophyll fluorescence (NPQ) was highest at low air temperatures. Differences among species in susceptibility to winter photoinhibition resulted from their specific light preferences and led to different mechanisms to cope with photoinhibitory stress. The more shade-tolerant A. alba sustained a higher photosynthetic capacity in winter than P. abies and P. mugo. Winter photoinhibition in P. abies, P. mugo and, to a lesser extent, in A. alba may reflect adaptive photoprotection of the photosynthetic apparatus in winter.
SofoA, DichiB, MontanaroG, XiloyannisC ( 2009). Photosynthetic performance and light response of two olive cultivars under different water and light regimes Photosynthetica, 47, 602-608. DOIURL [本文引用: 1] The olive tree ( Olea europaea L.) is commonly grown in the Mediterranean area, where it is adapted to resist periods characterized by severe drought and high irradiance levels. Photosynthetic efficiency (in terms of F v /F m and PSII ), photochemical (q P ) and nonphotochemical quenching (NPQ) were determined in two-year-old olive plants (cultivars Coratina and Biancolilla) grown under two different light levels (exposed plants, EP, and shaded plants, SP) during a 21-day controlled water deficit. After reaching the maximum level of drought stress, plants were rewatered for 23 days. During the experimental period, measurements of gas exchange and chlorophyll (Chl) fluorescence were carried out to study the photosynthetic performance of olive plants. The synergical effect of drought stress and high irradiance levels caused a reduction of gas exchange and photosynthetic efficiency and these decreases were more marked in EP. EP showed a higher degree of photoinhibition, a higher NPQ and a lower q P if compared to SP. Coratina was more sensitive to high light and drought stress but also showed a slower recovery during rewatering, whereas Biancolilla showed a less marked photosynthesis depression during drought and a considerable resilience during rewatering. The results confirm that photoinhibition due to high light intensity and water deficit can be an important factor that affects photosynthetic productivity in this species.
TangWZ, LiXS, HuangHY, CaiCE, HuoYZ, HePM ( 2009). Effects of different light intensity and temperature treatment on photosynthesis and chlorophyll fluorescence in Ulva linza Journal of Fisheries of China, 33, 752-769. DOIURL [本文引用: 3] 利用脉冲振幅调制叶绿素荧光仪对不同光照和温度条件下长石莼(缘管浒苔)光合作用和培养条件进行了优化.结果表明,长石莼(缘管浒苔)在25℃和72μmol/(m2·s)条件下,其Fv/Fm、Fm、Fv和α值最高,分别高达0.74、4 567:3 406和0.305,低于该点为光不饱和,高于该点为光抑制,偏离越大,下降越显著(P20>15>30>10>5>35℃,光照强度排序为72>54/108>36/162>18/216μmol/(m2·s). [ 汤文仲, 李信书, 黄海燕, 蔡春尔, 霍元子, 何培民 ( 2009). 不同光强和温度对长石莼(缘管浒苔)光合作用和叶绿素荧光参数的影响 水产学报, 33, 752-769.] DOIURL [本文引用: 3] 利用脉冲振幅调制叶绿素荧光仪对不同光照和温度条件下长石莼(缘管浒苔)光合作用和培养条件进行了优化.结果表明,长石莼(缘管浒苔)在25℃和72μmol/(m2·s)条件下,其Fv/Fm、Fm、Fv和α值最高,分别高达0.74、4 567:3 406和0.305,低于该点为光不饱和,高于该点为光抑制,偏离越大,下降越显著(P20>15>30>10>5>35℃,光照强度排序为72>54/108>36/162>18/216μmol/(m2·s).
WhiteAJ, CritchleyC ( 1999). Rapid light curves: A new fluorescence method to assess the state of the photosynthetic apparatus Photosynthesis Research, 59, 63-72. DOIURL [本文引用: 2]
XieJM, YuJH, HuangGB, FengZ ( 2011). Correlations between changes of absorption and transformation of light energy by PSII in pepper leaves and the variety tolerance under low temperature & weak light Scientia Agricultura Sinica, 44, 1855-1862. DOIURL [本文引用: 1] 【目的】研究低温弱光下辣椒叶片PSII光能吸收和转换规律,分析其与品种耐低温弱光性的相关性,筛选和建立快速、准确、无损伤的辣椒耐低温弱光性鉴定指标和方法。【方法】以耐低温弱光性不同的12个辣椒品种为试材,以冷害指数作为品种耐性鉴定的依据,应用调制荧光测定技术,采用相关分析的方法,研究低温弱光下辣椒叶片PSII光能吸收和转换规律及其与品种耐低温弱光性的关系。【结果】15℃/5℃、100μmol.m-2.s-1低温弱光处理使辣椒叶片PSⅡ的实际光能转化效率ΦPSⅡ、开放的PSⅡ有效光能转化效率Fv′/Fm′、光化学猝灭系数qP、光下最大荧光Fm′、光下最小荧光Fo′和光下最大可变荧光Fv′均降低,非光化学猝灭系数NPQ和稳态荧光Fs增大,最大光能转化效率Fv/Fm变化不显著。15℃/5℃、100μmol.m-2.s-1低温弱光处理15 d辣椒叶片的ΦPSⅡ和Fv′与辣椒的冷害指数呈极显著负相关,Fv′/Fm′、qP和Fm′呈显著负相关,NPQ呈显著正相关。【结论】15℃/5℃、100μmol.m-2.s-1低温弱光处理15 d辣椒叶片的ΦPSⅡ、Fv′/Fm′、qP、NPQ、Fm′和Fv′等叶绿素荧光参数可用于辣椒耐低温弱光性快速、准确、无损伤的鉴定指标,其中ΦPSⅡ和Fv′为最敏感的指标。 [ 颉建明, 郁继华, 黄高宝, 冯致 ( 2011). 低温弱光下辣椒叶片PSII光能吸收和转换变化及与品种耐性的关系 中国农业科学, 44, 1855-1862.] DOIURL [本文引用: 1] 【目的】研究低温弱光下辣椒叶片PSII光能吸收和转换规律,分析其与品种耐低温弱光性的相关性,筛选和建立快速、准确、无损伤的辣椒耐低温弱光性鉴定指标和方法。【方法】以耐低温弱光性不同的12个辣椒品种为试材,以冷害指数作为品种耐性鉴定的依据,应用调制荧光测定技术,采用相关分析的方法,研究低温弱光下辣椒叶片PSII光能吸收和转换规律及其与品种耐低温弱光性的关系。【结果】15℃/5℃、100μmol.m-2.s-1低温弱光处理使辣椒叶片PSⅡ的实际光能转化效率ΦPSⅡ、开放的PSⅡ有效光能转化效率Fv′/Fm′、光化学猝灭系数qP、光下最大荧光Fm′、光下最小荧光Fo′和光下最大可变荧光Fv′均降低,非光化学猝灭系数NPQ和稳态荧光Fs增大,最大光能转化效率Fv/Fm变化不显著。15℃/5℃、100μmol.m-2.s-1低温弱光处理15 d辣椒叶片的ΦPSⅡ和Fv′与辣椒的冷害指数呈极显著负相关,Fv′/Fm′、qP和Fm′呈显著负相关,NPQ呈显著正相关。【结论】15℃/5℃、100μmol.m-2.s-1低温弱光处理15 d辣椒叶片的ΦPSⅡ、Fv′/Fm′、qP、NPQ、Fm′和Fv′等叶绿素荧光参数可用于辣椒耐低温弱光性快速、准确、无损伤的鉴定指标,其中ΦPSⅡ和Fv′为最敏感的指标。
XuJZ, YuYM, PenSZ, YangSH, LiaoLX ( 2014). A modified nonrectangular hyperbola equation for photosynthetic light-response curves of leaves with different nitrogen status Photosynthetica, 52, 117-123. DOIURL [本文引用: 1] Chlorophyll index and leaf nitrogen status (SPAD value) was incorporated into the nonrectangular hyperbola (NRH) equation for photosynthetic light-response (PLR) curve to establish a modified NRH equation to overcome the parameter variation. Ten PLR curves measured on rice leaves with different SPAD values were collected from pot experiments with different nitrogen (N) dosages. The coefficients of initial slope of the PLR curve and the maximum net photosynthetic rate in NRH equation increased linearly with the increase of leaf SPAD. The modified NRH equation was established by multiplying a linear SPAD-based adjustment factor with the NRH equation. It was sufficient in describing the PLR curves with unified coefficients for rice leaf with different SPAD values. SPAD value, as the indicator of leaf N status, could be used for modification of NRH equation to overcome the shortcoming of large coefficient variations between individual leaves with different N status. The performance of the SPAD-modified NRH equation should be further validated by data collected from different kinds of plants growing under different environments.
XuL, GaoZQ, AnW, LiYL, JiaoXF, WangCY ( 2016). Flag leaf photosynthetic characteristics, change in chlorophyll fluorescence parameters, and their relationship with yield of winter wheat sowed in spring Chinese Journal of Applied Ecology, 27, 133-142. DOIURL [本文引用: 1] 以长江中下游、西南麦区的5个优良冬播小麦品种为材料,在忻定盆地春播(早播、适播)条件下,于 2013-2014年对开花期、灌浆期的旗叶光合指标、叶绿素含量及叶绿素荧光参数进行测定,并分析这些参数与产量的相关性.结果表明:品种间大多荧光参 数(除叶绿素外)差异显著,且相关性显著;叶绿素变异系数较小(0.12~0.17),吸收光能为基础的性能指数(PIabs)变异系数较大 (0.32~0.39),两参数与产量偏相关系数为0.70~0.81;早播条件下,籽粒产量与PIabs(灌浆期、开花期)、灌浆期叶绿素呈显著正效 应,与灌浆期旗叶在Ⅰ点的相对可变荧光强度(Vi)呈显著负效应,且产量的81.1%~82.8%(2013、2014年)可由这3因素的变异决定;不同 品种表现出不同的播期效应,且两年变化趋势基本一致:扬麦13(春性、中早熟)旗叶光合速率、叶绿素及绝大多数荧光参数和产量均显著高于其他品种,且适宜 早播.早播条件下灌浆期旗叶叶绿素含量、PIabs、光合速率可作为选择高光效小麦资源的重要评价指标. [ 徐澜, 高志强, 安伟, 李彦良, 焦雄飞, 王创云 ( 2016). 冬麦春播条件下旗叶光合特性、叶绿素荧光参数变化及其与产量的关系 应用生态学报, 27, 133-142.] DOIURL [本文引用: 1] 以长江中下游、西南麦区的5个优良冬播小麦品种为材料,在忻定盆地春播(早播、适播)条件下,于 2013-2014年对开花期、灌浆期的旗叶光合指标、叶绿素含量及叶绿素荧光参数进行测定,并分析这些参数与产量的相关性.结果表明:品种间大多荧光参 数(除叶绿素外)差异显著,且相关性显著;叶绿素变异系数较小(0.12~0.17),吸收光能为基础的性能指数(PIabs)变异系数较大 (0.32~0.39),两参数与产量偏相关系数为0.70~0.81;早播条件下,籽粒产量与PIabs(灌浆期、开花期)、灌浆期叶绿素呈显著正效 应,与灌浆期旗叶在Ⅰ点的相对可变荧光强度(Vi)呈显著负效应,且产量的81.1%~82.8%(2013、2014年)可由这3因素的变异决定;不同 品种表现出不同的播期效应,且两年变化趋势基本一致:扬麦13(春性、中早熟)旗叶光合速率、叶绿素及绝大多数荧光参数和产量均显著高于其他品种,且适宜 早播.早播条件下灌浆期旗叶叶绿素含量、PIabs、光合速率可作为选择高光效小麦资源的重要评价指标.
YanLX, YuZW, ShiY, ZhaoJY, ZhangYL ( 2017). Effects of supplemental irrigation based on soil moisture measurement on flag leaf chlorophyll fluorescence and senescence characteristics in two wheat cultivars Scientia Agricultura Sinica, 50, 1416-1429. DOIURL [本文引用: 1] 【目的】探讨测墒补灌和定量灌溉对2个小麦品种旗叶叶绿素荧光、衰老特性及籽粒产量的影响,为小麦节水高产提供理论依据。【方法】于2013—2015两年度,在大田条件下,选用泰农18(T18)和济麦22(J22)2个小麦品种,设置3个水分处理:W0(全生育期不灌水)、W1(依据0—40 cm土层土壤相对含水量进行测墒补灌,拔节期和开花期目标土壤相对含水量均为65%)、W2(定量灌溉,拔节期和开花期分别灌溉60 mm),研究测墒补灌和定量灌溉对2个小麦品种旗叶叶绿素荧光特性及衰老特性的影响。【结果】W1处理通过调节拔节期和开花期灌水量,保持灌水后0—40 cm土层土壤相对含水量在65%,可防止灌水过多或过少,为小麦生长发育创造适宜的土壤水分环境。W1处理条件下,两小麦品种开花后14、21和28 d的旗叶电子传递速率、光化学猝灭系数、PSⅡ实际光化学效率及旗叶蔗糖含量均显著高于W2处理,磷酸蔗糖合成酶活性在花后14和21 d显著高于W2处理;两小麦品种开花后14、21和28 d的超氧化物歧化酶、过氧化氢酶活性均显著高于W2,但同期旗叶丙二醛含量显著低于W2并保持较高的旗叶可溶性蛋白含量。两年度T18和J22两品种W1处理的籽粒产量、水分利用效率和灌溉效益均显著高于W2。品种间比较可知,T18两灌水处理的旗叶电子传递速率、光化学猝灭系数、PSⅡ实际光化学效率及旗叶蔗糖含量在花后21和28 d均显著高于J22,磷酸蔗糖合成酶活性在花后7、14和21 d亦显著高于J22;T18开花后21和28 d的超氧化物歧化酶、过氧化氢酶活性、可溶性蛋白含量均显著高于J22,但同期旗叶丙二醛含量显著低于J22。同一年度同一处理条件下,T18和J20总耗水量和水分利用效率均无显著差异;在W0处理条件下,J22的籽粒产量显著高于T18;但在W1和W2处理条件下,T18的籽粒产量、灌溉效益均显著高于J22。【结论】在17 [ 闫丽霞, 于振文, 石玉, 赵俊晔, 张永丽 ( 2017). 测墒补灌对2个小麦品种旗叶叶绿素荧光及衰老特性的影响 中国农业科学, 50, 1416-1429.] DOIURL [本文引用: 1] 【目的】探讨测墒补灌和定量灌溉对2个小麦品种旗叶叶绿素荧光、衰老特性及籽粒产量的影响,为小麦节水高产提供理论依据。【方法】于2013—2015两年度,在大田条件下,选用泰农18(T18)和济麦22(J22)2个小麦品种,设置3个水分处理:W0(全生育期不灌水)、W1(依据0—40 cm土层土壤相对含水量进行测墒补灌,拔节期和开花期目标土壤相对含水量均为65%)、W2(定量灌溉,拔节期和开花期分别灌溉60 mm),研究测墒补灌和定量灌溉对2个小麦品种旗叶叶绿素荧光特性及衰老特性的影响。【结果】W1处理通过调节拔节期和开花期灌水量,保持灌水后0—40 cm土层土壤相对含水量在65%,可防止灌水过多或过少,为小麦生长发育创造适宜的土壤水分环境。W1处理条件下,两小麦品种开花后14、21和28 d的旗叶电子传递速率、光化学猝灭系数、PSⅡ实际光化学效率及旗叶蔗糖含量均显著高于W2处理,磷酸蔗糖合成酶活性在花后14和21 d显著高于W2处理;两小麦品种开花后14、21和28 d的超氧化物歧化酶、过氧化氢酶活性均显著高于W2,但同期旗叶丙二醛含量显著低于W2并保持较高的旗叶可溶性蛋白含量。两年度T18和J22两品种W1处理的籽粒产量、水分利用效率和灌溉效益均显著高于W2。品种间比较可知,T18两灌水处理的旗叶电子传递速率、光化学猝灭系数、PSⅡ实际光化学效率及旗叶蔗糖含量在花后21和28 d均显著高于J22,磷酸蔗糖合成酶活性在花后7、14和21 d亦显著高于J22;T18开花后21和28 d的超氧化物歧化酶、过氧化氢酶活性、可溶性蛋白含量均显著高于J22,但同期旗叶丙二醛含量显著低于J22。同一年度同一处理条件下,T18和J20总耗水量和水分利用效率均无显著差异;在W0处理条件下,J22的籽粒产量显著高于T18;但在W1和W2处理条件下,T18的籽粒产量、灌溉效益均显著高于J22。【结论】在17
YanXH, YinJH, DuanSH, ZouhB, HuWH, LiuS ( 2013). Photosynthesis light response curves of four rice varieties and model fitting Chinese Journal of Ecology, 32, 604-610. URL [本文引用: 1] 应用4种典型的光响应模型对4个水稻品种“02428”、“淦鑫688”、“JR8892.1”和“JR8892.2”叶片的光合光响应曲线进行了拟合。结果表明:直角双曲线模型、非直角双曲线模型、指数函数以及修正的直角双曲线模型对4个水稻品种的光合光响应曲线都可以进行拟合,决定系数均为R^2〉0.99。但只有修正的直角双曲线模型可以拟合发生光抑制部分的光响应曲线,且可直接计算其饱和光强(Isat);由各模型计算的光合参数可知,直角双曲线模型、非直角双曲线模型求得的最大净光合速率(Pnmax)远高于实测值,指数函数与修正的直角双曲线模型求得的Pnmax,与实测值最为接近;直角双曲线、非直角双曲线模型以及指数函数求得的Lsat均与实测值相差较大,除了“JR8892—1”采用指数函数求得的,sat大于实测值外,其余3个品种用上述3种模型拟合求得的Isat均小于实测值,唯有修正的直角双曲线模型求得的,sat与实测值最为相符;对于低光强部分的参数光补偿点(Ic)与暗呼吸速率(Rd),修正的直角双曲线模型最接近其测量值;不同水稻品种的光合特性,“JR8892—1”饱和光强与最大净光合速率明显高于高光效品种“02428”;处于分蘖盛期的“JR8892—2”也表现出较高的饱和光强和最大净光合速率,而超级稻“淦鑫688”相对于其他品种,则具有较低的暗呼吸速率,光合产物消耗少。 [ 闫小红, 尹建华, 段世华, 周兵, 胡文海, 刘帅 ( 2013). 四种水稻品种的光合光响应曲线及其模型拟合 生态学杂志, 32, 604-610.] URL [本文引用: 1] 应用4种典型的光响应模型对4个水稻品种“02428”、“淦鑫688”、“JR8892.1”和“JR8892.2”叶片的光合光响应曲线进行了拟合。结果表明:直角双曲线模型、非直角双曲线模型、指数函数以及修正的直角双曲线模型对4个水稻品种的光合光响应曲线都可以进行拟合,决定系数均为R^2〉0.99。但只有修正的直角双曲线模型可以拟合发生光抑制部分的光响应曲线,且可直接计算其饱和光强(Isat);由各模型计算的光合参数可知,直角双曲线模型、非直角双曲线模型求得的最大净光合速率(Pnmax)远高于实测值,指数函数与修正的直角双曲线模型求得的Pnmax,与实测值最为接近;直角双曲线、非直角双曲线模型以及指数函数求得的Lsat均与实测值相差较大,除了“JR8892—1”采用指数函数求得的,sat大于实测值外,其余3个品种用上述3种模型拟合求得的Isat均小于实测值,唯有修正的直角双曲线模型求得的,sat与实测值最为相符;对于低光强部分的参数光补偿点(Ic)与暗呼吸速率(Rd),修正的直角双曲线模型最接近其测量值;不同水稻品种的光合特性,“JR8892—1”饱和光强与最大净光合速率明显高于高光效品种“02428”;处于分蘖盛期的“JR8892—2”也表现出较高的饱和光强和最大净光合速率,而超级稻“淦鑫688”相对于其他品种,则具有较低的暗呼吸速率,光合产物消耗少。
YanXH, ZhouB, WangN, YeZP, YinZF ( 2016). Differences of photosynthetic characteristics based on a mechanistic model of light-response for three herbaceous plants Journal of Nanjing Forestry University (Natural Sciences Edition), 40, 63-69. DOIURL [本文引用: 1] 利用植物光响应机理模型,比较了大狼把草(Bidens frondosa)、山莴苣(Lactuca indica)和酸模叶蓼(Polygonum lapathifolium)光合特性的差异。结果表明,大狼把草和酸模叶蓼在中、高光强部分的电子传递速率(J)明显高于山莴苣,并且在高光强部分二者没有出现明显的光抑制,而山莴苣则光抑制较明显。大狼把草和酸模叶蓼在各光强下的净光合速率(P_n)也明显高于山莴苣。3种植物成熟叶片叶绿素的含量依次是大狼把草〉酸模叶蓼〉山莴苣,类胡萝卜素含量依次为酸模叶蓼〉大狼把草〉山莴苣。山莴苣的叶绿素含量虽然最少,但它的捕光色素分子本征光能吸收截面(σ_(ik))最大;与之相反,大狼把草的叶绿素含量最多,但σ_(ik)最小,酸模叶蓼的叶绿素含量和σ_(ik)则介于二者之间。大狼把草和酸模叶蓼的最大电子传递速率(Jmax)明显高于山莴苣,二者是通过增加有效光能吸收截面,以及缩短τ_(min)值提高电子传递速率。相对于山莴苣,大狼把草和酸模叶蓼具有更高的最大净光合速率(P_(n,max))、光饱和点(I_(sat))和暗呼吸速率(R_d)。大狼把草更高的非光化学猝灭系数(q_(NPQ))有利于热耗散,避免光损伤。酸模叶蓼更高的类胡萝卜素含量,在耗散过剩的激发能上具有积极作用,有助于植物体免受高温、高光、干旱等逆境伤害。综上所述,相较于山莴苣,大狼把草和酸模叶蓼应该对环境胁迫有更好的耐受性和适应性。 [ 闫小红, 周兵, 王宁, 叶子飘, 尹增芳 ( 2016). 基于光响应机理模型的3种草本植物光合特性差异解析 南京林业大学学报(自然科学版), 40, 63-69.] DOIURL [本文引用: 1] 利用植物光响应机理模型,比较了大狼把草(Bidens frondosa)、山莴苣(Lactuca indica)和酸模叶蓼(Polygonum lapathifolium)光合特性的差异。结果表明,大狼把草和酸模叶蓼在中、高光强部分的电子传递速率(J)明显高于山莴苣,并且在高光强部分二者没有出现明显的光抑制,而山莴苣则光抑制较明显。大狼把草和酸模叶蓼在各光强下的净光合速率(P_n)也明显高于山莴苣。3种植物成熟叶片叶绿素的含量依次是大狼把草〉酸模叶蓼〉山莴苣,类胡萝卜素含量依次为酸模叶蓼〉大狼把草〉山莴苣。山莴苣的叶绿素含量虽然最少,但它的捕光色素分子本征光能吸收截面(σ_(ik))最大;与之相反,大狼把草的叶绿素含量最多,但σ_(ik)最小,酸模叶蓼的叶绿素含量和σ_(ik)则介于二者之间。大狼把草和酸模叶蓼的最大电子传递速率(Jmax)明显高于山莴苣,二者是通过增加有效光能吸收截面,以及缩短τ_(min)值提高电子传递速率。相对于山莴苣,大狼把草和酸模叶蓼具有更高的最大净光合速率(P_(n,max))、光饱和点(I_(sat))和暗呼吸速率(R_d)。大狼把草更高的非光化学猝灭系数(q_(NPQ))有利于热耗散,避免光损伤。酸模叶蓼更高的类胡萝卜素含量,在耗散过剩的激发能上具有积极作用,有助于植物体免受高温、高光、干旱等逆境伤害。综上所述,相较于山莴苣,大狼把草和酸模叶蓼应该对环境胁迫有更好的耐受性和适应性。
YaoCY, LuXS, XuDW, WuJ ( 2011). Study on the relations of six forage’s photo-response characteristics and aboveground biomass under orchard Chinese Agricultural Science Bulletin, 27, 69-73. URL [本文引用: 1] 在果林下间作菊苣、紫花苜蓿(新疆大叶)、紫花苜蓿(驯鹿)、红三叶、鸭茅、无芒雀麦,对6种牧草的产量和光响应特性进行测定分析,研究不同牧草在果林的生长状况。结果表明:6种牧草产量的高低依次为菊苣驯鹿苜蓿新疆大叶鸭茅红三叶无芒雀麦。利用叶子飘新模型,对6种牧草的光响应曲线拟合表明菊苣、紫花苜蓿(新疆大叶)、紫花苜蓿(驯鹿)的光合能力较强,最大净光合速率均大于15μmolCO2/(m2·s)。6种牧草的光补偿点的大小为鸭茅菊苣新疆大叶驯鹿苜蓿无芒雀麦红三叶,光饱和点为驯鹿苜蓿新疆大叶无芒雀麦鸭茅红三叶菊苣;6种牧草在光量子小于400μmol/(m2·s)时Pn均值与第1茬鲜草产量存在显著相关关系。 [ 姚春艳, 卢欣石, 徐大伟, 吴健 ( 2011). 果林下6种牧草的光响应特性与地上生物量关系的研究 中国农学通报, 27, 69-73.] URL [本文引用: 1] 在果林下间作菊苣、紫花苜蓿(新疆大叶)、紫花苜蓿(驯鹿)、红三叶、鸭茅、无芒雀麦,对6种牧草的产量和光响应特性进行测定分析,研究不同牧草在果林的生长状况。结果表明:6种牧草产量的高低依次为菊苣驯鹿苜蓿新疆大叶鸭茅红三叶无芒雀麦。利用叶子飘新模型,对6种牧草的光响应曲线拟合表明菊苣、紫花苜蓿(新疆大叶)、紫花苜蓿(驯鹿)的光合能力较强,最大净光合速率均大于15μmolCO2/(m2·s)。6种牧草的光补偿点的大小为鸭茅菊苣新疆大叶驯鹿苜蓿无芒雀麦红三叶,光饱和点为驯鹿苜蓿新疆大叶无芒雀麦鸭茅红三叶菊苣;6种牧草在光量子小于400μmol/(m2·s)时Pn均值与第1茬鲜草产量存在显著相关关系。
YaoJP, ChenL, CuiHX, YaoT ( 2010). Responses of photosynthesis and heat-tolerance in seedling of four lilacs to high-temperature Grassland & Turf,( 5), 50- 55, 58. DOIURL [本文引用: 1] 为研究植物迁地保育的引种适应机制,以丁香属广布种和狭域种4种代表种质为试验材料,研究了丁香属种间叶片光合气体交换、叶绿素荧光的热响应特征。结果显示,引种地旱季37℃高温使4种丁香净光合速率(Pn)呈不同变化趋势,气孔导度(Gs)和胞间CO2浓度(Ci)均呈上升趋势;与冷凉处理相比,适度高温下4种丁香叶片羧化效率(CE)不同程度下降,而蒸腾速率(Tr)与之相反;高温下,4种丁香光系统II(PSII)的激发能捕获效率(Fv′/Fm′)、光化学淬灭(qP)、实际光化学效率(ΦPSII)和电子传递速率(ETR)与净光合速率变化相一致。与紫丁香和暴马丁香相比,红丁香和羽叶丁香的非光化学淬灭(NPQ)均显著升高。综合分析表明:(1)在适度高温下,非气孔因素是丁香狭域种光合速率下降的主要原因;(2)较高的蒸腾速率和较强的光能利用效率可能是丁香广布种在适度高温下保持光合机构热稳定性的重要原因;(3)紫丁香、暴马丁香较耐热,羽叶丁香的耐热性最差。 [ 姚军朋, 陈莉, 崔洪霞, 姚拓 ( 2010). 丁香属植物光合机构的热响应特征研究 草原与草坪,( 5), 50- 55, 58.] DOIURL [本文引用: 1] 为研究植物迁地保育的引种适应机制,以丁香属广布种和狭域种4种代表种质为试验材料,研究了丁香属种间叶片光合气体交换、叶绿素荧光的热响应特征。结果显示,引种地旱季37℃高温使4种丁香净光合速率(Pn)呈不同变化趋势,气孔导度(Gs)和胞间CO2浓度(Ci)均呈上升趋势;与冷凉处理相比,适度高温下4种丁香叶片羧化效率(CE)不同程度下降,而蒸腾速率(Tr)与之相反;高温下,4种丁香光系统II(PSII)的激发能捕获效率(Fv′/Fm′)、光化学淬灭(qP)、实际光化学效率(ΦPSII)和电子传递速率(ETR)与净光合速率变化相一致。与紫丁香和暴马丁香相比,红丁香和羽叶丁香的非光化学淬灭(NPQ)均显著升高。综合分析表明:(1)在适度高温下,非气孔因素是丁香狭域种光合速率下降的主要原因;(2)较高的蒸腾速率和较强的光能利用效率可能是丁香广布种在适度高温下保持光合机构热稳定性的重要原因;(3)紫丁香、暴马丁香较耐热,羽叶丁香的耐热性最差。
YeZP, HuWH, XiaoYA, FanDY, YinJH, DuanSH, YanXH, HeL, ZhangSS ( 2014). A mechanistic model of light-response of photosynthetic electron flow and its application Chinese Journal of Plant Ecology, 38, 1241-1249. DOIURL [本文引用: 2] AimsMethodsImportant findings (1) The mechanistic model of light-response of photosynthetic electron flow not only well described the light-response curves of photosynthetic electron flow in L. sibiricum, E. annuus and A. tataricus, but also obtained some key photosynthetic parameters, e.g. maximum photosynthetic electron flow, saturation irradiance and initial slope of the light-response curve; the fitted photosynthetic parameters were similar to the measured values. (2) The effective light absorption cross-section of light-harvesting pigment molecules quickly decreased with increasing irradiance in L. sibiricum, and showed slowest rate of decrease in E. annuus. (3) The light-harvesting pigment molecules in the lowest excited state increased most rapidly with increasing irradiance in L. sibiricum, and most slowly in E. annuus. In conclusion, compelling evidence indicates that decrease in effective absorption cross-section and increase in the number of light-harvesting pigments in the lowest excited state would reduce light energy absorption. [ 叶子飘, 胡文海, 肖宜安, 樊大勇, 尹建华, 段世华, 闫小红, 贺俐, 张斯斯 ( 2014). 光合电子流对光响应的机理模型及其应用 植物生态学报, 38, 1241-1249.] DOIURL [本文引用: 2] AimsMethodsImportant findings (1) The mechanistic model of light-response of photosynthetic electron flow not only well described the light-response curves of photosynthetic electron flow in L. sibiricum, E. annuus and A. tataricus, but also obtained some key photosynthetic parameters, e.g. maximum photosynthetic electron flow, saturation irradiance and initial slope of the light-response curve; the fitted photosynthetic parameters were similar to the measured values. (2) The effective light absorption cross-section of light-harvesting pigment molecules quickly decreased with increasing irradiance in L. sibiricum, and showed slowest rate of decrease in E. annuus. (3) The light-harvesting pigment molecules in the lowest excited state increased most rapidly with increasing irradiance in L. sibiricum, and most slowly in E. annuus. In conclusion, compelling evidence indicates that decrease in effective absorption cross-section and increase in the number of light-harvesting pigments in the lowest excited state would reduce light energy absorption.
YeZP, HuWH, YanXH ( 2016 a). Comparison on light-response models of actual photochemical efficiency in photosystem II Chinese Journal of Plant Ecology, 40, 1208-1217. DOIURL [本文引用: 1] 为了比较光系统Ⅱ实际光化学量子效率(FPSⅡ)对光的响应机理模型(简称机理模型)、负指数模型和指数模型的优缺点,用LI-6400-40B光合作用测定仪控制CO2浓度和温度,测量了剑叶金鸡菊(Coreopsis lanceolata)、黄荆(Vitex negundo)和大狼杷草(Bidens frondosa)的电子传递速率(ETR)对光的响应曲线(ETR-I)和FPSⅡ对光的响应曲线(FPSⅡ-I),然后用这3个模型分别拟合了这些数据。拟合结果表明:3个模型都可以较好地拟合这3种植物的ETR-I的响应数据和FPSⅡ-I的响应数据,但由指数模型拟合ETR-I和FPSⅡ-I的响应数据得到相应的饱和光强(PARsat)和光系统Ⅱ最大光能利用效率(Fv/Fm)之间存在显著差异,且估算的饱和光强远低于实测值。由机理模型可知,FPSⅡ不仅与光强的函数有关,还与植物的内禀特性有关,即与天线色素分子的本征光能吸收截面、激子的传递效率、能级的简并度、光化学反应常数、热耗散常数和处于最低激发态的平均寿命等参数有关。此外,由机理模型还可知,FPSⅡ随光强的增加而下降的原因是捕光色素分子的有效光能吸收截面随光强增加而降低。 [ 叶子飘, 胡文海, 闫小红 ( 2016 a). 光系统II实际光化学量子效率对光的响应模型的比较 植物生态学报, 40, 1208-1217.] DOIURL [本文引用: 1] 为了比较光系统Ⅱ实际光化学量子效率(FPSⅡ)对光的响应机理模型(简称机理模型)、负指数模型和指数模型的优缺点,用LI-6400-40B光合作用测定仪控制CO2浓度和温度,测量了剑叶金鸡菊(Coreopsis lanceolata)、黄荆(Vitex negundo)和大狼杷草(Bidens frondosa)的电子传递速率(ETR)对光的响应曲线(ETR-I)和FPSⅡ对光的响应曲线(FPSⅡ-I),然后用这3个模型分别拟合了这些数据。拟合结果表明:3个模型都可以较好地拟合这3种植物的ETR-I的响应数据和FPSⅡ-I的响应数据,但由指数模型拟合ETR-I和FPSⅡ-I的响应数据得到相应的饱和光强(PARsat)和光系统Ⅱ最大光能利用效率(Fv/Fm)之间存在显著差异,且估算的饱和光强远低于实测值。由机理模型可知,FPSⅡ不仅与光强的函数有关,还与植物的内禀特性有关,即与天线色素分子的本征光能吸收截面、激子的传递效率、能级的简并度、光化学反应常数、热耗散常数和处于最低激发态的平均寿命等参数有关。此外,由机理模型还可知,FPSⅡ随光强的增加而下降的原因是捕光色素分子的有效光能吸收截面随光强增加而降低。
YeZP, HuWH, YanXH, DuanSH ( 2016 b). Photosynthetic characteristics of different plant species based on a mechanistic model of light-response of photosynthesis Chinese Journal of Ecology, 35, 2544-2552. DOIURL [本文引用: 6] 为了研究不同植物的光合特性,用LI-6400-40B荧光仪同时测量了葎草(Humulus scandens)、白泡桐(Paulownia fortunei)、龙葵(Solanum nigrum)的电子传递速率和光合作用对光的响应曲线。利用植物光合电子流对光响应和光合作用对光响应的机理模型研究了这3种植物光合特性的差异及其产生的原因。结果表明:葎草的叶绿素含量最低,但其捕光色素分子的本征光能吸收截面最大;白泡桐的叶绿素含量是龙葵的1.2倍,但这两种植物的最大电子传递速率没有差异。产生这种差异的原因是:龙葵的捕光色素分子的本征光能吸收截面要比白泡桐大,而它处于激发态的最小平均寿命要比白泡桐短,且龙葵的电子利用效率要比白泡桐大。此外,捕光色素分子的有效光能吸收截面和处于激发态的捕光色素分子对光响应曲线的差异也有可能影响它们的光合特性。 [ 叶子飘, 胡文海, 闫小红, 段世华 ( 2016 b). 基于光响应机理模型的不同植物光合特性 生态学杂志, 35, 2544-2552.] DOIURL [本文引用: 6] 为了研究不同植物的光合特性,用LI-6400-40B荧光仪同时测量了葎草(Humulus scandens)、白泡桐(Paulownia fortunei)、龙葵(Solanum nigrum)的电子传递速率和光合作用对光的响应曲线。利用植物光合电子流对光响应和光合作用对光响应的机理模型研究了这3种植物光合特性的差异及其产生的原因。结果表明:葎草的叶绿素含量最低,但其捕光色素分子的本征光能吸收截面最大;白泡桐的叶绿素含量是龙葵的1.2倍,但这两种植物的最大电子传递速率没有差异。产生这种差异的原因是:龙葵的捕光色素分子的本征光能吸收截面要比白泡桐大,而它处于激发态的最小平均寿命要比白泡桐短,且龙葵的电子利用效率要比白泡桐大。此外,捕光色素分子的有效光能吸收截面和处于激发态的捕光色素分子对光响应曲线的差异也有可能影响它们的光合特性。
YeZP, KangHJ, TaoYL, WangLX ( 2011). Comparative analysis on the fitting effect of different models of Koelreuteria bipinnata var. integrifoliola rapid light curves Chinese Journal of Ecology, 30, 1662-1667. URL [本文引用: 2] 用Li-6400光合仪同时测定了CO2浓度为380和600μmol·mol-1条件下黄山栾树的光响应曲线和快速光曲线,分别采用不同模型进行了拟合。结果表明:直角双曲线模型和双指数方程拟合得到的最大电子传递速率远大于实测值;直角双曲线模型、非直角双曲线模型、单指数方程不能拟合黄山栾树存在PSⅡ动力学下调的快速光曲线,只有双指数方程和直角双曲线修正模型可以拟合黄山栾树存在PSⅡ动力学下调的快速光曲线,且可以计算它的饱和光强。综合拟合结果可知,直角双曲线修正模型不仅可以很好地拟合黄山栾树的快速光曲线,而且得到的最大电子传递速率和饱和光强与实测值相符合。此外,通过拟合植物的快速光曲和光响应曲线,还可以判断在饱和光强时它的电子传递速率与碳同化是否同时达到最大值。 [ 叶子飘, 康华靖, 陶月良, 王立新 ( 2011). 不同模型对黄山栾树快速光曲线拟合效果的比较 生态学杂志, 30, 1662-1667.] URL [本文引用: 2] 用Li-6400光合仪同时测定了CO2浓度为380和600μmol·mol-1条件下黄山栾树的光响应曲线和快速光曲线,分别采用不同模型进行了拟合。结果表明:直角双曲线模型和双指数方程拟合得到的最大电子传递速率远大于实测值;直角双曲线模型、非直角双曲线模型、单指数方程不能拟合黄山栾树存在PSⅡ动力学下调的快速光曲线,只有双指数方程和直角双曲线修正模型可以拟合黄山栾树存在PSⅡ动力学下调的快速光曲线,且可以计算它的饱和光强。综合拟合结果可知,直角双曲线修正模型不仅可以很好地拟合黄山栾树的快速光曲线,而且得到的最大电子传递速率和饱和光强与实测值相符合。此外,通过拟合植物的快速光曲和光响应曲线,还可以判断在饱和光强时它的电子传递速率与碳同化是否同时达到最大值。
YeZP, RobakowskiP, SuggettDJ ( 2013 a). A mechanistic model for the light response of photosynthetic electron transport rate based on light harvesting properties of photosynthetic pigment molecules Planta, 237, 837-847. DOIURLPMID [本文引用: 3] Models describing the light response of photosynthetic electron transport rate (ETR) are routinely used to determine how light absorption influences energy, reducing power and yields of primary productivity; however, no single model is currently able to provide insight into the fundamental processes that implicitly govern the variability of light absorption. Here we present development and application of a new mechanistic model of ETR for photosystem II based on the light harvesting (absorption and transfer to the core 'reaction centres') characteristics of photosynthetic pigment molecules. Within this model a series of equations are used to describe novel biophysical and biochemical characteristics of photosynthetic pigment molecules and in turn light harvesting; specifically, the eigen-absorption cross-section and the minimum average lifetime of photosynthetic pigment molecules in the excited state, which describe the ability of light absorption of photosynthetic pigment molecules and retention time of excitons in the excited state but are difficult to be measured directly. We applied this model to a series of previously collected fluorescence data and demonstrated that our model described well the light response curves of ETR, regardless of whether dynamic down-regulation of PSII occurs, for a range of photosynthetic organisms (Abies alba, Picea abies, Pinus mugo and Emiliania huxleyi). Inherent estimated parameters (e.g. maximum ETR and the saturation irradiance) by our model are in very close agreement with the measured data. Overall, our mechanistic model potentially provides novel insights into the regulation of ETR by light harvesting properties as well as dynamical down-regulation of PSII.
YeZP, SuggettJD, RobakowskiP, KangHJ ( 2013 b). A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species New Phytologist, 199, 110-120. DOIURLPMID [本文引用: 3] A new mechanistic model of the photosynthesis090009light response is developed based on photosynthetic electron transport via photosystem II (PSII) to specifically describe light-harvesting characteristics and associated biophysical parameters of photosynthetic pigment molecules. This model parameterizes 090004core090005 characteristics not only of the light response but also of difficult to measure physical parameters of photosynthetic pigment molecules in plants.Application of the model to two C3 and two C4 species grown under the same conditions demonstrated that the model reproduced extremely well (r2 > 0.992) the light response trends of both electron transport and CO2 uptake.In all cases, the effective absorption cross-section of photosynthetic pigment molecules decreased with increasing light intensity, demonstrating novel operation of a key mechanism for plants to avoid high light damage.In parameterizing these previously difficult to measure characteristics of light harvesting in higher plants, the model provides a new means to understand the mechanistic processes underpinning variability of CO2 uptake, for example, photosynthetic down-regulation or reversible photoinhibition induced by high light and photoprotection. However, an important next step is validating this parameterization, possibly through application to less structurally complex organisms such as single-celled algae.
YeZP, YanXH, DuanSH ( 2015). Investigation on the relationship between saturation irradiance and chlorophyll contents of flag leaves and intrinsic characteristics of light-harvesting pigment molecules in high-yielding rice Journal of Jinggangshan University (Natural Science), 36, 25-32. DOIURL [本文引用: 1] 叶绿素是水稻叶片进行光合作用 的重要物质之一。为了研究叶片叶绿素的含量、捕光色素分子的内禀特性与饱和光强的关系,利用LI-6400XT便携式光合测定系统测量了6个水稻品种的光 响应曲线,并利用光合作用对光响应的机理模型进行了拟合。结果表明:供试的6种水稻品种的饱和光强只与植物捕光色素分子的物理参数如本征光能吸收截面、激 子传递到光反应中心的速率(kp)、热耗散速率(kD)和激发态的平均寿命(?)等有关,与叶片的叶绿素含量无关。 [ 叶子飘, 闫小红, 段世华 ( 2015). 高产水稻剑叶的叶绿素含量、捕光色素分子的内禀特性与饱和光强关系的研究 井冈山大学学报(自然科学版), 36, 25-32.] DOIURL [本文引用: 1] 叶绿素是水稻叶片进行光合作用 的重要物质之一。为了研究叶片叶绿素的含量、捕光色素分子的内禀特性与饱和光强的关系,利用LI-6400XT便携式光合测定系统测量了6个水稻品种的光 响应曲线,并利用光合作用对光响应的机理模型进行了拟合。结果表明:供试的6种水稻品种的饱和光强只与植物捕光色素分子的物理参数如本征光能吸收截面、激 子传递到光反应中心的速率(kp)、热耗散速率(kD)和激发态的平均寿命(?)等有关,与叶片的叶绿素含量无关。
YeZP, ZhaoZH ( 2009). Effects of shading on the photosynthesis and chlorophyll content of Bidens pilosa Chinese Journal of Ecology, 28, 19-22. URL [本文引用: 1] 采用盆栽试验,研究了遮光对三叶鬼针草(Bidens pilosa)光合参数的影响。结果表明:遮光处理提高了三叶鬼针草叶绿素a+b含量,但叶绿素a/b的比值没有什么改变;高光强条件下生长的三叶鬼针草的最大光合速率和饱和光分别是遮光处理的3.79和2.71倍;遮光处理的初始斜率比对照高,而表观量子效率却比对照低;遮光处理的三叶鬼针草在光合有效辐射低于100μmol.m-2.s-1时,其光合速率比对照组要高;当光合有效辐射超过650μmol.m-2.s-1时,光合速率随光强的增加而下降;对照组在光强约为1780μmol.m-2.s-1时才出现光合速率随光强的增加而降低的现象;遮光处理不利于三叶鬼针草的生长。 [ 叶子飘, 赵则海 ( 2009). 遮光对三叶鬼针草光合作用和叶绿素含量的影响 生态学杂志, 28, 19-22.] URL [本文引用: 1] 采用盆栽试验,研究了遮光对三叶鬼针草(Bidens pilosa)光合参数的影响。结果表明:遮光处理提高了三叶鬼针草叶绿素a+b含量,但叶绿素a/b的比值没有什么改变;高光强条件下生长的三叶鬼针草的最大光合速率和饱和光分别是遮光处理的3.79和2.71倍;遮光处理的初始斜率比对照高,而表观量子效率却比对照低;遮光处理的三叶鬼针草在光合有效辐射低于100μmol.m-2.s-1时,其光合速率比对照组要高;当光合有效辐射超过650μmol.m-2.s-1时,光合速率随光强的增加而下降;对照组在光强约为1780μmol.m-2.s-1时才出现光合速率随光强的增加而降低的现象;遮光处理不利于三叶鬼针草的生长。
YunJY, YangJD, ZhaoHL ( 2006). Research progress in the mechanism for drought and high temperature to affect plant photosynthesis Acta Botanica Boreali-Occidentalia Sinica, 26, 641-648. DOIURL [本文引用: 1] 干旱(水分亏缺)和高温是我国北方沙漠化地区植物生长季的两个主要环境胁迫因子.本文主要就干旱和高温在生理水平对植物光合作用影响机制的最新研究进展进行了综述,并对以后的相关研究进行了一些分析. [ 云建英, 杨甲定, 赵哈林 ( 2006). 干旱和高温对植物光合作用的影响机制研究进展 西北植物学报, 26, 641-648.] DOIURL [本文引用: 1] 干旱(水分亏缺)和高温是我国北方沙漠化地区植物生长季的两个主要环境胁迫因子.本文主要就干旱和高温在生理水平对植物光合作用影响机制的最新研究进展进行了综述,并对以后的相关研究进行了一些分析.
ZengZF, YanXH, HuWH ( 2014). Effects of NaCl stress on plant growth and photosynthetic characteristics in the cucumber seedling Journal of Jinggangshan University (Natural Science), 35, 39-42. DOIURL [本文引用: 1] 以碧春黄瓜为材料,研究了不同浓度 NaCl 处理对黄瓜幼苗生长、叶片光合作用特性、以及脯氨酸和丙二醛含量的影响。结果表明:10 mmol/L NaCl处理促进了黄瓜幼苗生长,但高浓度NaCl(≥25 mmol/L)处理下,黄瓜幼苗的生物量、净光合速率(Pn)、气孔导度( Gs)、胞间CO2浓度( Ci)、光系统II电子传递量子效率(ΦPSⅡ)和叶绿素含量均随着浓度增加下降,气孔限制值(LS)、MDA和脯氨酸含量则随着浓度增加而上升;但NaCl处理对光化学效率(Fv/Fm)影响不大。由此说明,低浓度NaCl处理促进了黄瓜幼苗的生长,且NaCl浓度低于100 mmol/L 时也未引起黄瓜幼苗叶片光抑制的发生,其光合作用下降的主要原因是叶绿素含量下降以及气孔限制,并且脯氨酸含量的增加也起到增强其抗性的作用。 [ 曾志峰, 严小红, 胡文海 ( 2014). NaCl胁迫对黄瓜幼苗生长及光合作用特征的影响 井冈山大学学报(自然科学版), 35, 39-42.] DOIURL [本文引用: 1] 以碧春黄瓜为材料,研究了不同浓度 NaCl 处理对黄瓜幼苗生长、叶片光合作用特性、以及脯氨酸和丙二醛含量的影响。结果表明:10 mmol/L NaCl处理促进了黄瓜幼苗生长,但高浓度NaCl(≥25 mmol/L)处理下,黄瓜幼苗的生物量、净光合速率(Pn)、气孔导度( Gs)、胞间CO2浓度( Ci)、光系统II电子传递量子效率(ΦPSⅡ)和叶绿素含量均随着浓度增加下降,气孔限制值(LS)、MDA和脯氨酸含量则随着浓度增加而上升;但NaCl处理对光化学效率(Fv/Fm)影响不大。由此说明,低浓度NaCl处理促进了黄瓜幼苗的生长,且NaCl浓度低于100 mmol/L 时也未引起黄瓜幼苗叶片光抑制的发生,其光合作用下降的主要原因是叶绿素含量下降以及气孔限制,并且脯氨酸含量的增加也起到增强其抗性的作用。
ZhangGS, HaoL, YanZJ, ZhaoX, WangY, BaiYR, LiXL ( 2017). The responses of chlorophyll fluorescence kinetics parameters of six tree species to soil moisture changes Chinese Journal of Ecology, 36, 3079-3085. DOIURL [本文引用: 1] 采用便携式调制叶绿素荧光仪(Mini-PAM),于2015年生长季节对6种固沙树种:沙柳(Salix psammophila C.Wang et Ch.Y.Yang)、黄柳(Salix gordejevii Y.L.Chang et Skv.)、沙枣(Elaeagnus angustifolia L.)、掌裂草葡萄(Ampelopsis aconitifolia var.glabra Diels)、紫穗槐(Amorpha fruticosa L.)和沙冬青(Ammopiptanthus mongolicus(Maxim.ex Kom.)S.H.Cheng)叶片叶绿素荧光动力学参数的日变化进行了测定,探讨其对土壤水分胁迫的响应。结果显示:量子产量(Yield)、相对光合电子传递速率(ETR)、最大量子产量(Fv/Fm)和非光化学淬灭系数(NPQ)的日变化呈单峰型,Yield、ETR和Fv/Fm值随着土壤体积含水量的降低而降低,NPQ随着土壤体积含水量的降低而增加,这意味着叶绿素荧光动力学参数可作为抗逆性评价的指标。Fv/Fm值变化量对土壤水分变化响应最为敏感,6种树种呈现沙冬青〉沙柳〉黄柳〉掌裂草葡萄〉紫穗槐〉沙枣的趋势,即旱生树种对土壤水分变化最为敏感,其次是中生或湿生树种。 [ 张国盛, 郝蕾, 闫子娟, 赵鑫, 王颖, 白玉荣, 李小龙 ( 2017). 6种树种叶片叶绿素荧光动力学参数对土壤水分变化的响应 生态学杂志, 36, 3079-3085.] DOIURL [本文引用: 1] 采用便携式调制叶绿素荧光仪(Mini-PAM),于2015年生长季节对6种固沙树种:沙柳(Salix psammophila C.Wang et Ch.Y.Yang)、黄柳(Salix gordejevii Y.L.Chang et Skv.)、沙枣(Elaeagnus angustifolia L.)、掌裂草葡萄(Ampelopsis aconitifolia var.glabra Diels)、紫穗槐(Amorpha fruticosa L.)和沙冬青(Ammopiptanthus mongolicus(Maxim.ex Kom.)S.H.Cheng)叶片叶绿素荧光动力学参数的日变化进行了测定,探讨其对土壤水分胁迫的响应。结果显示:量子产量(Yield)、相对光合电子传递速率(ETR)、最大量子产量(Fv/Fm)和非光化学淬灭系数(NPQ)的日变化呈单峰型,Yield、ETR和Fv/Fm值随着土壤体积含水量的降低而降低,NPQ随着土壤体积含水量的降低而增加,这意味着叶绿素荧光动力学参数可作为抗逆性评价的指标。Fv/Fm值变化量对土壤水分变化响应最为敏感,6种树种呈现沙冬青〉沙柳〉黄柳〉掌裂草葡萄〉紫穗槐〉沙枣的趋势,即旱生树种对土壤水分变化最为敏感,其次是中生或湿生树种。
ZhangLG, ZengFJ, LiuB, LiuZ, AnGX, LiHF, YuanN ( 2012). Study of the photosynthesis characteristics and physical signs of four plants at the desert-oasis ecoton Acta Prataculturae Sinica, 21, 103-111. DOIURL [本文引用: 1] 绿洲—荒漠过渡带植被的恢复和更新在过渡带生态环境建设中具有重 要意义.通过对多枝柽柳,梭梭,头状沙拐枣和沙枣的光合特性、水势(φ)以及生理指标的研究,结果表明,1)4种植物光响应曲线变化趋势基本一致,均表现 为净光合速率(A)随着光合有效辐射(PAR)的增大迅速增大,当PAR达到了植物的光饱和点之后,A出现下降的趋势;作为C4植物的头状沙拐枣和梭梭都 具有较高的光饱和点(LSP)和光补偿点(LCP),体现出了C4植物特有的高光合的优势,而其他一些光合参数K,Amax,Rday的变化规律并不完全 一致;2)4种植物中,梭梭具有较低的清晨和正午水势,说明梭梭主要依靠夜间水分恢复能力来应对干旱环境;清晨水势除头状沙拐枣和沙枣之间差异不显著外, 其他植物种之间的差异均达到了极显著水平(P<0.01),正午水势除多枝柽柳和沙枣之间差异不显著,其他植物种之间的差异也都达到了极显著水平 (P<0.01);3)4种植物中脯氨酸和可溶性糖的变化较为一致,头状沙拐枣均显著高于其他3种植物(P<0.05);而丙二醛含量最高的是沙枣,最低 的为头状沙拐枣和梭梭,除头状沙拐枣和梭梭之间差异不显著之外,其他植物种之间的差异均达到显著水平(P<0.05);4)在相关分析的基础上运用平均隶 属函数法对这4种植物进行抗旱性排序,结果表明,4种植物的抗旱性强弱顺序依次是梭梭>多枝柽柳>沙枣>头状沙拐枣. [ 张利刚, 曾凡江, 刘波, 刘镇, 安桂香, 李海峰, 袁娜 ( 2012). 绿洲-荒漠过渡带四种植物光合及生理特征的研究 草业学报, 21, 103-111.] DOIURL [本文引用: 1] 绿洲—荒漠过渡带植被的恢复和更新在过渡带生态环境建设中具有重 要意义.通过对多枝柽柳,梭梭,头状沙拐枣和沙枣的光合特性、水势(φ)以及生理指标的研究,结果表明,1)4种植物光响应曲线变化趋势基本一致,均表现 为净光合速率(A)随着光合有效辐射(PAR)的增大迅速增大,当PAR达到了植物的光饱和点之后,A出现下降的趋势;作为C4植物的头状沙拐枣和梭梭都 具有较高的光饱和点(LSP)和光补偿点(LCP),体现出了C4植物特有的高光合的优势,而其他一些光合参数K,Amax,Rday的变化规律并不完全 一致;2)4种植物中,梭梭具有较低的清晨和正午水势,说明梭梭主要依靠夜间水分恢复能力来应对干旱环境;清晨水势除头状沙拐枣和沙枣之间差异不显著外, 其他植物种之间的差异均达到了极显著水平(P<0.01),正午水势除多枝柽柳和沙枣之间差异不显著,其他植物种之间的差异也都达到了极显著水平 (P<0.01);3)4种植物中脯氨酸和可溶性糖的变化较为一致,头状沙拐枣均显著高于其他3种植物(P<0.05);而丙二醛含量最高的是沙枣,最低 的为头状沙拐枣和梭梭,除头状沙拐枣和梭梭之间差异不显著之外,其他植物种之间的差异均达到显著水平(P<0.05);4)在相关分析的基础上运用平均隶 属函数法对这4种植物进行抗旱性排序,结果表明,4种植物的抗旱性强弱顺序依次是梭梭>多枝柽柳>沙枣>头状沙拐枣.
ZhangSS ( 2014). Effect of Simulated Warming on Photosynthetic Adaptability and Reproductive Ecology of Invasive Solidago Canadensis, Bidens frondosa and Native Pterocypsela laciniata. Master degree dissertation, Southwest University Chongqing. [本文引用: 1]
ZhongCF, WuXY, YaoHJ, ShiZ, GaoRF ( 2008). Characteristics of chlorophyll a fluorescence kinetics of Euonymus japonicus during early spring in Beijing region Journal of Beijing Forestry University, 30, 9-15. URL [本文引用: 1] 该文利用调制式荧光仪与氧电极 技术,研究北京地区大叶黄杨在春初返青过程中的荧光动力学及光合放氧的变化,探讨常绿阔叶植物在北方越冬过程中抵御逆境的适应机制,同时借助稳态荧光动力 学曲线的变化揭示植物返青过程光暗反应的动态变化。结果表明:冬季大叶黄杨阳生叶与阴生叶的PSⅡ光化学效率、光合速率均很低;光合电子传递不通畅,低温 强光双重胁迫导致阳生叶比阴生叶的电子链阻塞更严重;捕光天线系统遭遇破坏,这是植物减少过多激发能对PSⅡ反应中心破坏的一种光保护机制;PSⅡ保持较 高的开放程度,借此减小激发能在供体侧积累对反应中心造成的伤害;同时非线性电子传递和天线系统之外的能量耗散是大叶黄杨越冬过程重要的光保护机制。初春 随着温度的回升,捕光天线系统、光合电子链的活性、光合速率逐渐恢复。倒春寒发生时,大叶黄杨再次发生光抑制现象。 [ 钟传飞, 武晓颖, 姚洪军, 施征, 高荣孚 ( 2008). 北京地区大叶黄杨春初返青过程的叶绿素荧光动力学研究 北京林业大学学报, 30, 9-15.] URL [本文引用: 1] 该文利用调制式荧光仪与氧电极 技术,研究北京地区大叶黄杨在春初返青过程中的荧光动力学及光合放氧的变化,探讨常绿阔叶植物在北方越冬过程中抵御逆境的适应机制,同时借助稳态荧光动力 学曲线的变化揭示植物返青过程光暗反应的动态变化。结果表明:冬季大叶黄杨阳生叶与阴生叶的PSⅡ光化学效率、光合速率均很低;光合电子传递不通畅,低温 强光双重胁迫导致阳生叶比阴生叶的电子链阻塞更严重;捕光天线系统遭遇破坏,这是植物减少过多激发能对PSⅡ反应中心破坏的一种光保护机制;PSⅡ保持较 高的开放程度,借此减小激发能在供体侧积累对反应中心造成的伤害;同时非线性电子传递和天线系统之外的能量耗散是大叶黄杨越冬过程重要的光保护机制。初春 随着温度的回升,捕光天线系统、光合电子链的活性、光合速率逐渐恢复。倒春寒发生时,大叶黄杨再次发生光抑制现象。 Temperature response of photosynthesis, excitation energy dissipation and alternative electron sinks to carbon assimilation in Beta vulgaris L 1 2006
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
克里雅河流域荒漠-绿洲交错带3种不同生活型植物的光合特性 1 2015
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
The rate of nitrite reduction in leaves as indicated by O2 and CO2 exchange during photosynthesis 1 2011
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Physiological response of the symbiotic gorgonian Eunicella singularis to a longterm temperature increase 1 2009
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Relationships between leaf conductance to CO2 diffusion and photosynthesis in micropropagated grapevine plants, before and after ex vitro acclimatization 1 2006
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri var. rupestris) 1 2009
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
A role for a light-harvesting antenna complex of photosystem II in photoprotection 1 2002
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
UV-B辐射增强对粳稻光合电子传递与光响应特性的影响 1 2016
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
UV-B辐射增强对粳稻光合电子传递与光响应特性的影响 1 2016
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Modelling photosynthesis of cotton grown in elevated CO2 1 1992
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
高CO2浓度和干旱胁迫对4种树苗光合特性的影响 1 2015
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error 1 2003
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
绿洲-沙漠过渡带引种植物光合生理特征研究 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
绿洲-沙漠过渡带引种植物光合生理特征研究 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Improving the estimation of mesophyll conductance to CO2: On the role of electron transport rate correction and respiration 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Structure and function of photosystems I and II 1 2006
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Regulation of guard cell photosynthetic electron transport by nitric oxide 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Low temperature effects on leaf physiology and survivorship in the C3 and C4 subspecies of Alloteropsis semialata 1 2008
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
On the mechanism underlying photosynthetic limitation upon trigger hair irritation in the carnivorous plant venus flytrap (Dionaea muscipula Ellis) 1 2011
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton 1 1980
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
2012年中国植物科学若干领域重要研究进展 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
2012年中国植物科学若干领域重要研究进展 1 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Rapid light curves: A powerful tool to assess photosynthetic activity 1 2005
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer onsite in the field 1 2000
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
A mechanistic model for the light response of photosynthetic electron transport rate based on light harvesting properties of photosynthetic pigment molecules 3 2013
... 光合作用作为地球上最重要的一个生理生化过程(许大全, 2013), 为植物的生长和发育提供了大部分物质和能量.植物的光合特性常用光合生理指标来衡量, 如光合参数: 净光合速率(Pn)、气孔导度(Gs)、光补偿点(Ic)、羧化效率(η)等(Lawlor & Cornic, 2002; 张永征等, 2013; 丁俊祥等, 2015), 荧光参数: 潜在光化学量子效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光系统II (PSII)的实际光化学量子效率(ΦPSII)等, 用光合电子流或电子传递速率(J)表示光合生理特征的较少.前人对以上参数的大小、变化趋势进行了比较研究, 主要用以探讨环境因素对植物光合特性的影响(曾志峰等, 2014)、不同品种间(姚春艳等, 2011)或同种群植物(简在友等, 2010)的光合特性差异, 或用来分析不同植物的光合特性差异(张利刚等, 2012; 罗维成等, 2013).光合作用的原初反应是将叶绿素吸收的光能转变成活跃的化学能储存在腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷磷酸中(Govindjee, 2002; Nelson & Yocum, 2006).光合电子流是植物光合能力形成的基础和碳同化的能量源泉.因此, 开展电子传递速率-光响应的研究对完善植物光合作用机理、提高光合效率具有重要的意义(Pavlovic et al., 2011; Martins et al., 2013; Ördög et al., 2013; 钱前等, 2013).植物发射出的叶绿素荧光信号中含有丰富的光合信息(康华靖等, 2015).通过叶绿素荧光测量仪器获取数据, 利用相关模型可估算出表示植物光合特性的相关参数, 如初始斜率(α)、饱和光强(Isat)、最大电子传递速率(Jmax)等(Fila et al., 2006; Ferrier-Pagès et al., 2009; Flexas et al., 2009; Eichelmann et al., 2011).α表示光能转化为电子流的最大能力; Isat的计算有利于了解植物发生PSII动力学下调或者光抑制的光照强度临界点, 可以为生产实践提供理论依据; 准确估算植物叶片的Jmax对研究植物叶片光呼吸过程在光保护中的作用是非常必要的.若高估Jmax, 将高估光合电子流分配到光呼吸的量, 从而高估光呼吸对植物的光保护作用(康华靖等, 2015; 郭巍等, 2016).植物叶片电子传递速率-光响应模型主要有: 双指数函数模型(Platt et al., 1980)、单指数函数模型(Harrison & Platt, 1986)、直角双曲线模型(Baly, 1935)和非直角双曲线模型(Thornley, 1976).这些经验模型大都使用测量的数据, 利用数学方法建模.不同的经验模型适应不同的植物: 双指数函数模型主要用于藻类电子传递速率-光响应曲线的拟合(Ralph & Gademann, 2005; Brading et al., 2011); 单指数函数模型主要用于藻类和高等植物(Rascher et al., 2000; Robakowski, 2005; Reynolds et al., 2012); 非直角双曲线模型主要用于高等植物(Caemmerer, 2000; Long & Bernacchi, 2003).经验模型的数学表达式较为简单, 使用方便.然而, 此类模型不涉及植物光合作用的原初反应, 也不涉及叶绿素分子的光能吸收、激发、电子传递和转化等过程.因此, 经验模型不能反映植物叶片电子传递对光响应的机理, 也不能直接计算光饱和点(闫小红等, 2013).此外, 经验模型不能拟合发生光抑制部分的光响应曲线, 同时这类模型无法解释植物的PSII动力学下调、光适应和光保护等现象(叶子飘等, 2014).Ye等(2013a, 2013b)提出了叶子飘模型, 该模型涉及叶绿素分子对光能的吸收、激发、退激发及电子传递等过程, 可拟合光抑制部分, 为更加准确分析植物的光合能力提供了可靠的检测方法.植物的光合作用除受自身生理特性的影响之外, 还受水分、光照、温度、湿度、CO2浓度等外界环境的影响(Bernacchi et al., 2005; Osborne et al., 2008; 刘娟娟等, 2015). ...
 ,1,*, 孟平3, 张劲松3, 张静茹1
,1,*, 孟平3, 张劲松3, 张静茹1
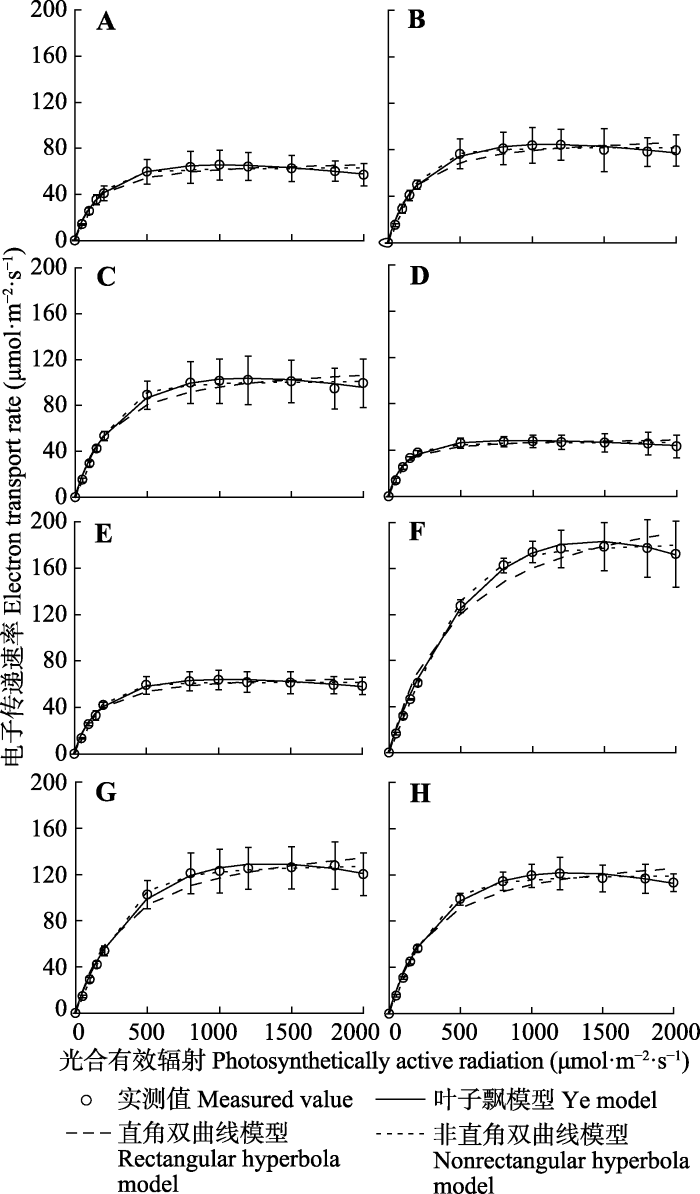 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT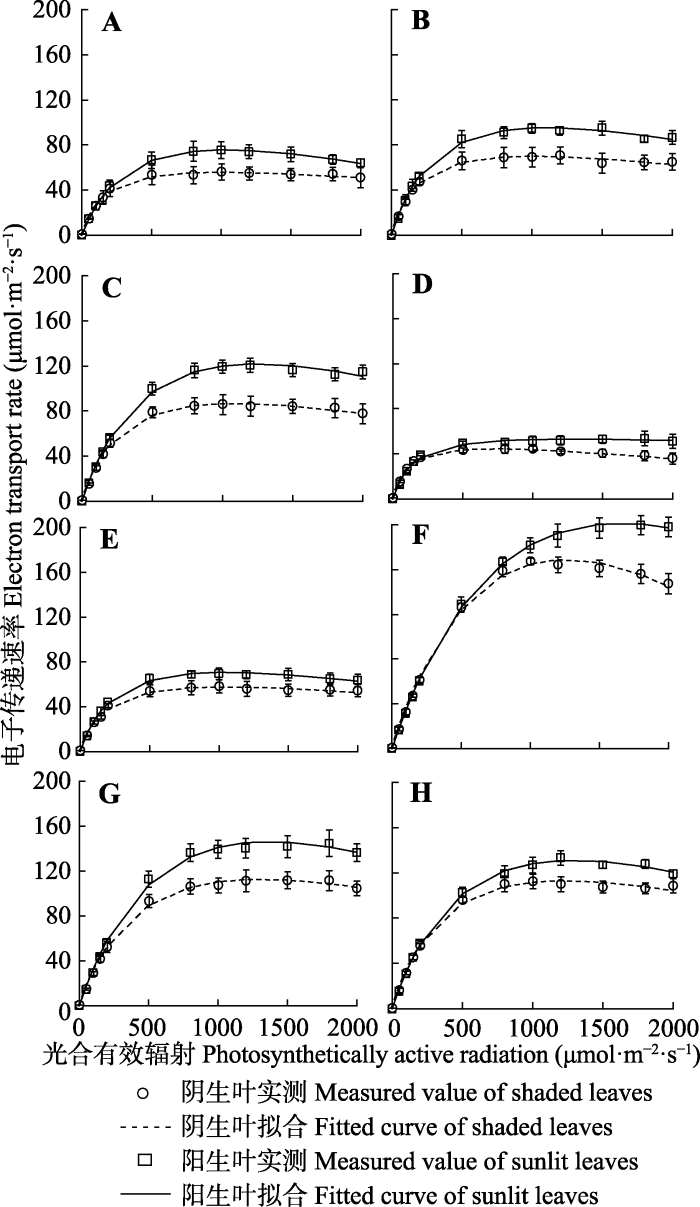 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}