Correlation between endogenous hormone and the adaptability of Chinese fir with high phosphorus-use efficiency to low phosphorus stress
Xian-Hua ZOU1,2, Ya-Nan HU1, Dan WEI1, Si-Tong CHEN1, Peng-Fei WU1,2, Xiang-Qing MA,1,2,*1 Forestry College, Fujian Agriculture and Forestry University, Fuzhou 350002, China 2 Chinese Fir Engineering Technology Research Center of State Forestry Administration, Fuzhou 350002, China
Abstract Aims Hormones are important signals for plants adaption to environmental stresses. To understand the mechanism of plants adaptation to nutrient deficiency from the perspective of hormone regulation is of great significance for breeding the genotypes with high phosphorus (P)-use efficiency. Methods This study investigated the correlation between hormone content and the adaptability of Chinese fir (Cunninghamia lanceolata) to low P stress by examining the changes of hormone content, root morphology, root dry matter and root P distribution patterns in the passive tolerance (M1) and active activation (M4) genotypes under low P stress at different treatment periods. Important findings No correlation was found between the foliar hormone contents and the adaptive characteristics of M1 and M4 under low P stress, although the root hormone content was significantly correlated with the growth index of roots. Low P stresses increased root IAA contents in M1 and M4 after 27 h of treatments and increased continuously with the prolongation of time. The IAA contents were positively correlated with surface area, volume and length of roots in both M1 and M4 (p < 0.05), suggesting that the increase of IAA induced root growth in both genotypes. Specifically, we observed an obvious phenomenon of IAA transportation from leaves to roots in M4, along with stronger root growth of M4 compared with that of M1. Meanwhile, low P stress increased the root-shoot ratio of M4, suggesting that root growth prompted more dry matter distribution to roots. Similarly, the ABA and GA3 contents in both M1 and M4 roots also increased as P availability decreased, but they showed a trend toward decrease over time and a negative correlation with root growth. The ZT contents in the root lower under low P treatment, yet there was no significant correlation between its contents and the low P adaptive characteristics of M1 and M4. Our results indicated that the contents of root IAA, ABA, and GA3 in Chinese fir clones with high P-use efficiency were closely related to the morphological changes of the roots. These comprehensive regulations of different organs is an essential survival strategy for plants to adapt to low P stress. Keywords:low phosphorus stress;endogenous hormone;high phosphorus-use efficiency;Chinese fir;root morphology;root shoot ratio;nutrient distribution
PDF (1631KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 邹显花, 胡亚楠, 韦丹, 陈思同, 吴鹏飞, 马祥庆. 磷高效利用杉木对低磷胁迫的适应性与内源激素的相关性. 植物生态学报, 2019, 43(2): 139-151. DOI: 10.17521/cjpe.2018.0201 ZOU Xian-Hua, HU Ya-Nan, WEI Dan, CHEN Si-Tong, WU Peng-Fei, MA Xiang-Qing. Correlation between endogenous hormone and the adaptability of Chinese fir with high phosphorus-use efficiency to low phosphorus stress. Chinese Journal of Plant Ecology, 2019, 43(2): 139-151. DOI: 10.17521/cjpe.2018.0201
杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005)。近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011)。研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率。此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018)。在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显。激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究。低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关?
Fig. 2Endogenous hormone contents of Cunninghamia lanceolata M1 at different sampling periods (mean ± SD). A, IAA content. B, ABA content. C, GA3 content. D, ZT content. L-P and H-P represent the low and high phosphorus treatments, respectively. Different lower- and upper-case letters indicate significant differences (p < 0.05) in each variables across different treatment periods under L-P and H-P conditions, respectively.
Fig. 3Endogenous hormone content of Cunninghamia lanceolata M4 at different sampling periods (mean ± SD). A, IAA content. B, ABA content. C, GA3 content. D, ZT content. L-P and H-P represent the low and high phosphorus treatments, respectively. Different lower- and upper-case letters indicate significant differences (p < 0.05) in each variables across different treatment periods under L-P and H-P conditions, respectively.
Fig. 4Root morphological changes of different Cunninghamia lanceolata with different high phosphorus-use efficiency at different sampling periods (mean ± SD). A, Root length increments of M1. B, Root length increments of M4. C, Root surface area increments of M1. D, Root surface area increments of M4. E, Root volume increments of M1. F, Root volume increments of M4. G, Averaged root diameter of M1. H, Averaged root diameter of M4. L-P and H-P represent the low and high P treatments, respectively. Different lower- and upper-case letters indicate significant differences (p < 0.05) in each variables across different treatment periods under L-P and H-P conditions, respectively.
Fig. 5Root/shoot ratio of different Cunninghamia lanceolata with different high P-use efficiency at different sampling periods (mean ± SD). A, Root/shoot ratio of M1. B, Root/shoot ratio of M4. L-P and H-P represent the low and high P treatments, respectively. Different lower- and upper-case letters indicate significant differences (p < 0.05) across different treatment periods under L-P and H-P conditions, respectively.
Fig. 6Phosphorous distribution patterns of Cunninghamia lanceolata with different high P-use efficiency at different sampling periods (mean ± SD). A, Phosphorous distribution patterns in the aerial part and roots of M1. B, Phosphorous distribution patterns in the aerial part and roots of M4. L-P and H-P represent the low and high P treatments, respectively. Different lower- and upper- case letters indicate significant differences (p < 0.05) across different treatment periods under L-P and H-P conditions, respectively.
Table 1 表1 表1不同磷高效利用杉木内源激素与苗木生长特性的相关性分析 Table 1Correlation between endogenous hormones and growth characteristics of Chinese fir clones with high phosphorus-use efficiency under different phosphorus levels
家系 Clone
部位 Organ
供磷处理 Phosphorus supply level
内源激素 Endogenous hormone
根体积 Root volume
根平均直径 Average root diameter
根表面积 Root surface area
根长 Root length
根冠比 Root/shoot ratio
地上部磷养分含量 P content in the aerial parts
根磷养分含量 P content in the roots
M1
叶片 Leaves
L-P
ABA
-0.159
-0.161
-0.186
-0.306
0.289
0.313
0.103
IAA
-0.075
0.100
0.046
0.136
-0.190
-0.072
0.046
GA3
0.525
-0.674
0.550
0.221
-0.655
0.518
-0.018
ZT
-0.644
0.443
-0.663
-0.495
0.382
-0.170
0.604
H-P
ABA
-0.019
0.372
-0.103
-0.473
0.157
-0.540
0.352
IAA
0.250
-0.548
0.364
0.435
-0.186
0.546
0.006
GA3
0.313
-0.571
0.365
0.131
0.045
0.545
0.517
ZT
-0.391
0.701
-0.423
-0.210
0.589
-0.208
0.237
根系 Roots
L-P
ABA
-0.958**
0.442
-0.921**
-0.863*
0.077
-0.073
0.682
IAA
0.891**
-0.777*
0.880**
0.627
-0.245
0.381
-0.675
GA3
-0.794*
0.558
-0.797*
-0.715
-0.316
-0.317
0.660
ZT
-0.554
0.276
-0.564
-0.482
-0.415
-0.346
0.693
H-P
ABA
-0.612
0.416
-0.604
-0.695
0.538
-0.690
0.302
IAA
0.672
-0.456
0.619
0.128
-0.599
-0.180
-0.230
GA3
-0.370
0.427
-0.360
-0.409
0.029
-0.402
-0.035
ZT
-0.383
0.364
-0.360
-0.374
-0.010
-0.359
-0.110
M4
叶片 Leaves
L-P
ABA
-0.106
0.430
-0.205
-0.170
-0.258
-0.167
-0.602
IAA
-0.045
-0.239
0.006
-0.205
-0.027
-0.099
0.572
GA3
-0.130
0.302
-0.186
0.066
0.120
-0.068
-0.341
ZT
-0.344
0.563
-0.432
-0.376
-0.071
-0.281
-0.336
H-P
ABA
-0.146
0.494
-0.138
-0.106
0.107
-0.123
-0.310
IAA
0.338
-0.394
0.393
0.453
-0.278
0.012
0.219
GA3
-0.524
0.584
-0.466
-0.331
0.440
0.336
0.065
ZT
0.054
-0.361
0.078
0.103
0.230
-0.038
-0.357
根系 Roots
L-P
ABA
-0.079
0.368
-0.114
-0.120
-0.170
-0.248
-0.625
IAA
0.772*
-0.876**
0.963**
0.810*
-0.647
0.481
-0.906**
GA3
-0.380
0.248
-0.357
-0.429
0.081
-0.542
0.223
ZT
-0.434
0.286
-0.402
-0.473
0.157
-0.533
0.282
H-P
ABA
0.450
0.003
0.425
0.184
-0.242
-0.606
-0.474
IAA
0.692
-0.475
0.735
0.689
-0.512
-0.313
-0.694
GA3
-0.291
0.138
-0.356
-0.285
0.280
-0.506
0.086
ZT
-0.497
0.111
-0.481
-0.266
0.362
0.126
0.644
L-P represents the low-P treatment and H-P represents the high-P control treatment. * for a significant correlation at 0.05 level (bilateral), ** for a significant correlation at 0.01 level (bilateral). L-P代表低磷处理, H-P代表高磷对照。*表示在0.05水平(双侧)上显著相关, **表示在0.01水平(双侧)上极显著相关。
AlexovaR, MillarAH ( 2013). Proteomics of phosphate use and deprivation in plants Proteomics, 13, 609-623. DOI:10.1002/pmic.v13.3-4URL [本文引用: 1]
BrennerC, DeplusR, DidelotC, LoriotA, ViréE, SmetCD, GutierrezA, DanoviD, BernardD, BoonT ( 2005). Myc represses transcription through recruitment of DNA methyltransferase corepressor EMBO Journal, 24, 336-346. DOI:10.1038/sj.emboj.7600509URL [本文引用: 2]
ChenBL, LuoJ, JiangPA ( 2016). Effects of different phosphorus concentration on endogenous hormones of cotton Southwest China Journal of Agricultural Sciences, 29, 1839-1843. [本文引用: 5]
CiereszkoI, KleczkowskiLA ( 2002). Effects of phosphate deficiency and sugars on expression of rab18 in Arabidopsis: Hexokinase-dependent and okadaic acid-sensitive transduction of the sugar signal. Biochimica et Biophysica Acta (BBA)—Gene Structure and Expression, 1579, 43-49. [本文引用: 3]
DevaiahBN, MadhuvanthiR, KarthikeyanAS, RaghothamaKG ( 2009). Phosphate starvation responses and gibberellic acid biosynthesis are regulated by the MYB62 transcription factor in Arabidopsis. Molecular Plant, 2, 43-58. [本文引用: 3]
DongJ, MouP ( 2012). Root nutrient foraging of morphological plasticity and physiological mechanism in Callistephus chinensis. Chinese Journal of Plant Ecology, 36, 1172-1183. [本文引用: 2]
Franco-ZorrillaJM, MartinAC, SolanoR, RubioV, LeyvaA, Paz-AresJ ( 2002). Mutations at CRE1 impair cytokinin-?induced repression of phosphate starvation responses in Arabidopsis. Plant Journal, 32, 353-360. [本文引用: 4]
FuYP, YangY, XueJB ( 2005). Influence under low P-deficient stress on endogenous hormone and root activities of dark sun-cured tobacco Chinese Agricultural Science Bulletin , 21, 227-229. [本文引用: 3]
GiehlRFH, GruberBD, von WirenN ( 2014). Its time to make changes: Modulation of root system architecture by nutrient signals Journal of Experimental Botany, 65, 769-778. DOI:10.1093/jxb/ert421URL [本文引用: 1]
HuangRH, YangHL, HuangW, LuYM, ChenK ( 2015). Effects of Funneliformis mosseae on endogenous hormones and photosynthesis of Sorghum haipense under Cs stress. Chinese Journal of Applied Ecology, 26, 2146-2150. [本文引用: 1]
LiuH, WangSG ( 2003). Influences of P deficiency stress on endogenous hormones in Barley. Journal of Southwest Agricultural University, 25, 48-51. [本文引用: 2]
LiuHC, KuangYH, ChenRY ( 2003). Changes of IAA contents in different Asparagus Bean cultivars under phosphorus- deficient stress. Plant Physiology Communications, 39, 125-127. [本文引用: 1]
MartinAC, Del PozoJC, IglesiasJ, RubioV, SolanoR, de La PenaA, LeyvaA, Paz-AresJ ( 2000). Influence of cytokinins on the expression of phosphate starvation responsive genes in Arabidopsis. The Plant Journal, 24, 559-567. [本文引用: 2]
MissonJ, RaghothamaKG, JainA, JouhetJ, BlockMA, BlignyR, OrtetP, CreffA, SomervilleS, RollandN ( 2005). A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. Proceedings of the National Academy of Sciences of the United States of America, 102, 11934-11939. [本文引用: 1]
MiuraK, LeeJ, GongQ, MaS, JinJB, YooCY, MiuraT, SatoA, BohnertHJ, HasegawaPM ( 2011). SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation Plant Physiology, 155, 1000-1012. DOI:10.1104/pp.110.165191URL [本文引用: 4]
MorcuendeR, BariR, GibonY, ZhengW, PantBD, BlasingO, UsadelB, CzechowskiT, UdvardiMK, StittM, ScheibleW ( 2007). Genome-wide reprogramming of metabolism and regulatory networks of Arabidopsis in response to phosphorus. Plant, Cell & Environment, 30, 85-112. [本文引用: 1]
NacryP, CanivencG, MullerB, AzmiA, Van OnckelenH, RossignolM, DoumasP ( 2005). A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. Plant Physiology, 138, 2061-2074. [本文引用: 1]
Pérez-TorresCA, López-BucioJ, Cruz-RamírezA, Ibarra- LacletteE, DharmasiriS, EstelleM, Herrera-EstrellaL ( 2008). Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. Plant Cell, 20, 3258-3272. [本文引用: 2]
PostmaJA, LynchJP ( 2011). Theoretical evidence for the functional benefit of root cortical aerenchyma in soils with low phosphorus availability Annals of Botany, 107, 829-841. DOI:10.1093/aob/mcq199URL [本文引用: 1]
RadinJW, ParkerLL, GuinnG ( 1982). Water relations of cotton plants under nitrogen deficiency: V. Environmental control of abscisic acid accumulation and stomatal sensitivity to abscisic acid Plant Physiology, 70, 1066-1070. DOI:10.1104/pp.70.4.1066URL [本文引用: 1]
ShenY, ZhangY, LinH, GaoS, PanG ( 2012). Effect of low phosphorus stress on endogenous hormone levels of different maize genotypes in seedling stage Journal of Biological Sciences, 12, 208-314. [本文引用: 3]
ShengWT, FanSH (2005). Long-term Productivity of Chinese fir Plantations. Science Press, Beijing. [本文引用: 3]
WeiXW, GouC, XuMZ, XuHW, ZhouXF ( 2013). Study on the improvement of HPLC method for analyzing endogenous hormone in Maize. Journal of Maize Sciences, 21, 144-148. [本文引用: 1]
WuPF, MaXQ, TigabuMT, WangCW, LiuAQ, OdenPC ( 2011). Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress Canadian Journal of Forest Research, 41, 228-234. DOI:10.1139/X10-198URL [本文引用: 6]
WuPF, WangGY, El-KassabyYA, WangP, ZouXH, MaXQ ( 2016). Solubilization of aluminum-bound phosphorus by root cell walls: Evidence from Chinese fir, Cunninghamia lanceolata(Lamb.) Hook. Canadian Journal of Forest Research, 47, 419-423. [本文引用: 1]
WuPF, WangGY, FarooqTH, LiQ, ZouXH, MaXQ ( 2017). Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? Journal of Soils and Sediments, 17, 2775-2785. DOI:10.1007/s11368-017-1852-8URL [本文引用: 1]
WuPF, ZangGZ, MaXQ ( 2006). Advances in the mechanism of chemical communication of plants under stress Subtropical Agriculture Research , 2, 271-277. [本文引用: 1]
YamagishiM, ZhouK, OsakiM, MillerSS, VanceCP ( 2011). Real-time RT-PCR profiling of transcription factors including 34 MYBs and signaling components in white lupin reveals their P status dependent and organ-specific expression Plant and Soil, 342, 481-493. DOI:10.1007/s11104-010-0711-9URL [本文引用: 1]
YangCC, HuangQJ, SuXH ( 2013). Correlation between endogenous IAA, ABA contents and height growth of black poplar at the seedling stage Scientia Silvae Sinicae , 49(8), 35-42. DOI:10.11707/j.1001-7488.20130806Magsci [本文引用: 1] <p>以黑杨派3个无性系为材料对其生长进行定位观测,用石蜡组织切片技术研究伸长节间和成熟节间的结构区别,并用GC-MS-SIM法测定幼叶等有代表性组织的内源IAA和ABA含量。结果表明: 1) 节间数月增量与苗高月增量间呈显著正相关,第1和第2节间总长与苗高月增量、IAA含量、ABA含量及IAA/ABA无显著相关; 2) 伸长节间细胞较小,排列紧密,次生木质部不明显; 成熟节间细胞较大,排列松散,有明显的次生木质部; 3) 5—7月,凌丰1号3个幼嫩组织部位的内源IAA含量、ABA含量、IAA/ABA、节间数月增量和苗高月增量都在增大; 7月,3个无性系的节间数月增量和苗高月增量的排列顺序与幼嫩组织的IAA含量没有相关性,与幼叶的ABA含量呈显著负相关,与顶芽的IAA/ABA呈显著正相关。不能用幼叶等组织的内源IAA含量来预测多个无性系苗高顺序,幼叶的ABA含量可作为预测多个无性系苗高顺序的参考指标,建议用顶芽的IAA/ABA对杨树苗高进行早期预测。</p> [ 杨成超, 黄秦军, 苏晓华 ( 2013). 内源激素IAA和ABA含量与黑杨苗期高生长关系 林业科学, 49(8), 35-42.] DOI:10.11707/j.1001-7488.20130806Magsci [本文引用: 1] <p>以黑杨派3个无性系为材料对其生长进行定位观测,用石蜡组织切片技术研究伸长节间和成熟节间的结构区别,并用GC-MS-SIM法测定幼叶等有代表性组织的内源IAA和ABA含量。结果表明: 1) 节间数月增量与苗高月增量间呈显著正相关,第1和第2节间总长与苗高月增量、IAA含量、ABA含量及IAA/ABA无显著相关; 2) 伸长节间细胞较小,排列紧密,次生木质部不明显; 成熟节间细胞较大,排列松散,有明显的次生木质部; 3) 5—7月,凌丰1号3个幼嫩组织部位的内源IAA含量、ABA含量、IAA/ABA、节间数月增量和苗高月增量都在增大; 7月,3个无性系的节间数月增量和苗高月增量的排列顺序与幼嫩组织的IAA含量没有相关性,与幼叶的ABA含量呈显著负相关,与顶芽的IAA/ABA呈显著正相关。不能用幼叶等组织的内源IAA含量来预测多个无性系苗高顺序,幼叶的ABA含量可作为预测多个无性系苗高顺序的参考指标,建议用顶芽的IAA/ABA对杨树苗高进行早期预测。</p>
YuanY, HuangLQ, LüDM, MaoY, FuGF ( 2008). Effect of low pH on endogenous hormones and root development in Chinese medicine Atractylodes lancea(Thunb.) DC. Chinese Pharmaceutical Journal, 43, 101-104. [本文引用: 1]
ZouXH, WeiD, WuPF, ZhangY, HuYN, ChenST, MaXQ ( 2018). Strategies of organic acid production and exudation in response to low-phosphorus stress in Chinese fir genotypes differing in phosphorus-use efficiencies Trees, 32, 897-912. DOI:10.1007/s00468-018-1683-2 [本文引用: 2]
ZouXH, WuPF, ChenNL, WangP, MaXQ ( 2015). Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil Canadian Journal of Forest Research, 45, 402-410. DOI:10.1139/cjfr-2014-0384URL [本文引用: 2]
ZouXH, WuPF, JiaYY, MaJ, MaXQ ( 2016). Periodical response of Chinese fir root to the phosphorus concentrations in patches and heterogeneous distribution in different growing stages Journal of Plant Nutrition and Fertilizer, 22, 1056-1063. [本文引用: 1]
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
Signaling network in sensing phosphate availability in plants 1 2011
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
Effects of phosphate deficiency and sugars on expression of rab18 in Arabidopsis: Hexokinase-dependent and okadaic acid-sensitive transduction of the sugar signal. 3 2002
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
翠菊根系养分捕获形态塑性及其生理机制 2 2012
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
Mutations at CRE1 impair cytokinin-?induced repression of phosphate starvation responses in Arabidopsis. 4 2002
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... ).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
低磷胁迫对晒红烟内源激素和根系活力的影响 3 2005
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
... )、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
低磷胁迫对晒红烟内源激素和根系活力的影响 3 2005
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
... )、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
Hormone interactions during lateral root formation 1 2009
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
Its time to make changes: Modulation of root system architecture by nutrient signals 1 2014
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
核素铯胁迫下接种摩西球囊霉对宿根高粱内源激素和光合的影响 1 2015
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
核素铯胁迫下接种摩西球囊霉对宿根高粱内源激素和光合的影响 1 2015
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. 4 2007
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... ).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
Influence of cytokinins on the expression of phosphate starvation responsive genes in Arabidopsis. 2 2000
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. 1 2005
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation 4 2011
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
Genome-wide reprogramming of metabolism and regulatory networks of Arabidopsis in response to phosphorus. 1 2007
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. 1 2005
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. 2 2008
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
Theoretical evidence for the functional benefit of root cortical aerenchyma in soils with low phosphorus availability 1 2011
Water relations of cotton plants under nitrogen deficiency: V. Environmental control of abscisic acid accumulation and stomatal sensitivity to abscisic acid 1 1982
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
Plant hormones and nutrient signaling 4 2009
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
... )等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
... ), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
Cytokinins: Activity, biosynthesis, and translocation 1 2006
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
1 1986
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
1 1986
... 植物激素是由植物细胞接受一定的信号诱导, 在植物体内合成的微量生理活性有机物质, 通常是在植物体内合成, 从合成部位运往作用部位, 对植物的生长发育、物质代谢和形态建成等各个方面均起着重要的信号传导和调节作用, 是植物适应逆境的重要信号物质(邵莉楣和郝斌, 1986).研究表明, 植物体内磷的缺乏将导致激素平衡的改变(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过多种形态及生理途径来实现对低磷环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012).IAA (Miura et al., 2011)、ZT (Franco-Zorrilla et al., 2002; Brenner et al., 2005)、GA3 (Jiang et al., 2007)、ABA (Ciereszko & Kleczkowski, 2002)等常见激素均被证实参与了磷饥饿反应. ...
Effect of low phosphorus stress on endogenous hormone levels of different maize genotypes in seedling stage 3 2012
... 其中, IAA信号与缺磷诱导的根构型重塑有着紧密的联系(Miura et al., 2011), 是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008).孙海国和张福锁(2000)、符云鹏等(2005)、Shen等(2012)、陈洁等(2013)、陈智裕等(2016)相继报道低磷环境下, IAA含量增加, 从而诱导根系生长.有****认为, 这主要是因为IAA的极性运输提高了根部生长素的浓度, 并增强了化合物向根中的分配力度(李俊华和种康, 2006), IAA又能诱导细胞周期蛋白基因cyclAt的表达, 促进根分生组织的分裂, 加速根的伸展(孙海国和张福锁, 2000).本研究中, 低磷处理条件下磷高效利用杉木M1与M4根系中的IAA含量自27 h起高于高磷对照, 且呈明显的逐渐上升趋势.M1与M4的根系IAA含量均表现为与根表面积、根体积及根长等根系生长指标呈显著正相关关系, 与平均直径呈显著负相关关系.IAA含量的增加诱导了根系增长, M1与M4均表现出一定的根系增长量, 但M4的根系增生表现得比M1更明显.M4的根长、根表面积及根体积增量均高于高磷对照, 平均直径小于高磷对照.同时, 生长素自上而下向基部积累也促使更多的干物质量分配到根系, 在整个处理过程中, M4的根冠比均高于高磷对照.与生长素相反, 细胞分裂素被证明消极地调节大量的磷饥饿基因的表达, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).本研究中M1与M4叶及根系中的ZT含量表现为低磷处理逐渐低于高磷对照, 且随时间延长呈明显下降趋势, 其中, M1的ZT含量降低先于M4.与前人在大麦(刘辉和王三根, 2003)、烟草(符云鹏等, 2005)、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
... )、玉米(Shen et al., 2012; 陈洁等, 2013)、欧洲油菜(Shi et al., 2012)、棉花(陈波浪等, 2016)等的研究结果一致.有研究认为, 磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-Zorrilla et al., 2002), 从而抑制了细胞分裂素的活性(Brenner et al., 2005).鉴于细胞分裂素的移动特性(Sakakibara, 2006)及磷饥饿诱导基因影响范围的广泛性(Martin et al., 2000), 细胞分裂素被认为是无机磷酸盐信号的系统抑制(Lai et al., 2007).在本研究中M1的根系生长量低于高磷对照.可见, 虽然低磷条件下M1与M4根系的IAA含量均呈增加趋势, 但M4的增加强度高于M1, 而M1的ZT含量降低却先于M4.由此, 我们推测, M1中ZT的负调节作用大于IAA的正调控作用, 最终表现为根系增生能力并未显著提高.已有研究证实, 植物的生长发育受到各内源激素在植物体内的含量水平及相互平衡状况的调节.在营养胁迫条件下, 植株也是通过改变其体内的各种激素含量及其比例来维持正常生长(师晨娟等, 2006). ...
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress 6 2011
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... ), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... ).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
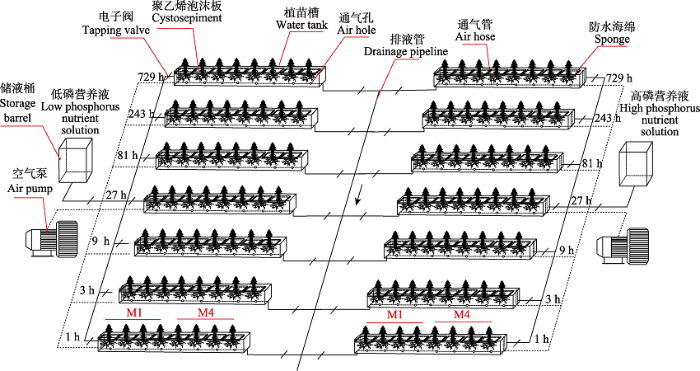
... 本研究选择课题组前期筛选出的1号(M1)与4号(M4)杉木苗木为试验材料, 其中, M1为“被动忍受低磷”基因型杉木, 其在土壤有效磷低于一般基因型所需的浓度时仍有较高的产量, 主要通过加快体内磷素的内循环来抵抗磷胁迫逆境; M4为“主动捕获土壤磷”基因型杉木, 主要通过根系拓展增生和分泌化学物质来适应环境的磷胁迫(Wu et al., 2011).供试苗木为福建漳平五一国有林场杉木无性系种子园按单系采种培育的半同胞家系苗.在温室内培育5个月, 温室平均气温20.3 ℃, 相对湿度78%.供试苗木生长势均一, 根系完整, 无病害, 平均地径3.30 mm, 平均苗高15.96 cm. ...
Solubilization of aluminum-bound phosphorus by root cell walls: Evidence from Chinese fir, Cunninghamia lanceolata(Lamb.) Hook. 1 2016
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? 1 2017
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
逆境中植物化学通讯机制的研究进展 1 2006
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
逆境中植物化学通讯机制的研究进展 1 2006
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
植物激素调控研究进展 1 2009
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
植物激素调控研究进展 1 2009
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
Real-time RT-PCR profiling of transcription factors including 34 MYBs and signaling components in white lupin reveals their P status dependent and organ-specific expression 1 2011
... 生长素(IAA)(Miura et al., 2011)、细胞分裂素(ZT)(Franco-Zorrilla et al., 2002)、赤霉素(GA3)(Jiang et al., 2007)、脱落酸(ABA)(Ciereszko & Kleczkowski, 2002)等植物激素均被研究证实参与了磷饥饿反应.其中, IAA是缺磷条件下刺激侧根原基出现的必须条件(Pérez-Torres et al., 2008), 在侧根初始发育和侧根原基形成时期, IAA起着正调控作用(Fukaki, 2009).有研究报道, 低磷环境下, 生长素含量的增加诱导了根系的增生(陈洁等, 2013).Nacry等(2005)通过对白羽扇豆(Lupinus albus)外施生长素发现, 外施生长素引起了白羽扇豆根系形态的变化, 这与植物在低磷逆境中的表现相同, 反之, 白羽扇豆的簇生根则在施用生长素转运抑制剂的情况下受到了抑制.Yamagishi等(2011)也证实部分参与生长素合成与信号转导的基因在白羽扇豆簇生根形成过程中大量表达.可见, 低磷胁迫条件下, 植物生长素的合成、运输与信号传导对其根系形态的改变起着十分重要的作用(Miura et al., 2011).有研究表明, 细胞分裂素信号在调节磷饥饿应答中也起着关键性的作用.磷饥饿通过降低细胞分裂素的浓度及减少细胞分裂素受体CRE1的表达(Franco-?Zorrilla et al., 2002), 抑制了细胞分裂素的活性.同时, 细胞分裂素消极地调节大量的磷饥饿基因的表达.研究发现, 增加细胞分裂素的浓度显著抑制了磷饥饿诱导基因(PSI)的表达, 且对侧根的生长产生一定的影响(Martin et al., 2000).在大麦(Hordeum vulgare)(刘辉和王三根, 2003)、烟草(Nicotiana tabacum)(符云鹏等, 2005)、玉米(Zea mays)(陈洁等, 2013)、欧洲油菜(Brassica napus)(Shi et al., 2012)、棉花(Gossypium hirsutum)(陈波浪等, 2016)等大量植物中, 细胞分裂素浓度均被证实在低磷胁迫条件下显著降低, 在植物适应低磷胁迫过程中起负调节作用(Li et al., 2010).此外, 研究认为赤霉素及其下游蛋白组成的信号途径与生长素共同构成的信号途径在低磷胁迫条件下侧根的形成过程中起正调节作用(Jiang et al., 2007).在低磷信号转导过程中, 磷饥饿导致GA生物活性水平下降和DELLA蛋白的积累(Devaiah et al., 2009), 并导致磷吸收、酸性磷酸酶活性和茎中磷含量下降(Devaiah et al., 2009).另有推断认为ABA信号参与了磷饥饿反应(Ciereszko & Kleczkowski, 2002).Radin等(1982)发现低磷胁迫下, ABA在棉花叶片中大量累积.陈波浪等(2016)的研究也表明, 低磷胁迫促使棉株体内的ABA含量显著提升. ...
内源激素IAA和ABA含量与黑杨苗期高生长关系 1 2013
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
内源激素IAA和ABA含量与黑杨苗期高生长关系 1 2013
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
低pH对苍术根组织内源激素水平和生长发育的影响 1 2008
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
低pH对苍术根组织内源激素水平和生长发育的影响 1 2008
... 磷是植物生长发育所必需的大量营养元素之一, 不仅参与组成植物体内众多重要的化合物, 也是植物体内能量载体的主要组成成分和能量的提供者(Vitousek et al., 2010).然而, 由于土壤中大量的磷易被金属氧化物或碳酸盐化合物吸附固定, 大量的磷以无效态储备在土壤中, 而植物可利用的有效磷含量亏缺(Johnston et al., 2014).在土壤缺磷条件下, 植物在长期的进化过程中为了增加在低磷环境完成生命周期的机会, 表现出对低磷环境的多种适应方式和适应特性(Chiou & Lin, 2011; Alexova & Millar, 2013; Giehl et al., 2014).其中, 在植物的磷反应调控体系中, 激素的作用被大量研究所证实(Wu et al., 2003; Misson et al., 2005; Morcuende et al., 2007).研究发现植物个体抵抗营养胁迫的化学通讯现象主要是以激素为化学信号物质(吴鹏飞等, 2006), 植物激素处于诸多调控信号所构成的网络系统的中心(熊国胜等, 2009).植物体内缺乏磷会影响激素的产生、敏感性和转运(Rubio et al., 2009), 而激素平衡的改变, 促使植物通过调节根系形态(袁媛等, 2008)、降低光合速率(黄仁华等, 2015)、调整生物量分配(杨成超等, 2013)等途径来实现对养分匮乏环境的适应(Rubio et al., 2009; 董佳和牟溥, 2012). ...
Strategies of organic acid production and exudation in response to low-phosphorus stress in Chinese fir genotypes differing in phosphorus-use efficiencies 2 2018
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil 2 2015
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
... ; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
杉木根系对不同磷斑块浓度与异质分布的阶段性响应 1 2016
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
杉木根系对不同磷斑块浓度与异质分布的阶段性响应 1 2016
... 杉木(Cunninghamia lanceolata)作为中国南方的主要造林树种, 其连栽地力衰退导致生产力下降的问题引起大量****的重视, 鉴于南方酸性土壤对磷的强烈固定作用, 研究认为土壤有效磷含量低是造成这一问题的重要因素(盛炜彤和范少辉, 2005).近年来, 针对低磷胁迫条件下杉木的适应策略已开展了大量研究(Wu et al., 2011, 2016; Zou et al., 2015; 陈智裕等, 2016), 并筛选出不同的磷高效利用基因型杉木(Wu et al., 2011).研究表明, 磷高效利用基因型杉木在低磷环境中, 可通过根系形态的变化(Wu et al., 2011; Zou et al., 2015; 邹显花等, 2016), 增强其对土壤有效磷的寻觅能力, 并通过分泌有机酸(Wu et al., 2017; Zou et al., 2018)等化学物质提高对土壤难溶性磷的活化效率.此外, 还可通过将根系中的磷转运至茎叶重新利用, 从而加速植物体内的磷循环使植株对有限磷资源的利用更趋经济高效(Zou et al., 2018).在以上对低磷环境的形态生理适应过程中, 不同杉木基因型存在显著差异, 以致土壤磷利用效率的种内差异明显.激素作为植物适应逆境的重要信号物质, 目前对这些基因型在低磷胁迫条件下的激素调控机制却尚未开展相关研究.低磷条件下, 不同磷高效利用杉木基因型体内的激素平衡是否被打破? 而激素平衡的改变与各适应性反应之间是否密切相关? ...
 ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT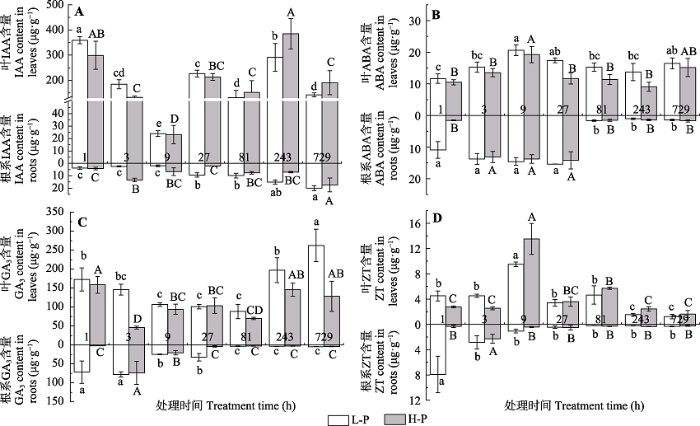 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT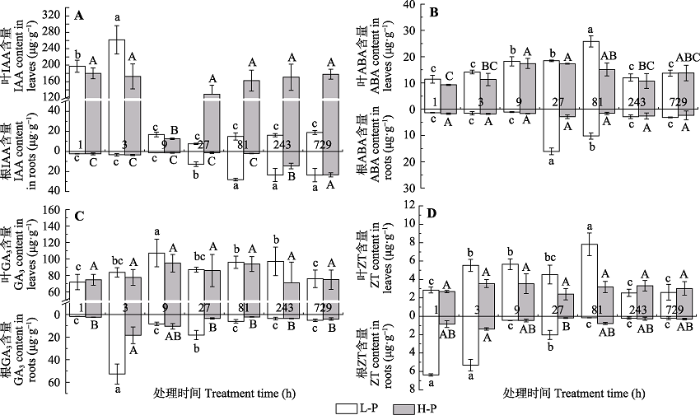 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT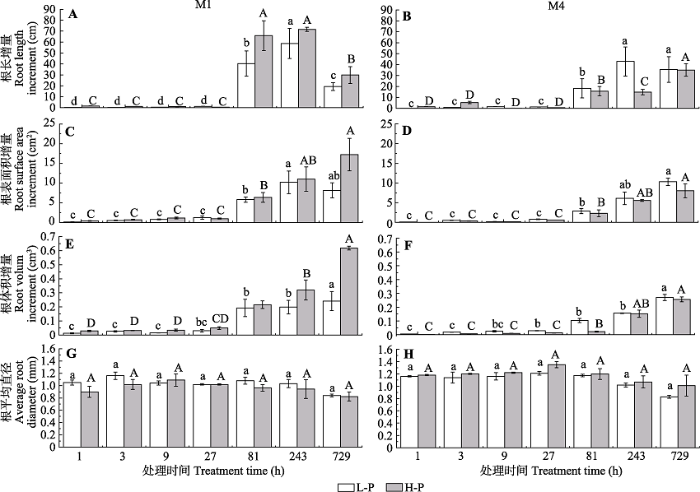 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT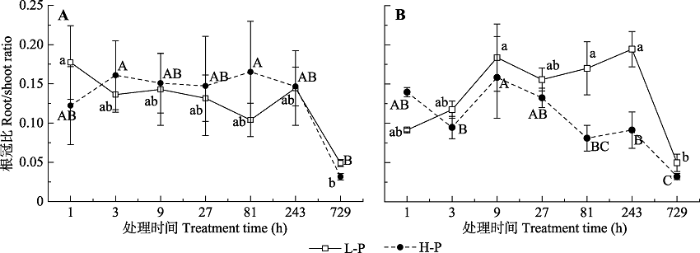 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT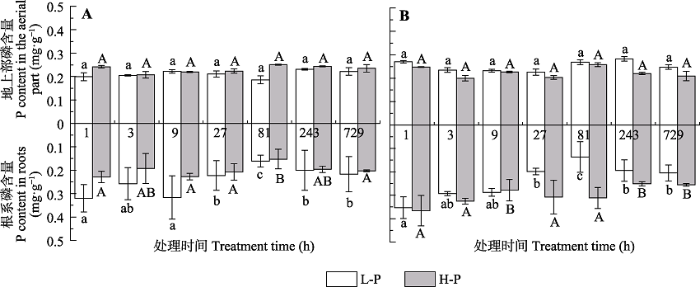 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}