 , 李俐蓉, 焦骄, 肖卫
, 李俐蓉, 焦骄, 肖卫西北师范大学地理与环境科学学院, 兰州 730070
Exploring the community assembly of subalpine meadow communities based on functional traits and community phylogeny
CHEYing-Di, LIUMin-Xia, LILi-Rong, JIAOJiao, XIAOWei通讯作者:
收稿日期:2017-08-10
接受日期:2017-09-29
网络出版日期:2017-11-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods We selected five slope-oriented plots to construct a super-tree representing the species pool. We surveyed the leaf functional traits and soil environmental factors in different slopes. Then we tested the phylogenetic signal of leaf dry matter content (LDMC), specific leaf area (SLA), leaf nitrogen content (LNC) and leaf phosphorus content (LPC).
Important findings The changes of slope aspect had significant influence on soil water content (SWC) and soil nutrient content. Most of the plants leaf functional traits had significant difference along different slope aspects. The LDMC was higher in south and southwest slope than north slope, while SLA, LNC and LPC were relatively high in north and northwest slope. The LPC showed feeble phylogenetic signal, while LDMC, SLA, LNC did not have a significant phylogenetic signal. With changes in the slope aspect from south to north, community phylogenetic structure shifted from over-dispersion to clustered dispersion. In south and southwest slope, habitat filtering was the driving force for community assembly. Interspecific competition was the main driving factor for community assembly in north and northwest slope aspects. But in west slope, two indices showed contrary consequence. This means the process of community assembly in west slope was more complicated and its phylogenetic index may be the result of several mechanisms working together.
Keywords:
-->0
PDF (1243KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(Rosindell et al., 2011)。群落构建理论已有近一个世纪的历史, 但我们对群落构建机理仍然不很清楚(牛克昌等, 2009)。生态位理论和中性理论是两个试图从不同的角度解释群落物种多样性, 从而揭示群落构建机制的基本理论(Hubbell, 2001)。近年来, 尽管已有大量的基于这两个理论来探究物种多样性的研究,但对局域群落构建机制的认识仍不清晰(柴永福和岳明, 2016)。现如今, 大多数生态学家致力于将这两个理论的要素结合起来构建综合模型(Gravel et al., 2006; 牛克昌等, 2009), 探究群落构建中随机和生态位过程。起初研究者常常采用物种多样性指标, 后来有些****提出采用种属比来反映群落物种的组成(Simberloff, 1970)。但这些方法都忽略了物种间进化历史对功能性状的影响, 仍不能准确地揭示群落构建的成因(牛红玉等, 2011)。
Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构。植物功能性状是植物为适应生存环境形成的植物形态和生理特征(孟婷婷等, 2007), 其差异反映了植物自身生理过程及其对外部环境异质性的适应策略, 而且能将群落结构与群落环境、生态系统过程等联系起来(孟婷婷等, 2007; 柴永福和岳明, 2016)。植物功能性状不同导致群落构建有所不同(Gerhold et al., 2013)。Webb曾在群落系统发育研究中, 将生态性状划分为保守性状和趋同性状(Webb et al., 2002; Kraft et al., 2007) (表1)。由于物种功能性状的种内和种间变异能更有效地描述物种间的相互关系, 越来越多的证据表明基于功能性状的研究方法有助于阐明生物多样性效应的潜在机制(刘晓娟和马克平, 2015)。Felsenstein (1985)提出, 除了外界环境, 物种进化是影响功能性状的重要因素, 亲缘关系近的物种间性状差异较小, 亲缘关系较远的物种间性状差异较大。因此, 在物种间性状的相关性研究中需考虑物种间的亲缘关系, 即检验物种的功能性状是否表现出系统发育信号。如杨洁等(2014)关于中山常绿阔叶林的研究, 选取的功能性状均具有显著的系统发育信号, 表明该森林植物受系统发育进化历史的影响。功能性状是植物某一个功能方面的表现, 而系统发育表现了种间总体的差异, 是多个性状的综合, 二者不能相互替代, 故在研究中应选择将两者结合的方式推断群落的构建过程。
Table 1
表1
表1不同生态性状和群落构建过程结合的群落期望系统发育结构(引自Webb et al., 2002; Kraft et al., 2007)
Table 1The excepted distribution of community phylogenetic, given various community assembly processes and different evolutionary characteristic of ecological traits after( Webb et al., 2002; Kraft et al., 2007)
| 群落构建过程 Community assembly processes | 生态性状的进化特征 Evolutionary characteristic of ecological trait | ||
|---|---|---|---|
| 性状保守 Traits conserved | 性状趋同 Traits convergent | ||
| 生态位理论 Neutral theory | 生境过滤 Habitat filtering | 系统发育聚集 Cluster dispersion | 系统发育发散 Over dispersion |
| 极限相似 Limiting similarity | 系统发育发散 Over dispersion | 系统发育聚集或随机 Clustered or random dispersion | |
| 中性理论 Neutral theory | 中性作用 Neutral assembly | 系统发育随机 Random dispersion | 系统发育随机 Random dispersion |
新窗口打开
甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区。但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓。随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注。与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化。这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等。植物的生长及群落结构又会反作用于土壤。因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义。近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(刘梦雪等, 2010)、物种多样性(董世魁等, 2017)、物种丰富度(Zhang et al., 2012)、功能多样性(王海东等, 2013)、群落生产力(Liu et al., 2015; 朱桂丽等, 2017)、环境因子对群落构建的影响, 如土壤环境、刈割、施肥(吕美强等, 2014)、营养元素的添加、人为干扰放牧(李红琴等, 2017)等方面。也有一些关于高寒草甸群落构建机制的研究, 但多选用功能性状或系统发育单一的指标(Zhang et al., 2015; 肖欣爽和王刚, 2015), 鲜有结合两者来揭示群落构建机制的研究。本文将结合植物的功能性状、环境因子和群落的系统发育结构, 从进化历史和生态过程的角度, 更深入地探讨甘南亚高寒草甸群落沿坡向梯度的构建机制。这将为了解群落发展的本质, 预测群落未来的演替方向, 为当地生态环境进行保护以及合理的开发利用提供理论依据。
1 材料和方法
1.1 试验样地概况
野外试验样地位于青藏高原东缘的兰州大学高寒草甸与湿地生态系统定位研究站附近(34.54° N, 102.49° E)。这里平均海拔2 900-3 000 m, 气候寒冷湿润。年平均气温2 ℃, 年降水量为557.8 mm, 降水集中于6-8月, 雨热同期。主要植被类型为亚高寒草甸, 土壤为亚高寒草甸土。1.2 样方设计和调查
2016年在植物生长的旺盛期7、8月进行野外取样。通过360°电子罗盘坡向定位, 依次将所选山坡分为5个坡向, 即南坡(0°)、西南坡(45°)、西坡(90°)、西北坡(135°)、北坡(180°)。在各坡向样地顺着山体垂直的方向设置两行样方, 间距3 m, 每行4个, 大小为50 cm × 50 cm, 每个样方间的距离是1 m。调查统计样方内植物的数量、高度、盖度等。选取各样地的优势物种, 测定其叶片性状。在每个样方内取0-20 cm深的土壤用于后续试验。1.3 研究方法
1.3.1 植物功能性状的测定本文选用4个与光照和土壤含水量相关的植物功能性状, 即比叶面积、叶干物质含量、叶片氮含量、叶片磷含量。其中比叶面积是衡量物种生长状况和光能利用效率的重要指标, 叶干物质含量主要反映植物营养元素的保持能力, 氮、磷元素在植物的新陈代谢中起着非常重要的作用, 对光合速率有重要影响。在所选样地取每个物种3-5片成熟展开的叶(取样物种覆盖度为90%-95%), 用扫描仪扫描后保存图像, 用软件ImageJ进行叶面积计算。比叶面积(SLA)的计算公式如下: SLA = Areal/Drymassl, 其中Areal为叶面积, Drymassl为叶片干质量。叶干物质含量(LDMC) =叶片干质量/叶片鲜质量。其余叶片烘干磨成细粉, 用凯氏法测定叶片全氮含量; 用钼锑抗比色法测定叶片全磷含量。每个样方中所测物种的功能性状都用相对多度来计算, 物种功能性状的群落加权平均值(CWM)的计算公式如下:
$CW{{M}_{i}}=\sum\limits_{i=1}^{n}{{{D}_{i}}}\times Trai{{t}_{i}}$ (1)
式中, CWMi表示所测物种i功能性状的群落加权平均值, Di表示物种i的相对多度, Traiti表示物种i的平均功能性状值。
1.3.2 土壤理化性质的测定
用烘干法测定土壤含水量, 并重复3次。用凯氏法测定土壤全氮含量; 用钼锑抗比色法测定土壤全磷含量; 用重铬酸钾容量法(外加热法)测定土壤有机碳含量。
1.4 数据计算和分析
1.4.1 群落系统发育以Webb (2000)的试验方法构建群落系统发育树。将样方中所有物种的科属种名称按所要求的格式输入phylomatic在线软件中, 会自动输出由其样方物种构建的系统发育树(Webb, 2000; Webb & Donoghue, 2005; Webb et al., 2008)。运用这个软件可对已输出的进化树拟合枝长, 生成一个矫正过分枝长度的谱系树。然后计算每个样地的Faith (1992)谱系多样性指数(PD)。分析物种在进化树中的位置,
计算物种间的谱系距离。假设物种分布随机, 在随机模型下将谱系距离标准化, 从而获得群落系统发育结构指数, 即净亲缘关系指数(NRI)和最近亲缘关系指数(NTI)。其中NRI是指群落中所有物种对的平均谱系距离(MPD), 而NTI是群落中所有物种的最近相邻谱系距离的平均值(MNTD)。群落系统发育结构指数的计算公式如下:
$NR{{I}_{\text{s}}}=-1\times \frac{MP{{D}_{\text{s}}}-MP{{D}_{\text{rnds}}}}{SD(MP{{D}_{\text{rnds}}})}$ (2)
$NT{{I}_{\text{s}}}=-1\times \frac{MNT{{D}_{\text{s}}}-MNT{{D}_{\text{rnds}}}}{SD(MP{{D}_{\text{rnds}}})}$ (3)
式中, NRIs和NTIs是样方群落系统发育结构指数, MPDs和MNTDs分别表示平均谱系距离的观测值和最近相邻谱系距离的观测平均值, 而MPDrnds和MNTDrnds表示在软件间中随机模拟出的平均谱系距离和最近相邻谱系距离的平均值。
Table 2
表2
表2研究区样地概况
Table 2The information of plots in study area
| 坡向 Aspect | 海拔 Altitude (m) | 纬度 Latitude (° N) | 经度 Longitude (° E) | 坡度 Gradient (°) | 坡向定位 Aspect orientation (°) |
|---|---|---|---|---|---|
| 南坡 South slope | 3 034 | 34.65 | 102.53 | 32 | 44 |
| 西南坡 Southwest slope | 3 032 | 34.65 | 102.53 | 27 | 88 |
| 西坡 West slope | 3 031 | 34.65 | 102.53 | 24 | 132 |
| 西北坡 Northwest slope | 3 038 | 34.65 | 102.53 | 20 | 176 |
| 北坡 North slope | 3 042 | 34.65 | 102.53 | 22 | 220 |
新窗口打开
为了衡量群落的谱系结构, 通常采用与零模型比较的方法。本文选用Phylocom软件中的2号模型。该模型中每一个样地中的种均为随机从谱系库中抽取, 保留了每个样地的物种丰富度, 但是每个样地出现的种被随机化。例如, 对于一块样地, 物种是从谱系库中不放回抽取。若NRIs和NTIs值大于零模型, 则表示该样方群落的系统发育结构聚集; 若NRIs和NTIs值小于零模型, 则表示该样方群落的系统发育结构发散; 若NRIs和NTIs值等于零模型, 则表示该样方群落的系统发育结构随机。Phylocom软件在计算NRIs和NTIs值时, 会默认将观测值与模拟值比较999次, 如果观测值大于或小于模拟值的次数达到975次以上, 就认为所测样方具有显著的系统发育结构, 即p < 0.05。最后运用student t检验判断每个坡向群落系统发育结构不显著的样方偏离零假设的显著性。
1.4.2 功能性状的系统发育信号检验
本文采用Blomberg等(2003)提出的K值法检验功能性状的系统发育信号强度。若K = 1, 则表明该功能性状表现出按布朗运动模型的方式进化; 若K < 1, 则表示功能性状表现出的系统发育信号比按布朗运动模型进化弱; 若K > 1, 则表示功能性状表现出的系统发育信号比按布朗运动模型方式进化强。功能性状系统发育信号的显著性采用与零模型比较的方式来衡量, 若是实际值大于零模型值的次数达到950次以上, 就认为功能性状表现出显著的系统发育信号(p < 0.05)。
2 结果和分析
2.1 不同坡向环境因子的变化
由图1A可知, 土壤含水量在各个坡向间差异显著。从南坡到西北坡, 土壤含水量依次上升, 在西北坡达到最大值(37.92%), 随后又开始递减, 其中, 南坡的土壤含水量最低(17.74%)。土壤有机碳和土壤全氮含量在各坡向上差异显著, 且具有相同的变化趋势, 西南坡的含量最低(土壤有机碳含量62.13 g·kg-1、土壤全氮含量3.85 g·kg-1), 北坡的含量最高(土壤有机碳含量101.63 g·kg-1、土壤全氮含量5.03 g·kg-1), 从西南坡到北坡土壤有机碳和土壤全氮含量呈上升趋势(图1B、1D)。从南坡到西坡土壤全磷含量先升后降(图1C), 到西坡值最低(0.4 g·kg-1), 随后呈增长趋势, 北坡含量最高, 为0.68 g·kg-1。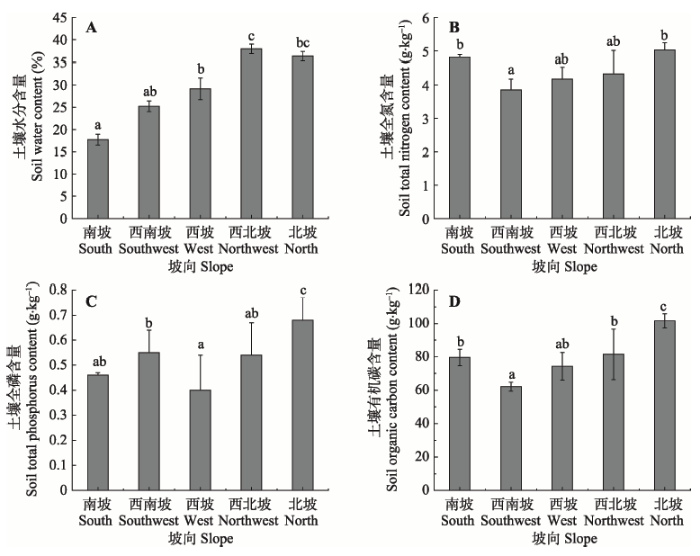 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1不同坡向土壤环境因子的变化(平均值±标准偏差)。不同小写字母表示坡向间差异显著(p < 0.05)。
-->Fig. 1Variation on soil environmental factors in different slope aspect (mean ± SD). Different lowercase letters indicate significant difference among different slope aspects (p < 0.05).
-->
2.2 植物群落组成
本研究共调查到21科48属60种物种, 其中南坡21种, 西南坡32种, 西坡35种, 西北坡39种, 北坡41种。南坡优势种有矮生嵩草(Kobresia humilis)和少花米口袋(Gueldenstaedtia verna), 西南坡优势种为矮生嵩草, 西坡优势种有圆穗蓼(Polygonum macrophyllum), 西北坡和北坡的优势种为金露梅(Potentilla fruticosa)。矮生嵩草、蒲公英(Taraxacum mongolicum)、少花米口袋和狼毒(Stellera chamaejasme)为5个坡向的共有种。2.3 植物的功能性状沿坡向梯度的变化
由图2可知, 在不同的坡向上, 植物的各功能性状差异显著。其中叶干物质含量在南坡、西南坡最高, 西北坡含量最低。SLA、叶片氮、磷含量沿坡向变化趋势相同, 从南坡到北坡, 其值呈先增大后减小的趋势, 在西北坡达到最大。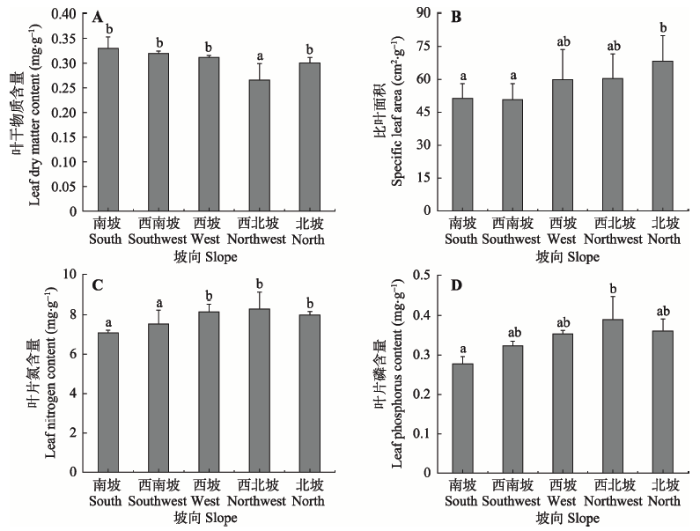 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2植物功能性状沿坡向变化(平均值±标准偏差)。不同小写字母表示坡向间差异显著(p < 0.05)。
-->Fig. 2Variation of plant functional traits in different slope aspects (mean ± SD). Different lowercase letters indicate significant difference among different slope aspects (p < 0.05).
-->
2.4 系统发育信号
由表3可知, 植物叶干物质含量、SLA、叶片氮含量、叶片磷含量等4个功能性状的K值均小于1, 说明功能性状表现出的系统发育信号强度比按布朗运动模型进化弱, 这4种功能性状都没有较强的系统发育保守性。叶片磷含量表现出微弱的系统发育信号, p < 0.05, 而其他所选功能性状均未检测到明显的系统发育信号(p > 0.05), 表明4种功能性状受物种的进化历史影响不大。Table 3
表3
表3甘南藏族自治州亚高寒草甸植物功能性状的系统发育信号
Table 3The phylogenetic signal of plants functional traits in subalpine meadow communities in Gannan Tibetan Autonomous Prefecture
| 功能性状 Functional traits | 物种数 Species No. | K值 K value | p |
|---|---|---|---|
| 叶干物质含量 Leaf dry matter content (mg·g-1) | 60 | 0.378 | 0.259 |
| 比叶面积 Specific leaf area (cm2·g-1) | 60 | 0.352 | 0.167 |
| 叶片氮含量 Leaf nitrogen content (mg·g-1) | 60 | 0.412 | 0.135 |
| 叶片磷含量 Leaf phosphorus content (mg·g-1) | 60 | 0.235 | 0.040 |
新窗口打开
2.5 群落系统发育结构沿坡向的变化
基于APGIII数据和60个物种构建的系统发育树如图3所示, 各分支长度代表物种的进化历史。由图4可知, 从南坡到北坡, 各样方生境内的物种数逐渐增多, 分别为21、32、35、39、41种。谱系多样性指数PD也随坡向变化逐渐增大。对谱系多样性指数PD与样方内的物种数进行相关性分析, 发现两者极显著正相关。系统发育指数NRI和NTI沿坡向有相同的变化趋势。从南坡到北坡, 样方群落的系统发育发散程度逐渐降低。西坡样方群落的系统发育指数结果表现不一致, 其中NRI表现出小于零假设的趋势, 代表西坡样方群落的系统发育结构发散, 而NTI表现出大于零假设的趋势, 代表系统发育结构聚集。西北坡与北坡的系统发育指数相同, 均大于零假设期望值, 系统发育结构聚集。而南坡、西南坡的系统发育指数均小于零假设期望值, 系统发育结构发散。对于各坡向样方群落系统发育结构不显著的群落进行t检验, 发现均未表现出显著偏离零假设的趋势。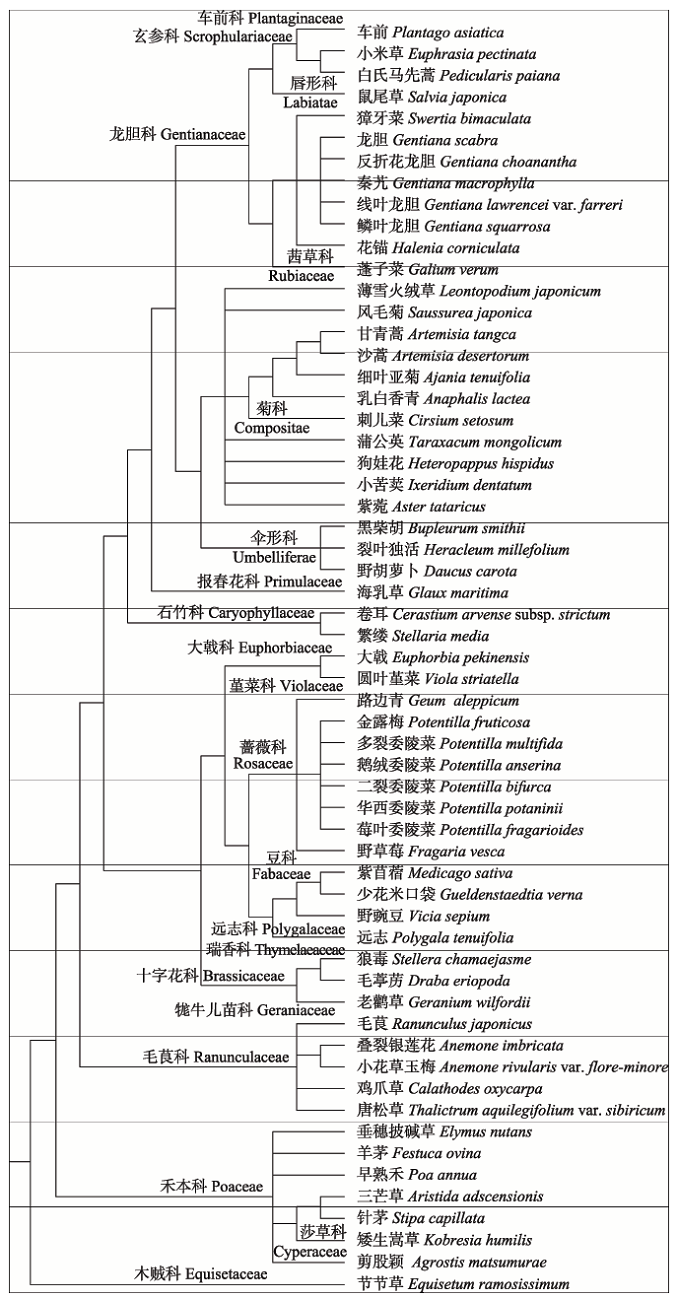 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3甘南藏族自治州亚高寒草甸基于坡向样地60个物种构建的群落系统发育树。
-->Fig. 3A super-tree representing the 60 species based on slope-oriented plots in subalpine meadow communities in Gannan Tibetan Autonomous Prefecture.
-->
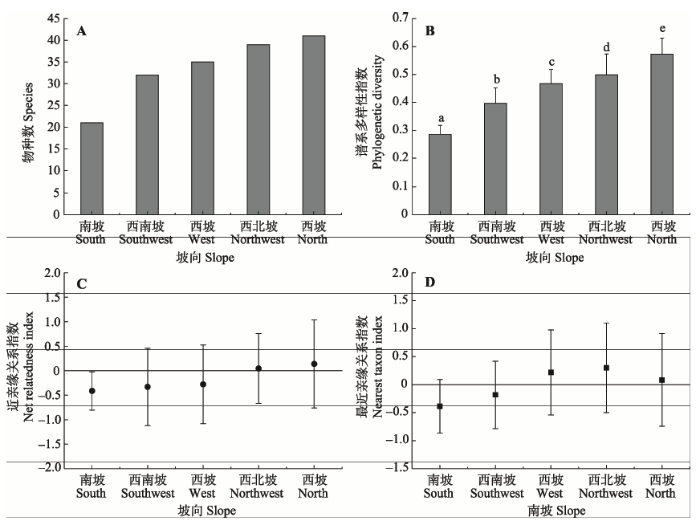 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4不同坡向生境内的物种数、谱系多样性指数及系统发育指数(平均值±标准偏差)。不同小写字母表示坡向间差异显著 (p < 0.05)。
-->Fig. 4Variation of species, phylogenetic diversity and community phylogenetic relatedness along slope aspect (mean ± SD). Different lowercase letters indicate significant difference among different slope aspects (p < 0.05).
-->
3 讨论和结论
环境因子和光照是影响亚高寒草甸坡向生境的主要环境条件。坡向的变化对土壤含水量、土壤养分(碳、氮、磷)等都有显著影响(图1)。土壤含水量从南坡到北坡先增加后减少, 在西北坡达到最大值, 这与宫骁和王刚(2015)、刘旻霞和马建祖(2012)指出的土壤含水量随着坡度的变化从南坡到北坡呈上升的趋势有所不同。这可能是因为西北坡光照较弱且光照时间短, 水分蒸发量较小, 且生长有金露梅等大量的灌木, 具有一定的保水性, 水分不易蒸发, 因而土壤含水量相对较多。北坡的土壤养分含量最高, 这与Gong等(2008)的研究结论一致。西南坡和西坡的碳、氮含量较低, 磷含量在南坡和西坡较低。这可能是因为西南坡和西坡为南北坡的生境过渡带, 受外界干扰大, 风蚀严重, 从而导致土壤养分流失严重。如图2可知, 大部分植物的叶片功能性状在不同坡向间差异显著, 其中叶干物质含量在南坡、西南坡较高, SLA和叶片氮、磷含量在北坡、西北坡较高。这是由于SLA是衡量物种的生长状况和光能利用效率的重要指标, 在阳坡, 因为土壤含水量较低, 温度和光照度较高, 蒸发量大, 养分含量低, 所以物种生长速率慢, SLA较小, 且叶片营养元素较低。土壤含水量与SLA以及叶片N含量在坡向上均有相同的变化趋势, 与叶干物质含量的变化趋势相反(图1, 图2)。从南坡到北坡各样方生境内的物种数逐渐增多(图4), 这与聂莹莹等(2010)的研究结果一致, 即物种的多样性与光照和土壤温度负相关, 与土壤含水量正相关。综上所述, 土壤含水量是坡向多种环境因子中最主要的限制因子, 这与刘旻霞等(2013)和卢孟孟等(2014)的研究结果一致。本文选用叶干物质含量、SLA、叶片氮含量、叶片磷含量4种功能性状, 来检验甘南藏族自治州亚高寒草甸植物群落的功能性状的系统发育信号。结果表明: 只有植物叶片磷含量表现出微弱的系统发育信号, 其余功能性状表现出的系统发育信号强度比按布朗运动模型进化弱, 都没有较强的系统发育保守性, 系统发育信号都不显著, 表明4种功能性状受物种进化历史的影响不大(表3)。Liu等(2015)对亚高寒草甸的研究表明, 只有部分功能性状, 如叶片磷含量、植物高度有微弱的系统发育信号, 叶片氮含量、SLA、平均种子质量等均未检测到显著的系统发育信号, 我们的研究结果与之较一致。宫骁和王刚(2015)在坡向梯度研究中发现SLA和稳定碳同位素都没有显著的系统发育信号。Ding等(2012)和Swenson (2013)的研究表明并不是所有的功能性状都会表现出显著的系统发育信号。而在沿海拔梯度对植物群落的研究中, 试验所选功能性状均具有显著的系统发育信号(曹科等, 2013; 房帅等, 2014; 杨洁等, 2014)。因此我们推测, 在坡向这样的微生境梯度上, 植物功能性状受物种进化历史的影响不是很大, 没有表现出明显的系统发育信号。
本研究结果表明, 各坡向物种数与系统发育多样性指数有相同的变化趋势, 即从南坡到北坡, 物种数增多, 系统发育指数也增大(图4A, 4B), 这与Honorio Coronado等(2015)的研究结果一致。由图4可知, 南坡的植物群落的系统发育结构聚集,西南坡的植物群落系统发育结构发散, 且从西南坡到北坡发散程度逐渐降低。在西坡, 两种群落的系统发育指数并不一致, NRI表现出西坡群落系统发育聚集, NTI则表现出系统发育发散。宫骁和王刚(2015)在分析沿坡向梯度植物群落的系统发育结构时发现西坡和西北坡的两种系统发育指数也不一致。西坡两种系统发育指数结果不同, 可能有两种解释: 可能是由于西坡属于南北坡的坡向过渡带, 环境条件和种间竞争共同影响了该坡群落构建过程; 也可能是由于两种系统发育指数的侧重点不同, NRI亲缘关系是基于群落整体水平的, 反映的是整个系统发育树的发育模式, 而NTI亲缘关系是基于系统进化树末端的最近的系统发育关系(Kraft et al., 2007)。
综合环境因子、功能性状和系统发育结果(图1- 4; 表3), 我们可以看到, 植物叶片功能性状随坡向呈现出有规律的变化, 系统发育结构随着坡向梯度的变化表现出发散程度逐渐降低的规律。从表3可知, 本研究中所选功能性状的K值均小于1, 故功能性状表现出的系统发育信号强度比按布朗运动模型进化弱, 都没有较强的系统发育保守性, 因此甘南藏族自治州高寒草甸植物群落的生态性状趋同。从系统发育角度看, 南坡和西南坡植物群落系统发育结构发散, 故该坡向植物群落的构建过程主要是生境过滤。从功能性状角度看, 南坡和西南坡土壤含水量低, 土壤养分含量较少, 且光照强, 生存环境恶劣不利于植物生长, 生境内的物种类型少, 植物叶片的SLA、叶片氮、磷含量较低, 表明物种生长状况受环境影响较大, 这与肖欣爽和王刚(2015)的研究结果一致, 即环境过滤是阳坡群落构建的主要过程。西北坡和北坡NRI和NTI指数都表现出了聚集的倾向, 表明极限相似性是西北坡和北坡植物群落构建的主驱动力。与南坡和西南坡相比, 西北坡和北坡土壤含水量和土壤养分含量高, 适宜大多数植物生存, 环境对植物共存方式的影响逐渐减弱, 生物间的相互作用愈加增强。西坡群落的系统发育指数不一致, 表现出随机的趋势, 但这不一定是中性作用导致的。因为除了生境过滤和竞争排斥, 其他的生态过程也会影响群落的物种多样性格局, 如生物间的捕食、互利共生、促进等作用, 这可能是由几种驱动机制共同作用的结果(牛红玉等, 2011)。
本研究结合植物功能性状和群落谱系结构揭示了亚高寒草甸群落物种的共存方式, 得出以下结论: 物种的多样性与光照和土壤温度呈负相关关系, 与土壤含水量呈正相关关系, 物种多样性与系统发育多样性往往有着显著的正相关关系。生境过滤是南坡和西南坡植物种共存的主要机制; 北坡和西北坡生境过滤作用逐渐减弱, 物种共存的主要机制是极限相似性; 而西坡的结果表明, 过渡区的群落构建过程比较复杂, 需做进一步的研究。利用群落系统发育结合植物功能性状探讨群落的构建过程, 对于了解物种共存及维持生物多样性具有重要意义。但本文仅从植物功能性状和系统发育方面来探讨群落的构建机制, 对于西坡这种生境过渡区域, 并不能准确地揭示其群落构建过程。在以后的研究中, 应结合其他的生态过程, 从多维度分析植物群落的构建机制。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . Abstract The primary rationale for the use of phylogenetically based statistical methods is that phylogenetic signal, the tendency for related species to resemble each other, is ubiquitous. Whether this assertion is true for a given trait in a given lineage is an empirical question, but general tools for detecting and quantifying phylogenetic signal are inadequately developed. We present new methods for continuous-valued characters that can be implemented with either phylogenetically independent contrasts or generalized least-squares models. First, a simple randomization procedure allows one to test the null hypothesis of no pattern of similarity among relatives. The test demonstrates correct Type I error rate at a nominal alpha = 0.05 and good power (0.8) for simulated datasets with 20 or more species. Second, we derive a descriptive statistic, K, which allows valid comparisons of the amount of phylogenetic signal across traits and trees. Third, we provide two biologically motivated branch-length transformations, one based on the Ornstein-Uhlenbeck (OU) model of stabilizing selection, the other based on a new model in which character evolution can accelerate or decelerate (ACDC) in rate (e.g., as may occur during or after an adaptive radiation). Maximum likelihood estimation of the OU (d) and ACDC (g) parameters can serve as tests for phylogenetic signal because an estimate of d or g near zero implies that a phylogeny with little hierarchical structure (a star) offers a good fit to the data. Transformations that improve the fit of a tree to comparative data will increase power to detect phylogenetic signal and may also be preferable for further comparative analyses, such as of correlated character evolution. Application of the methods to data from the literature revealed that, for trees with 20 or more species, 92% of traits exhibited significant phylogenetic signal (randomization test), including behavioral and ecological ones that are thought to be relatively evolutionarily malleable (e.g., highly adaptive) and/or subject to relatively strong environmental (nongenetic) effects or high levels of measurement error. Irrespective of sample size, most traits (but not body size, on average) showed less signal than expected given the topology, branch lengths, and a Brownian motion model of evolution (i.e., K was less than one), which may be attributed to adaptation and/or measurement error in the broad sense (including errors in estimates of phenotypes, branch lengths, and topology). Analysis of variance of log K for all 121 traits (from 35 trees) indicated that behavioral traits exhibit lower signal than body size, morphological, life-history, or physiological traits. In addition, physiological traits (corrected for body size) showed less signal than did body size itself. For trees with 20 or more species, the estimated OU (25% of traits) and/or ACDC (40%) transformation parameter differed significantly from both zero and unity, indicating that a hierarchical tree with less (or occasionally more) structure than the original better fit the data and so could be preferred for comparative analyses. |
| [2] | . . 植物功能性状(plant functional trait)的进化历史及其与群落结构的关系是近年来生态学研究的热点问题之一.为探究植物功能性状受系统进化历史的影响程度,以及功能性状是否通过影响物种多度分布进而影响群落结构等问题,本研究以古田山24 ha永久性监测样地中的156种木本植物为对象,收集了叶片氮含量、叶片磷含量、叶面积、木质密度、比叶面积和种子重量6种功能性状的数据,用K值法检验了各个功能性状的系统发育信号(phylogenetic signal),并运用系统发育独立比较(phylogenetic independent contrasts,PIC)的方法分析了各个性状与多度的关系.结果显示,所有6种功能性状都表现出较强的系统发育信号,表明本区域木本植物系统发育历史显著地影响这些功能性状;另一方面,叶氮含量、叶磷含量、叶面积、木质密度和比叶面积与物种多度显著相关,而种子重量与物种多度没有显著的相关性,表明群落内不同物种的资源获取方式可显著地影响群落的结构. |
| [3] | . . 群落构建研究对于解释物种共存和物种多样性的维持是至关重要的,因此一直是生态学研究的中心论题。尽管近年来关于生态位和中性理论的验证研究已经取得了显著的成果,但对于局域群落构建机制的认识仍存在很大争议。随着统计和理论上的进步使得用功能性状和群落谱系结构解释群落构建机制变为可能,主要是通过验证共存物种的性状和谱系距离分布模式来实现。然而,谱系和功能性状不能相互替代,多种生物和非生物因子同时控制着群落构建,基于中性理论的扩散限制、基于生态位的环境过滤和竞争排斥等多个过程可能同时影响着群落的构建。所以,综合考虑多种方法和影响因素探讨植物群落的构建机制,对于预测和解释植被对干扰的响应,理解生物多样性维持机制有重要意义。试图在简要回顾群落构建理论及研究方法发展的基础上,梳理其最新研究进展,并探讨整合功能性状及群落谱系结构的研究方法,解释群落构建和物种多样性维持机制的可能途径。在结合功能性状和谱系结构研究群落构建时,除了考虑空间尺度、环境因子、植被类型外,还应该关注时间尺度、选择性状的种类和数量、性状的种内变异、以及人为干扰等因素对群落构建的影响。 |
| [4] | . Abstract A rapidly increasing effort to merge functional community ecology and phylogenetic biology has increased our understanding of community assembly. However, studies using both phylogenetic- and trait-based methods have been mainly conducted in old-growth forests, with fewer studies in human-disturbed communities, which play an increasingly important role in providing ecosystem services as primary forests are degraded. We used data from 18 1-ha plots in tropical old-growth forests and secondary forests with different disturbance histories (logging and shifting cultivation) and vegetation types (tropical lowland and montane forests) on Hainan Island, southern China. The distributions of 11 functional traits were compared among these six forest types. We used a null model approach to assess the effects of disturbance regimes on variation in response and effect traits and community phylogenetic structure across different stem sizes (saplings, treelets, and adult trees) and spatial scales (10 50 m). We found significant differences in the distribution of functional traits in highly disturbed lowland sites versus other forest types. Many individuals in highly disturbed lowland sites were deciduous, spiny, with non-fleshy fruits and seeds dispersed passively or by wind, and low SLA. The response traits of coexisting species were clustered in all sites except for highly disturbed lowland sites where evenness was evident. There were different distributions of effect traits for saplings and treelets among different forest types but adult trees showed stronger clustering of trait values with increasing spatial scale among all forest types. Phylogenetic clustering predominated across all size classes and spatial scales in the highly disturbed lowland sites, and evenness in other forest types. High disturbance can lead to abiotic filtering, generating a community dominated by closely related species with disturbance-adapted traits, where biotic interactions play a relatively minor role. In lightly disturbed and old growth forests, multiple processes simultaneously drive the community assembly, but biotic processes dominate at the fine scale. |
| [5] | . . 物种多样性与功能多样性的关系是生态学当前研究的热点问题之一,不同区域典型生态系统物种多样性和功能多样性的关系研究有利于生物多样性保护理论的全面发展。以青藏高原地区的主要草地生态系统-高寒草甸和高寒草原为研究对象,采用4个物种多样性指数(Patrick丰富度指数、Shannon-Weiner多样性指数、Pielou均匀度指数和Simpson优势度指数)和9个功能多样性指数(FAD功能性状距离指数、MFAD功能性状平均距离指数、基于样地的FDp和基于群落的FDc功能树状图指数、FRic功能体积指数、FEve功能均匀度指数、Rao功能离散度常二次熵指数、FDiv功能离散指数、FDis功能分散指数),分析了高寒草地植物物种多样性、功能多样性关系及其与初级生产力的关系,以期阐明3个科学问题:不同草地类型的高寒草地生态系统植物物种多样性和功能多样性有何差异?高寒草地生态系统的植物物种多样性和功能多样性有何关系?高寒草地生态系统物种多样性、功能多样性对生态系统功能的影响有何异同?研究结果表明:(1)与高寒草原相比,高寒草甸具有更高的物种多样性、功能丰富度和功能离散度;(2)高寒草甸中,Patrick丰富度与功能丰富度指数(FAD、MFAD、FDp、FDc)和功能离散度指数(FDiv)的具有较强的相关性,最优拟合方程分别为幂函数和二次多项式函数;(3)高寒草原中,Patrick丰富度与功能丰富度指数(FAD、MFAD、FDp、FDc、FRic)、Shannon指数和Simpson指数与FEve指数的相关性较强,最优拟合方程为二次多项式函数,Pielou指数与FEve指数的相关性较强,最优拟合方程为指数函数;(4)高寒草甸的初级生产力分别与物种丰富度指数Patrick、功能离散指数FDiv具有较强的相关性;而高寒草原的初级生产力与4个物种多样性指数间均具有较强的相关性,与17 |
| [6] | . |
| [7] | . Protecting biological diversity with limited resources may require placing conservation priorities on different taxa. A system of priorities that reflects the value of taxonomic diversity can be achieved by setting priorities such that the subset of taxa that is protected has maximum underlying feature diversity. Such feature diversity of taxon subsets is difficult to estimate directly, but can be predicted by the cladistic/phylogenetic relationships among the taxa. In this study, a simple measure of phylogenetic diversity is defined based on cladistic information. The measure of phylogenetic diversity, PD, is contrasted with a measure of taxic diversity recently developed by Vane-Wright et al. (Biol. Conserv., 55, 1991). In re-examining reserve-selection scenarios based on a phylogeny of bumble bees (Apidae), PD produces quite different priorities for species conservation, relative to taxic diversity. The potential application of PD at levels below that of the species is then illustrated using a mtDNA phylogeny for populations of crested newts Triturus cristatus. Calculation of PD for different population subsets shows that protection of populations at either of two extremes of the geographic range of the group can significantly increase the phylogenetic diversity that is protected. |
| [8] | . |
| [9] | . ABSTRACT Question: There are contrasting opinions about how communities assemble along a productivity gradient, particularly in relation to competitive interactions. One view is that functionally similar, and closely related species, are less likely to co-exist (limiting similarity). Alternatively, competitive exclusion may act on dissimilar species because species bearing traits associated with low competitive ability are excluded (weaker competitor exclusion). We ask if patterns of functional and phylogenetic diversity are related to changes in species diversity in response to fertility manipulations. Location: Species-rich mesophytic grassland in Estonia. Methods: The grassland has been manipulated from 2002 to 2011 to increase (fertilizer addition) and decrease productivity (sucrose addition) in small-scale (50 cm 50 cm) plots. We linked annual increases and decreases in species diversity to changes in functional and phylogenetic diversity. We used abundance-weighted mean pair-wise functional or phylogenetic distance of all possible species pairs. Results: We found convergence in four traits (plant height, leaf distribution, lateral spread, type of reproduction) and a decrease in mean functional and phylogenetic diversity, in support of weaker competitor exclusion or habitat filtering. There was less support for limiting similarity, with divergence found for two traits associated with decreasing species diversity (leaf distribution in the sucrose treatment and lateral spread in the fertilizer treatment). Conclusions: Our results support the view that competition can lead to the exclusion of weaker competitors, rather than increasing functional and phylogenetic diversity, as expected from the principle of limiting similarity. However, multiple assembly processes, which are generally seen as mutually exclusive, are operating simultaneously, albeit on different traits and at different stages of community assembly. |
| [10] | . In order to investigate the productivity of hilly grassland of the Xilin River Basin as affected by slope aspect (North versus South), aboveground green biomass (AGB) and species composition were studied at four hill sites in 2005. Additionally, a detailed investigation combining vegetation parameters, physical and chemical soil characteristics was carried out at different slope positions (base, middle, and top) of north and south slope aspect on one of the hills in 2004 and 2005. Northern slopes exhibited a higher productivity and species diversity compared to southern slopes. Plant available soil K, P and N were not limiting the productivity at south- relative to north-oriented slopes, whereas soil available water was the primarily limiting factor for plant productivity, which was supported by results of leaf carbon isotope discrimination of main species. A negative correlation between productivity and relative abundance of C 4 plants indicates that the relative biomass of C 4 plants can be used as a water-stress indicator. |
| [11] | |
| [12] | . |
| [13] | . Abstract Aim To examine variation in the phylogenetic diversity (PD) of tree communities across geographical and environmental gradients in Amazonia. Location Two hundred and eighty-three c . 1 ha forest inventory plots from across Amazonia. Methods We evaluated PD as the total phylogenetic branch length across species in each plot (PDss), the mean pairwise phylogenetic distance between species (MPD), the mean nearest taxon distance (MNTD) and their equivalents standardized for species richness (ses.PDss, ses.MPD, ses.MNTD). We compared PD of tree communities growing (1) on substrates of varying geological age; and (2) in environments with varying ecophysiological barriers to growth and survival. Results PDss is strongly positively correlated with species richness (SR), whereas MNTD has a negative correlation. Communities on geologically young- and intermediate-aged substrates (western and central Amazonia respectively) have the highest SR, and therefore the highest PDss and the lowest MNTD. We find that the youngest and oldest substrates (the latter on the Brazilian and Guiana Shields) have the highest ses.PDss and ses.MNTD. MPD and ses.MPD are strongly correlated with how evenly taxa are distributed among the three principal angiosperm clades and are both highest in western Amazonia. Meanwhile, seasonally dry tropical forest (SDTF) and forests on white sands have low PD, as evaluated by any metric. Main conclusions High ses.PDss and ses.MNTD reflect greater lineage diversity in communities. We suggest that high ses.PDss and ses.MNTD in western Amazonia results from its favourable, easy-to-colonize environment, whereas high values in the Brazilian and Guianan Shields may be due to accumulation of lineages over a longer period of time. White-sand forests and SDTF are dominated by close relatives from fewer lineages, perhaps reflecting ecophysiological barriers that are difficult to surmount evolutionarily. Because MPD and ses.MPD do not reflect lineage diversity per se , we suggest that PDss, ses.PDss and ses.MNTD may be the most useful diversity metrics for setting large-scale conservation priorities. |
| [14] | |
| [15] | . Abstract Taxa co-occurring in communities often represent a nonrandom sample, in phenotypic or phylogenetic terms, of the regional species pool. While heuristic arguments have identified processes that create community phylogenetic patterns, further progress hinges on a more comprehensive understanding of the interactions between underlying ecological and evolutionary processes. We created a simulation framework to model trait evolution, assemble communities (via competition, habitat filtering, or neutral assembly), and test the phylogenetic pattern of the resulting communities. We found that phylogenetic community structure is greatest when traits are highly conserved and when multiple traits influence species membership in communities. Habitat filtering produces stronger phylogenetic structure when taxa with derived (as opposed to ancestral) traits are favored in the community. Nearest-relative tests have greater power to detect patterns due to competition, while total community relatedness tests perform better with habitat filtering. The size of the local community relative to the regional pool strongly influences statistical power; in general, power increases with larger pool sizes for communities created by filtering but decreases for communities created by competition. Our results deepen our understanding of processes that contribute to phylogenetic community structure and provide guidance for the design and interpretation of empirical research. |
| [16] | . . 植被生态化学计量特征是高寒草甸生态系统稳定性的基础,但其对放牧管理的响应仍不清楚.为此,以2011年在青海海北高寒矮嵩草(Kobresia humilis)草甸建立的不同放牧强度试验地为研究对象,于2014年植物生长季的5月-9月,通过测定禁牧(对照,CK)、轻度放牧(light grazing,LG)、中度放牧(moderate grazing,MG)和重度放牧(heavygrazing,HG)样地的地上植被生产力和植被群落结构及群落叶片碳(carbon,C)、氮(nitrogen,N)、磷(phosphorus,P)含量,研究放牧强度对高寒草甸群落生态化学计量的影响.结果表明,MG能显著提高地上生物量和优良牧草禾草类植物的生物量比例,C含量的变异均在MG处理下最大,此结果支持了中度干扰理论;重牧能显著提高群落叶片的N含量(P<0.05);放牧管理对叶片P含量的影响无明显规律;禁牧有利于系统N和P的周转.本研究结果显示,适度的放牧强度有利于高寒草甸生产力的提高,而禁牧可能通过改变群落优势种增强系统的N、P循环强度. |
| [17] | . Abstract Many community experiments have shown a positive relationship between plant biodiversity and community productivity, with biodiversity measured in multiple ways based on taxonomy, function, and phylogeny. Whether these different measures of biodiversity and their interactions explain variation in productivity in natural assemblages has rarely been tested. In a removal experiment using natural alpine assemblages in the Tibetan Plateau, we manipulated species richness and functional diversity to examine how different measures of biodiversity predict aboveground biomass production. We combined different biodiversity measures (functional, phylogenetic, richness, evenness) in generalized linear models to determine which combinations provided the most parsimonious explanations of variation in biomass production. Although multivariate functional diversity indices alone consistently explained more variation in productivity than other single measures, phylogenetic diversity and plant height represented the most parsimonious combination. In natural assemblages, single metrics alone cannot fully explain ecosystem function. Instead, a combination of phylogenetic diversity and traits with weak or no phylogenetic signal is required to explain the effects of biodiversity loss on ecosystem function. |
| [18] | . . |
| [19] | . . 研究甘南高寒草甸不同坡向植物功能性状和土壤因子及二者的关系.结果表明:禾草是阳坡的优势功能群,杂草和灌木是阴坡的优势功能群.阴坡植物群落生物量显著大于阳坡,但叶片氮磷比显著小于阳坡.不同坡向上,除叶片氮含量变化不显著外,叶片磷含量、比叶面积和物种平均高度大小均为阴坡偏阴坡阳坡.植被的生存策略在阴阳坡发生了很大变化,这种变化可以从不同坡向植物叶性状的差异和优势功能群的改变反映出来.土壤温度大小为阳坡偏阴坡阴坡;土壤含水量大小为阴坡偏阴坡阳坡;除了全磷含量自阳坡到阴坡出现递增趋势外,土壤全氮、有机质和速效氮的大小均为阴坡阳坡偏阴坡.比叶面积、物种平均高度与土壤全磷、有机质和土壤含水量均呈显著正相关,与土壤pH呈显著负相关.土壤水分和土壤pH共同影响着高寒草甸不同坡向植物功能性状和土壤养分的分布. |
| [20] | . . 以甘南高寒草甸为对象,研究了坡向梯度上环境因子的变化特点及其与地上生物量和物种丰富度的关系.结果表明:0~20 cm与20~40 cm的土壤含水量均是自阳坡到阴坡呈递增的变化趋势,且20~40 cm土壤的含水量高于0~20 cm的;两层土壤的全磷质量分数和w(C):w(N)与含水量的变化相同,土壤全氮、有机质、速效磷及速效氮质量分数的变化均呈先减小后增大的趋势,偏阳坡最小.土壤养分质量分数均是0~20 cm的大于20~40 cm的,表现出表层土壤养分富集的现象,土壤pH值沿阳坡到阴坡呈递减趋势.地上生物量和物种丰富度的变化特点均为阴坡偏阴坡偏阳坡阳坡,且地上生物量与物种丰富度显著正相关.同归分析表明:土壤含水量与土壤全磷,有机质质量分数是影响地上生物量和物种丰富度的主要因子,其中土壤含水量是关键限制性因子. |
| [21] | . . |
| [22] | . . |
| [23] | . . 针对目前关于植物群落功能性状构建模式及其驱动因素存在的争议,研究了33个物种10个功能性状的构建模式。研究结果显示:(1)在刈割一施肥复合梯度上,这些功能性状主要表现为随机构建模式,发生随机性和确定性构建的样方比例分别为82.7%和17.3%;(2)在10个功能性状中,生长型、生活周期、单株地上干质量、叶面积和叶干质量5个功能性状为随机构建模式,不受试验处理和群落特征(地上净初级生产力、刈割生物量损失、群落高度)变化的影响。植物倾斜度、繁殖方式、固氮性、株高和比叶面积5个功能性状的构建模式与试验处理或群落特征变化有关,表现出趋同或趋异构建响应。其中,植物倾斜度和比叶面积的构建模式仅受群落特征影响,而固氮性、繁殖方式和株高3个性状的构建模式同时还受刈割或施肥处理的影响,其构建模式因功能性状而异;(3)群落的生物量损失和地上净初级生产力是解释植物功能性状构建模式变化较理想的群落特征;(4)刈割和施肥处理对株高的构建模式具有相反效应,而刈割和施肥的交互作用对其无显著影响。上述结果说明该研究群落植物功能性状的构建存在不同模式,以随机构建模式为主,确定性构建模式居次要地位。确定性构建模式与试验处理和群落特征变化有关,而且是性状依赖的。相反选择力对趋同和趋异构建模式的平衡效应能引起功能性状发生随机构建。 |
| [24] | . . 植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、过程与功能联系起来(所谓 的"植物功能性状").该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地貌、生态 梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO2 浓度升高)对个体和群落植物功能性状的影响.植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态 保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国的植物功能性状研究还需要更加明朗化和系 统化. |
| [25] | . . 采用野外调查与室内实验相结合的方法,研究了甘南地区青藏高原高寒草甸不同坡向上的植物群落物种多样性变化特点及与环境因子的关系.结果表明:阳坡-阴坡生境梯度上,光照、土温和含水量总的变化趋势为阳坡阳坡,阳坡和阴坡稍高,半阳坡最低的趋势.阳坡-阴坡生境梯度上植物群落物种的α多样性的变化趋势为:阳坡<半阳坡<阴坡;β多样性从阳坡,半阳坡到阴坡为一单峰曲线,即呈现先升高后降低的变化趋势,这表明在土壤湿度和日照时间适中时,物种替代速率比较快.不同坡向上的植物群落α多样性和β多样性与光照和土温均呈负相关,与0~15,15~30cm土层的土壤含水量和土壤全磷质量分数均呈显著正相关关系.通过主成分分析可以得出:引起α多样性和β多样性沿生境梯度变化的最重要因子是土壤水分和全磷质量分数. |
| [26] | . . 群落如何构建是群落生态学中的重要问题。群落谱系结构研究将物种间的亲缘进化关系运用到群落生态学研究中,利用物种的系统发育状况推测历史因素对现有群落的影响,为推断影响群落组成的生态学机制提供了有效方法。群落谱系结构的研究方法是首先建立可代表群落物种库的超级系统进化树,然后计算群落内物种间的谱系距离,最后通过统计方法检测其与随机模型下的谱系距离是否有显著差异来获得谱系结构(如谱系聚集、谱系发散),从而揭示群落构建中的关键生态过程(如生境过滤、竞争作用)。群落谱系结构与空间尺度、分类群尺度、时间尺度等不同研究尺度有关。在小的空间尺度下,随着分类群尺度降低、树木年龄级增大,群落谱系结构从聚集逐渐转为发散;而随群落空间尺度的增大,谱系趋向于聚集。谱系结构受到环境因素影响,因此分析集合群落下的谱系可以揭示区域生态过程的影响。另外,群落谱系结构研究还有助于探讨中性理论、密度制约假说等生态学理论,并预测干扰作用下的群落演化趋势。在利用谱系结构深入探讨群落构建成因时,需要基于生态特征和环境变量共同分析,同时考虑小尺度局域过程(群落的微环境或群落内种间相互作用等)和大尺度区域过程(地史过程和物种形成等),并可结合生态控制实验,以确认群落构建的关键因素。在研究方法和手段上,今后需要注重通过选择合适的基因片段建立系统树,然后通过生态特征来加以校正,以更准确地反映物种间的亲缘距离。另外,获得谱系树后还需要寻找更加合理的统计模型和指数,增加统计分析和解决问题的能力。 |
| [27] | . . 物种共存和生物多样性维持一直是生态学研究的中心论题.基于物种生态位分化的群落构建理论已经发展了近一个世纪,但我们对群落构建和生物多样性维持的机理仍然不清楚.近年来,群落中性理论以其简约性和预测能力成为群落生态学研究的焦点,但由于其"物种在生态功能上等价"的假设与大量研究结果相悖,同时对自然群落结构的准确预测也只限于少数的生态系统,因而饱受质疑.如今,越来越多的生态学家认为群落构建的生态位理论与中性理论之争的最终归宿应该是二者的整合.在本文中,我们在简要回顾生态位理论和群落中性理论发展的基础上,分析二者之间的主要分歧和互补性,试图梳理二者整合的途径.我们认为,尽管中性理论的发展极大地丰富了群落构建理论,但二者的整合尚处于初级阶段;群落构建零模型假说、中性一生态位连续体假说、随机生态位假说等都不失为有价值的尝试,今后需要在其他类型的生态系统中进行实验验证,以更好地理解确定性过程和随机过程在决定群落构建和生物多样性维持中的作用. |
| [28] | . A decade has now passed since Hubbell published The Unified Neutral Theory of Biodiversity and Biogeography. Neutral theory highlights the importance of dispersal limitation, speciation and ecological drift in the natural world and provides quantitative null models for assessing the role of adaptation and natural selection. Significant advances have been made in providing methods for understanding neutral predictions and comparing them with empirical data. In this review, we describe the current state-of-the-art techniques and ideas in neutral theory and how these are of relevance to ecology. The future of neutral theory is promising, but its concepts must be applied more broadly beyond the current focus on species-abundance distributions. |
| [29] | . 1) For any taxon higher than genus, the expected mean number of species per genus (E(S/G)) for a randomly drawn subset is a non-decreasing function (approximately linear for many taxa) of the size of the subset, and is therefore lower than the mean number of species per genus for the entire taxon. 2) The relationship of actual S/G for an island biota to S/G expected for a random subset of identical size drawn from the species pool of the presumed source area is relatively insensitive to modifications of taxonomy and to use of a source area slightly larger than the real one. This relationship is subject to more drastic change upon use of incomplete species lists for island or source or a presumed source area smaller than or different from the real source. 3) Pseudo-random drawings of biotas of identical size to those of a number of islands show that, in general, the mean number of species per genus, though lower than in the source biota, is higher than would be expected on a hypothesis of random colonization. 4) The deviation of actual from expected S/G is not strongly correlated with island area, maximum elevation, and distance from source. 5) The claimed positive correlation between species range and genus size, even if it should be shown to exist, is probably insufficient to account for the magnitude of these deviations. 6) The main causes of the excess of insular S/G's over those predicted by chance are probably two simultaneous tendencies: a) similarity of congeneric species in ecological requirements. b) similarity of congeneric species in dispersal capabilities. |
| [30] | . |
| [31] | . . 植物群落中不同"功能身份"物种的多样性与特定生态系统功能之间具有何种关系及其作用机制尚不明确。通过在高寒矮嵩草(Kobresia humilis)草甸为期5年的刈割(不刈割、留茬3cm、留茬1cm)、施肥(施肥、不施肥)和浇水(浇水、不浇水)控制实验,研究了刈割与土壤资源获得性梯度上不同"功能身份"物种(群落中所有物种、响应物种、作用物种和共有物种)的多样性变化与群落地上净初级生产力和稳定性的关系以及稳定性机制。研究结果显示:群落中响应物种、作用物种和共有物种数分别占全部物种数的36.6%、18.3%和64.8%,物种多样性对生态系统功能具有不同的效应,净初级生产力主要受响应物种和作用物种的多样性变化影响,而稳定性则主要由共有物种的多样性变化决定;群落稳定性的维持主要依赖于共有物种的多样性增加,其作用机制是投资组合效应,而超产效应和异步性效应对稳定性并无作用;刈割和施肥对物种多样性、稳定性和净初级生产力具有相反的影响,前者能增加物种多样性和稳定性,并降低净初级生产力,而后者的作用正相反。这与群落中全部物种的多样性变化受刈割影响较大,而作用物种的多样性变化受资源获得性影响较大有关。上述结果表明高寒草甸生态系统地上净初级生产力主要由少数影响生产力的作用物种的多样性决定,而稳定性则由大量共有物种的多样性所掌控。投资组合效应是物种多样性导致稳定性的机制。由于群落中不同物种的多样性效应具有分异性,对于特定的生态系统功能而言,物种的"功能身份"可能比物种多样性本身更重要,不加区别地笼统定义物种多样性与生态系统功能的关系可能欠妥。 |
| [32] | . |
| [33] | . |
| [34] | . |
| [35] | . Abstract MOTIVATION: The increasing availability of phylogenetic and trait data for communities of co-occurring species has created a need for software that integrates ecological and evolutionary analyses. Capabilities: Phylocom calculates numerous metrics of phylogenetic community structure and trait similarity within communities. Hypothesis testing is implemented using several null models. Within the same framework, it measures phylogenetic signal and correlated evolution for species traits. A range of utility functions allow community and phylogenetic data manipulation, tree and trait generation, and integration into scientific workflows. AVAILABILITY: Open source at: http://phylodiversity.net/phylocom/. |
| [36] | |
| [37] | . . 群落构建机制一直是群落生态学研究的核心问题.群落系统发育和功能性状的α及β多样性为探讨 生态位过程和中性过程在局域群落构建中的相对作用提供了新的视角.本研究以哀牢山6 hm2中山湿性常绿阔叶林动态监测样地为研究平台,基于DNA条形码构建的群落系统发育树和树种8个关键功能性状,结合地形数据,分析不同生活史阶段树种 在不同空间尺度的系统发育和功能性状α及β多样性.研究表明:(1)各生活史阶段树种在不同空间尺度上均表现出非随机的系统发育和功能性状结构,系统发育 和功能性状聚集程度随空间尺度的增大而增大;(2)在所有空间尺度上,系统发育和功能性状的周转都为非随机,并且环境距离对系统发育和功能性状β多样性的 解释程度均高于空间距离.研究结果揭示了环境过滤在中山湿性常绿阔叶林群落构建中起主导作用. |
| [38] | . The relationship between species richness and evenness across communities remains an unsettled issue in ecology from both theoretical and empirical perspectives. As a result, we do not know the mechanisms that could generate a relationship between species richness and evenness, and how this responds to spatial scale. Here we examine the relationship between species richness(S) and evenness (Pielou's J' evenness) using a chronosequence of successional sub-alpine meadow communities in the eastern Qinghai-Tibetan Plateau. These meadows range from natural community (never farmed), to those that have been protected from agricultural exploitation for periods ranging from 1 to 10 years. A total of 30 sampling quadrats with size of 0.5 m×0.5 m were laid out along two transects at each meadow. Using correlation analyses we found a consistent negative correlation between S and J' in these communities along the successional gradient at the sampling scale of 0.5 m×0.5 m. We also explored the relationship between S and J' at different sampling scales (from 0.5 m×0.5 m to10 m×10 m) using properly measured ramet-mapped data of a10 m×10 m quadrat in the natural community. We found that S was negatively corrected with J' at the scales of 0.5 m×0.5 m to 2 m×2 m, but such a relationships disappeared at relative larger scales (≥2 m×4 m). When fitting different species abundance models combined with trait-specific methods, we found that niche preemption may be the determining mechanism of species evenness along the succession gradient. Considering all results together, we can conclude that such niche differentiation and spatial scale effects may help to explain the maintenance of high species richness in sub-alpine meadow communities. |
| [39] | . Successional chronosequences provide a unique opportunity to study the effects of multiple ecological processes on plant community assembly. Using a series of 0.5 0.5 m2 plots (n = 30) from five successional sub-alpine meadow plant communities (ages 3, 5, 9, 12, and undisturbed) in the Qinghai-Tibetan Plateau, we investigated whether community assembly is stochastic or deterministic for species and functional traits. We tested directional change in species composition, functional trait composition, and then functional trait diversity measured by Rao's quadratic entropy for four traits plant height, leaf dry matter content, specific leaf area, and seed mass along two comparable successional chronosequences. We then evaluated the importance of species interactions, habitat filtering and stochasticity by comparing with random communities and partitioning the environmental and spatial components of Rao's quadratic entropy. We found no directional change in species composition, but clear directionality in functional trait composition. None of the abiotic environmental variables (except P) showed linear change with successional age, but soil moisture and nitrogen were positively related to functional diversity within meadows. Functional trait diversity increased significantly with the increase in successional age. Comparison with random communities showed a significant shift from trait divergence in early stages of succession (3- and 5-yr) to convergence in the later stages of succession 9-, 12-yr and undisturbed). The relative importance of abiotic variables and spatial structure for functional trait diversity changed in a predictable manner with successional age. Stochasticity at the species level may indicate dispersal limitation, but deterministic effects on functional trait distributions show the role of both habitat effects and biotic interactions. |
| [40] | . . |
1
2003
... 本文采用
古田山木本植物功能性状的系统发育信号及其对群落结构的影响
1
2013
... 如
植物群落构建机制研究进展
2
2016
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests
2012
高寒草地植物物种多样性与功能多样性的关系
1
2017
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
长白山阔叶红松林木本植物系统发育与功能性状结构
1
2014
... 如
Conservation evaluation and phylogenetic diversity
1992
Phylogenies and the comparative method
1985
Functional and phylogenetic community assembly linked to changes in species diversity in a long-term resource manipulation experiment
1
2013
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China
1
2008
... 环境因子和光照是影响亚高寒草甸坡向生境的主要环境条件.坡向的变化对土壤含水量、土壤养分(碳、氮、磷)等都有显著影响(
1
2015
... 环境因子和光照是影响亚高寒草甸坡向生境的主要环境条件.坡向的变化对土壤含水量、土壤养分(碳、氮、磷)等都有显著影响(
Reconciling niche and neutrality: The continuum hypothesis
1
2006
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
Phylogenetic diversity of Amazonian tree communities
2015
1
2001
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
Trait evolution, community assembly, and the phylogenetic structure of ecological communities
2
2007
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
... 本研究结果表明, 各坡向物种数与系统发育多样性指数有相同的变化趋势, 即从南坡到北坡, 物种数增多, 系统发育指数也增大(
放牧强度对高寒草甸群落碳氮磷化学计量特征的影响
1
2017
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
Explaining maximum variation in productivity requires phylogenetic diversity and single functional traits
1
2015
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
亚高寒草甸不同生境植物群落物种多度分布格局的拟合
1
2010
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
甘南高寒草甸植物功能性状和土壤因子对坡向的响应
2012
高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系
2013
植物功能性状研究进展
1
2015
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
沿海拔梯度变化的哀牢山亚热带森林群落系统发育结构
2014
刈割与施肥干扰下高寒草甸植物功能性状的构建模式
1
2014
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
植物功能性状与环境和生态系统
2
2007
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
... ), 其差异反映了植物自身生理过程及其对外部环境异质性的适应策略, 而且能将群落结构与群落环境、生态系统过程等联系起来(
阳坡—阴坡生境梯度上植物群落α多样性与β多样性的变化模式与环境因子的关系
2010
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究
2
2011
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
... 综合环境因子、功能性状和系统发育结果(
群落构建的中性理论和生态位理论
2
2009
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
... ;
The unified neutral theory of biodiversity and biogeography, at age ten
1
2011
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
Taxonomic diversity of island biotas
1
1970
... 群落构建问题一直是群落生态学研究的重点, 群落构建机制的研究旨在说明群落中物种共存和物种多样性的维持机制(
The assembly of tropical tree communities—The advances and shortcomings of phylogenetic and functional trait analyses
2013
刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制
1
2013
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
Exploring the phylogenetic structure of ecological communities: An example for rain forest trees
1
2000
... 以Webb (2000)的试验方法构建群落系统发育树.将样方中所有物种的科属种名称按所要求的格式输入phylomatic在线软件中, 会自动输出由其样方物种构建的系统发育树(
Phylogenies and community ecology
1
2002
... Webb等(2002)在研究热带雨林群落结构中提出群落系统发育的研究方法, 评估群落是否具有系统发育结构, 具有什么样的系统发育结构.植物功能性状是植物为适应生存环境形成的植物形态和生理特征(
Phylomatic: Tree assembly for applied phylogenetics
1
2005
... 以Webb (2000)的试验方法构建群落系统发育树.将样方中所有物种的科属种名称按所要求的格式输入phylomatic在线软件中, 会自动输出由其样方物种构建的系统发育树(
Phylocom: Software for the analysis of phylogenetic community structure and trait evolution
1
2008
... 以Webb (2000)的试验方法构建群落系统发育树.将样方中所有物种的科属种名称按所要求的格式输入phylomatic在线软件中, 会自动输出由其样方物种构建的系统发育树(
1
2015
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
中山湿性常绿阔叶林系统发育和功能性状的α及β多样性
1
2014
... 如
The relationship between species richness and evenness in plant communities along a successional gradient: A study from sub-alpine meadows of the eastern Qinghai-Tibetan Plateau, China
1
2012
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
Using functional trait diversity to evaluate the contribution of multiple ecological processes to community assembly during succession
1
2015
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(
青藏高寒草地植被生产力与生物多样性的经度格局
1
2017
... 甘南亚高寒草甸位于青藏高原东部, 生物资源丰富, 是当地牧草的主产区.但由于其特殊的地理位置, 极其严酷的自然环境, 加之频繁的人类活动的影响, 草场退化十分严重, 生物多样性不断丧失, 使得其生态系统非常脆弱, 一旦破坏就很难恢复, 因此保护亚高寒草甸草地生态系统刻不容缓.随着环境的变化, 亚高寒草甸群落组成, 物种多样性和物种共存方式即群落构建机制也会发生变化, 这引起群落生态学家的关注.与大尺度上的纬度和海拔梯度类似, 坡向梯度在数十米至数百米的小尺度上使得生境条件(光照、温度、水分及土壤养分等)发生有规律的变化.这些生境条件的变化影响植物群落的生长和分布, 进而影响植被类型、群落的物种多样性等.植物的生长及群落结构又会反作用于土壤.因此, 研究植物群落功能性状及功能多样性随坡向的变化规律, 对于认识不同坡向上植物群落的形成和植物对复杂环境的适应, 以及对高寒草甸生态系统功能和结构的维持都有重要的意义.近年来, 对亚高寒草甸植物群落的研究逐渐增多, 主要集中在植物分布格局(


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}