Phenotypic variations in natural populations of Amygdalus pedunculata
LIUJiang-Qun, YINMing-Yu, ZUOSi-Yu, YANGShao-Bing, WUYUNTana*, Non-Timber Forest Research and Development Center, Chinese Academy of Forestry, Zhengzhou 450003, China 通讯作者:* 通信作者Author for correspondence (E-mail: tanatanan@163.ccom) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中央级公益性科研院所基本科研业务费专项资金(CAFYBB2017ZA004-7)
关键词:长柄扁桃;天然种群;表型变异;相关分析 Abstract Aims Our objectives were to determine the phenotypic variations, adaption and distribution patterns in seven natural Amygdalus pedunculata populations.Methods We analyzed 14 phenotypic traits from 120 individuals in seven populations of A. pedunculata by variance analysis, correlation analysis, and cluster analysis.Important findings Results showed that there were plentiful phenotypic variation within and among populations. In particular, the phenotypic variation within population was 40.91%, higher than that among populations (35.29%), which indicated that the phenotypic variation within population was the main source of the phenotypic variation in A. pedunculata. Mean differentiation coefficient was 45.90%, and mean coefficient of variation of 14 traits was 15.59%, ranged from 9.39% to 31.98%. Mean annual temperature, latitude, length of frost-free period, longitude and altitude appear to be prominent ecological factors influencing phenotypic traits. Mean annual temperature and length of frost-free period were key indicators to phenotypic of A. pedunculata in different site conditions. According to principal component analysis and unweighted pair-group method with arithmetic means (UPGMA) cluster analysis, the seven populations of A. pedunculata could be divided into two groups. In mountainous region, A. pedunculata’s leaf blade was usually rotund to oblong, fruit nearly spherical shape with shorter fruit stem, stone was usually ovoid to spherical shape. In contrast, in sandy region, leaf blade was long oval to ovate-lanceolate, fruit and stone was usually flat ovoid with longer fruit stem. Our results provide critical information for the resource collection and breeding of this ecologically important species.
FLD, fruit longitudinal diameter; FTD, fruit transverse diameter; FSD, fruit side diameter; FSI, fruit shape index; FSL, fruit stem length; LL, leaf length; LSI, leaf shape index; LW, leaf width; NLD, nutlet longitudinal diameter; NSD, nutlet side diameter; NSI, nutlet shape index; NTD, nutlet transverse diameter; PL, petiole length; PT, pulp thickness. **, p < 0.01. 新窗口打开 Table 3 表3 表3长柄扁桃7个种群的表型性状及多重比较(平均值±标准偏差) Table 3Phenotypic traits and multiple comparison of the seven Amygdalus pedunculata populations (mean ± SD)
性状 Trait
种群 Population
YYe
SYe
HJG
SSC
LMD
QD
XJG
LL (mm)
22.91 ± 4.43c
24.65 ± 5.84d
20.89 ± 3.89ab
24.15 ± 3.47cd
21.23 ± 1.99b
19.58 ± 2.28a
21.14 ± 3.172b
LW (mm)
7.70 ± 1.97a
8.19 ± 1.73a
11.41 ± 2.34b
13.09 ± 1.95c
14.63 ± 2.34d
13.75 ± 1.90c
12.15 ± 2.49b
PL (mm)
6.19 ± 1.50abc
6.96 ± 1.52d
5.95 ± 1.28ab
6.54 ± 1.31cd
5.77 ± 1.67a
6.38 ± 1.52bc
6.42 ± 1.31bc
LSI
3.11 ± 0.85c
3.06 ± 0.65c
1.86 ± 0.30b
1.86 ± 0.26b
1.48 ± 0.21a
1.44 ± 0.16a
1.80 ± 0.38b
FLD (mm)
12.81 ± 1.22b
13.35 ± 1.29c
11.79 ± 1.42a
12.62 ± 1.43b
13.59 ± 1.32c
12.14 ± 0.97a
12.74 ± 1.09b
FTD (mm)
10.11 ± 0.89a
11.32 ± 1.37c
9.98 ± 1.56a
11.00 ± 1.00bc
12.55 ± 1.31d
10.57 ± 1.03b
11.06 ± 1.29c
FSD (mm)
9.37 ± 0.85a
10.07 ± 1.00b
10.51 ± 1.47bc
11.41 ± 0.90d
12.49 ± 1.29e
10.67 ± 1.07c
11.48 ± 1.31d
FSI
1.38 ± 0.16b
1.34 ± 0.20b
1.13 ± 0.14a
1.11 ± 0.09a
1.09 ± 0.09a
1.14 ± 0.09a
1.12 ± 0.12a
FSL (mm)
6.19 ± 1.21cd
6.46 ± 1.55d
5.20 ± 1.66b
5.80 ± 1.92c
4.30 ± 1.11a
4.05 ± 1.17a
4.01 ± 1.21a
PT (mm)
1.03 ± 0.35bc
0.94 ± 0.10ab
0.86 ± 0.38a
1.16 ± 0.46c
1.15 ± 0.41c
1.46 ± 0.46d
1.40 ± 0.41d
NLD (mm)
11.88 ± 1.26b
11.74 ± 0.91b
11.03 ± 1.22a
11.46 ± 1.38a
11.13 ± 1.08a
11.00 ± 1.16a
11.64 ± 1.13b
NTD (mm)
8.48 ± 1.12a
8.98 ± 0.94b
9.38 ± 0.97cd
9.70 ± 0.72de
9.55 ± 0.56de
9.04 ± 0.95bc
9.79 ± 0.94e
NSD (mm)
7.47 ± 0.64a
7.72 ± 0.89ab
8.22 ± 1.07c
8.70 ± 0.73d
8.67 ± 0.48d
7.85 ± 0.74b
8.30 ± 0.78c
NSI
1.60 ± 0.16a
1.53 ± 0.12a
1.37 ± 0.34bc
1.32 ± 0.12ab
1.28 ± 0.11a
1.41 ± 0.13c
1.41 ± 0.17c
Table 1 and Table 2 indicated the numbers of populations and the abbreviations of phenotypic traits, respectively. Different letters after the number in the same column indicated significant difference at 0.05 level.种群缩写及表型性状缩写分别见表1和表2。表中同列数字后的不同字母表示差异显著(p < 0.05)。 新窗口打开
种群缩写及表型性状缩写分别见表1和表2。 新窗口打开 Table 5 表5 表5长柄扁桃表型性状与地理生态因子间的相关分析 Table 5Analysis of correlation between phenotypic traits and geo-ecological factors in Amygdalus pedunculata populations
性状 Trait
经度 Longitude (E)
纬度 Latitude (N)
海拔 Altitude (m)
年平均气温 AAT (℃)
年日照时间 AAS (h)
年降水量 AP (mm)
无霜期 Frost-free season (d)
LL
-0.344
-0.593
-0.439
0.661
-0.095
0.189
0.606
LW
0.455
0.910**
0.356
-0.958**
0.625
-0.426
-0.797*
PL
0.022
-0.365
0.134
0.513
-0.123
0.377
0.416
LSI
-0.521
-0.959**
-0.478
0.982**
-0.573
0.436
0.862*
FLD
0.001
-0.345
-0.851*
0.269
-0.399
0.537
0.336
FTD
0.292
0.229
-0.566
-0.281
-0.026
0.223
-0.123
FSD
0.594
0.779*
-0.075
-0.810*
0.336
-0.118
-0.707
FSL
-0.676
-0.787*
-0.347
0.842*
-0.119
-0.006
0.813*
PT
0.555
0.493
0.471
-0.566
0.149
0.243
-0.594
FSI
-0.600
-0.988**
-0.419
0.971**
-0.586
0.430
0.899**
NLD
-0.086
-0.729
-0.436
0.732
-0.610
0.721
0.494
NTD
0.778*
0.845*
0.187
-0.755*
0.337
-0.116
-0.801*
NSD
0.486
0.788*
0.043
-0.772*
0.531
-0.354
-0.706
NSI
-0.417
-0.893**
-0.218
0.889**
-0.656
0.529
0.739
Total
5.827
9.701
5.019
10.003
5.165
4.705
8.894
Abbreviations see Table 1 and Table 2. * and ** indicate significant correlation at p < 0.05 and p < 0.01, respectively.缩写同表1、表2。*与**分别表示在p < 0.05和p < 0.01的显著水平。 新窗口打开
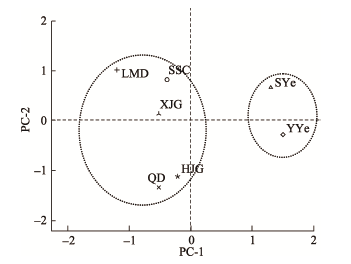
Abbreviations of phenotypic traits was indicated in Table 2.表型性状缩写同表2。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图1基于表型性状(PC-1, PC-2)的长柄扁桃种群关系。种群缩写见表1。 -->Fig. 1The relationship among different Amygdalus pedunculata populations based on PC-1 and PC-2. Table 1 indicated the numbers of populations. -->
2.6 种群聚类分析及不同立地条件下的长柄扁桃表型变异
对7个种群进行UPGMA聚类分析(图2), 基于表型性状将种群划分为两大类, 第一大类包括HJG、XJG、QD、LMD和SSC, 主要立地为山地; 第二大类包括YYe和SYe, 主要立地为沙地。通过比较山地、沙地这两种不同立地条件下生长的长柄扁桃表型性状(表7), 可知立地条件对长柄扁桃不同表型性状具有不同程度的影响, 其中, 对叶长、叶宽、叶形指数、果侧径、果柄长、肉厚、果形指数、核横径、核侧径及核形指数等性状的影响较大。相对于山地上生长的长柄扁桃, 沙地的长柄扁桃叶更长更窄、叶形指数更大, 山地的长柄扁桃叶形为近圆形至长圆形, 而沙地的多为长椭圆形或卵状披针形(图3); 沙地的长柄扁桃的果纵径、果柄、果形指数都大于山地的长柄扁桃, 果横径小于山地的长柄扁桃, 沙地的果实多为卵球形, 而山地的果实多为近球形, 沙地的长柄扁桃果柄普遍长于山地(图4); 沙地的长柄扁桃核横径和核侧径均小于山地的长柄扁桃, 核形指数大于山地的长柄扁桃, 沙地的长柄扁桃核近宽卵形, 山地的长柄扁桃核近宽卵形至近球形。不同立地的长柄扁桃表型特征差异对比见表8。 显示原图|下载原图ZIP|生成PPT 图2基于表型性状的长柄扁桃种群聚类分析。种群缩写同表1。 -->Fig. 2Cluster analysis based on the phenotypic traits of Amygdalus pedunculata populations. Table 1 indicated the numbers of populations. -->
显示原图|下载原图ZIP|生成PPT 图3不同立地下长柄扁桃叶片形态变异。A, 山地。B, 沙地。网格大小为1 cm × 1 cm。 -->Fig. 3The phenotype difference of leaves in Amygdalus pedunculata between different stands. A, Mountainous region. B, Sand. The size of the grid was 1 cm × 1 cm. -->
显示原图|下载原图ZIP|生成PPT 图4不同立地下长柄扁桃果实形态变异。A, 山地果枝。B, 山地果实。C, 山地果肉。D, 沙地果枝。E, 沙地果实。F, 沙地果肉。网格大小为1 cm × 1 cm。 -->Fig. 4The phenotype difference of fruits in Amygdalus pedunculata between different stands. A, Fruit spur from mountainous region. B, Fruit from mountainous region. C, Pulp from mountainous region. D, Fruit spur from sand. E, Fruit from sand. F, Pulp from sand. The size of the grid was 1 cm × 1 cm. -->
Table 7 表7 表7不同立地对长柄扁桃表型的影响 Table 7The influence of different stands on Amygdalus pedunculata populations
性状 Trait
山地 Mountainous region
沙地 Sand
t检验 t-test
分布范围 Rangeability
均值 Mean value
标准偏差 Standard deviation
分布范围 Rangeability
均值 Mean value
标准偏差 Standard deviation
LL (mm)
9.21-33.19
20.75
4.04
12.16-38.91
23.45
4.96
**
LW (mm)
5.21-19.52
12.01
2.61
4.23-13.95
7.85
1.90
**
PL (mm)
2.68-12.22
6.28
1.43
2.87-10.44
6.44
1.57
-
LSI
0.84-3.23
1.77
0.35
1.96-5.93
3.08
0.72
**
FLD (mm)
7.14-19.22
12.65
1.45
9.75-16.21
12.98
1.26
*
FTD (mm)
5.58-14.77
10.68
1.39
8.30-14.65
10.49
1.19
-
FSD (mm)
5.92-15.73
11.20
1.33
7.24-12.06
9.59
0.95
**
FSL (mm)
1.16-10.15
4.47
1.54
3.08-10.07
6.27
1.32
**
PT (mm)
0.15-2.99
1.23
0.47
0.38-2.21
1.00
0.30
**
FSI
0.75-1.66
1.14
0.14
0.94-1.82
1.36
0.17
**
NLD (mm)
7.28-18.15
11.61
1.44
9.89-15.27
11.84
1.16
-
NTD (mm)
7.04-13.45
9.67
0.97
5.82-10.97
8.64
1.09
**
NSD (mm)
2.48-11.93
8.27
0.85
6.09-9.64
7.54
0.73
**
NSI
0.81-4.06
1.42
0.25
1.23-2.01
1.58
0.15
**
Abbreviations of phenotypic traits was indicated in Table 2. * and ** indicate significant correlation at p < 0.05 and p < 0.01, respectively.表型性状缩写同表2。*与**分别表示在p < 0.05和p < 0.01的显著水平。 新窗口打开 Table 8 表8 表8不同立地长柄扁桃形态特征对比 Table 8Comparisons of morphological characteristics of Amygdalus pedunculata populations between different sites
山地 Mountainous region
沙地 Sand
叶片形态 Leaf morphology
叶片近圆形至长圆形, 长9.21-33.19 mm, 宽5.21-19.52 mm; 叶柄长2.68-12.22 mm。 Blade rotund to oblong, 9.21-33.19 mm long, 5.21-19.52 mm wide; petioles 2.68-12.22 mm long.
叶长椭圆形或卵状披针形, 长12.16-38.91 mm, 宽4.23-13.95 mm; 叶柄长2.87-10.44 mm。 Blade long oval to ovate-lanceolate, 12.16-38.91 mm long, 4.23- 13.95 mm wide; petioles 2.87-10.44 mm long.
果实形态 Fruit morphology
果实近球形, 直径5.92-15.73 mm; 果柄较短, 长1.16-10.15 mm; 果肉较厚, 0.15-2.99 mm。 Fruit subglobose, 5.92-15.73 mm in diameter; fruit stem short, 1.16-10.15 mm; pulp thick, 0.15-2.99 mm.
果实卵球形, 直径7.24-12.06 mm; 果柄较长, 长3.08-10.07 mm; 果肉较薄, 0.38-2.21 mm。 Fruit ovoid, 7.24-12.06 mm in diameter; fruit stem long, 3.08-10.07 mm; pulp thin, 0.38-2.21 mm.
核形态 Stone form
核宽卵形至近球形, 直径2.48-11.93 mm。 Stone wide oval to subglobose, 2.48-11.93 mm in diameter.
核宽卵形, 直径6.09-9.64 mm。 Stone wide oval to subglobose, 6.09-9.64 mm in diameter.
新窗口打开 对不同立地条件各地理生态因子进行差异显著性检验(表9), 发现年平均气温和无霜期差异均极显著(p < 0.01), 纬度差异显著(p < 0.05), 表明年平均气温、无霜期是不同立地长柄扁桃表型差异的主要影响因子。 Table 9 表9 表9不同立地各地理生态因子的比较分析 Table 9Comparisons on geographical and ecological factors between different sites
地理生态因子 Geo-ecological factor
山地 Mountainous region
沙地 Sand
t检验 t-test
均值 Mean value
标准偏差 Standard deviation
均值 Mean value
标准偏差 Standard deviation
经度 Longitude (E)
110.48
0.76
109.535
0.47
-
纬度 Latitude (N)
40.81
0.10
38.65
0.28
*
海拔 Altitude (m)
1 394.00
107.60
1 281.00
18.38
-
年平均气温 AAT (℃)
3.78
0.52
8.45
0.49
**
年日照时间 AAS (h)
2 974.80
78.85
2 877.50
2.12
-
年降水量 AP (mm)
352.58
82.62
425.25
16.05
-
无霜期 Frost-free season (d)
113.80
10.64
161.50
10.60
**
AAT, AAS, AP see Table 1. * and ** indicate significant correlation at p < 0.05 and p < 0.01, respectively.AAT、AAS和AP同表1。*与**分别表示在p < 0.05和p < 0.01的显著水平。 新窗口打开
DengLL, SunQ, XuYL, ZhouL, XuY, LiDL, LuoY, ChenS, LiGQ, CaiNH (2016). Compare on the needle phenotypic variations between the different type of trunk populations of Pinus yunnanensis. Journal of Southwest forestry University, 36(3), 30-37. (in Chinese with English abstract)[邓丽丽, 孙琪, 许玉兰, 周丽, 徐杨, 李德龙, 罗元, 陈诗, 李根前, 蔡年辉 (2016). 云南松不同茎干类型群体针叶性状表型多样性比较 . 西南林业大学学报, 36(3), 30-37.]https://doi.org/10.11929/j.issn.2095-1914.2016.03.006URL [本文引用: 1]摘要 在宜良、新平和禄丰3个地点各选择1个有代表性的林分,每个林分从林冠层和更新层分别按通直和扭曲的树干表型性状采集针叶,并对7个针叶表型性状进行测量和比较。结果表明:在林冠层和更新层中,通直群体和扭曲群体间并不存在明显差异,云南松林冠层通直群体和扭曲群体的针叶性状(针叶长度、针叶粗度、针叶束粗、叶鞘长度)平均值均大于更新层,而云南松针叶的形状指数(针叶长度/针叶粗度、针叶长度/叶鞘长度、针叶束粗/针叶粗度)在林冠层和更新层间差异不大;云南松针叶性状在群体间和群体内均存在极其丰富的变异,7个针叶性状指标的变异系数按大小顺序为NL/FSL(22.20%)〉NL/NW(19.15%)〉FSL(19.04%)〉NW(19.00%)〉NL(16.61%)〉FW(15.10%)〉FW/NW(13.20%);云南松针叶性状大部分表现为群体间变异小于群体内变异,群体间总体平均表型分化系数为20.90%,群体内的变异占79.10%,说明群体内变异是云南松针叶性状变异的主要来源。
FengQH, ShiZM, XuZJR, MiaoN, TangJC, LiuXL, ZhangL (2017). Phenotypic variations in cones and seeds of natural Cupressus chengiana populations in China. Chinese Journal of Applied Ecology, 28, 748-756. (in Chinese with English abstract)[冯秋红, 史作民, 徐峥静茹, 缪宁, 唐敬超, 刘兴良, 张雷 (2017). 岷江柏天然种群种实表型变异特征 . 应用生态学报, 28, 748-756.]
[4]
GarcíaD, ZamoraR, GómezJM (2000). Geographical variation in seed production, predation and abortion in Juniperus communis throughout its range in Europe. Journal of Ecology, 88, 435-446. [本文引用: 1]
[5]
GeS, WangMX, ChenYW (1988). An analysis of population genetic structure of masson pine by isozyme technique. Scientia Silvae Sinicae, 24, 399-410. (in Chinese with English abstract)[葛颂, 王明庥, 陈岳武 (1988). 用同工酶研究马尾松群体的遗传结构 . 林业科学, 24, 399-410.] [本文引用: 1]
[6]
GilL, ClimentJ, NanosN, MutkeS, OrtizI, SchillerG (2002). Cone morphology variation in Pinus canariensis Sm. Plant Systematics and Evolution, 235, 35-51. [本文引用: 1]
GuYJ, LuoJX, WuYW, CaoXJ (2009). Phenotypic diversity in natural populations of Picea balfouriana in Sichuan, China. Chinese Journal of Plant Ecology, 33, 291-301. (in Chinese with English abstract)[辜云杰, 罗建勋, 吴远伟, 曾小军 (2009). 川西云杉天然种群表型多样性 . 植物生态学报, 33, 291-301.] [本文引用: 1]
[9]
GuoCH, LuoM, MaYH, MaXW (2005). Study on the salt tolerance of three woody ground cover plants. Journal of Northwest A & F University (Natural Sciences Edition), 33(12), 125-129. (in Chinese with English abstract)[郭春会, 罗梦, 马玉华, 马小卫 (2005). 沙地濒危植物长柄扁桃特性研究进展 . 西北农林科技大学学报(自然科学版), 33(12), 125-129.] [本文引用: 3]
[10]
GuoGG, FengB, MaBL, JingZB, ZhangYL, GuoCH (2013a). Studies on drought resistance of different regional Amygdalus pedunculata Pall. Plant Science Journal, 31, 360-369. (in Chinese with English abstract)[郭改改, 封斌, 麻保林, 井赵斌, 张应龙, 郭春会 (2013a). 不同区域长柄扁桃抗旱性的研究 . 植物科学学报, 31, 360-369.]
[11]
GuoGG, WeiY, FengB, MaBL, ZhangYL, GuoCH (2013b). Gold-resistance of Amygdalus pedunculata Pall from different provenances in China. Journal of Northwest Forestry University, 28(4), 11-15. (in Chinese with English abstract)[郭改改, 魏钰, 封斌, 麻保林, 张应龙, 郭春会 (2013b). 我国几个不同地域长梗扁桃苗木的抗寒性研究 . 西北林学院学报, 28(4), 11-15.] [本文引用: 1]
[12]
HamrickJL, GodtMJW (1990). Allozyme diversity in plant species. In: Brown HD, Clegg MT, Kahler AL, Weir BS eds. Plant Population Genetics, Breeding, and Genetic Resources . Sinauer Associates, Sunderland, USA. 43-63. [本文引用: 1]
[13]
HuangDJ, FengG, LiuJH, WangZB (2016). Study on samara phenotypic variation of Acer palmatum. Journal of Heilongjiang Bayi Agricultural University, 28(5), 36-41. (in Chinese with English abstract)[黄东静, 冯刚, 刘继虎, 万志兵 (2016). 鸡爪槭翅果表型性状变异 . 黑龙江八一农垦大学学报, 28(5), 36-41.] [本文引用: 1]
[14]
HouGF, LiC, ChenB, ShenYH, QianZJ, ZhangYL (2014). Composition analysis of Amygdalus pedunculata Pall seed from different regions. Acta Botanica Boreali-Occidentalia Sinica, 34, 1843-1848. (in Chinese with English abstract)[侯国峰, 李聪, 陈邦, 申烨华, 钱振杰, 张应龙 (2014). 不同产地长柄扁桃种仁成分分析 .西北植物学报, 34, 1843-1848.] [本文引用: 1]
[15]
JiZL, QianAD (1981). Amygdalus pedunculata Pall and A. mongolia Maxim natural distribution area in China. China Fruits, (2), 38-39. (in Chinese)[姬钟亮, 钱安东 (1981). 长柄扁桃和蒙古扁桃在我国自然分布区的调查 . 中国果树, (2), 38-39.] [本文引用: 2]
[16]
JiangB, GuoCH, MeiLX, ShenYH, WangYJ (2008). Studies on cold-resistance of sand Amygdalus pedunculata Pall. Journal of Northwest A & F University (Natural Sciences Edition), 36(5), 92-96. (in Chinese with English abstract)[蒋宝, 郭春会, 梅立新, 申烨华, 王亚俊 (2008). 沙地植物长柄扁桃抗寒性的研究 . 西北农林科技大学学报(自然科学版), 36(5), 92-96. [本文引用: 1]
[17]
JiangXB, GongBC, LiuQZ, ChenX, WuKY, DengQE, TangD (2014). Phenotypic diversity of important agronomic traits of local cultivars of Chinese chestnut. Acta Horticulturae Sinica, 41, 641-652. (in Chinese with English abstract)[江锡兵, 龚榜初, 刘庆忠, 陈新, 吴开云, 邓全恩, 汤丹 (2014). 中国板栗地方品种重要农艺性状的表型多样性 . 园艺学报, 41, 641-652.] [本文引用: 2]
[18]
JiangZM, WuyunTN, WangS, ZhuXC (2016). Amino acid composition and nutritional quality evaluation of wild Amygdalus pedunculatus Pall. kernels from different growing regions. Food Science, 37(4), 77-82. (in Chinese with English abstract)[姜仲茂, 乌云塔娜, 王森, 朱绪春 (2016). 不同产地野生长柄扁桃仁氨基酸组成及营养价值评价 . 食品科学, 37(4), 77-82.] [本文引用: 1]
[19]
LiB, GuWC, LuBM (2002). A study on phenotypic diversity of seeds and cones characteristics in Pinus bungeana. Biodiversity Science, 10, 181-188. (in Chinese with English abstract)[李斌, 顾万春, 卢宝明 (2002). 白皮松天然群体种实性状表型多样性研究 . 生物多样性, 10, 181-188.] [本文引用: 3]
[20]
LiB, LiY, XuNX, ZhangC, ShenYH, ZhangYL (2010). Preparation of activated carbon from amygdalus shell with zinc chloride. Journal of Northwest University (Natural Science Edition), 40, 806-810. (in Chinese with English abstract)[李冰, 李洋, 许宁侠, 张弛, 申烨华, 张应龙 (2010). 氯化锌活化法制备长柄扁桃壳活性炭 . 西北大学学报(自然科学版), 40, 806-810.] [本文引用: 2]
[21]
LiC, LiGP, ChenQ, BaiB, ShenYH, ZhangYL (2010). Fatty acid composition analysis of the seed oil of Amygdalus pedunculatus Pall. China Oils and Fats, 35(4), 77-79. (in Chinese with English abstract)[李聪, 李国平, 陈俏, 白斌, 申烨华, 张应龙 (2010). 长柄扁桃油脂肪酸成分分析 . 中国油脂, 35(4), 77-79.] [本文引用: 1]
[22]
LiDW, DangKL, WenZM (2004). Research of rare and endangered plants of seed plant flora in Loess Plateau. Acta Botanica Boreali-Occidentalia Sinica, 24, 2321-2328. (in Chinese with English abstract)[李登武, 党坤良, 温仲明 (2004). 黄土高原地区种子植物区系中的珍稀濒危植物研究 . 西北植物学报, 24, 2321-2328.] [本文引用: 1]
[23]
LiW, LinFR, ZhengYQ, LiB (2013). Phenotypic diversity of pods and seeds in natural populations of Gleditsia sinensis in southern China. Chinese Journal of Plant Ecology, 37, 61-69. (in Chinese with English abstract)[李伟, 林富荣, 郑勇奇, 李斌 (2013). 皂荚南方天然群体种实表型多样性 . 植物生态学报, 37, 61-69.] [本文引用: 1]
[24]
LinL, WangJH, LuoJ, ChenS (2014). Phenotypic diversity of seed and fruit traits in natural populations of Sophora moorcroftiana. Scientia Silvae Sinicae, 50(4), 137-143. (in Chinese with English abstract)[林玲, 王军辉, 罗建, 陈帅 (2014). 砂生槐天然群体种实性状的表型多样性 . 林业科学, 50(4), 137-143.] [本文引用: 1]
[25]
LiuMJ (1998).Wild Fruits of China. China Agricultural Press, Beijing. 1-8. (in Chinese)[刘孟军 (1998).中国野生果树. 中国农业出版社, 北京. 1-8.] [本文引用: 1]
[26]
LiuYH, GaoGQ, CuiW, ChengCH, YangPH, FanJF (2010). Study on phenotypic diversity of seeds and cones characteristics in Pinus tabuleaformis Carr. Seed, 29(9), 44-48. (in Chinese with English abstract)[刘永红, 高桂琴, 崔嵬, 程春红, 杨培华, 樊军锋 (2010). 油松天然群体种实性状表型多样性分析 . 种子, 29(9), 44-48.] [本文引用: 1]
[27]
LuoJX, GuWC (2005). Study on phenotypic diversity of natural population in Picea asperata. Scientia Silvae Sinicae, 41(2), 66-73. (in Chinese with English abstract)[罗建勋, 顾万春 (2005). 云杉天然群体表型多样性研究 . 林业科学, 41(2), 66-73.] [本文引用: 1]
[28]
MengC, ZhengX, JiZF, LinLL, ZhangCQ, WangYL (2013). Phenotypic diversity of natural populations of Acer grosseri in Shanxi. Acta Botanica Boreali-Occidentalia Sinica, 33, 2232-2240. (in Chinese with English abstract)[孟超, 郑昕, 姬志峰, 林丽丽, 张翠琴, 王祎玲 (2013). 山西葛萝槭天然种群表型多样性研究 . 西北植物学报, 33, 2232-2240.] [本文引用: 2]
[29]
MingJ, GuWC (2006). Phenotypic variation of Syringa oblate Lindl. Forest Research, 19, 199-204. (in Chinese with English abstract)[明军, 顾万春 (2006). 紫丁香表型多样性研究 . 林业科学研究, 19, 199-204.] [本文引用: 1]
[30]
PigliucciM, MurrenCJ, SchlichtingCD (2006). Phenotypic plasticity and evolution by genetic assimilation. Journal of Experimental Biology, 209, 2362-2367.
[31]
SuGX, YaoYQ (1983). Wild almond resources of China. Study on Wild Plant, (2), 7-11. (in Chinese)[苏贵兴, 姚玉卿 (1983). 我国的野生扁桃资源 . 野生植物研究, (2), 7-11.] [本文引用: 1]
[32]
SuYX, ZhangX, WangWL, ZhaoYY, WangYH, ShenSK (2017). Phenotypic diversity of Rhododendron rubiginosum populations at different altitudes. Acta Botanica Boreali-Occidentalia Sinica, 37, 356-362. (in Chinese with English abstract)[苏应雄, 张雪, 王文礼, 赵云勇, 王跃华, 申仕康 (2017). 红棕杜鹃不同海拔种群的表型多样性研究 . 西北植物学报, 37, 356-362.] [本文引用: 1]
[33]
ViaS (1993). Adaptive phenotypic plasticity: Target or by- product of selection in a variable environment? The American Naturalist, 142, 352-365. [本文引用: 1]
[34]
WangW, ChuJM, TangXQ, LiYF, XuXQ (2014). Morphological diversity and correlativity analysis of nut traits of Amygdalus pedunculata. Forest Research, 27, 854-859. (in Chinese with English abstract)[王伟, 褚建民, 唐晓倩, 李毅夫, 许新桥 (2014). 长柄扁桃坚果表型多样性及其相关关系研究 . 林业科学研究, 27, 854-859.]
[35]
WangYL, LiYH, WangY, ZhuQ, WangL (2012). Analysis of nutritional components of three kind of Amygdalus plant. Guangdong Agricultural Sciences, 39(7), 127-129. (in Chinese with English abstract)[王娅丽, 李永华, 王钰, 朱强, 王丽 (2012). 3种扁桃属植物营养成分分析 . 广东农业科学, 39(7), 127-129.] [本文引用: 1]
XuJJ, SuYQ, ZhangQ, GuoCH, XueG, YuquanXYL (2011). Process of the extraction of Amygdalus pedunculata pall. oil. Journal of Northwest Forestry University, 26(4), 184-187. (in Chinese with English abstract)[徐金娟, 苏印泉, 张强, 郭春会, 薛刚, 玉泉幸一郎 (2011). 溶剂法提取长柄扁桃油的工艺研究 . 西北林学院学报, 26(4), 184-187.] [本文引用: 1]
[38]
XuL (2014). Study on Preparation of Lube Base Oil from the Desert Amygdalus pedunculata Pall. Oil. Master degree dissertation, Northwest University, Xi’an. (in Chinese with English abstract)[许龙 (2014). 沙生植物长柄扁桃油改性制备润滑油基础油的研究 . 硕士学位论文, 西北大学, 西安.] [本文引用: 1]
[39]
XuXQ, WangW, ChuJM (2015). Variation analysis on kernel oil content and its fatty acid composition in 31 superior Amygdalus pedunculata individuals form Mu Us Desert. Scientia Silvae Sinicae, 51(7), 142-147. (in Chinese with English abstract)[许新桥, 王伟, 褚建民 (2015). 毛乌素沙地长柄扁桃31个优良单株坚果核仁脂肪酸组成变异分析 . 林业科学, 51(7), 142-147.] [本文引用: 2]
[40]
XuY, ZhouL, CaiNH, DengLL, WangDW, DuanAA, HeCZ, XuYL (2016). Needle phenotypic variation among populations of Pinus yunnanensis at different altitude. Journal of Yunnan Agricultural University (Natural Science), 31(1), 109-114. (in Chinese with English abstract)[徐杨, 周丽, 蔡年辉, 邓丽丽, 王大伟, 段安安, 何承忠, 许玉兰 (2016). 云南松不同海拔群体的针叶性状表型多样性研究 . 云南农业大学学报: 自然科学, 31(1), 109-114.] [本文引用: 2]
[41]
XuYJ, HanHB, WangH, ChenLN, MaQG, PeiD (2016). Phenotypic and genetic diversities of nuts of walnut (Juglans regia) populations originated from seedlings in Daba Mountains. Scientia Silvae Sinicae, 52(5), 111-117. (in Chinese with English abstract)[徐永杰, 韩华柏, 王滑, 陈凌娜, 马庆国, 裴东 (2016). 大巴山区核桃实生居群的坚果表型和遗传多样性 . 林业科学, 52(5), 111-117.] [本文引用: 2]
[42]
YinMY, JiangZM, ZhuXC, BaoWQ, ZhaoH, WuyunT (2016). High-level phenotypic variations in populations of siberian apricot (Armeniaca sibirica) in Nei Mongol. Chinese Journal of Plant Ecology, 40, 1090-1099. (in Chinese with English abstract)[尹明宇, 姜仲茂, 朱绪春, 包文泉, 赵罕, 乌云塔娜 (2016). 内蒙古山杏种群表型变异 . 植物生态学报, 40, 1090-1099.] [本文引用: 2]
[43]
ZhangDH (2009).Descriptors and Data Standard for Almond (Amygdalus).China Agriculture Press, Beijing. (in Chinese)[张大海 (2009). 扁桃种质资源描述规范和数据标准. 中国农业出版社, 北京.] [本文引用: 1]
[44]
ZhangP, ShenYH, GuoCH, KongXH, HeXW, WangJW (2004). HPLC determination of α-tocopherol in almond kernel. Food Science, 25(1), 142-144. (in Chinese with English abstract)[张萍, 申烨华, 郭春会, 孔祥宏, 何学文, 王继武 (2004). 扁桃种仁中维生素E的高效液相色谱法测定 . 食品科学, 25(1), 142-144.] [本文引用: 1]
[45]
ZhangY, CaoYF, HuoHL, TianLM, DongXG, QiD, ZhangXB (2016). Research on diversity of pear germplasm resources based on flowers phenotype traits. Acta Horticulturae Sinica, 43, 1245-1256. (in Chinese with English abstract)[张莹, 曹玉芬, 霍宏亮, 田路明, 董星光, 齐丹, 张小双 (2016). 基于花表型性状的梨种质资源多样性研究 . 园艺学报, 43, 1245-1256.] [本文引用: 1]
[46]
ZhaoZY (1992).Rare and Endangered Plants in Inner Mongolia. China Agriculture Science and Technology Press,Beijing. (in Chinese)[赵一之 (1992).内蒙古珍稀濒危植物图谱. 中国农业科技出版社, 北京.]
[47]
ZhengX, MengC, JiZF, WangYL (2013). Phenotypic diversity of leaves morphologic characteristics of Ulmus lamellosa natural populations in Shanxi. Acta Horticulturae Sinica, 40, 1951-1960. (in Chinese with English abstract)[郑昕, 孟超, 姬志峰, 王祎玲 (2013). 脱皮榆山西天然居群叶性状表型多样性研究 . 园艺学报, 40, 1951-1960.] [本文引用: 2]
[48]
ZuoSY, WuyunT, ZhuXC, DuXL (2015). Diversity of leaves phenotype traits of endangered and wild species Amygdalus pcdunculata. Journal of Central South University of Forestry & Technology, 35(11), 60-67. (in Chinese with English abstract)[左丝雨, 乌云塔娜, 朱绪春, 杜笑林 (2015). 濒危野生长柄扁桃叶片表型性状的多样性 . 中南林业科技大学学报, 35(11), 60-67.]

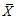
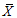
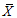
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT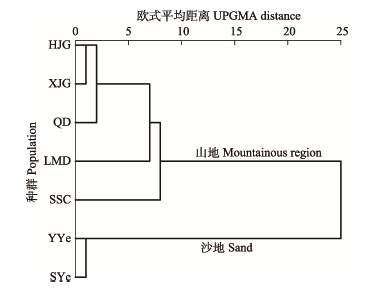 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}