Water sources of Populus euphratica and Tamarix ramosissima in Ejina Delta, the lower reaches of the Heihe River, China
LIYa-Fei1,2,3,4, YUJing-Jie1,5,*, LUKai1,4, WANGPing1, ZHANGYi-Chi1, DUChao-Yang1 1Key Laboratory of Water Cycle & Related Land Surface Processes, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;2University of Chinese Academy of Sciences, Beijing 100049, China3Sino-Danish Center for Education andResearch, Beijing 100190, China4 Sino-Danish College, University of Chinese Academy of Sciences, Beijing 100190, China5College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 国家自然科学基金面上项目(41571029、41371059和41671023)
关键词:干旱区;河岸带;植物水分来源;氧稳定同位素;生态输水 Abstract Aims We aim to evaluate the water sources of typical riparian arbor species (Populus euphratica) and shrub species (Tamarix ramosissima), and analyze the spatial and temporal dynamics of plant water source in Ejina Delta, the lower reaches of the Heihe River, China.Methods Eight sampling sites were selected in the riparian zones along the East River and West River in Ejina. The plant xylem water, soil moisture, rainwater, stream water and groundwater were taken and pretreated during the growing season in 2015-2016, and the stable oxygen isotope ratio (δ18O) for each water sample was measured. The δ18O of plant xylem water and soil moisture were compared to estimate the dominant depth of root water uptake, and the linear-mixed model called “IsoSource” were applied to determine plant water sources and quantify their proportions.Important findings This study indicated that the main recharge sources for P. euphratica and T. ramosissima were stream water and groundwater. The contributions of rain water to them was negligible due to the limited amount and the shallow infiltration depth of local rainfall. As affected by groundwater level fluctuation, soil physical properties, as well as lateral and vertical recharge of stream water on soil moisture, the dominant depth of root water uptake spatially varied. However, the relative contributions of stream water or groundwater to plant water sources did not change significantly across space. Populus euphratica used more stream water (68%), while T. ramosissima used more groundwater (65%). Plant water sources were sensitive to environmental flow controls. The contributions of stream water to the water sources of the two species went up to 84% and 48% for P. euphratica and T. ramosissima respectively during the discharge period, but dropped to 63% and 30% during the non-discharge period. On the other hand, the contributions of groundwater decreased to 16% and 52% during the discharge period, but increased to 37% and 70% during non-discharge period. It is noteworthy that the high similarity of δ18O between stream water and groundwater due to extensive water exchange in the riparian zone made increase the uncertain in quantifying plant water sources.
Keywords:arid region;riparian zone;plant water source;stable oxygen isotope;environmental flow controls -->0 PDF (2483KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 李亚飞, 于静洁, 陆凯, 王平, 张一驰, 杜朝阳. 额济纳三角洲胡杨和多枝柽柳水分来源解析. 植物生态学报, 2017, 41(5): 519-528 https://doi.org/10.17521/cjpe.2016.0381 LIYa-Fei, YUJing-Jie, LUKai, WANGPing, ZHANGYi-Chi, DUChao-Yang. Water sources of Populus euphratica and Tamarix ramosissima in Ejina Delta, the lower reaches of the Heihe River, China. Chinese Journal of Plant Ecology, 2017, 41(5): 519-528 https://doi.org/10.17521/cjpe.2016.0381 额济纳三角洲位于中国第二大干旱内陆河黑河流域的下游, 降水稀少, 蒸发强烈(Wang et al., 2011)。该地区的植被主要依赖于黑河上、中游下泄来水及其补给的土壤水和地下水(Wei et al., 2012), 因此多沿河流两岸及尾闾湖岸分布(朱军涛等, 2011)。自2000年实施生态输水工程以来, 额济纳三角洲河道过水过程和地下水埋深等发生了很大变化(司建华等, 2013; Wang et al., 2013), 该地区植被, 尤其是河岸带植被得到了恢复(张一驰等, 2011)。已有研究表明, 生长在干旱环境中的植物能随水分条件的变化调整其水分利用策略, 以适应干旱缺水环境(Huxman et al., 2004)。实施生态输水工程后, 河岸带植物如何响应水分利用?对这一问题的科学阐释将有助于评估和改进生态输水方案。进行额济纳三角洲代表性植物的水分来源解析是回答这个问题的前提。 利用氧同位素开展植物水分来源解析, 被公认为一种有效的科研手段。****们已利用氧同位素在非洲干旱区(Schachtschneider & February, 2010)、美国西南干旱区(Ehleringer et al., 1991; Snyder & Williams, 2000)以及中国西北干旱区(巩国丽等, 2011; 邢星等, 2014)开展了植物水分来源的研究。土壤水分被植物根系吸收并向木质部运输的过程中不发生氧同位素分馏(Ehleringer & Dawson, 1992), 为利用氧同位素研究植物水分来源提供了理论基础。氧同位素亦被应用于额济纳三角洲的植物水分来源研究(褚建民, 2007; 赵良菊等, 2008; 尹力等, 2012; 陈小丽等, 2014), 但因已有研究选取样点单一和未进行连续采样等, 导致我们对“额济纳三角洲生态输水对植物水分来源的影响”问题认识不足, 亟待进行样点加密和生长期的连续采样, 为系统探究植物对水分的利用提供科学依据。 本研究在额济纳三角洲东、西河沿岸选取8个样点, 于植物生长期连续采集代表性植物胡杨(Populus euphratica)和多枝柽柳(Tamarix ramosissima)木质部、土壤以及各水源样品, 利用氧同位素直接对比法和IsoSource线性混合模型, 研究了额济纳三角洲胡杨和多枝柽柳的水分来源及其时空变化规律。研究所获得的胡杨和多枝柽柳对水分的利用以及对水源变化的响应结果, 可丰富干旱区植物水分来源研究, 并为基于绿洲生态保护的生态输水方案制定提供依据。
1 研究区概况和研究方法
1.1 研究区概况
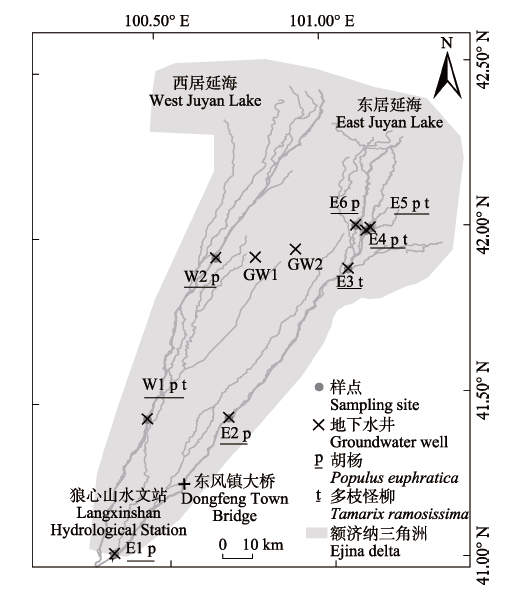
研究区额济纳三角洲(图1)面积约8 022 km2, 属内陆干燥气候, 年平均气温为9.1 ℃, 极端高温为43.7 ℃, 极端低温为-35.3 ℃; 年降水量为34.5 mm (1961-2014年) (杜朝阳, 2016)。黑河经狼心山水文站入额济纳后, 分为东、西河两大支流。据狼心山水文站1991-2016年实测资料, 生态输水工程实施前(1991-1999年)黑河入额济纳的径流量平均为4.78亿m3·a-1, 实施生态输水工程后(2000-2016年)径流量增至5.88亿m3·a-1 (杜朝阳, 2016)。研究区地下水埋深较浅, 南部为1.0-2.0 m, 中部为2.0-3.5 m, 北部为3.5-8.0 m (Wang et al., 2011)。该地区植被稀疏、种群单一, 以旱生、耐盐碱的荒漠植被为主, 绿洲主要分布于东、西河沿岸和低洼地带, 沿河乔灌木植被主要为胡杨、多枝柽柳等(朱军涛等, 2011)。
1.2 采样与数据收集
考虑到研究区东河与西河沿岸水源条件和土壤物理特性的差异、胡杨和多枝柽柳在河岸带的分布(朱军涛等, 2011)以及采样的便利性, 在东、西河沿岸自上而下设置了8个样点(图1)。每月采样1次, 于2015年7-9月和2016年6-9月采集胡杨和多枝柽柳木质部样品和土壤样品。其中2015年设置了样点E2、E3、E4、E5、E6, 2016年增设样点E1和W1、W2。2015年7月和2016年9月, 在输水期(即狼心山水文站向下放水的时期, 包括2015年7月6日到7月22日和2016年8月22日到11月16日)进行采样, 其余月份采样均在非输水期进行; 样点E3和W1因在输水期被河水淹没, 未采集到对应时段的植物和土壤样品。 显示原图|下载原图ZIP|生成PPT 图1研究区位置及样点。图中所标示各地点地理坐标如下: 东风镇大桥(100.55° E, 41.25° N), E1 (100.33° E, 41.04° N), E2 (100.69° E, 41.44° N), E3 (101.07° E, 41.88° N), E4 (101.12° E, 41.99° N), E5 (101.14° E, 42.00° N), E6 (101.09° E, 42.01° N), GW1 (100.79° E, 41.92° N), GW2 (100.91° E, 41.94° N), 狼心山水文站(100.32° E, 41.04° N), W1 (100.44° E, 41.44° N), W2 (100.67° E, 41.92° N)。 -->Fig. 1Location of the study area and sampling sites. The coordinates of the sites in the figure are as follows: Dongfeng Town Bridge (100.55° E, 41.25° N), E1 (100.33° E, 41.04° N), E2 (100.69° E, 41.44° N), E3 (101.07° E, 41.88° N), E4 (101.12° E, 41.99° N), E5 (101.14° E, 42.00° N), E6 (101.09° E, 42.01° N), GW1 (100.79° E, 41.92° N), GW2 (100.91° E, 41.94° N), Langxinshan Hydrological Station (100.32° E, 41.04° N), W1 (100.44° E, 41.44° N), W2 (100.67° E, 41.92° N). -->
植物木质部和土壤样品经室内低温真空抽提(全自动真空冷凝抽提系统LI-2100, LICA United, 北京)获取植物木质部水分和土壤水, 与降水、河水、地下水一起经过滤后, 利用液态水同位素分析仪(LGR DLT-100, Los Gatos Research, Los Gatos, USA)分析其样品的氧稳定同位素比率(δ18O); δ18O的测量精度为0.3‰。 $\delta_{sample}=[\frac{R_{sample}}{R_{standar}}-1]\times 1000 \quad \quad \quad \quad(‰ VSMOW)$ 式中, δsample是样品的氢(或氧)的同位素组成, Rsample是样品中氢(或氧)的重同位素和轻同位素丰度之比(2H/1H或18O/16O), Rstandard是维也纳标准平均海水(VSMOW)的氢(或氧)的重同位素和轻同位素丰度之比(2H/1H或18O/16O), 其中2H/1H值为0.000 155 76, 18O/16O值为0.002 005 2。 Table 1 表1 表1各样点土壤质地剖面 Table 1Soil textural profiles at each sampling site
样点 Sampling site
土壤埋深 Soil depth (cm)
土壤质地 Soil texture
样点 Sampling site
土壤埋深 Soil depth (cm)
土壤质地 Soil texture
E1
0-80
砂土 Sand
E5
0-50
粉壤土 Silt loam
80-85
壤质砂土 Loamy sand
50-100
壤土 Loam
85-90
砂土 Sand
100-130
粉壤土 Silt loam
90-100
细砂 Fine sand
E6
0-30
壤质细砂 Loamy fine sand
100-130
砂土 Sand
30-90
细砂 Fine sand
130-170
细砂 Fine sand
90-120
砂黏土 Sandy clay
170-185
砂壤土 Sandy loam
120-160
细砂 Fine sand
185-200
壤质砂土 Loamy sand
160-180
壤土 Loam
E2
0-20
壤质细砂 Loamy fine sand
180-210
砂土 Sand
20-50
细砂壤土 Fine sandy loam
210-230
黏土 Clay
50-70
极细砂壤土 Very fine sandy loam
230-320
细砂 Fine sand
70-100
粉壤土 Silt loam
W1
0-90
壤质砂土 Loamy sand
100-200
砂土 Sand
90-110
壤质粗砂 Loamy coarse sand
E3
0-30
粉壤土 Silt loam
110-130
砂土 Sand
30-80
细砂壤土 Very fine sandy loam
130-160
粗砂 Coarse sand
80-105
细砂 Fine sand
160-210
砂土 Sand
105-220
粗砂 Coarse sand
W2
0-60
壤质砂土 Loamy sand
E4
0-15
细砂壤土 Fine sandy loam
60-80
砂土 Sand
15-60
砂壤土 Very fine sandy loam
80-120
粉壤土 Silt loam
60-120
粉壤土 Silt loam
120-200
砂土 Sand
120-170
砂土 Sand
200-210
粗砂 Coarse sand
210-230
壤质砂土 Loamy sand
230-260
粗砂 Coarse sand
The coordinates of each sampling site are the same as in Fig.1.各样点地理坐标见图1。 新窗口打开 植物和土壤样品的氧稳定同位素比率(δ18O)采用经光污染校正后的δ18O值, 校正方法与校正公式参见孟宪菁等(2012); 降水、河水和地下水的δ18O直接采用仪器所测δ18O值。
降水的氧稳定同位素比率(δ18O)为-8.89‰ ± 5.74‰ (平均值±标准误差, 下同)(图2A), 河水的δ18O为-6.67‰ ± 1.03‰ (图2B)。 地下水的δ18O为-6.74‰ ± 1.21‰, 但具有空间差异性(表2); 其中各样点处地下水的δ18O为-6.31‰ ± 1.08‰, 距西河10 km的GW1和距东河13 km的GW2处地下水的δ18O为-9.84‰ ± 0.00‰; 各样点位于河岸带, 因此所采集的地下水δ18O波动较大, 且与河水的δ18O较为接近; 远离河道的GW1和GW2处地下水的δ18O较为稳定, 且低于样点处(即河岸带)地下水的δ18O。 显示原图|下载原图ZIP|生成PPT 图2研究区降水(A)与河水(B)的氧稳定同位素比率(δ18O)。VSMOW, 维也纳标准平均海水。 -->Fig. 2The stable oxygen isotope ratio (δ18O) of rainwater (A) and stream water (B) in the study area. VSMOW, Vienna standard mean ocean water. -->
Table 2 表2 表2研究区地下水的氧稳定同位素比率(δ18O) Table 2The stable oxygen isotope ratio (δ18O) of groundwater in the study area
日期 Date
样点 Spot
δ18O (‰)
日期 Date
样点 Spot
δ18O (‰)
2016-5-31
E1
-7.39
2015-8-16
E5
-6.29
2015-7-22
E2
-5.94
2015-9-13
E5
-6.69
2015-8-16
E2
-5.39
2015-8-16
E6
-5.56
2015-9-13
E2
-5.60
2016-5-29
E6
-7.02
2016-5-31
E2
-6.94
2015-7-21
E6
-6.47
2015-8-16
E3
-5.73
2015-9-13
E6
-6.12
2015-9-13
E3
-5.57
2016-5-31
GW1
-9.84
2016-5-30
E3
-6.96
2016-5-31
GW2
-9.84
2015-7-19
E4
-5.77
2016-5-31
W1
-5.95
2015-8-16
E4
-7.35
2016-5-31
W2
-7.08
2015-9-13
E4
-5.22
The coordinates of each sampling site are the same as in Fig.1.各样点地理坐标见图1。 新窗口打开 沿土壤剖面, 土壤水的δ18O表现为从上到下贫化的特点, 即表层土壤水同位素富集程度较高, 深层土壤水同位素富集程度较低, δ18O最高可达7.62‰, 最低为-11.77‰ (图3)。样点E3处沿土壤剖 面土壤水的δ18O变化较小, 表层为-4.66‰, 富集程度较其他样点偏低, 与该处土壤接受河水漫溢补给有关。沿土壤剖面土壤水的δ18O变化存在明显转折点, 转折点以上, δ18O从富集到贫化, 变化较大; 转折点以下, δ18O相对较为稳定, 在小幅范围内波动。 各样点所采集胡杨木质部水分的δ18O为-7.30‰ ± 0.08‰, 最小值为-8.22‰, 最大值为-6.57‰; 多枝柽柳木质部水分的δ18O为-8.58‰ ± 0.08‰, 最小值为-9.27‰, 最大值为-7.56‰ (图3)。 显示原图|下载原图ZIP|生成PPT 图3各样点土壤植物木质部水分的氧稳定同位素比率(δ18O)。各样点地理坐标见图1。VSMOW, 维也纳标准平均海水。 -->Fig. 3The stable oxygen isotope ratio (δ18O) of soil moisture and plant xylem water at each sampling site. The coordinates of each sampling site are the same as in Fig.1. “VSMOW” is the abbreviation of “Vienna standard mean ocean water”. -->
2.2 植物吸收水分的主要土壤层位
植物吸收水分的主要土壤层位采用氧同位素直接对比法得出。图3中与胡杨和多枝柽柳木质部水分的δ18O较为接近的土壤层范围, 即为胡杨和多枝柽柳的主要吸水层位(表3)。植物主要吸水层位的下边界为饱和土壤含水层的上边界, 因各样点处的地下水位埋深而异。植物主要吸水层位的上边界平均为100 cm埋深处, 但各样点有所不同: 样点E3的上边界明显较其他样点浅, 主要是因该样点位于河漫滩, 漫溢河水的补给作用远大于蒸发富集作用对浅层土壤水δ18O的影响; 样点W2的上边界明显较其他样点深, 东河沿岸植物吸水层位上边界较西河沿岸浅, 东河中游沿岸(E2)较东河上游沿岸(E1)浅, 东河下游沿岸E6处的上边界明显深于样点E4和E5。植物吸水层位可能受地下水位波动、土壤物理特性、河水侧向补给的渗透系数以及河水漫溢补给等的影响。 Table 3 表3 表3各样点胡杨和多枝柽柳的主要吸水层位 Table 3Dominant soil depths of root water uptake by Populus euphratica and Tamarix ramosissima at each sampling site
样点1) Sampling site1)
植物主要吸水层位 Dominant soil depths of root water uptake 2) (cm)
2015-7
2015-8
2015-9
2016-6
2016-7
2016-8
2016-9
E1
\
\
\
130-210
80-200
80-260
120-180
E2
100-175
100-200
70-200
80-140
90-140
80-180
80-130
E3
30-220
80-220
50-195
5-170
25-180
\
E4
60-220
60-270
60-220
100-200
110-210
110-230
100-260
E5
50-185
50-185
50-185
90-170
75-240
90-230
110-240
E6
80-330
115-330
115-330
140-370
140-280
115-255
100-235
W1
\
\
\
100-230
110-230
90-?
\
W2
\
\
\
150-260
100-185
170-210
100-150
1) The coordinates of each sampling site are the same as in Fig.1. 2) The values on the left side of the symbol “-” represents the upper most depths of root water uptake, while the values on the right side of the signal “-” represents the lower most depths of root water uptake. The symbol “?” indicates that the sampling did not reach the saturated soil depth due to the impeding of large gravel on soil sampler. “\” indicated that the samples were not taken at the corresponding time, and thus the corresponding results were missing.1) 各样点地理坐标见图1。2) “-”左侧数字为植物吸水层位的上边界, 右侧数字为植物吸水层位的下边界。“?”表示因砾石过大, 土钻无法取样到饱和土壤层。“\”表示对应时段未采集样品, 因此无统计结果。 新窗口打开
2.3 植物的补给水源
该研究区植物可能的补给水源包括降水、河水与地下水。深层土壤受河水侧渗补给和地下水垂向补给, 而降水主要补给表层土壤, 且补给的土壤层范围取决于降水入渗的深度。 样点E5土壤体积含水量与降水数据表明: 在降水开始后, 2 cm埋深的土壤含水量最先开始升高, 4 cm和10 cm埋深土壤含水量升高稍有延迟, 且峰值较低; 20 cm埋深的土壤含水量仅在降水末期略有上升; 40 cm埋深的土壤含水量保持稳定(图4)。由此说明, 降水入渗仅补给0-20 cm埋深范围内的土层; 各样点表层土壤质地类似(壤质砂土、粉壤土、砂壤土, 表1), 因此研究区降水入渗最大深度在20 cm范围内; 除样点E3外, 降水入渗最大深度均未达到其余样点处胡杨和多枝柽柳吸收水分的主要土壤层位。因此对该研究区的胡杨和多枝柽柳而言, 降水的补给可忽略不计, 而将河水与地下水作为胡杨和多枝柽柳的主要补给水源, 也即河水与地下水对胡杨和多枝柽柳的水分贡献率之和为100%, 降水贡献率记为0。 显示原图|下载原图ZIP|生成PPT 图4样点E5 (101.14° E, 42.00° N)的10 min降水与土壤含水量变化(2015-9-9-2015-9-10)。 -->Fig. 4Dynamics of 10-min-gridded precipitation and soil water content at Site E5 (101.14° E, 42.00° N) (2015-9-9-2015-9-10). -->
2.4 补给水源对植物水分的贡献率
利用IsoSource线性混合模型计算河水/地下水对胡杨/多枝柽柳的水分贡献率, 首先需要确定输入模型的河水/地下水的δ18O。考虑到河水补给作用对河岸带地下水δ18O的影响, 以远离河道的GW1和GW2处地下水δ18O值-9.84‰作为供给植物水分的地下水δ18O值代入模型计算。河水的δ18O为-6.67‰ ± 1.03‰ (平均值±标准误差), 分别以河水的“平均值” (-6.67‰)、“(平均值-标准误差)” (-6.67‰ - 1.03‰)和“平均值+标准误差” (-6.67‰ + 1.03‰)作为河水的δ18O值代入模型计算, 得到河水(和地下水)贡献率的平均值以及最大值、最小值。 河水与地下水对胡杨和多枝柽柳的水分贡献率结果如图5所示。河水对胡杨的水分贡献率表现为: 东河(70%)略高于西河(64%); 输水期(2015年7月和2016年9月)河水对东、西河沿岸胡杨贡献率差距较大, 分别为78%和64%; 非输水期差距较小, 分别为66%和64%。东河上(E1)、中(E2)、下游(E3-E6)沿岸河水对胡杨贡献率分别为68%、69%和71%, 输水期分别为85%、82%和86%, 非输水期分别为62%、64%和64%。西河中游沿岸(W1)未采集输水期胡杨样品, 非输水期河水对胡杨贡献率为65%; 西河下游沿岸(W2)输水期和非输水期河水对胡杨贡献率分别为79%和58%。 显示原图|下载原图ZIP|生成PPT 图5河水与地下水对胡杨和多枝柽柳的贡献率。误差棒代表最小值和最大值。各样点地理坐标见图1。 -->Fig. 5Proportional contribution (%) of stream water and groundwater to the water sources of Populus euphratica and Tamarix ramosissima respectively. Bars indicate the minimum and maximum values. The coordinates of each sampling site are the same as in Fig.1. -->
ChenXL, ChenYN, ChenYP (2014). Relationship among water use of different plants in Heihe River riparian forests. Chinese Journal of Eco-Agriculture, 22, 972-979. (in Chinese with English abstract)[陈小丽, 陈亚宁, 陈亚鹏 (2014). 黑河下游荒漠河岸林植物水分利用关系研究 . 中国生态农业学报, 22, 972-979.] [本文引用: 2]
[2]
ChuJM (2007). Studies on Selective Utilization of Water by Plants in Aridland Region. PhD dissertation, Chinese Academy of Forestry, Beijing. (in Chinese with English abstract)[褚建民 (2007). 干旱区植物的水分选择性利用研究 . 博士学位论文, 中国林业科学研究院, 北京.] [本文引用: 1]
[3]
DuCY (2016). Simulation for Coupled Water-Vapor-Air-Heat Flow Transport in Vadose Zone and Estimation of Groundwater Evaporation in Arid Region—A Case Study of Ejina Delta. PhD dissertation, University of Chinese Academy of Sciences, Beijing. (in Chinese with English abstract)[杜朝阳 (2016). 干旱区包气带水-汽-气热耦合模拟及潜水蒸发估算——以额济纳三角洲为例 . 博士学位论文, 中国科学院大学, 北京.] [本文引用: 2]
[4]
EhleringerJ, DawsonT (1992). Water uptake by plants: Perspectives from stable isotope composition. Plant, Cell & Environment, 15, 1073-1082. [本文引用: 1]
[5]
EhleringerJR, PhillipsSL, SchusterWS, SandquistDR (1991). Differential utilization of summer rains by desert plants. Oecologia, 88, 430-434. [本文引用: 1]
[6]
GongGL, ChenH, DuanDY (2011). Comparison of the methods using stable hydrogen and oxygen isotope to distinguish the water source of Nitraria tangutorum. Acta Ecologica Sinica, 31, 7533-7541. (in Chinese with English abstract)[巩国丽, 陈辉, 段德玉 (2011). 利用稳定氢氧同位素定量区分白刺水分来源的方法比较 . 生态学报, 31, 7533-7541.] [本文引用: 1]
[7]
HuxmanTE, SmithMD, FayPA, KnappAK, ShawMR, LoikME, SmithSD, TissueDT, ZakJC, WeltzinJF (2004). Convergence across biomes to a common rain-use efficiency. Nature, 429, 651-654. [本文引用: 1]
[8]
LiX, ChengG, LiuS, XiaoQ, MaM, JinR, CheT, LiuQ, WangW, QiY (2013). Heihe watershed allied telemetry experimental research (HiWATER): Scientific objectives and experimental design. Bulletin of the American Meteorological Society, 94, 1145-1160. [本文引用: 1]
[9]
LiuSM, XuZW, WangWZ, JiaZZ, ZhuMJ, BaiJ, WangJM (2011). A comparison of eddy-covariance and large aperture scintillometer measurements with respect to the energy balance closure problem. Hydrology and Earth System Sciences, 15, 1291-1306. [本文引用: 1]
[10]
MengXJ, WenXF, ZhangXY, HanJJ, SunXM, LiXB (2012). Potential impacts of organic contaminant on δ18O and δD in leaf and xylem water detected by isotope ratio infrared spectroscopy. Chinese Journal of Eco-Agriculture, 20, 1359-1365. (in Chinese with English abstract)[孟宪菁, 温学发, 张心昱, 韩佳音, 孙晓敏, 李晓波 (2012). 有机物对红外光谱技术测定植物叶片和茎秆水δ18O和δD的影响 . 中国生态农业学报, 20, 1359-1365.] [本文引用: 1]
[11]
PengXM, XiaoSC, ChengGD, XiaoHL, TianQY, ZhangQB (2015). Human activity impacts on the stem radial growth of Populus euphratica riparian forests in China’s Ejina Oasis, using tree-ring analysis. Trees, 31, 379-392. [本文引用: 1]
[12]
QianYP, LinXU, QinDJ, WangL (2005). Study on groundwater of the Ejina Basin at the lower reaches of the Heihe River using isotopes. Arid Land Geography, 28, 574-580. (in Chinese with English abstract)[钱云平, 林学钰, 秦大军, 王玲 (2005). 应用同位素研究黑河下游额济纳盆地地下水 . 干旱区地理, 2005, 28, 574-580.] [本文引用: 1]
[13]
SchachtschneiderK, FebruaryEC (2010). The relationship between fog, floods, groundwater and tree growth along the lower Kuiseb River in the hyperarid Namib. Journal of Arid Environments, 74, 1632-1637. [本文引用: 1]
[14]
SiJH, FengQ, XiHY, YuTF, LiW (2013). Determination of critical period and requirement of ecological water demanded in the Ejina Oasis in lower reaches of the Heihe River. Journal of Desert Research, 33, 560-567. (in Chinese with English abstract)[司建华, 冯起, 席海洋, 鱼腾飞, 李炜 (2013). 黑河下游额济纳绿洲生态需水关键期及需水量 . 中国沙漠, 33, 560-567.] [本文引用: 1]
[15]
SnyderKA, WilliamsDG (2000). Water sources used by riparian trees varies among stream types on the San Pedro River, Arizona. Agricultural and Forest Meteorology, 105, 227-240. [本文引用: 1]
[16]
WangP, YuJ, ZhangY, FuG, MinL, AoF (2011). Impacts of environmental flow controls on the water table and groundwater chemistry in the Ejina Delta, northwestern China. Environmental Earth Sciences, 64, 15-24. [本文引用: 2]
[17]
WangP, YuJ, ZhangY, LiuC (2013). Groundwater recharge and hydrogeochemical evolution in the Ejina Basin, northwest China. Journal of Hydrology, 476, 72-86. [本文引用: 1]
[18]
WeiY, FangJ, ZhaoX, ZhangR, LiS (2012). Isotopic model estimate of relative contribution of potential water pools to water uptake of Pinus sylvestris var. mongolica in Horqin Sandy Land. Journal of Resources and Ecology, 3, 308-315. [本文引用: 1]
[19]
XingX, ChenH, ZhuJJ, ChenTT (2014). Water sources of five dominant desert plant species in Nuomuhong area of Qaidam Basin. Acta Ecologica Sinica, 34, 6277-6286. (in Chinese with English abstract)[邢星, 陈辉, 朱建佳, 陈同同 (2014). 柴达木盆地诺木洪地区5种优势荒漠物水分来源 . 生态学报, 34, 6277-6286.] [本文引用: 1]
[20]
YinL, ZhaoLJ, RuanYF, XiaoHL, ChengGD, ZhouMX, WangF, LiCZ (2012). Study of the replenishment sources of typical ecosystems water and dominant plant water in the lower reaches of the Heihe, China. Journal of Glaciology & Geocryology, 34, 1478-1486. (in Chinese with English abstract)[尹力, 赵良菊, 阮云峰, 肖洪浪, 程国栋, 周茅先, 王芳, 李彩芝 (2012). 黑河下游典型生态系统水分补给源及优势植物水分来源研究 . 冰川冻土, 34, 1478-1486.] [本文引用: 2]
[21]
ZhangYC, YuJJ, QiaoMY, YangHW (2011). Effects of eco-water transfer on changes of vegetation in the lower Heihe River Basin. Journal of Hydraulic Engineering, 42, 757-765. (in Chinese with English abstract)[张一驰, 于静洁, 乔茂云, 杨宏伟 (2011). 黑河流域生态输水对下游植被变化影响研究 . 水利学报, 42, 757-765.] [本文引用: 1]
[22]
ZhangYH, WuYQ (2007). Variation of δ18O in water in Heihe River Basin. Advances in Water Science, 18, 864-870. (in Chinese with English abstract)[张应华, 仵彦卿 (2007). 黑河流域不同水体中δ18O的变化 . 水科学进展, 18, 864-870.] [本文引用: 1]
[23]
ZhaoLJ, XiaoHL, ChengGD, SongYX, ZhaoL, LiCZ, YangQ (2008). A preliminary study of water sources of riparian plants in the lower reaches of the Heihe Basin. Acta Geoscientica Sinica, 29, 709-718. (in Chinese with English abstract)[赵良菊, 肖洪浪, 程国栋, 宋耀选, 赵亮, 李彩芝, 杨秋 (2008). 黑河下游河岸林植物水分来源初步研究 . 地球学报, 29, 709-718.] [本文引用: 1]
[24]
ZhuJT, YuJJ, WangP, WangZY (2011). Quantitative classification and analysis of relationships between plant communities and their groundwater environment in the Ejina Desert Oasis of China. Chinese Journal of Plant Ecology, 35, 480-489. (in Chinese with English abstract)[朱军涛, 于静洁, 王平, 王志勇 (2011). 额济纳荒漠绿洲植物群落的数量分类及其与地下水环境的关系分析 . 植物生态学报, 35, 480-489.] [本文引用: 3]
... 研究区额济纳三角洲(图1)面积约8 022 km2, 属内陆干燥气候, 年平均气温为9.1 ℃, 极端高温为43.7 ℃, 极端低温为-35.3 ℃; 年降水量为34.5 mm (1961-2014年) (杜朝阳, 2016).黑河经狼心山水文站入额济纳后, 分为东、西河两大支流.据狼心山水文站1991-2016年实测资料, 生态输水工程实施前(1991-1999年)黑河入额济纳的径流量平均为4.78亿m3·a-1, 实施生态输水工程后(2000-2016年)径流量增至5.88亿m3·a-1 (杜朝阳, 2016).研究区地下水埋深较浅, 南部为1.0-2.0 m, 中部为2.0-3.5 m, 北部为3.5-8.0 m (Wang et al., 2011).该地区植被稀疏、种群单一, 以旱生、耐盐碱的荒漠植被为主, 绿洲主要分布于东、西河沿岸和低洼地带, 沿河乔灌木植被主要为胡杨、多枝柽柳等(朱军涛等, 2011). ...
... (杜朝阳, 2016).研究区地下水埋深较浅, 南部为1.0-2.0 m, 中部为2.0-3.5 m, 北部为3.5-8.0 m (Wang et al., 2011).该地区植被稀疏、种群单一, 以旱生、耐盐碱的荒漠植被为主, 绿洲主要分布于东、西河沿岸和低洼地带, 沿河乔灌木植被主要为胡杨、多枝柽柳等(朱军涛等, 2011). ...
Water uptake by plants: Perspectives from stable isotope composition. 1 1992
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT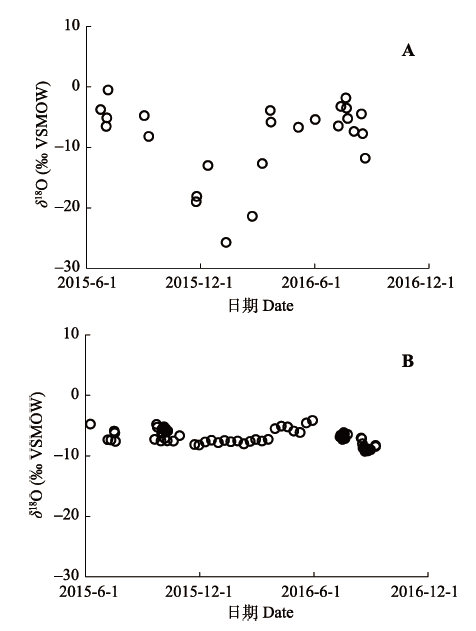 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT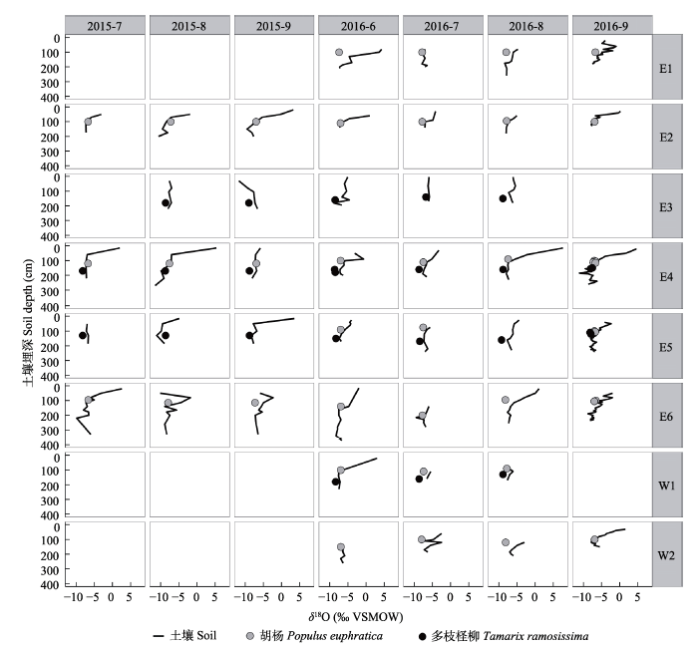 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT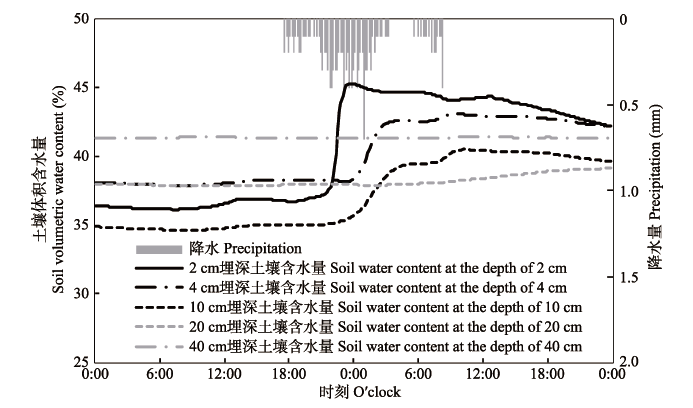 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT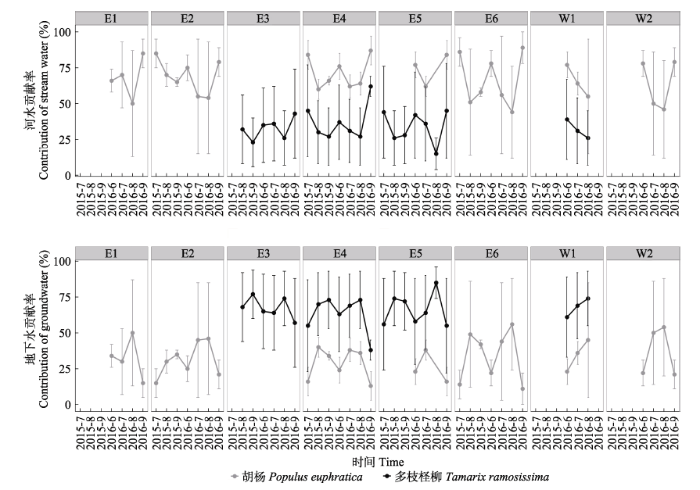 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}