Chemical composition of volatiles from the syconia of Ficus microcarpa and host recognition behavior of pollinating fig wasps
ZHANGTing1,2, WANGBo1, MIAOBai-Ge1, PENGYan-Qiong1,* 1Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan 666303, China2University of Chinese Academy of Sciences, Beijing 100049, China 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31300318、31570418和31600302)和云南省应用基础研究计划项目(2016FA0069)
关键词:榕树;传粉榕小蜂;化学通讯;隐头花序;挥发物 Abstract Aims Chemical communication plays a key role in host plant recognition of pollinators. There are two recognized types of chemical communication between syconia and their pollinating fig wasps: one is “generalization”, of which the wasps respond to the relative ratio of multiple compounds, and the other is “specialization”, of which the key signal is a single uncommon, possibly unique, compound. The aims of this study were to identify the chemical composition of volatiles from the syconia of Ficus microcarpa at different developmental phases, and to determine if the signaling between F. microcarpa and its pollinating fig wasp, Eupristina verticillata, is of generalized type, or of specialized type.Methods The volatiles from syconia of F. microcarpa were extracted using solid-phase micro extraction (SPME) at different developmental phases (pre-female, female (before and after pollination), interfloral, male and postfloral phases) and the chemical compounds were identified by gas chromatography mass spectrometry (GC-MS). We then tested the behavioral responses of E. verticillata to fresh syconia at different developmental phases using two-choice olfactometers.Important findings There were 21 volatile compounds identified from the syconia at different developmental phases, which were mainly fatty acid derivatives, terpenoids and aromatic compounds. The components of the volatiles apparently differed among the developmental stages. The contents of terpenoids declined, but the contents of fatty acid derivatives increased, from before the pollination to after the pollination. Especially, the characteristic compounds of 2-heptanone and 3-octanone before the pollination disappeared, D-limonene decreased after the pollination, but copanene, cyclohexane and 2-hexenal increased. The results of the two-choice olfactometer experiment showed that the pollinating fig wasps had higher selection ratio to chemicals found in the female phase syconia than those in other phases; whereas the volatile compounds from the male phase syconia had the function pushing the pollinating fig wasps to leave the natal syconia so that there existed the “push-pull” responses by fig wasps to volatiles released by their host syconia. We conclude that there are multiple chemical compounds playing the roles in host recognition of pollinating fig wasp E. verticillata. The mutualistic relationship between F. microcarpa and E. verticillata is maintained by the chemical communication of “generalization” strategy.
Keywords:Ficus microcarpa; pollinating fig wasps;chemical communication;syconia;volatiles -->0 PDF (3578KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 张亭, 王波, 苗白鸽, 彭艳琼. 榕树隐头花序挥发物组成及其传粉榕小蜂寄主识别行为. 植物生态学报, 2017, 41(5): 549-558 https://doi.org/10.17521/cjpe.2016.0250 ZHANGTing, WANGBo, MIAOBai-Ge, PENGYan-Qiong. Chemical composition of volatiles from the syconia of Ficus microcarpa and host recognition behavior of pollinating fig wasps. Chinese Journal of Plant Ecology, 2017, 41(5): 549-558 https://doi.org/10.17521/cjpe.2016.0250 昆虫触角是其重要的嗅觉器官, 传粉昆虫的嗅觉在定位寄主植物过程中发挥着重要的作用(钦俊德和王琛柱, 2001)。昆虫的触角并不能感知所有挥发物, 而是仅仅接受特定的信息化合物, 并通过行为上的趋性来定位寄主(杜家纬, 2001)。植物挥发物可由植物的花、叶、果或者根部产生(Whittaker & Feeny, 1971)。除少数植物利用叶的挥发物吸引传粉者(Dufaÿ & Anstett, 2003)外, 绝大多数植物是依赖花的气味向传粉昆虫传递信息。花的挥发物是一种古老的传粉信号, 是植物与传粉昆虫之间交流的语言(Pellmyr & Thien, 1986)。对专性较强的传粉者而言, 其拜访的植物一般都具有区别于其他物种的专一性化学信息物质, 因此利用植物的专一性挥发物, 一些传粉昆虫与植物之间便建立了专性稳定的互利共生关系(Bronstein, 1987; Sakai, 2002)。 植物挥发物以两种方式实现对传粉者的特异性吸引(Grison-Pigé et al., 2002; Hossaert-McKey et al., 2010): 第一种是“泛化” (generalization)策略, 即通过一些常见的挥发性物质以区别于其他物种的组成比例形成特异性, 这些化合物一般是通过生物途径合成, 普遍存在于植物体中, 包括各种脂肪酸衍生物和萜类物质(Degenhardt et al., 2003); 第二种是“专化” (specialization)策略, 即通过释放不常见的单一挥发性物质来实现其特异性, 这些化合物往往是植物次生代谢分解产生的, 如一些芳香族化合物(Dötterl et al., 2005)。已知大多数植物种类采取“泛化”策略吸引传粉者, 采取“专化”策略的植物种类极为少见(Knudsen et al., 1993; Chen et al., 2009)。根据系统发育保守性(phylogenetic conservatism)假设, 亲缘关系较近的植物应该具有相似的挥发物种类和组成(Grison-Pigé et al., 1999), 并在分布于非洲的两种榕树上获得验证(Cornille et al., 2012)。目前, 在大约800种榕树中, 仅提取分析了25种隐头花序雌花期释放的挥发物, 还不足以分析以植物挥发物构建的聚类树和分子数据构建的系统发育树之间的关系(与Proffit Magali个人交流)。因此, 增加榕树种类, 分析其挥发物组成和传粉者的识别机制, 对于掌握榕-蜂化学通讯、了解传粉昆虫-植物协同进化具有重要的意义。 榕树(Ficus spp.)是植物界唯一具有隐头花序(syconium或fig)结构的类群, 依赖专一的榕小蜂(Agaonidae)传粉, 并为传粉榕小蜂的繁衍提供场所, 构成动植物间典型的互惠共生关系, 是研究物种间互作和协同进化的理想材料(Harrison & Shanahan, 2005; Herre et al., 2008; Cook & Segar, 2010; Cruaud et al., 2012)。榕树隐头花序俗称榕果, 它有5个发育时期, 即: 1)雌花前期(pre-female phase, 从发芽到传粉榕小蜂进隐头花序之前这段时期, 在此阶段, 苞片口紧闭着, 雌性小花还未开放); 2)雌花期(female phase, 在此期间, 雌性小花开放, 苞片通道变松, 传粉榕小蜂可以进入隐头花序内传粉、产卵); 3)间花期(interfloral phase, 在此期间, 被传粉、产卵的雌花开始发育成种子及培育子代传粉榕小蜂, 雄花也逐渐发育成熟); 4)雄花期(male phase, 在此阶段, 传粉榕小蜂和雄花同步发育成熟, 羽化、交配后的雌蜂采集花粉离开隐头花序); 5)花后期(postfloral phase, 在此阶段, 种子发育成熟, 成熟隐头花序脱落或被动物取食) (Galil & Eisikowitch, 1968)。雌花期隐头花序能释放特异性挥发物吸引专一的传粉榕小蜂到访。在西双版纳, 同域分布的榕树种类多达46种2个亚种19个变种(许再富等, 1996), 分布区高度重叠, 雌花期短, 传粉榕小蜂成虫寿命也较短, 因此, 雌花期隐头花序特异性化学信号是榕树和传粉榕小蜂成功相遇、稳定繁殖的关键。传粉榕小蜂寻找到专一的寄主是后者挥发性化合物的引诱及隐头花序表面物理特征综合作用的结果(Ware et al., 1993)。目前已对25种榕树隐头花序雌花期释放的挥发物进行了研究, 只发现鸡嗉子榕(Ficus semicordata)执行“专化”策略, 其余种类采用“泛化”策略(Chen et al., 2009)。进一步对不同榕树种类的挥发物及其对传粉昆虫的吸引性进行广泛地鉴定, 了解这两种策略与榕-蜂系统发育或生态特征上的相关性, 将有助于我们深入了解榕-蜂专性互惠关系的演化和维持机制。 榕树(Ficus microcarpa)是隶属于桑科(Moraceae)榕属(Ficus)榕亚属(Subg. Urostigma)榕亚组(Sect. Urostigma)环纹榕亚组(Subsect. Conosycea)的高大乔木(张秀实等, 1998)。榕树种群内全年有隐头花序, 通常树内花期同步, 树间花期异步, 每株榕树每年可多次开花。榕树自然分布于亚洲热带地区。由于它是优良的绿化树种, 被广泛引种到世界各地。榕树隐头花序内寄生有最为多样的榕小蜂种类, 全球共记录43种(Wang et al., 2015)。西双版纳是榕树的原产地, 其榕小蜂种类丰富, 目前已知20种, 包括一种传粉榕小蜂Eupristina verticillata, 一种尚未描述的欺骗性传粉榕小蜂Eupristina sp., 以及18种非传粉小蜂(孔月, 2014)。已知隐头花序释放的挥发物对于维持榕-蜂互惠、共生系统扮演着重要角色(Ware et al., 1993; Proffit et al., 2007), 但在榕树上还未开展相关研究。本研究采用固相微萃取法提取榕树各发育时期隐头花序的挥发物, 并进行分析鉴定, 再配合Y型嗅觉仪检测其传粉榕小蜂对不同发育期隐头花序挥发物的行为反应, 希望回答如下问题: (1)榕树不同发育阶段隐头花序释放的挥发物成分在雌花期传粉前后是如何变化的?(2)榕树不同发育期隐头花序释放的挥发物是否存在相似性? (3)榕树传粉榕小蜂对不同发育期隐头花序的行为反应怎样?本文研究结果不但可以确定榕树及其传粉榕小蜂之间的化学信息交流策略, 而且可为深入探索榕-蜂互惠、欺骗性演化, 以及榕小蜂群落稳定共存提供化学生态学的支持依据。
数据分析中, 各化合物相对含量用峰面积表示, 利用Excel 2010进行峰面积比例计算。采用统计分析软件SPSS 19.0进行传粉榕小蜂对不同发育期隐头花序选择的χ2检验。利用R 3.0.3软件(R Development Core Team, 2015)中Vegan、MASS和lattice数据包进行非度量多维尺度(NMDS)分析。NMDS分析是一种非参数降维方法, 可在二维平面显示不同隐头花序发育期化合物组成的相似性(基于Bray-Curtis距离)。应力值表示NMDS距离与观察矩阵的拟合程度, 该值越小表明拟合越好。同时, 我们使用非加权配对算术平均法(UPGMA)进行聚类分析, 将聚类树(基于Bray-Curtis距离)表示在NMDS二维平面上, 以更直观地表现不同发育期隐头花序释放挥发物的相似性(Wang et al., 2014)。
2 结果
2.1 榕树隐头花序不同发育期释放的挥发物成分
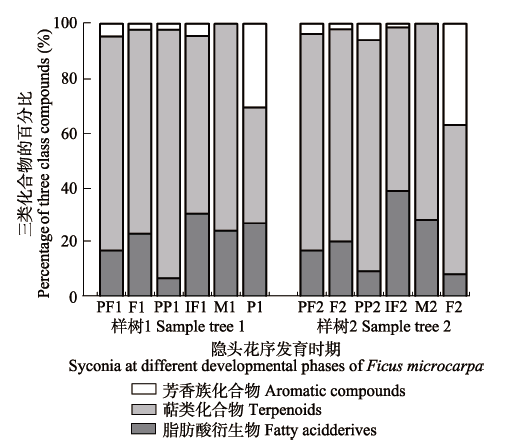
对榕树雌花前期、雌花期传粉前、雌花期传粉后、间花期、雄花期和花后期隐头花序的挥发物进行提取、鉴定和分析。从6个发育时期的隐头花序中共鉴定出21种化合物, 雌花前期、雌花期传粉前、雌花期传粉后、间花期、雄花期和花后期隐头花序挥发物中分别含9、12、10、10、7和5种化合物(表1)。21种化合物可归为3类, 即脂肪酸类衍生物12种、萜类化合物7种、芳香族类化合物2种, 这3类化合物的比例随着隐头花序发育而有明显变化, 在雌花前期、雌花期传粉前、雌花期传粉后的榕树隐头花序挥发物中萜类化合物较多, 样树1上分别为78.89%、75.17%和91.28%, 样树2上分别为79.96%、78.19%和85.26%; 而脂肪酸类衍生物在这3个时期的隐头花序挥发物中所占比例均少于25%。脂肪酸类衍生物在间花期、雄花期和花后期的隐头花序挥发物中含量上升, 样树1上分别占30.69%、23.96%和27.22%, 样树2上分别占39.01%、28.60%和7.98% (图1), 与前3个时期相比, 萜类化合物含量均下降。花后期隐头花序中芳香族化合物明显增加, 样树1和样树2上分别达30.43%和36.80%。其他时期芳香族化合物含量均小于5%,总之, 两株榕树隐头花序释放的化合物随发育期的变化呈现相同规律。 Table 1 表1 表1榕树不同发育期隐头花序释放挥发物的相对含量 Table 1The relative contents of volatile compounds released by syconia of Ficus microcarpa at different developmental phases
挥发物 Volatile
分子式 Formula
相对含量 Relative content (%)
PF1
PF2
F1
F2
PP1
PP2
IF1
IF2
M1
M2
P1
P2
脂肪酸衍生物 Fatty acid derivatives
3,7-二甲基-1,6辛二烯-3-醇 3,7-dimethyl-1,6-octadien-3-ol
C10H18O
0
0
0
0
0
0
0
0
19.55
13.74
0
0
2-庚酮 2-Heptanone
C7H14O
0
0
11.94
8.29
0
0
0
0
0
0
0
0
2-己烯醛 2-Hexenal
C6H10O
2.96
6.24
1.24
0.84
1.78
1.29
4.41
10.13
0
0
0
0
3-己烯-1-醇 3-Hexen-1-ol
C6H12O
13.24
9.93
5.31
7.28
3.95
6.53
13.65
18.17
0
0
0
0
3-辛酮 3-Octanone
C8H16O
0
0
0.58
0.44
0
0
0
0
0
0
0
0
癸醛 Decanal
C10H20O
0
0
0
0
0
0
6.71
2.60
0
0
0
0
癸烷 Decane
C10H22
0
0
0
0
0
0
4.16
6.12
0
0
0
0
十二烷 Dodecane
C12H26
0
0
0
0
0
0
0
0
2.22
1.55
0
0
己醛 Hexanal
C6H12O
0
0
3.67
2.95
0.77
1.13
0
0
0
0
0
0
壬醛 Nonanal
C9H18O
0
0
0
0
0
0
0
0
2.19
13.31
0
0
辛酸乙酯 Octanoic acid, ethyl ester
C10H20O2
0
0
0
0
0
0
1.76
1.99
0
0
0
0
十三烷 Tridecane
C13H28
0.32
0.30
0
0
0
0
0
0
0
0
27.22
7.98
萜类化合物 Terpenoids
α-荜澄茄油烯 α-cubebene
C15H24
0
0
1.09
0.45
1.76
1.83
0
0
0
0
0
0
1,2,4-甲桥-苯并环戊烯 1,2,4-Metheno-1H-indene
C15H24
9.23
6.46
7.89
8.84
0.99
1.88
4.37
3.85
8.26
8.49
0
0
3,7-二甲基-1,3,6环己烯 3,7-dimethyl-1,3,6-octatriene
C10H16
0
0
0.47
0.55
2.34
1.09
11.63
13.75
12.33
11.17
0
0
反式-2,4-己二烯 (E,E)-2,4-hexadiene,
C6H10
4.43
7.53
0
0
0
0
0
0
0
0
0
0
石竹烯 Caryophyllene
C15H24
8.04
7.95
5.92
8.19
2.71
1.45
6.70
8.89
6.94
8.71
6.06
11.44
可巴烯 Copaene
C15H24
54.85
50.85
44.33
52.82
71.08
76.85
42.21
33.23
48.51
43.03
36.28
43.77
D-柠檬烯 D-limonene
C10H16
2.34
7.17
15.46
7.34
12.40
2.16
0
0
0
0
0
0
芳香族化合物 Aromatic compounds
苯甲酸乙酯 Benzoic acid, ethyl ester
C9H10O2
0
0
0
0
0
0
4.40
1.27
0
0
15.39
18.67
环己烷 Cyclohexane
C6H12
4.59
3.57
2.09
2.00
2.21
5.78
0
0
0
0
15.04
18.13
F, IF, M, P, PF and PP represent the syconia at female phase before pollination, interfloral phase, male phase, postfloral phase, pre-female phase, and post-pollination phase, respectively. No.1 indicates sample tree 1 and No. 2 indicates sample tree 2.F、IF、M、P、PF和PP分别表示榕树雌花期传粉前、间花期、雄花期、花后期、雌花前期、雌花期传粉后的隐头花序。带1表示样树1, 带2表示样树2。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图1脂肪酸类衍生物、萜类和芳香族化合物在榕树隐头花序不发育期间的变化情况。缩写同表1。 -->Fig. 1Changes in fatty acid derivatives, terpenoids and aromatics compounds from syconia of Ficus microcarpa with developmental phases. Abbreviations are the same as in Table 1. -->
虽然有些化合物同时存在于2个或多个发育时期隐头花序的挥发物中, 如可巴烯和石竹烯在6个发育时期中都有, 但它们在各时期所占的相对含量不同。隐头花序每个发育时期挥发物的化学特性均由少数几种化合物表征, 例如, 雌花期传粉前隐头花序共检测到12种挥发物, 主要是可巴烯、D-柠檬烯和2-庚酮, 样树1和样树2上的含量分别占该时期总量的71.73%和68.54%; 传粉后的隐头花序只发现10种挥发物成分, 以可巴烯、D-柠檬烯和3-己烯-1-醇为主, 样树1和样树2上的含量分别占该时期总量的87.43%和85.54%; 间花期主要以可巴烯和3,7-二甲基-1,3,6环己烯为主, 样树1和样树2上的含量分别占该时期总量的53.84%和46.98% (表1)。在榕树隐头花序整个发育期, 样树1和样树2上隐头花序挥发物释放量在雌花期传粉前均最多, 该期含量分别达39%和40%; 传粉后化合物释放量比传粉前明显降低, 分别为15%和24%; 花后期都最少, 化合物释放量分别为3%和2%; 花前期、间花期和雄花期化合物释放量较接近(图2)。 显示原图|下载原图ZIP|生成PPT 图2隐头花序总挥发物在榕树隐头花序6个发育期的变化。缩写同表1。 -->Fig. 2Changes in the total volatiles from syconia of Ficus microcarpa at six developmental phases. Abbreviations are the same as in Table 1. -->
2.2 雌花期传粉前后挥发物的变化
雌花期传粉前, 榕树隐头花序挥发物主要以萜类化合物和脂肪酸衍生物为主, 传粉后萜类和芳香族化合物含量有所增加, 脂肪酸衍生物含量则下降。雌花期传粉前, 可巴烯、D-柠檬烯和2-庚酮含量相对较高。传粉24 h后2-庚酮和3-辛酮消失, D-柠檬烯、1,2,4-甲桥-苯并环戊烯、石竹烯、己烯醛和3-己烯-1-醇含量则下降, 推测这些物质主要起吸引传粉榕小蜂定位雌花期隐头花序的作用。而另外一些化合物如可巴烯、环己烷、2-己烯醛、3,7-二甲基-1,3,6环己烯和α-荜澄茄油烯, 在传粉后含量有所增加, 推测这些挥发物可能利于趋避传粉榕小蜂选择已进过蜂的隐头花序(图3)。 显示原图|下载原图ZIP|生成PPT 图3榕树隐头花序挥发物中的12个主要化合物在雌花期传粉前后的变化。A, 2-庚酮; B, 2-己烯醛; C, 3-己烯-1-醇; D, 3-辛酮; E, 己醛; F, α-荜澄茄油烯; G, 1,2,4-甲桥-苯并环戊烯; H, 3,7-二甲基-1,3,6环己烯; I, 石竹烯; J, 可巴烯; K, D-柠檬烯; L, 环己烷。 -->Fig. 3Changes in the 12 major compounds from syconia of Ficus microcarpa between the female phases before and after pollination. A, 2-heptanone; B, 2-hexenal; C, 3-hexen-1-ol; D, 3-octanone; E, hexanal; F, α-cubebene; G, 1,2,4-metheno-1H- indene; H, 1,3,6-octatriene,3,7-dimethyl; I, caryophyllene; J, copaene; K, D-limonene; L, cyclohexane. F, female phase before pollination; PP, post-pollination phase. -->
2.3 榕树各发育时期隐头花序挥发物组成的相似性
通过对榕树不同发育期隐头花序挥发物进行聚类分析和NMDS分析(stress = 0.08, R2 = 0.99), 结果发现: 花后期与其他时期隐头花序挥发物距离最远, 单独成支; 间花期与雄花期聚为一支; 花前期、雌花期传粉前后聚为一支, 雌花前期和雌花期传粉前距离最近, 雌花期传粉前后隐头花序挥发物组成有明显距离(图4)。 显示原图|下载原图ZIP|生成PPT 图4榕树隐头花序各发育时期所对应的挥发物多维尺度分析和聚类树。F1、F2、IF1、IF2、M1、M2、P1、P2、PF1、PF2、PP1和PP2见表1。实线代表根据两株树上不同发育期隐头花序的化学物质组成, 基于Bray-Curtis距离, 使用UPGMA法生成的聚类树。 -->Fig. 4Non-metric multidimensional scaling (ordination) of chemical composition of volatiles from syconia of Ficus microcarpa at different developmental phases. F1, F2, IF1, IF2, M1, M2, P1, P2, PF1, PF2, PP1 and PP2 are the same as in Table 1. The solid line shows the dendrogram based on distance calculated from chemical percentage data using Bray-Curtis measures of dissimilarity. Each datum point represents a specific phase of a tree. -->
2.4 榕树传粉榕小蜂对不同发育期隐头花序的行为反应
当榕树传粉榕小蜂在各发育时期隐头花序挥发物与空气对照之间做选择时, 榕树传粉榕小蜂最偏爱选择雌花期传粉前的隐头花序(χ2 = 17.19, N = 21, p < 0.001), 对雌花前期(χ2 = 9.97, N = 29, p < 0.01)和间花期隐头花序(χ2 = 4, N = 19, p < 0.05)也有一定的趋向性, 而显著地避开雄花期隐头花序(χ2 =16.20, N = 18, p < 0.001), 对雌花期传粉后的隐头花序(χ2 = 1.80, N = 20, p = 0.18)无明显的趋向性。当榕树传粉榕小蜂在对雌花前期与雌花期传粉后(χ2 = 0.80, N = 20, p = 0.371)及间花期隐头花序(χ2 = 0.18, N = 49, p = 0.668)间作选择时, 没有明显的趋向性, 而在雌花前期与雄花期隐头花序间做选择时, 显著地趋向于选择雌花前期隐头花序(χ2 = 12.46, N = 26, p < 0.001)。当榕树传粉榕小蜂在雌花期传粉前与其他时期隐头花序间作选择时, 除对雌花前期隐头花序(χ2 = 2.38, N = 42, p = 0.123)趋向性不明显外, 它们都显著地趋向于选择雌花期传粉前隐头花序; 当榕树传粉榕小蜂在雌花期传粉后与间花期(χ2 = 9.32, N = 31, p = 0.002)及雄花期隐头花序(χ2 = 20.17, N = 22, p < 0.001)间做选择时, 都显著趋向于雌花期传粉后隐头花序。传粉榕小蜂在间花期和雄花期隐头花序间选择时, 也显著地避开雄花期隐头花序(图5)。总之, 榕树雌花期隐头花序对传粉榕小蜂的吸引能力最强, 雄花期隐头花序则能趋避传粉榕小蜂, 表现出雌花期“拉”及雄花期“推”的作用模式。 显示原图|下载原图ZIP|生成PPT 图5用Y型嗅觉仪检测榕树传粉榕小蜂对榕树各发育期隐头花序的选择性行为反应结果。卡方检验: *, p < 0.05; **, p < 0.01; ***, p < 0.001, NS, p > 0.05。CA, 空气对照; F, 雌花期传粉前隐头花序; IF, 间花期隐头花序; M, 雄花期隐头花序; NC, 无反应的榕小蜂数量; PF, 雌花前期隐头花序; PP, 雌花期传粉后隐头花序。 -->Fig. 5Behavioural responses of Eupristina verticillata to syconia of Ficus microcarpa at different developmental phases in Y-tube olfactometer tests. χ2 test: *, p < 0.05; **, p < 0.01; ***, p < 0.001, NS, p > 0.05. CA, clean air; F, female syconia before pollination; IF, interfloral syconia; M, male phase syconia; NC, number of irresponsive fig wasps; PF, pre-female syconia; PP, post-pollination syconia. -->
BronsteinJL (1987). Maintenance of species-specificity in a neotropical fig: Pollinator wasp mutualism. Oikos, 48, 39-46. [本文引用: 1]
[2]
ChangXS, WuCY, CaoZY (1998). Flora of China 23 (1). Science Press, Beijing. 112-113. (in Chinese)[张秀实, 吴征镒, 曹子余 (1998). 中国植物志第23卷第1分册, 科学出版社, 北京. 112-113.]
[3]
ChenC, SongQS, ProffitM, BessièreJM, LiZB, Hossaert-McKeyM (2009). Private channel: A single unusual compound assures specific pollinator attraction in Ficus semicordata. Functional Ecology, 23, 941-950. [本文引用: 3]
[4]
ChenYR, ChouLS, WuWJ (2001). Regulation of fig wasp entry and egress: The role of ostiole of Ficus microcarpa L. Formosan Entomologist, 21, 171-182. [本文引用: 1]
CornilleA, UnderhillJG, CruaudA, Hossaert-McKeyM, JohnsonSD, TolleyKA, KjellbergF, van NoortS, ProffitM (2012). Floral volatiles, pollinator sharing and diversification in the fig-wasp mutualism: Insights from Ficus natalensis, and its two wasp pollinators (South Africa). Proceedings of the Royal Society B: Biological Sciences, 279, 1731-1739. [本文引用: 1]
[7]
CruaudA, RønstedN, ChantarasuwanB, ChouLS, ClementWL, CoulouxA, CousinsB, GensonG, HarrisonRD, HansonPE, Hossaert-McKeyM, Jabbour-ZahabR, JousselinE, KerdelhuéC, KjellbergF, Lopez-VaamondeC, PeeblesJ, PengYQ, PereiraRA, SchrammT, UbaidillahR, van NoortS, WeiblenGD, YanDR, YodpinyaneeA, Libeskind-HadasR, CookJM, RasplusJY, SavolainenV (2012). An extreme case of plant-insect codiversification: Figs and fig-pollinating wasps. Systematic Biology, 61, 1029-1047. [本文引用: 1]
[8]
DegenhardtJ, GershenzonJ, BaldwinIT, KesslerA (2003). Attracting friends to feast on foes: Engineering terpene emission to make crop plants more attractive to herbivore enemies. Current Opinion in Biotechnology, 14, 169-176. [本文引用: 1]
[9]
DötterlS, FüsselU, JürgensA, AasG (2005). 1, 4-Dimethoxybenzene, a floral scent compound in willows that attracts an oligolectic bee. Journal of Chemical Ecology, 31, 2993-2998. [本文引用: 1]
[10]
DuJW (2001). Plant-insect behavior and chemical communication and the behavior control. Plant Physiology Communications, 27(3), 193-200. (in Chinese with English abstract)[杜家纬 (2001). 植物-昆虫间的化学通讯及其行为控制 . 植物生理学报, 27(3), 193-200.] [本文引用: 1]
[11]
DufaÿM, AnstettMC (2003). Conflicts between plants and pollinators that reproduce within inflorescences: Evolutionary variations on a theme. Oikos, 100, 3-14. [本文引用: 1]
[12]
GalilJ, EisikowitchD (1968). On the pollination ecology of Ficus sycomorus in East Africa. Ecology, 49, 259-269. [本文引用: 1]
[13]
GengXZ, WangB, JiaAQ, WangRW (2014). Roles of semiochemicals in regulating intraspecific competition of pollinating wasps of Ficus racemosa. Biodiversity Science, 22, 189-195. (in Chinese with English abstract)[耿向宗, 王波, 贾爱群, 王瑞武 (2014). 聚果榕小蜂种内竞争的化学信息调节机制初探 . 生物多样性, 22, 189-195.] [本文引用: 1]
[14]
Grison-PigéL, EdwardsAA, Hossaert-McKeyM (1999). Interspecies variation in floral fragrances emitted by tropical Ficus species. Phytochemistry, 52, 1293-1299. [本文引用: 1]
GuD, ComptonSG, PengYQ, YangDR (2012). ‘Push’ and ‘pull’ responses by fig wasps to volatiles released by their host figs. Chemoecology, 22, 217-227. [本文引用: 1]
[17]
HarrisonRD, ShanahanM (2005). Seventy-seven ways to be a fig: Overview of a diverse plant assemblage. In: Roubik DW, Sakai S, Hamid AA eds . Pollination Ecology and the Rain Forest Canopy: Sarawak Studies. Springer-Verlag, New York.111-127.
[18]
HerreEA, JandérKC, MachadoCA (2008). Evolutionary ecology of figs and their associates: Recent progress and outstanding puzzles. Annual Review of Ecology, Evolution, and Systematics, 39, 439-458. [本文引用: 1]
[19]
Hossaert-McKeyM, SolerC, SchatzB, ProffitM (2010). Floral scents: Their roles in nursery pollination mutualisms. Chemoecology, 20, 75-88. [本文引用: 1]
[20]
KnudsenJT, TollstenL, BergströmLG (1993). Floral scents—A checklist of volatile compounds isolated by head-space techniques. Phytochemistry, 33, 253-280. [本文引用: 1]
[21]
KongY (2014). The Structure and Function of Fig Wasp Community on Ficus microcarpa in Different Locations in Yunnan. Master degree dissertation, Institute of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Kunming. 13-14. (in Chinese with English abstract)[孔月 (2014). 云南不同分布区榕树(Ficus microcarpa)果内榕小蜂群落结构和功能的研究 . 硕士学位论文, 中国科学院西双版纳热带植物园, 昆明. 13-14.] [本文引用: 1]
[22]
LiZB (2010). The Role of Fig Volatiles on the Host Selecting Behavior of the Pollinating Fig Wasps. PhD dissertation, Institute of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Kunming. 106-108. (in Chinese with English abstract)[李宗波 (2010). 榕树隐头果挥发性化合物在传粉小蜂寄主选择过程中的作用 . 博士学位论文, 中国科学院西双版纳热带植物园, 昆明. 106-108.]
[23]
LiZB, YangP, PengYQ, YangDR (2012). Analysis on chemical compounds and characteristics of fig volatiles emitted from Ficus curtipes Corner. Journal of Tropical and Subtropical Botany, 20, 292-298. (in Chinese with English abstract)[李宗波, 杨培, 彭艳琼, 杨大荣 (2012). 钝叶榕隐头果挥发物成分及其构成特征分析, 热带亚热带植物学报, 20, 292-298.] [本文引用: 2]
[24]
LiuC, YangDR, ComptonSG, PengYQ (2013). Larger fig wasps are more careful about which figs to enter -- With good reason. PLOS ONE, 8, e74117. doi: 10.1371/journal. pone.0074117. [本文引用: 2]
[25]
LuRC, ChenJJ, LinYJ, WenHF, ShiWJ (2015). The analysis of the volatile commounds which collected from Pelargonium citrosum Van Leenii. Journal of Northwest Forestry University, 30, 213-216. (in Chinese with English abstract)[路荣春, 陈佳佳, 林宇杰, 闻海峰, 施文健 (2015). 驱蚊草( Pelargonium citrosum Van Leenii)挥发物成分的采集与分析. 西北林学院学报, 30, 213-216.] [本文引用: 1]
[26]
ProffitM, SchatzB, BorgesRM, Hossaert-McKeyM (2007). Chemical mediation and niche partitioning in non-pollinating fig-wasp communities. Journal of Animal Ecology, 76, 296-303. [本文引用: 1]
[27]
ProffitM, SchatzB, BessièreJM, ChenC, SolerC, Hossaert- McKeyM (2008). Signalling receptivity: Comparison of the emission of volatile compounds by figs of Ficus hispida before, during and after the phase of receptivity to pollinators. Symbiosis, 45, 15-24. [本文引用: 1]
[28]
PellmyrO, ThienLB (1986). Insect reproduction and floral fragrances: Keys to the evolution of the angiosperms? Taxon, 35, 76-85. [本文引用: 1]
[29]
QinJD, WangCZ (2001). The relation of interaction between insects and plants to evolution. Acta Entomologica Sinica, 44, 360-365.(in Chinese with English abstract). [钦俊德, 王琛柱 (2001). 论昆虫与植物的相互作用和进化的关系. 昆虫学报, 44, 360-365.] [本文引用: 1]
SakaiS (2002). Aristolochia spp. (Aristolochiaceae) pollinated by flies breeding on decomposing flowers in Panama. American Journal of Botany, 89, 527-534. [本文引用: 2]
[32]
SolerC, Hossaert-McKeyM, BuatoisB, BessièreJM, SchatzB, ProffitM (2011). Geographic variation of floral scent in a highly specialized pollination mutualism. Phytochemistry, 72, 74-81. [本文引用: 1]
[33]
SongQS, YangDR, ZhangGM, YangCR (2001). Volatiles from Ficus hispida and their attractiveness to fig wasps. Journal of Chemical Ecology, 27, 1929-1942. [本文引用: 1]
[34]
VerkerkeV (1989). Structure and function of the fig. Experientia, 45, 612-622. [本文引用: 1]
[35]
WangB, GengXZ, MaLB, CookJM, WangRW (2014). A trophic cascade induced by predatory ants in a fig-fig wasp mutualism. Journal of Animal Ecology, 83, 1149-1157. [本文引用: 1]
[36]
WangR, AylwinR, LouiseB, ChenXY, ChenY, ChouLS, CobbJ, ColletteD, CaraineL, GiblinR, GhanaS, HarperM, HarrisonR, McphersonJ, PengYQ, PereiraR, ReyesA, RodriguezLJV, StrangeE, van NoortS, YangHW, YuH, ComptonSG (2015). The fig wasp followers and colonists of a widely introduced fig tree, Ficus microcarpa. Insect Conservation & Diversity, 8, 322-336. [本文引用: 2]
[37]
WareAB, KayePT, ComptonSG, van NoortS (1993). Fig volatiles: Their role in attracting pollinators and maintaining pollinator specificity. Plant Systematics & Evolution, 186, 147-156. [本文引用: 1]
[38]
WhittakerRH, FeenyPP (1971). Allelochemics: Chemical interactions between species. Science, 171, 757-770.
[39]
XuZF, ZhuH, YangDR (1996). Species diversity and ecological significance of Ficus in tropical rain forest of southern Yunnan Province. In: Chen XW, Zhang YH eds,. Collected Research Papers on the Tropical Botany (Ⅳ) .Yunnan University Press . Kunming . 1-15. (in Chinese)[许再富, 朱华, 杨大荣(1996). 滇南热带雨林榕树类群多样性及生态学意义. 见: 陈新文, 张永宏编.热带植物研究论文报告集, 第4集. 云南大学出版社, 昆明. 1-15.] [本文引用: 1]
[40]
ZhangKY (1963). A preliminary analysis of the characteristics and formation factors of climate in south Yunnan. Journal of Meteorology, 33, 218-230. (in Chinese)[张克映 (1963). 滇南气候的特征及其形成因子的初步分析 . 气象学报, 33, 218-230.] [本文引用: 1]
[41]
ZhangY, YangDR, PengYQ, ComptonSG (2012). Costs of inflorescence longevity for an Asian fig tree and its pollinator. Evolutionary Ecology, 26, 513-527.
Maintenance of species-specificity in a neotropical fig: Pollinator wasp mutualism. 1 1987
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT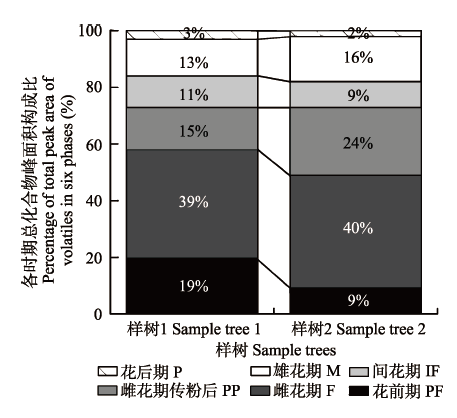 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT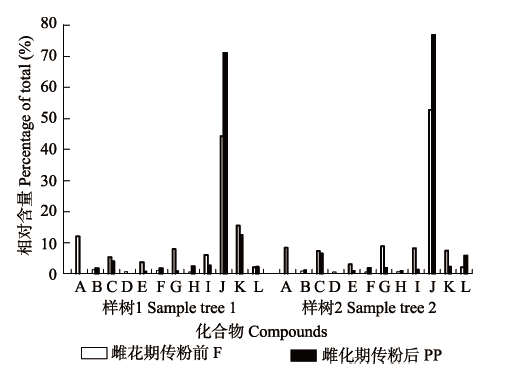 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT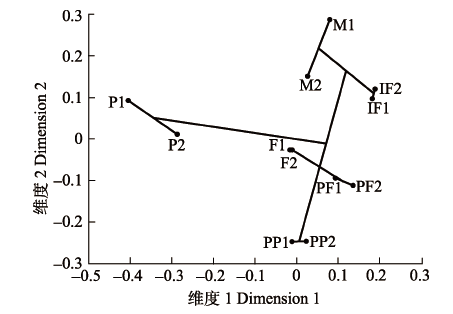 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT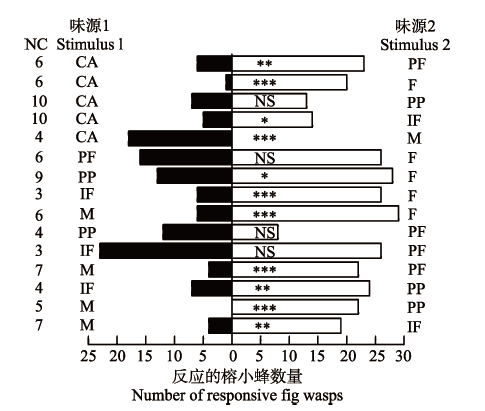 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}