关键词:树干CO2释放通量;季节变化;种间差异;驱动因子;温带森林;温度 Abstract Aims Stem CO2 efflux (Es) is an important component of annual carbon budget in forest ecosystems, but how biotic and environmental factors regulate seasonal and inter-specific variations in Es is poorly understood. The objectives of this study were: (1) to compare seasonal dynamics in Es for four temperate coniferous tree species in northeastern China, including Korean pine (Pinus koraiensis), Korean spruce (Picea koraiensis), Mongolian pine (Pinus sylvestris var. mongolica), and Dahurian larch (Larix gmelinii); and (2) to explore factors driving the inter-specific variability in Es during the growing and non-growing seasons. Methods Ten to twelve trees for each tree species were sampled for Es and stem temperature at 1 cm depth beneath the bark (Ts) measurements in situ with an infrared gas analyzer (LI-6400 IRGA) and a digital thermometer, respectively, from July to October 2013 and March to July 2014. The daily stem circumference increment (Si), sapwood nitrogen concentration ([N]), and related environmental factors were monitored simultaneously. Important findings The temporal variation in Es for the four tree species overall followed the changes in Ts throughout the study period, with the maxima occurring in the summer months (late May to early July) characterized by higher temperature and more rapid stem growth and the minima in spring (late March to April) or autumn (October) having lower temperature. Ts accounted for 42%-91% and 56%-89% of variations in Es during the growing (May to September) and non-growing (other months) seasons, respectively. Furthermore, apart from Ts, we also found significant regression relationships between Es and Si, relative air humidity and [N] during the growing season, but their forms and correlation coefficients were species-dependent. These results indicated that Ts was the dominant environmental factor affecting seasonal variations in Es, but the magnitude of the effect varied with tree species and growth rhythm. Mean Es for each of the four tree species was significantly higher in the growing season than in the non-growing season, whereas within the season there were also significant differences in mean Es among the tree species (all p < 0.05). The temperature sensitivity of Es (Q10 value) did not differ significantly among the tree species during the growing season, ranging from 1.64 for Dahurian larch to 2.09 for Mongolian pine, but did differ during the non-growing season which varied from 1.80 for Korean pine to 3.14 for Dahurian larch. Moreover, Korean spruce, Mongolian pine and Dahurian larch had significantly greater Q10 values in the non-growing season than in the growing season (p < 0.05). These findings suggested that the differences of the response of Es to temperature change for different tree species were mainly from the non-growing season. Because the seasonality and inter-specific variability in Es for these temperate coniferous tree species were primarily controlled by multiple factors such as temperature, we conclude that using a single annual temperature response curve to estimate the annual Es may lead to more uncertainty.
研究地设在黑龙江帽儿山森林生态站(45.40° N, 127.67° E)。平均海拔400 m, 平均坡度3°-15°, 地带性土壤为暗棕色森林土。该地区气候属大陆性季风气候, 夏季短促湿热, 冬季寒冷干燥。1989-2009年 平均年降水量和潜在蒸发量分别为629 mm和 864 mm; 年平均气温、1月份和7月份的平均气温分别为3.1 ℃、-18.5 ℃和22.0 ℃; 无霜期为120-140天。现有植被是原地带性植被——阔叶红松林经干扰后形成的天然次生林和人工林, 代表着东北东部山区典型的森林类型。有关林分特征和立地状况参见文献(Wang et al., 2006; Zhang et al., 2014)。 在红松、红皮云杉、樟子松和落叶松等4个冠层已经郁闭的人工林内, 根据林分内林木的径级分布情况, 按照均匀取样的原则, 每个树种随机选取生长良好的样树10-12株(基本涵盖本地区各树种的最大径级), 以进行Es测定, 其基本属性见表1。 Table 1 表1 表1测定样树的基本特征 Table 1Basic characteristics of the sampled trees
树种 Species
叶性状 Leaf trait
样本数 Sample size
胸径 Diameter at breast height (cm)
边材体积 Sapwood volume (cm3)
范围 Range
平均值±标准误差 Mean ± SE
范围 Range
平均值±标准误差 Mean ± SE
红松 Pinus koraiensis
常绿 Evergreen
11
8.0-30.1
18.7 ± 2.2b
35.7-297.5
115.5 ± 26.2b
红皮云杉 Picea koraiensis
常绿 Evergreen
10
17.2-43.4
29.1 ± 2.9a
115.1-389.0
285.7 ± 30.5a
樟子松 Pinus sylvestris var. mongolica
常绿 Evergreen
11
18.9-34.3
25.6 ± 1.6ab
187.2-381.3
287.4 ± 17.7a
落叶松 Larix gmelinii
落叶 Deciduous
12
11.7-46.4
28.2 ± 3.2a
34.2-208.6
131.2 ± 15.6b
a and b in the table represent the significant difference group at α = 0.05 level.a和b表示显著性差异组别(α = 0.05)。 新窗口打开
1.2 Es及其影响因子测定
采用红外气体分析法原位测定Es (Xu et al., 2000)。实验开始前, 先在样树树干北面的胸径(DBH, 约离地1.3 m)处安装高5.0-6.0 cm、内径10.4 cm的可连接LI-6400便携式CO2/H2O分析仪(LI-COR, Lincoln, USA) 6400-09同化室的PVC呼吸环。将呼吸环的一端打磨成与树干表面相吻合的弧形, 再利用100%中性硅酮防水胶将其固定在已除去松脱的树皮等表面杂物的树干表面上, 并同步测定呼吸环所覆盖的树干表面积; 呼吸环完全固定后, 采用注水法测定其体积, 用于计算Es。整个测定期间呼吸环位置保持不变, 以便连续测定。 2013年7-10月和2014年3-7月, 采用LI-6400便携式CO2/H2O分析系统(IRGA, LI-COR, Lincoln, USA)几乎每月测定1次Es的昼间动态。测定从日出(6:00)开始, 日落(19:00)结束, 每2 h测定1次, 连续测定5-7次Es。限于人力、仪器和样地距离, 每天只能测定一个树种的昼间变化; 而且在某些月份, 频繁的降雨(图1)导致个别树种未能进行昼间动态测定, 造成数据缺失, 如落叶松缺少2013年8月和2014年5月的数据。测定前, 检查并确保呼吸环不漏气; 测定时, 将LI-6400-09同化室扣在呼吸环上, 采用自动测定设置, 每5 s (生长季)或10 s (非生长季)记录一个CO2浓度值、连续取样2 min, 然后根据同化室中CO2浓度随时间的变化曲线计算Es。同步利用连接在IRGA上的温度计测定呼吸环上方约3 cm处、树皮下约1 cm深处的树干温度(Ts, ℃)。 显示原图|下载原图ZIP|生成PPT 图12013年7月1日至2014年7月20日日降水总量(Pre)、日平均气温(Ta)和空气相对湿度(RH)的季节动态。 -->Fig. 1Seasonal changes in daily sums of precipitation (Pre) and daily means of air temperature (Ta) and relative air humidity (RH) between July 1, 2013 and July 20, 2014. -->
采用下式计算Es (许飞和王传宽, 2015): $\begin{align} & {{E}_{\text{s}}}\text{ =}\frac{\partial C}{\partial t}\times \frac{V+{V}'-\pi {{(d/2)}^{2}}h}{S}\times \frac{\text{10}}{R}\times \frac{P}{T+273.2}\times \\ & \ \ \ \ \ \ \ \left( 1-\frac{W}{1000} \right) \\ \end{align}$ (1) 式中: Es为单位树干表面积的树干CO2释放通量(μmol·m-2·s-1), $\frac{\partial C}{\partial t}$为采样时气体浓度随时间变化的直线斜率(μmol·mol-1·s-1), V为呼吸环的实际体积(cm3), V′为系统默认的同化室体积(991.0 cm3), S为呼吸环所覆盖的树干表面积(cm2), d和h分别为插入呼吸环的同化室的外径和深度(cm), R为气体常数(8.3 Pa·m3·K-1·mol-1), P为采样点的大气压(kPa), T为同化室内气温(℃), W为同化室内水浓度(mmol·mol-1)。 为深入了解调控年内Es季节变化的主要的生物和非生物因子, 我们将测定时期按照物候和树木生长(图2)情况划分为生长季(5-9月)和非生长季(3、4、10月)。采用指数模型拟合生长季和非生长季Es与Ts之间的关系: Es = α eβTs (2) 式中: α和β为回归系数。通过下式确定Es的Q10值: Q10 = e10β (3) 采用独立样本t检验比较同一树种生长季和非生长季之间的平均Es、Ts和[N]的差异; 采用单因素方差分析(ANOVA)和Duncan多重比较检验(方差齐性)或者独立样本Kruskal-Wallis检验(方差不齐性)比较同一季节不同树种之间的平均Es、Ts、[N]、年胸围生长量和Q10值的差异; 采用配对样本t检验比较同一树种生长季和非生长季之间的Q10值的差异。采用回归分析法建立Es与生物和环境因子之间关系的模型。所有数据均采用SPSS统计软件进行分析。 显示原图|下载原图ZIP|生成PPT 图22013年5月10日至2014年7月3日4个树种的树干CO2释放通量(Es) (A)、树干温度(Ts) (B)、日胸围生长量(Si) (C)和边材氮浓度([N]) (D)的季节动态(平均值±标准误差; n = 6-12)。Es和Ts是一天内测定5-7次10-12株树的平均值。 -->Fig. 2Seasonal changes in stem CO2 efflux (Es) (A), stem temperature (Ts) (B), daily stem circumference increment (Si) (C), and sapwood nitrogen concentration ([N]) (D) for the four tree species (mean ± SE; n = 6-12) between May 10, 2013 and July 3, 2014. The values of Es or Ts are means of 10 to 12 trees measured five to seven times a day. -->
2 结果
2.1 Es与生物和非生物因子的季节动态
2013年7月至2014年6月, 降水主要集中在生长季的5月份(132.8 mm)、7月份(312.8 mm)和8月份(166.4 mm), 占全年降水总量(860.8 mm)的71%; 其余月份的降水量均小于80 mm。日平均RH波动在31.7% (2014年4月15日)-95.4% (2013年10月24日)之间, 频繁的降水导致日平均RH维持在较高的水平(平均78%)。日平均Ta从春季开始迅速增加, 到夏季达到最大值(24.5 ℃), 于秋冬季节下降到年最小值(-30.2 ℃) (图1)。 整个测定期间4个树种Es的年内季节(四季)变化大体上与Ts的变化趋势一致, 但变化格局因树种而异(图2A、2B)。2013年7-10月, 随着Ts的快速下降, 4个树种的Es也迅速由生长季的峰值状态减小到非生长季的维持代谢程度。至次年早春(2014年3月末至4月初), 红松、红皮云杉和樟子松的Ts与2013年10月基本持平, 其Es也几乎维持在2013年秋季的水平(数值间无显著差异); 而落叶松由于Ts的大幅下降, 导致Es亦显著低于2013年秋季的代谢量。之后, 随着Ts的升高, Es也快速增加: 红松的Es和Ts在5月末同时达到高峰值; 红皮云杉和落叶松的Es在7月初再次达到高峰值, 但滞后Ts峰值1个月; 樟子松的Es和Ts到5月末均略微有所下降, 随后同步持续增加到7月初的高峰值。实验期间红松、红皮云杉、樟子松和落叶松的Es分别波动在(0.46 ± 0.07) - (1.62 ± 0.27)(平均值±标准误差; 下同)、(0.81 ± 0.05) - (3.21 ± 0.29)、(0.33 ± 0.02) - (2.10 ± 0.22)和(0.23 ± 0.03) - (3.82 ± 0.49) μmol·m-2·s-1之间; 对应的年平均值分别为(1.06 ± 0.19)、(1.94 ± 0.18)、(1.29 ± 0.13)和(1.98 ± 0.25) μmol·m-2·s-1, 生长季平均值分别为(1.29 ± 0.24)、(2.39 ± 0.23)、(1.58 ± 0.17)和(2.76 ± 0.35) μmol·m-2·s-1, 非生长季平均值分别为(0.49 ± 0.06)、(0.83 ± 0.06)、(0.73 ± 0.06)和(0.42 ± 0.05) μmol·m-2·s-1。ANOVA分析表明, 各树种生长季的平均Es和Ts均显著高于非生长季, 且同一季节(生长季或非生长季)不同树种间的平均Es和Ts也存在显著差异(图3A、3B)。生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松, 而非生长季则是红皮云杉和樟子松显著高于红松和落叶松。但是, 同一季节不同树种间平均Ts和Es的差异并不一致。生长季和非生长季4个树种的平均Ts间均存在显著差异, 其大小排序分别为: 红松>落叶松>红皮云 杉>樟子松和樟子松>红松>红皮云杉>落叶松。 显示原图|下载原图ZIP|生成PPT 图3不同季节4个树种树干CO2释放通量(Es) (A)、树干温度(Ts) (B)、年胸围生长量(Gi) (C)和边材氮浓度([N]) (D)的均值比较(平均值±标准误差; n = 6-12)。不同小写字母(a-d)表示同一季节不同树种间的平均Es、Ts、Gi和[N]存在显著差异(p < 0.05), 星号(*)表示同一树种不同季节间的均值存在显著差异(p < 0.05)。 -->Fig. 3Comparisons of mean values of stem CO2 efflux (Es) (A), stem temperature (Ts) (B), annual stem circumference increment (Gi) (C), and sapwood nitrogen concentration ([N]) (D) between the growing and non-growing seasons for the four tree species (mean ± SE; n = 6-12). Different lowercase letters (a-d) stand for significant differences (p < 0.05) in mean values of Es, Ts, Gi, and [N] among different species in the same season, while stars (*) represent significant differences (p < 0.05) in those of Es, Ts, and [N] between the two seasons for the same species. -->
生长季和非生长季4个树种的Es与Ts之间均存在显著的指数关系(p < 0.01), 但是Ts对Es变异的解释率(即决定系数R2)随树种和季节而变化(图4)。生长季各树种Es与Ts关系的R2以红松(0.91)最高、落叶松最低(0.42), 而非生长季的R2则波动在0.56 (红松) -0.89 (樟子松)之间。 显示原图|下载原图ZIP|生成PPT 图4不同季节4个树种树干CO2释放通量(Es)与树干温度(Ts)的关系。每个点表示10-12株样木的平均值。 -->Fig. 4Relationships between stem CO2 efflux (Es) and stem temperature (Ts) for the four tree species during the growing and non-growing seasons. Each point is the mean of 10 to 12 trees. -->
4个树种生长季和非生长季的Q10值分别在1.64 (落叶松) - 2.09 (樟子松)和1.80 (红松) - 3.14 (落叶松)之间波动(图5)。所有树种的Q10值均呈现出显著的季节变化。红皮云杉、樟子松和落叶松生长季的Q10值均显著低于非生长季, 而红松则展现出相反的规律, 其生长季的Q10值显著高于非生长季。不同树种间Q10值的差异也随季节而变化。生长季红松和樟子松(二者差异不显著)的Q10值显著高于落叶松, 而红皮云杉与它们之间的差异均不显著; 非生长季4个树种的Q10值间都存在显著差异, 大小排序为: 落叶松>樟子松>红皮云杉>红松。 显示原图|下载原图ZIP|生成PPT 图5不同季节4个树种树干CO2释放通量的温度系数(Q10)比较(平均值±标准误差; n = 10-12)。不同小写字母(a-d)表示同一季节不同树种间的Q10值存在显著差异(p < 0.05), 星号(*)表示同一树种不同季节间的Q10值存在显著差异(p < 0.05)。 -->Fig. 5Comparisons of temperature sensitivity (Q10) of stem CO2 efflux between the growing and non-growing seasons for the four tree species (mean ± SE; n = 10-12). Different lowercase letters (a-d) stand for significant differences (p < 0.05) in Q10 values among different species in the same season, while stars (*) represent significant differences (p < 0.05) in Q10 values between the two seasons for the same species. -->
此外, 生长季期间4个树种的平均Es与Si、RH或[N]间亦存在相关关系, 但其相关程度和显著性因树种而异(图6)。所有树种的平均Es与Si间均呈显著的线性正相关关系(p < 0.05, 其中红松在p < 0.10显著, 樟子松去掉2014年5月末的最大值; 图6A)。樟子松和落叶松的平均Es与RH间则分别呈现显著的二次和线性函数(p = 0.01)关系(图6B)。樟子松的平均Es与[N]间也显著线性正相关(去掉2013年9月中旬的测定值: p = 0.03; 图6C), 而其他3个树种均不存在明显的函数关系。 显示原图|下载原图ZIP|生成PPT 图6生长季4个树种树干CO2释放通量(Es)与日胸围生长量(Si) (A)、空气相对湿度(RH) (B)和边材氮浓度([N]) (C)的关系(平均值±标准误差; n = 6-12)。黑色三角(▼)表示樟子松2014年5月末的最大Si (A)和2013年9月中旬生长缓慢时的[N] (C), 二者在拟合模型时去除。 -->Fig. 6Relationships of stem CO2 efflux (Es) with daily stem circumference increment (Si) (A), air relative humidity (RH) (B), and sapwood nitrogen concentration ([N]) (C) during the growing season for the four tree species (mean ± SE; n = 6-12). The black triangles (▼) indicate the maximum Si at the end of May 2014 (A) and [N] at the time of slow growth in mid-September 2013 (C) for Pinus sylvestris var. mongolica, both of which are excluded when fitting models. -->
3 讨论
3.1 Es的季节动态及其驱动因子
4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性。本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015)。综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的。 然而, 温度在解释4个树种生长季Es动态变化时还存在局限性(如落叶松: R2 = 0.42)(图2, 图4), 说明除了温度以外, 还有其他的生物(树干生长、[N]等)或非生物(空气湿度等)因子在调控其变化。树干生长不仅与Es组分之一的生长呼吸紧密相关(Ryan, 1990; Zha et al., 2004), 而且还能通过影响树干活细胞量(数量或体积)而影响Es的另一组分维持呼吸(Stockfors & Linder, 1998; Gruber et al., 2009), 因此可能是造成生长季Es变化的重要因子。本研究发现生长季4个树种Es的变化格局大体上表现出与树干生长(Si)一致的变化趋势(图2A、2C), 而且各树种的Es与Si间均存在显著的线性相关关系(图6A)。其他研究也报道了类似的结果。Maier (2001)发现火炬松(Pinus taeda)的标准化Es (即校正到某一温度下的Es)与树干生长具有相似的季节动态, 并且树干生长能够解释标准化Es季节变异的51%。日本扁柏(Chamaecyparis obtusa)日总Es的季节动态与日直径生长显著相关, 而且采用Ts和日直径生长的综合模型能更好地解释Es的季节变化(Araki et al., 2015)。所以, 树干生长是引起4个针叶树种生长季Es波动的一个重要因素。 但是, 我们也发现个别树种的Es与树干生长在某些情况下的匹配性较差。例如, 红松和樟子松的Es峰值出现时间滞后树干生长峰值20-35天; 樟子松在2014年5月末伴随着Ts和[N]的下降、Es同步下降, 而此时树干生长却处于峰值(图2A、2C)。这些现象说明: 一方面, 由于木材生产总是发生在细胞扩增之后(Sprugel & Benecke, 1991), 所以Es峰值与树干生长峰值之间很可能存在一定的滞后期。因此, 有必要采用时间分辨率更高的测定(如日测定)来匹配生长季Es与树干生长的变化(Ceschia et al., 2002; Brito et al., 2010), 将树干生长纳入Es模型, 提高季节内或年内季节性Es的估测精度。另一方面, 其他因子(如Ts和[N]等)对Es的强烈影响可能掩盖或者超过了树干生长对Es的作用。樟子松生长季Es与[N]近乎平行的动态变化(图2A、2D)以及二者间较强的相关关系(图6C)表明[N]对樟子松Es季节内变化有显著的影响, 而且这种影响可能独立于温度, 并超过树干生长的影响。Maier (2001)发现[N]解释了火炬松标准化Es季节动态24%的变异, 并指出[N]的时间变异能影响不依赖温度的Es的季节变化。 目前明确提出空气湿度对Es季节变化有影响的研究比较少见。本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素。Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es。有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响。随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008)。限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015)。(2) RH对树干生长的影响。RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加。落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点。同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应。(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003)。这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B)。我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小。综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种。 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4)。但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998)。本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991))。Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系。此外, 测定时期也可能影响非生长季Es对温度的响应。树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004)。本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用。这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力。 其他因子也可能影响Es的季节(内)变化。一是碳水化合物供应。环割(Maier et al., 2010)和同位素示踪实验(Wertin & Teskey, 2008)均指出, 新近固定的光合产物的转运能够改变树干的底物浓度, 进而影响Es的动态变化。二是树干水分状况及其内部结构。树干含水量和树干内部结构(木质部、形成层和树皮层)及其引起的树干径向CO2扩散阻碍的动态变化可引起Es的时间变异。Sorz和Hietz (2006)报道较高的树干含水量与内部较低的O2含量相对应, 因此会极大地抑制径向气体交换速率; 而O2在充满饱和水的木质部中的扩散系数总是低于在水中的扩散系数, 则说明木质部细胞壁是阻碍气体扩散的一个主要障碍。第三, 树干皮层光合作用能够通过固定树干内活细胞呼吸释放或者蒸腾流中运输的CO2的方式来减小Es (Wittmann et al., 2006), 因此也可能造成Es的季节性波动。本研究中4个针叶林林分已经郁闭, 到达地表的光照较少; 而且即使有光照能够穿透冠层直接照射树干表面, 因为我们测定所采用的呼吸环是不透光的PVC材质, 且Es测定部位及其周边部位的树皮或致密或较厚, 因此可以忽略树干光合作用对被测树种Es的影响。
3.2 Es的季节和种间差异
所有树种生长季的平均Es均显著高于非生长季(图3), 与其他温带和北方地区的树种的研究结果相符(Zha et al., 2004; Yang et al., 2012a), 表明生长季的代谢活动要高于非生长季。季节间Es的差异主要源于底物供应、温度、树干生长等影响因子的差异。相较于仅以维持呼吸为主的非生长季, 本研究地的生长季雨热同期(图1), 底物充足, 温度较高(图2B), 树木生长活动较快(图2C), 生理代谢活动增强, 因此此时期不仅需要较多的呼吸能量用于维持自身正常的生命活动(维持呼吸), 而且需要更多的能量和呼吸产物以构建自身生长所需的碳骨架等物质(生长呼吸)(Zha et al., 2004), 因而Es较高。 同一季节Es的种间差异显著(图3A), 但是引起种间差异的原因比较复杂。温度是造成生长季或非生长季4个树种Es种间差异的主要因素, 因为Ts解释了各树种季节内Es大多数的变异(>55%; 图4), 并且生长季4个树种的Es对温度几乎具有同等的敏感性(Q10值; 图5)。但是生长季或非生长季被测树种间平均Es与Ts的差异不一致(图3B), 说明温度并不是导致同一季节种间Es差异的唯一因素, 其他因素也在同时影响各树种的Es。 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关。红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者。樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D)。[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998)。本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es。有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007)。本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示。红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007)。 但在非生长季, 4个树种间Es差异的规律略有不同。红皮云杉和樟子松的平均Es均显著高于红松和落叶松(图3A), 这可能是红皮云杉和樟子松边材中活细胞比例及其生理活性、非结构性碳水化合物数量(Ceschia et al., 2002; Rodríguez-Calcerrada et al., 2015)等较高的缘故。红皮云杉和樟子松的边材体积(表征活细胞量)均显著高于红松和落叶松(表1), 而边材体积与维持呼吸呈显著正相关关系(Ryan et al., 1995)。Rodríguez-Calcerrada等(2015)总结边材的非结构性碳水化合物浓度是引起秋季Es (维持呼吸)种间差异的主要因素。其他影响因素可能还包括[N], 需要进一步研究。 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b)。由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003)。Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节。类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990)。但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5)。其他地区的研究亦报道了相似的结果。生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010)。导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010)。
4 结论
4种温带针叶树种的Es具有明显而相似的季节动态格局, 高峰值均出现在生长旺盛的夏季。Ts是驱动生长季和非生长季4个树种Es变化的主导环境因子, 但是树干生长、边材氮浓度、空气湿度等因子也不同程度地影响生长季Es的动态变化, 而其影响程度因树种而异。不同树种、不同季节的Es及其Q10值差异显著。如果忽略上述Es的季节和种间变化, 将会增大本地区Es年通量估测的不确定性。因此, 为了提高本地区乃至区域森林碳收支模型的估测精度, 未来研究需要综合且深入地探明各树种不同季节的Es对各生物和环境因子动态变化的响应, 建立简单、实用、高效且具有生物学意义的Es模拟预测模型。 致谢 感谢帽儿山森林生态站提供野外基础支持。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
AmthorJS (1991). Respiration in a future, higher-CO2 world .Plant, Cell & Environment, 14, 13-20. [本文引用: 1]
[2]
ArakiMG, KajimotoT, HanQM, KawasakiT, UtsugiH, GyokusenK, ChibaY (2015). Effect of stem radial growth on seasonal and spatial variations in stem CO2 efflux of Chamaecyparis obtusa .Trees, 29, 499-514. [本文引用: 7]
[3]
ArakiMG, UtsugiH, KajimotoT, HanQM, KawasakiT, ChibaY (2010). Estimation of whole-stem respiration, incorporating vertical and seasonal variations in stem CO2 efflux rate, of Chamaecyparis obtusa trees .Journal of Forest Research, 15, 115-122. [本文引用: 1]
[4]
AtkinOK, TjoelkerMG (2003). Thermal acclimation and the dynamic response of plant respiration to temperature .TRENDS in Plant Science, 8, 343-351. [本文引用: 1]
[5]
BowmanWP, BarbourMM, TurnbullMH, TissueDT, WhiteheadD, GriffinKL (2005). Sap flow rates and sapwood density are critical factors in within- and between-tree variation in CO2 efflux from stems of mature Dacrydium cupressinum trees .New Phytologist, 167, 815-828. [本文引用: 2]
[6]
BritoP, MoralesD, WieserG, JiménezMS (2010). Spatial and seasonal variations in stem CO2 efflux of Pinus canariensis at their upper distribution limit .Trees, 24, 523-531. [本文引用: 10]
[7]
CavaleriMA, OberbauerSF, RyanMG (2006). Wood CO2 efflux in a primary tropical rain forest .Global Change Biology, 12, 2442-2458. [本文引用: 3]
[8]
CeschiaÉ, DamesinC, LebaubeS, PontaillerJ-Y, DufrêneÉ (2002). Spatial and seasonal variations in stem respiration of beech trees (Fagus sylvatica) .Annals of Forest Science, 59, 801-812. [本文引用: 3]
[9]
GruberA, WieserG, OberhuberW (2009). Intra-annual dynamics of stem CO2 efflux in relation to cambial activity and xylem development in Pinus cembra .Tree Physiology, 29, 641-649. [本文引用: 5]
[10]
JiangLF, ShiFC, ZuYG, WangWJ, KoikeT (2003). Study on stem respiration of Larix gmelinii of different ages and its relationship to environmental factors .Bulletin of Botanical Research, 23, 296-301. [本文引用: 1]
[11]
JosephG, KelseyRG (2004). Ethanol synthesis and aerobic respiration in the laboratory by leader segments of Douglas-fir seedlings from winter and spring .Journal of Experimental Botany, 55, 1095-1103. [本文引用: 1]
[12]
KatayamaA, KumeT, OhashiM, MatsumotoK, NakagawaM, SaitoT, KumagaiT, OtsukiK (2016). Characteristics of wood CO2 efflux in a Bornean tropical rainforest .Agricultural and Forest Meteorology, 220, 190-199. [本文引用: 1]
[13]
KöcherP, HornaV, LeuschnerC (2012). Environmental control of daily stem growth patterns in five temperate broad-leaved tree species .Tree Physiology, 32, 1021-1032. [本文引用: 1]
[14]
LavigneMB, LittleCHA, RidingRT (2004). Changes in stem respiration rate during cambial reactivation can be used to refine estimates of growth and maintenance respiration .New Phytologist, 162, 81-93. [本文引用: 1]
[15]
LavigneMB, RyanMG (1997). Growth and maintenance respiration rates of aspen, black spruce and jack pine stems at northern and southern BOREAS sites .Tree Physiology, 17, 543-551. [本文引用: 1]
[16]
MaierCA (2001). Stem growth and respiration in loblolly pine plantations differing in soil resource availability .Tree Physiology, 21, 1183-1193. [本文引用: 2]
[17]
MaierCA, AlbaughTJ, AllenHL, DoughertyPM (2004). Respiratory carbon use and carbon storage in mid-rotation loblolly pine (Pinus taeda L.) plantations: The effect of site resources on the stand carbon balance .Global Change Biology, 10, 1335-1350. [本文引用: 2]
[18]
MaierCA, JohnsenKH, ClintonBD, LudoviciKH (2010). Relationships between stem CO2 efflux, substrate supply, and growth in young loblolly pine trees .New Phytologist, 185, 502-513. [本文引用: 3]
[19]
MaierCA, ZarnochSJ, DoughertyPM (1998). Effects of temperature and tissue nitrogen on dormant season stem and branch maintenance respiration in a young loblolly pine (Pinus taeda) plantation .Tree Physiology, 18, 11-20. [本文引用: 3]
[20]
Rodríguez-CalcerradaJ, LópezR, SalomónR, GordalizaGG, Valbuena-CarabañaM, OleksynJ, GilL (2015). Stem CO2 efflux in six co-occurring tree species: Underlying factors and ecological implications .Plant, Cell & Environment, 38, 1104-1115. [本文引用: 2]
[21]
Rodríguez-CalcerradaJ, Martin-StPaulNK, LempereurM, OurcivalJ-M, del ReyMdC, JoffreR, RambalS (2014). Stem CO2 efflux and its contribution to ecosystem CO2 efflux decrease with drought in a Mediterranean forest stand .Agricultural and Forest Meteorology, 195, 61-72. [本文引用: 1]
[22]
RyanMG (1990). Growth and maintenance respiration in stems of Pinus contorta and Picea engelmannii .Canadian Journal of Forest Research, 20, 48-57. [本文引用: 3]
[23]
RyanMG (1991). Effects of climate change on plant respiration .Ecological Applications, 1, 157-167. [本文引用: 1]
[24]
RyanMG, GowerST, HubbardRM, WaringRH, GholzHL, Cropper Jr.WP, RunningSW (1995). Woody tissue maintenance respiration of four conifers in contrasting climates .Oecologia, 101, 133-140. [本文引用: 1]
[25]
RyanMG, LinderS, VoseJM, HubbardRM (1994). Dark respiration of pines .Ecological Bulletins, 43, 50-63. [本文引用: 3]
[26]
SaveynA, SteppeK, LemeurR (2007). Drought and the diurnal patterns of stem CO2 efflux and xylem CO2 concentration in young oak (Quercus robur) .Tree Physiology, 27, 365-374. [本文引用: 1]
[27]
SaveynA, SteppeK, McGuireMA, LemeurR, TeskeyRO (2008). Stem respiration and carbon dioxide efflux of young Populus deltoides trees in relation to temperature and xylem carbon dioxide concentration .Oecologia, 154, 637-649. [本文引用: 1]
[28]
ShiXL, WangCK, XuF, WangXC (2010). Temporal dynamics and influencing factors of stem respiration for four temperate tree species .Acta Ecologica Sinica, 30, 3994-4003. (in Chinese with English abstract)[石新立, 王传宽, 许飞, 王兴昌 (2010). 四个温带树种树干呼吸的时间动态及其影响因子 . 生态学报, 30, 3994-4003.] [本文引用: 1]
[29]
SorzJ, HietzP (2006). Gas diffusion through wood: Implications for oxygen supply .Trees, 20, 34-41. [本文引用: 1]
[30]
SprugelDG, BeneckeU (1991). Measuring woody-tissue respiration and photosynthesis . In: Lassoie JP, Hinckley TM eds. Techniques and Approaches in Forest Tree Ecophysiology. CRC Press, Boca Raton, USA. 329-355. [本文引用: 2]
[31]
StahlC, BurbanB, GoretJ-Y, BonalD (2011). Seasonal variations in stem CO2 efflux in the neotropical rainforest of French Guiana .Annals of Forest Science, 68, 771-782. [本文引用: 2]
[32]
SteppeK, SaveynA, McGuireMA, LemeurR, TeskeyRO (2007). Resistance to radial CO2 diffusion contributes to between-tree variation in CO2 efflux of Populus deltoides stems .Functional Plant Biology, 34, 785-792. [本文引用: 3]
[33]
StockforsJ, LinderS (1998). Effect of nitrogen on the seasonal course of growth and maintenance respiration in stems of Norway spruce trees .Tree Physiology, 18, 155-166. [本文引用: 3]
[34]
SunT, LiuRP, LiXH, MaoZJ, HuangW, DongLL (2015). Effects of simulated nitrogen deposition on stem respiration of Larix gmelinii Rupr .in Northeastern China. Acta Ecologica Sinica, 35, 3684-3691. (in Chinese with English abstract)[孙涛, 刘瑞鹏, 李兴欢, 毛子军, 黄唯, 董利利 (2015). 模拟氮沉降对东北地区兴安落叶松树干呼吸的影响 . 生态学报, 35, 3684-3691.] [本文引用: 2]
[35]
WangCK, YangJY (2005). Carbon dioxide fluxes from soil respiration and woody debris decomposition in boreal forests .Acta Ecologica Sinica, 25, 633-638. (in Chinese with English abstract)[王传宽, 杨金艳 (2005). 北方森林土壤呼吸和木质残体分解释放出的CO2通量 . 生态学报, 25, 633-638.] [本文引用: 1]
[36]
WangCK, YangJY, ZhangQZ (2006). Soil respiration in six temperate forests in China .Global Change Biology, 12, 2103-2114. [本文引用: 1]
[37]
WangM, WuYX, WuJL (2008). Stem respiration of dominant tree species in broad-leaved Korean pine mixed forest in Changbai Mountains .Chinese Journal of Applied Ecology, 19, 956-960. (in Chinese with English abstract)[王淼, 武耀祥, 武静莲 (2008). 长白山红松针阔叶混交林主要树种树干呼吸速率 . 应用生态学报, 19, 956-960.] [本文引用: 1]
[38]
WangWJ, YangFJ, ZuYG, WangHM, TakagiK, SasaK, KoikeT (2003). Stem respiration of a larch ( Larix gmelini) plantation in Northeast China .Acta Botanica Sinica, 45, 1387-1397. [本文引用: 1]
[39]
WangXW, MaoZJ, SunT, WuHJ (2011). Effects of temperature and sap flow velocity on CO2 efflux from stems of three tree species in spring and autumn in Northeast China .Acta Ecologica Sinica, 31, 3358-3367. (in Chinese with English abstract)[王秀伟, 毛子军, 孙涛, 吴海军 (2011). 春、秋季节树干温度和液流速度对东北3树种树干表面CO2释放通量的影响 . 生态学报, 31, 3358-3367.] [本文引用: 1]
[40]
WertinTM, TeskeyRO (2008). Close coupling of whole-plant respiration to net photosynthesis and carbohydrates .Tree Physiology, 28, 1831-1840. [本文引用: 1]
[41]
WittmannC, PfanzH, LoretoF, CentrittoM, PietriniF, AlessioG (2006). Stem CO2 release under illumination: Corticular photosynthesis, photorespiration or inhibition of mitochondrial respiration? Plant, Cell & Environment, 29, 1149-1158. [本文引用: 1]
[42]
XiaoFM, WangSL, DuTZ, ChenLC, YuXJ (2005). Respiration of Chinese fir in plantations in Huitong, Hu’nan Province .Acta Ecologica Sinica, 25, 2514-2519. (in Chinese with English abstract)[肖复明, 汪思龙, 杜天真, 陈龙池, 于小军 (2005). 湖南会同林区杉木人工林呼吸量测定 . 生态学报, 25, 2514-2519.] [本文引用: 1]
[43]
XuF, WangCK (2015). Diurnal dynamics in stem maintenance respiration in spring and autumn for four temperate coniferous tree species in northeastern China .Acta Ecologica Sinica, 35, 3233-3243. (in Chinese with English abstract)[许飞, 王传宽 (2015). 温带4种针叶树种春、秋季节树干维持呼吸的日动态 . 生态学报, 35, 3233-3243.] [本文引用: 4]
[44]
XuF, WangCK, WangXC (2011). Intra- and inter-specific variations in stem respiration for 14 temperate tree species in northeastern China .Acta Ecologica Sinica, 31, 3581-3589. (in Chinese with English abstract)[许飞, 王传宽, 王兴昌 (2011). 东北东部14个温带树种树干呼吸的种内种间变异 . 生态学报, 31, 3581-3589.] [本文引用: 2]
[45]
XuM, DeBiaseTA, QiY (2000). A simple technique to measure stem respiration using a horizontally oriented soil chamber .Canadian Journal of Forest Research, 30, 1555-1560. [本文引用: 1]
[46]
YanYP, ShaLQ, CaoM (2008). Stem respiration rates of dominant tree species in a tropical seasonal rain forest in Xishuangbanna, Yunnan, southwest China .Journal of Plant Ecology (Chinese Version), 32, 23-30. (in Chinese with English abstract)[严玉平, 沙丽清, 曹敏 (2008). 西双版纳热带季节雨林优势树种树干呼吸特征 . 植物生态学报, 32, 23-30.] [本文引用: 1]
[47]
YangJY, HeYJ, AubreyDP, ZhuangQL, TeskeyRO (2016). Global patterns and predictors of stem CO2 efflux in forest ecosystems .Global Change Biology, 22, 1433-1444. [本文引用: 1]
[48]
YangJY, TeskeyRO, WangCK (2012a). Stem CO2 efflux of ten species in temperate forests in Northeastern China .Trees, 26, 1225-1235. [本文引用: 4]
[49]
YangQP, XuM, ChiYG, ZhengYP, ShenRC, LiPX, DaiHT (2012b). Temporal and spatial variations of stem CO2 efflux of three species in subtropical China .Journal of Plant Ecology, 5, 229-237. [本文引用: 2]
[50]
ZabugaVF, ZabugaGA, Zinov'evaVP (1983). Ecological characteristics of respiratory CO2 exchange in the growing trunk of Scotch pine .Soviet Plant Physiology, 29, 934-939.
[51]
ZachA, HornaV, LeuschnerC, ZimmermannR (2010). Patterns of wood carbon dioxide efflux across a 2,000-m elevation transect in an Andean moist forest .Oecologia, 162, 127-137. [本文引用: 2]
[52]
ZhaTS, KellomäkiS, WangKY, RyyppöA, NiinistöS (2004). Seasonal and annual stem respiration of Scots pine trees under boreal conditions .Annals of Botany, 94, 889-896. [本文引用: 12]
[53]
ZhangHY, WangCK, WangXC (2014). Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species .Trees, 28, 77-89. [本文引用: 1]
Respiration in a future, higher-CO2 world 1 1991
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
Effect of stem radial growth on seasonal and spatial variations in stem CO2 efflux of Chamaecyparis obtusa 7 2015
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... ; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... ; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... ; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
Estimation of whole-stem respiration, incorporating vertical and seasonal variations in stem CO2 efflux rate, of Chamaecyparis obtusa trees 1 2010
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
Thermal acclimation and the dynamic response of plant respiration to temperature 1 2003
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Sap flow rates and sapwood density are critical factors in within- and between-tree variation in CO2 efflux from stems of mature Dacrydium cupressinum trees 2 2005
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
Spatial and seasonal variations in stem CO2 efflux of Pinus canariensis at their upper distribution limit 10 2010
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... ; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
... 值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
... ).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
... -温度关系(Brito et al., 2010). ...
Wood CO2 efflux in a primary tropical rain forest 3 2006
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Spatial and seasonal variations in stem respiration of beech trees (Fagus sylvatica) 3 2002
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 但在非生长季, 4个树种间Es差异的规律略有不同.红皮云杉和樟子松的平均Es均显著高于红松和落叶松(图3A), 这可能是红皮云杉和樟子松边材中活细胞比例及其生理活性、非结构性碳水化合物数量(Ceschia et al., 2002; Rodríguez-Calcerrada et al., 2015)等较高的缘故.红皮云杉和樟子松的边材体积(表征活细胞量)均显著高于红松和落叶松(表1), 而边材体积与维持呼吸呈显著正相关关系(Ryan et al., 1995).Rodríguez-Calcerrada等(2015)总结边材的非结构性碳水化合物浓度是引起秋季Es (维持呼吸)种间差异的主要因素.其他影响因素可能还包括[N], 需要进一步研究. ...
Intra-annual dynamics of stem CO2 efflux in relation to cambial activity and xylem development in Pinus cembra 5 2009
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
Study on stem respiration of Larix gmelinii of different ages and its relationship to environmental factors 1 2003
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
Ethanol synthesis and aerobic respiration in the laboratory by leader segments of Douglas-fir seedlings from winter and spring 1 2004
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Characteristics of wood CO2 efflux in a Bornean tropical rainforest 1 2016
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Environmental control of daily stem growth patterns in five temperate broad-leaved tree species 1 2012
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
Changes in stem respiration rate during cambial reactivation can be used to refine estimates of growth and maintenance respiration 1 2004
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
Growth and maintenance respiration rates of aspen, black spruce and jack pine stems at northern and southern BOREAS sites 1 1997
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Stem growth and respiration in loblolly pine plantations differing in soil resource availability 2 2001
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Respiratory carbon use and carbon storage in mid-rotation loblolly pine (Pinus taeda L.) plantations: The effect of site resources on the stand carbon balance 2 2004
... 树干CO2释放通量(Es)占树木自养呼吸的15%- 45% (Ryan et al., 1994; Maier et al., 2004; 肖复明等, 2005), 是树木和森林生态系统年碳平衡的重要组分.Es能消耗森林生态系统总初级生产力的8%-13%, 对森林生产力有重要的影响(Ryan et al., 1994; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
... ; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
Relationships between stem CO2 efflux, substrate supply, and growth in young loblolly pine trees 3 2010
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Effects of temperature and tissue nitrogen on dormant season stem and branch maintenance respiration in a young loblolly pine (Pinus taeda) plantation 3 1998
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
... ; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
Stem CO2 efflux in six co-occurring tree species: Underlying factors and ecological implications 2 2015
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 但在非生长季, 4个树种间Es差异的规律略有不同.红皮云杉和樟子松的平均Es均显著高于红松和落叶松(图3A), 这可能是红皮云杉和樟子松边材中活细胞比例及其生理活性、非结构性碳水化合物数量(Ceschia et al., 2002; Rodríguez-Calcerrada et al., 2015)等较高的缘故.红皮云杉和樟子松的边材体积(表征活细胞量)均显著高于红松和落叶松(表1), 而边材体积与维持呼吸呈显著正相关关系(Ryan et al., 1995).Rodríguez-Calcerrada等(2015)总结边材的非结构性碳水化合物浓度是引起秋季Es (维持呼吸)种间差异的主要因素.其他影响因素可能还包括[N], 需要进一步研究. ...
Stem CO2 efflux and its contribution to ecosystem CO2 efflux decrease with drought in a Mediterranean forest stand 1 2014
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
Growth and maintenance respiration in stems of Pinus contorta and Picea engelmannii 3 1990
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Effects of climate change on plant respiration 1 1991
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
Woody tissue maintenance respiration of four conifers in contrasting climates 1 1995
... 但在非生长季, 4个树种间Es差异的规律略有不同.红皮云杉和樟子松的平均Es均显著高于红松和落叶松(图3A), 这可能是红皮云杉和樟子松边材中活细胞比例及其生理活性、非结构性碳水化合物数量(Ceschia et al., 2002; Rodríguez-Calcerrada et al., 2015)等较高的缘故.红皮云杉和樟子松的边材体积(表征活细胞量)均显著高于红松和落叶松(表1), 而边材体积与维持呼吸呈显著正相关关系(Ryan et al., 1995).Rodríguez-Calcerrada等(2015)总结边材的非结构性碳水化合物浓度是引起秋季Es (维持呼吸)种间差异的主要因素.其他影响因素可能还包括[N], 需要进一步研究. ...
Dark respiration of pines 3 1994
... 树干CO2释放通量(Es)占树木自养呼吸的15%- 45% (Ryan et al., 1994; Maier et al., 2004; 肖复明等, 2005), 是树木和森林生态系统年碳平衡的重要组分.Es能消耗森林生态系统总初级生产力的8%-13%, 对森林生产力有重要的影响(Ryan et al., 1994; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
... 能消耗森林生态系统总初级生产力的8%-13%, 对森林生产力有重要的影响(Ryan et al., 1994; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Drought and the diurnal patterns of stem CO2 efflux and xylem CO2 concentration in young oak (Quercus robur) 1 2007
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Stem respiration and carbon dioxide efflux of young Populus deltoides trees in relation to temperature and xylem carbon dioxide concentration 1 2008
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
Seasonal variations in stem CO2 efflux in the neotropical rainforest of French Guiana 2 2011
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Resistance to radial CO2 diffusion contributes to between-tree variation in CO2 efflux of Populus deltoides stems 3 2007
... 目前明确提出空气湿度对Es季节变化有影响的研究比较少见.本研究发现樟子松和落叶松生长季的Es与RH间均存在显著的回归关系(图6B), 这说明空气湿度也是调控生长季Es动态变化的一个重要因素.Zabuga等(1983)报道欧洲赤松(Pinus sylvestris)的Es与RH也呈正相关关系, 而Jiang等(2003)发现较高的RH明显促进了生长季落叶松的Es.有关RH对Es影响机理的解释主要包括: (1) RH对液流的影响.随着RH增加, 树木蒸腾速率降低, 相应的液流速度也会下降, 结果导致Es增加(因为液流速度与Es显著负相关; Bowman et al., 2005; Saveyn et al., 2008).限于研究条件, 本研究未能同步测定样木的液流速度, 但有研究指出, 液流速度对季节性Es动态变化的影响较小(Gruber et al., 2009)或者无影响(Brito et al., 2010; Araki et al., 2015).(2) RH对树干生长的影响.RH增加引起蒸腾速率下降, 继而导致树木导水系统负压减小和树干形成层细胞膨压增加, 由此利于细胞分裂与膨胀, 引起树干生长(Köcher et al., 2012), 造成Es增加.落叶松树干生长与RH间的显著正相关关系(R2 = 0.91, p < 0.05; 数据未给出)部分支持以上论点.同时该论点也部分解释了落叶松生长季Es数据较为分散和温度对其变异解释率较低(图4)的原因: 树干生长和空气湿度对Es的混杂作用可能影响了Es对温度的响应.(3)高的RH (>70%)可能会促进树干表面死树皮的分解, 进而导致Es增强(Jiang et al., 2003).这个观点可能不被本实验结果所支持, 因为在RH超过70%以后, 樟子松的Es即随之下降(图6B).我们推测樟子松Es随高的RH下降的原因可能是: RH太高(>70%)的时候, 树木气孔关闭, 蒸腾速率下降, 这样很可能导致树干含水量过高, 进而引起树干内部的气体和水分含量的比例发生变化, 结果不仅增加了树干径向的扩散阻碍(Sorz & Hietz, 2006), 而且缩小了树干内部与大气间的CO2浓度梯度, 总体上不利于树干径向的气体扩散(Steppe et al., 2007), 导致Es减小.综合以上结果说明树干生长、[N]和空气湿度对于生长季Es的影响强度取决于树种. ...
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
... 交换(Steppe et al., 2007). ...
Effect of nitrogen on the seasonal course of growth and maintenance respiration in stems of Norway spruce trees 3 1998
... 与前人研究结果相符(Maier et al., 1998; Stockfors & Linder, 1998), 温度也是驱动本地区4个树种非生长季Es (相当于维持呼吸)动态变化的主导因子(图2, 图4).但是, 另有****报道, [N]亦与非生长季的Es密切相关(Ryan, 1991; Maier et al., 1998).本研究未发现非生长季Es与[N]间的耦合关系(动态变化不同; 图2), 可能是因为[N]取样树代表性不足(如种内变异较大; 图6C)、取样频率较低、或者这些林分下的土壤氮供应充足(因为在富氮生态系统中, 呼吸作用几乎不受氮有效性的影响(Amthor, 1991)).Lavigne和Ryan (1997)也发现几个北方树种非生长季的Es与[N]间没有任何关系.此外, 测定时期也可能影响非生长季Es对温度的响应.树干细胞超微结构和生物化学过程的改变会引起春季形成层再激活阶段的Es显著增加(Gruber et al., 2009); 而在秋冬季, 随着细胞分裂、木质部分化和抗寒锻炼的结束, 树木代谢活动减弱, 由此导致Es减小(Lavigne et al., 2004).本研究的非生长季数据即是这两个各异生理活性阶段——秋季和春季数据的组合, 因此有可能弱化温度对Es的作用.这些结果说明在非生长季的不同时段(如形成层的活动和休眠循环的过渡阶段、冬季沉寂期)控制Es的因子不同, 所以在构建基于过程的森林碳平衡模型时应考虑这些特殊时期(Gruber et al., 2009), 以提高模型预测能力. ...
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
模拟氮沉降对东北地区兴安落叶松树干呼吸的影响 2 2015
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 树干CO2释放通量(Es)占树木自养呼吸的15%- 45% (Ryan et al., 1994; Maier et al., 2004; 肖复明等, 2005), 是树木和森林生态系统年碳平衡的重要组分.Es能消耗森林生态系统总初级生产力的8%-13%, 对森林生产力有重要的影响(Ryan et al., 1994; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
温带4种针叶树种春、秋季节树干维持呼吸的日动态 4 2015
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
东北东部14个温带树种树干呼吸的种内种间变异 2 2011
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Global patterns and predictors of stem CO2 efflux in forest ecosystems 1 2016
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Stem CO2 efflux of ten species in temperate forests in Northeastern China 4 2012
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... 所有树种生长季的平均Es均显著高于非生长季(图3), 与其他温带和北方地区的树种的研究结果相符(Zha et al., 2004; Yang et al., 2012a), 表明生长季的代谢活动要高于非生长季.季节间Es的差异主要源于底物供应、温度、树干生长等影响因子的差异.相较于仅以维持呼吸为主的非生长季, 本研究地的生长季雨热同期(图1), 底物充足, 温度较高(图2B), 树木生长活动较快(图2C), 生理代谢活动增强, 因此此时期不仅需要较多的呼吸能量用于维持自身正常的生命活动(维持呼吸), 而且需要更多的能量和呼吸产物以构建自身生长所需的碳骨架等物质(生长呼吸)(Zha et al., 2004), 因而Es较高. ...
Temporal and spatial variations of stem CO2 efflux of three species in subtropical China 2 2012
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Ecological characteristics of respiratory CO2 exchange in the growing trunk of Scotch pine 1983
Patterns of wood carbon dioxide efflux across a 2,000-m elevation transect in an Andean moist forest 2 2010
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... )、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
Seasonal and annual stem respiration of Scots pine trees under boreal conditions 12 2004
... 树干CO2释放通量(Es)占树木自养呼吸的15%- 45% (Ryan et al., 1994; Maier et al., 2004; 肖复明等, 2005), 是树木和森林生态系统年碳平衡的重要组分.Es能消耗森林生态系统总初级生产力的8%-13%, 对森林生产力有重要的影响(Ryan et al., 1994; Maier et al., 2004; Zha et al., 2004).因此, 揭示Es的时空动态及其控制因子有助于正确评估Es在树木和森林年碳收支中的作用, 加深我们对森林碳循环过程的理解. ...
... Es存在季节变化已是毋庸置疑的事实.以往大量研究表明Es的季节动态随气候带、森林类型和立地特征而变化(Zha et al., 2004; Cavaleri et al., 2006; Brito et al., 2010; Araki et al., 2015).绝大多数温带和北方森林树木Es的经典季节变化模式为: 在温度高、树干生长快速的夏季达到高峰值, 在低温、树干生长停止的秋冬季减小到最小值(Lavigne & Ryan, 1997; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... ; Zha et al., 2004; Gruber et al., 2009; Yang et al., 2012a); 相反, 处于终年温暖、树干全年生长的热带地区树木的Es的季节动态格局更加复杂化, 比如干旱季节的Es低于(严玉平等, 2008; Stahl et al., 2011)或高于(Zach et al., 2010)湿润季节, 或者季节变化不明显(但有波动; Cavaleri et al., 2006).因此, 充分了解不同气候带下各树种Es的季节变异规律是准确估算该区域树木和林分水平Es的首要步骤.但是, 若要建立稳健实用高精度的Es模型则需要详细掌握驱动各森林树种Es时间变异的生物和环境因子的信息.温带和北方的Es研究表明, 温度是造成该区域树种Es季节性波动的主导因子.很多****发现Es的季节变化总体上追随温度的变化, 温度解释了Es季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 季节变化50%以上的变异(Zha et al., 2004; Brito et al., 2010; Araki et al., 2015).此外, 底物供应(Maier et al., 2010)、形成层活动(Gruber et al., 2009)、树干生长(Brito et al., 2010; Araki et al., 2015)、边材氮浓度(Maier, 2001; 孙涛等, 2015)、树干CO2透性(Joseph & Kelsey, 2004)等生物因子和土壤养分(Maier, 2001)、土壤含水量(许飞和王传宽, 2015)等环境因子也在一定程度上共同调节Es的季节变化.但对热带森林而言, 温度对Es季节变化的作用明显减弱(除严玉平等(2008)外).几个在热带低地或山地森林进行的研究指出, 温度对其森林树种Es季节变化的影响微乎其微, 而树种的物候(Cavaleri et al., 2006)和生长节律、液流速度(Katayama et al., 2016)、底物供应(Zach et al., 2010)、土壤含水量(Stahl et al., 2011)等生物或非生物因子才是主导该区域Es季节变化的主要因素.另外, 虽然目前有许多研究指出同一树种内不同树木个体间和同一树木个体内不同部位间的Es存在很大差异(Ceschia et al., 2002; Bowman et al., 2005; Yang et al., 2012b), 但是树种间的差异同样不可忽视.例如, 共存于同一气候区域下中国东北地区生长季(6-9月)的14个温带针阔叶树种间的Es差异可达2倍多(许飞等, 2011), 而西班牙中部地区非生长季(秋季)的6个温带阔叶树种的Es甚至可达到近3倍的差异(Rodríguez-Calcerrada et al., 2015).然而目前关于引起Es种间差异原因的探究实验比较少.Rodríguez-Calcerrada等(2015)报道边材的非结构性碳水化合物浓度是造成休眠季节6个阔叶树种Es种间变异的主要因子.许飞等(2011)指出树木生长节律、土壤养分状况和树皮的解剖结构等是引起生长季14个温带树种Es种间差异的重要因素.因此, 广泛而深入地研究不同气候带、不同地区各树种Es季节动态过程和种间差异以及相关的驱动因子, 全面了解各生物和环境因子对Es季节和种间变异的调控机理, 减少生态系统、区域乃至全球基于过程的森林碳平衡模型的不确定性(Yang et al., 2016), 提高其模拟和预测精度, 是当前迫切需要解决的问题. ...
... 4个针叶树种Es的年内季节变化格局总体上与温度(Ts (图2B)或Ta (图1))的变化格局相同, 在春季(3-4月)随温度升高而迅速增加, 于高温多雨(图1)的生长夏季(5月末-7月)达到峰值, 到秋冬季(9-10月)随温度的下降而减小至维持代谢水平(图2), 这与年内具有明显的生长和休眠季节动态特征的温带其他地区和北方地区的大多数针阔叶树种的规律一致(Zha et al., 2004; Yang et al., 2012a; Araki et al., 2015), 表明了温度对这些地区树木代谢活动潜在控制性影响的普遍性和树木生理活动调整对于温度变化的良好适应性.本研究显示Ts分别解释了4个树种生长季(除落叶松外)和非生长季Es 76%-91%和56%-89%的变异(图4), 说明温度是影响树木Es季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... 季节(内)变异最主要的环境因子, 这与许多前人的报道相符(Zha et al., 2004; Araki et al., 2015; 许飞和王传宽, 2015).综合以往研究, 这些结果共同表明把温度作为Es模型的主要预测参数来进行不同时间尺度(季节或年)下不同水平(呼吸环(Brito et al., 2010)、个体(Araki et al., 2010)或林分(Rodríguez-Calcerrada et al., 2014)的外推是合理的. ...
... 所有树种生长季的平均Es均显著高于非生长季(图3), 与其他温带和北方地区的树种的研究结果相符(Zha et al., 2004; Yang et al., 2012a), 表明生长季的代谢活动要高于非生长季.季节间Es的差异主要源于底物供应、温度、树干生长等影响因子的差异.相较于仅以维持呼吸为主的非生长季, 本研究地的生长季雨热同期(图1), 底物充足, 温度较高(图2B), 树木生长活动较快(图2C), 生理代谢活动增强, 因此此时期不仅需要较多的呼吸能量用于维持自身正常的生命活动(维持呼吸), 而且需要更多的能量和呼吸产物以构建自身生长所需的碳骨架等物质(生长呼吸)(Zha et al., 2004), 因而Es较高. ...
... ), 生理代谢活动增强, 因此此时期不仅需要较多的呼吸能量用于维持自身正常的生命活动(维持呼吸), 而且需要更多的能量和呼吸产物以构建自身生长所需的碳骨架等物质(生长呼吸)(Zha et al., 2004), 因而Es较高. ...
... 生长季红皮云杉和落叶松的平均Es均显著高于红松和樟子松(图3A), 这可能还与树干生长、[N]、径向CO2扩散阻碍等有关.红松的树干生长速率显著低于红皮云杉和落叶松(图3C), 因此与其生长密切相关的生长呼吸(Ryan, 1990; Zha et al., 2004)以及可能的维持呼吸(Sprugel & Benecke, 1991)也同样显著低于后者.樟子松[N]较低可能是其Es显著低于其他2个树种的一个原因(图3D).[N]与维持呼吸的相互关系已经被很多研究所证实(Maier et al., 1998; Stockfors & Linder, 1998).本研究虽然未发现红皮云杉和落叶松的[N]与Es间的相关关系, 但是[N]很有可能会通过影响其维持呼吸进而影响Es.有研究表明, 径向CO2扩散阻碍的差别也是引起个体(或树种)间Es差异的重要原因(Steppe et al., 2007).本研究未测定树干组织属性(如结构组成和生化指标), 但是从各树种可见的树皮属性差异可能得到一些启示.红皮云杉的树皮结构不如红松密实, 落叶松的树皮也比樟子松的更为松散, 所以红皮云杉和落叶松的树皮特性与红松和樟子松相比可能更易于树干和大气间的CO2交换(Steppe et al., 2007). ...
... 4个树种不同季节Es的Q10值(1.64-3.14; 图5)均处于前人报道的针叶树种的范围内(1.0-4.44; Ryan et al., 1994; Brito et al., 2010; Yang et al., 2012b).由于木本组织呼吸在低温时受最大酶活性控制、高温时受底物限制的缘故, 其Q10值通常随温度升高而降低(Atkin & Tjoelker, 2003).Maier (2001)报道火炬松Es对温度的敏感性在高温的夏季明显低于其他季节.类似的, 本研究中生长季红皮云杉(1.72 vs. 2.46)、樟子松(2.09 vs. 2.78)和落叶松(1.64 vs. 3.14)的Q10值均显著低于非生长季(图5), 表明生长季Es对温度的敏感性低于非生长季, 这与维持呼吸(非生长季)比生长呼吸(生长季)对温度变化更为敏感的论点一致(Ryan, 1990).但是我们也发现生长在阴坡的红松生长季的Q10值(2.05 vs. 1.80)显著高于非生长季(图5).其他地区的研究亦报道了相似的结果.生长在寒冷北方区域的欧洲赤松生长季的Q10值同样(1.89 vs. 1.79)显著高于非生长季(Zha et al., 2004); 而生长在半干燥地中海气候条件下的加那利松(Pinus canariensis)则呈现暖干季节的Q10值(3.89 vs. 2.17)显著高于冷湿季节的趋势(Brito et al., 2010).导致这种情况出现的原因尚不清楚, 推测可能与严酷的环境条件(如夏季干旱(Brito et al., 2010)或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
... )或寒冷(Zha et al., 2004)等)引起的碳水化合物的数量、组成和需求(Maier et al., 2010)、树干含水量(Saveyn et al., 2007)等的季节性变化有关, 从而掩盖了常见的Q10-温度关系(Brito et al., 2010). ...
Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species 1 2014
... 研究地设在黑龙江帽儿山森林生态站(45.40° N, 127.67° E).平均海拔400 m, 平均坡度3°-15°, 地带性土壤为暗棕色森林土.该地区气候属大陆性季风气候, 夏季短促湿热, 冬季寒冷干燥.1989-2009年 平均年降水量和潜在蒸发量分别为629 mm和 864 mm; 年平均气温、1月份和7月份的平均气温分别为3.1 ℃、-18.5 ℃和22.0 ℃; 无霜期为120-140天.现有植被是原地带性植被——阔叶红松林经干扰后形成的天然次生林和人工林, 代表着东北东部山区典型的森林类型.有关林分特征和立地状况参见文献(Wang et al., 2006; Zhang et al., 2014). ...

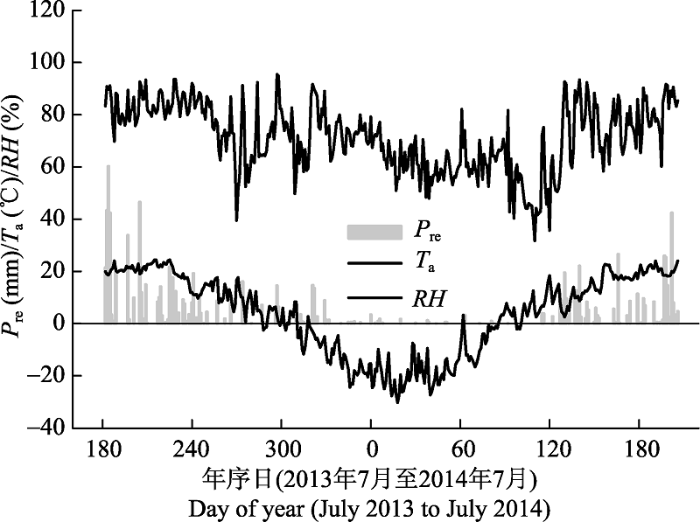 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT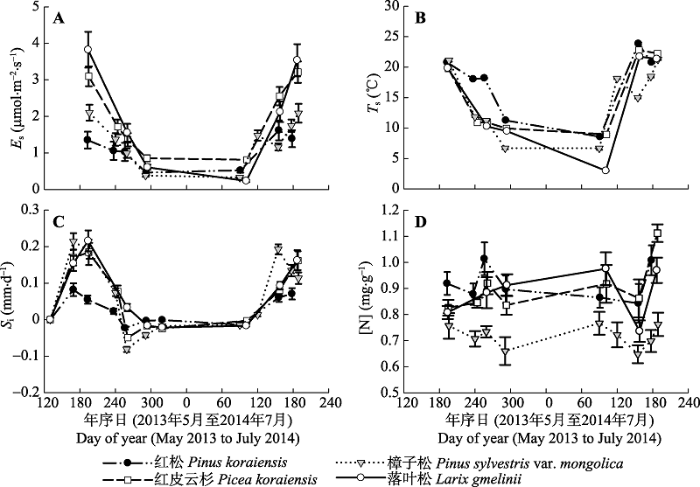 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT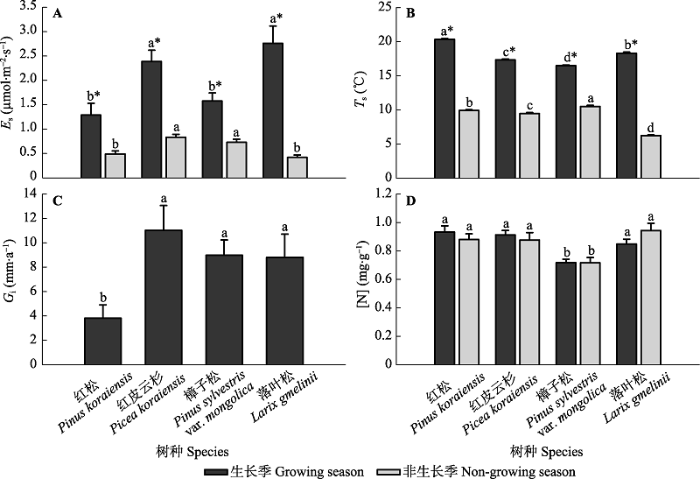 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT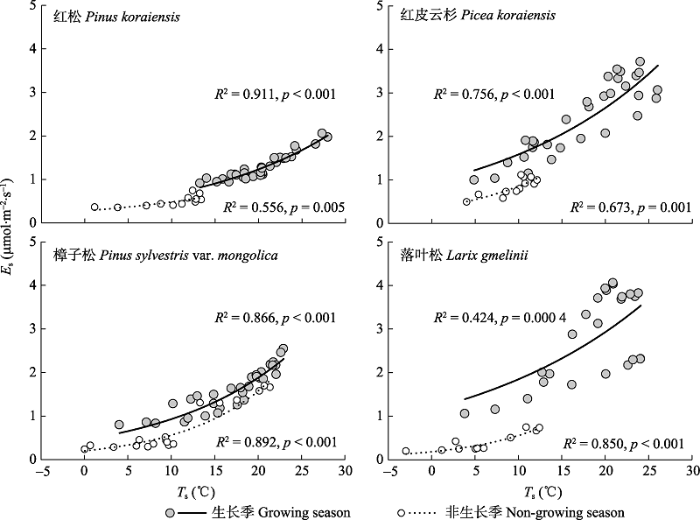 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT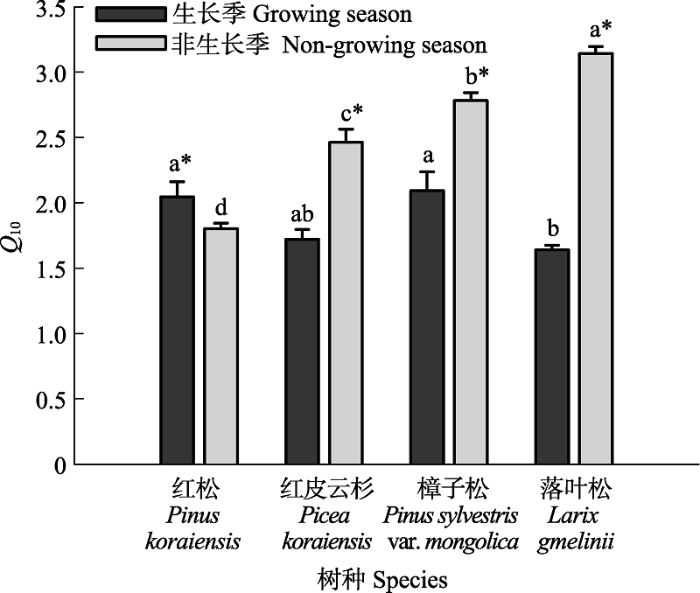 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT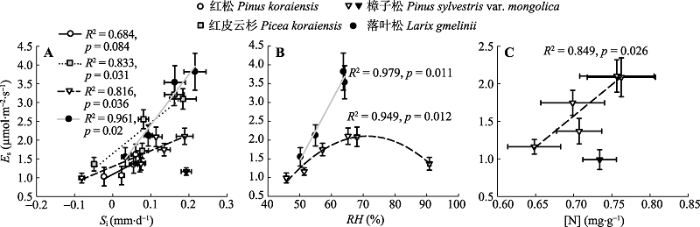 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}