Effects of nitrogen addition on photosynthetic characteristics of Leymus chinensis in the temperate grassland of Nei Mongol, China
ZHAIZhan-Wei1, GONGJi-Rui1,*,, LUOQin-Pu1, PANYan1, BAOYINTaogetao2, XUSha1, LIUMin1, YANGLi-Li1 1Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China2College of Life Sciences, Inner Mongolia University, Hohhot 010021, China 通讯作者:* 通信作者Author for correspondence (E-mail:jrgong@bnu.edu.cn) 收稿日期:2016-04-7 接受日期:2016-12-25 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41571048)、国家重点基础研究发展计划(973计划) (2014CB138803)和国家重点研发计划(2016YFC0500502)
关键词:氮添加;温带草原;羊草;光合特性;光合色素 Abstract Aims The increased atmospheric nitrogen (N) deposition due to human activity and climate change greatly causes grassland ecosystems shifting from being naturally N-limited to N-eutrophic or N-saturated, and further affecting the growth of grass species. The aims of this study are: 1) to evaluate the effects of different N addition levels on morphology and photosynthetic characteristics of Leymus chinensis; 2) to determine the critical N level to facilitate L. chinensis growth. Methods We conducted a different N addition levels experiment in dominant species in the temperate steppe of Nei Mongol. The aboveground biomass, morphological and leaf physiological traits, pigment contents, chlorophyll a fluorescence parameters and biochemical parameters of L. chinensis were investigated. Important findings Our results showed that aboveground biomass first increased and then decreased with the increased N, having the highest values at the 10 g N·m-2·a?1 treatment, but the 25 g N·m-2·a?1 still significantly increased the aboveground biomass relative to 0 g N·m-2·a?1. Leymus chinensis accommodate low N situation through allocating less N to carboxylation system and decreasing leaf mass per area (LMA) in order to get more light energy. Moderate N addition captured more light energy through increasing total chlorophyll (Chl) contents and decreasing the ratio of Chl a/b. Moderate N addition increased LMA, carboxylation efficiency, maximum carboxylation rate (Vcmax), maximum electron transport rate (Jmax) and decreased Jmax/Vcmax, thus allocating more N to carboxylation system to enhance carboxylation capability. Moreover, the photochemical activity of PSII was increased through higher effective quantum yield of PSII photochemistry, electron transport rate and photochemical quenching coefficient. Excessive N addition had negative effects on physiological variables of L. chinensis due to lower carboxylation capability and photochemical activity of PSII, further leading to decreased net photosynthetic rate, whereas increased non-photochemical quenching coefficient and carotenoids played the role in the dissipation of excess excitation energy. Overall, moderate N addition facilitated the photosynthetic characteristics of dominant species, but excessive N addition inhibited photosynthetic characteristics. The most appropriate N addition for the growth of L. chinensis was 5-10 g N·m-2·a?1 in the temperate steppe of Nei Mongol, China.
实验在内蒙古锡林浩特内蒙古大学毛登牧场实验站(116.03°?116.50° E, 44.80°?44.82° N)进行。该地属于温带大陆性气候, 年平均气温0?1 ℃, 年积温1800 ℃, 年降水量300-360 mm, 降水多集中在6?8月, 无霜期90?115天。土壤以栗钙土为主, 有少量褐色土, 栗钙土层15-80 cm, 土壤有机质含量为2%?3%, 土质比较肥沃。研究区域优势种为羊草和大针茅(Stipa grandis)。
1.2 实验设计
N添加试验设置6个水平, 分别为对照(CK)、N1 (0 g N·m-2·a?1)、N2 (2 g N·m-2·a?1)、N3 (5 g N·m-2·a?1)、N4 (10 g N·m-2·a?1)和N5 (25 g N·m-2·a?1), 其设置依据文献Liu等(2013)。2010年我国北方草原生态系统的N沉降量为22.6 kg N·hm-2·a-1, 平均每年的增加量为0.42 kg N·hm-2 (Liu et al., 2013)。N2基本上代表了2010年的N沉降量, 通过施加N肥来模拟未来几十年的N沉降。每个处理3组重复, 共计18个处理实验小区(面积为6 m × 6 m)。N肥为硝酸铵。为确保不受磷限制, 除了CK外, 各处理小区添加一定量的磷(1 g P2O5·m-2)。相邻小区之间设置1 m的缓冲带。从2014年开始, 每年6月一次性均匀喷洒。
TN, total nitrogen; TP, total phosphorus; TC, total carbon; TK, total kalium; TS, total sulfur. 新窗口打开 Table 2 表2 表2不同施氮水平下样地的土壤含水量(W) (平均值±标准误差, n = 6) Table 2Soil water content (W) of plots under different nitrogen (N) addition levels (mean ± SE, n = 6)
氮处理 N treatment
CK
N1
N2
N3
N4
N5
W (%)
12.73 ± 0.41a
12.83 ± 0.27a
10.92 ± 1.67bc
10.01 ± 0.53b
9.61 ± 0.92bc
9.11 ± 0.34c
不同小写字母表示处理间差异显著(p < 0.05)。CK, 不添加N和P。N1、N2、N3、N4、N5分别表示氮添加量为0、2、5、10、25 g N·m-2·a?1, 磷添加量1 g P2O5·m-2。Different lowercase letters indicate significant differences among treatments (p < 0.05). CK, without added N and phosphorus (P). N1, N2, N3, N4, N5 indicate N addition rates as 0, 2, 5, 10, 25 g N·m-2·a?1 , respectively. 1 g P2O5·m-2 was added to each treatment except CK. 新窗口打开
N添加对羊草地上生物量有显著影响(图1A; p < 0.05)。羊草地上生物量随着N添加量的增加而增加, 其中以N4处理促进作用最为明显, 极显著地促进了羊草地上生物量(p < 0.01)。而N5处理时有降低趋势, 但与CK之间差异性显著(p < 0.05); 与CK相比, N2、N3、N4和N5处理下羊草的地上生物量分别增加了23.38%、33.11%、45.96%和37.88%。 显示原图|下载原图ZIP|生成PPT 图1不同施氮水平羊草的地上生物量、叶片氮含量和叶片形态特性(平均值±标准误差)。不同小写字母表示处理间差异显著(p < 0.05)。处理同表2。 -->Fig. 1The aboveground biomass, leaf nitrogen content, and leaf morphological traits of Leymus chinensis under different nitrogen addition levels (mean ± SE). Different lowercase letters indicate significant differences among treatments (p < 0.05). Treatment see Table 2. -->
N添加对羊草的叶片N含量有显著影响(图1B; p < 0.05)。随着N添加, 叶片N含量也显著增加; 与CK相比, N2、N3、N4和N5处理下羊草的叶片N含量分别增加了21.08%、32.35%、82.84%和81.86%。 N添加对羊草LMA有显著影响(图1C; p < 0.05)。N2和N3处理显著促进了羊草的LMA (p < 0.05), 而N4和N5处理有下降趋势, 但与CK差异不显著(p > 0.05)。N2、N3和N4处理分别比CK增加了32.48%、41.66%和12.21%, 而N1和N5处理分别比CK降低了6.08%和0.98%。羊草叶片含水量和LMA变化趋势一致, 各处理间差异显著(图1D; p < 0.05), 只是变化程度比较平缓, 说明N添加在一定程度上改变了叶片的水分机制。
2.2 叶片气体交换特征对N添加的响应
N添加对羊草Pn有极显著影响(图2A; p < 0.01)。在低N水平, Pn随着N添加梯度而增加, 但在高N水平下有降低的趋势。N3和N4处理极显著地增加了28.07%和19.19% (p < 0.01), N5处理降低了2.31%。Gs、Ci和Tr随着N添加量的增加变化趋势基本一致, N1处理轻微增加, 而N2处理急剧增加, 随后逐渐降低, N2处理增加幅度最大, 并且都与CK的差异性达到了极显著水平(p < 0.01), 分别增加了216.8%、133.5%和137.5% (图2B-2D; p < 0.01)。WUE随着N添加先降低后升高, 呈现出开口向上的“抛物线”形(图2E; p < 0.01)。与CK相比, N2、N3和N4分别降低了54.5%、39.8%和38.0%, 差异性极显著(p < 0.01), 只有N5是增加了56.1%, 差异性极显著(p < 0.01)。不同的N添加水平下, 羊草的Pn和叶片N含量的线性关系存在差异(图3)。所有处理下的Pn和叶片N含量的关系不显著(图3A; R2 = 0.10, p > 0.05)。而除N5外, 其他处理下的Pn和叶片N含量显著的正相关(图3B; R2 = 0.43, p < 0.05)。 显示原图|下载原图ZIP|生成PPT 图2不同施氮水平羊草主要叶片气体交换参数(平均值±标准误差)。不同小写字母表示处理间差异显著(p < 0.05)。处理同表2。 -->Fig. 2The main leaf gas exchange parameters of Leymus chinensis under different nitrogen addition levels (mean ± SE). Different lowercase letters indicate significant differences among treatments (p < 0.05). Pn, net photosynthetic rate; Gs, stomatal conductance; Ci, intercellular CO2 concentration; Tr, transpiration rate; WUE, water use efficiency. Treatment see Table 2. -->
显示原图|下载原图ZIP|生成PPT 图3不同施氮水平羊草的净光合速率(Pn)和叶片氮含量的关系。A, 净光合速率和叶片氮含量的关系。B, 净光合速率和叶片氮含量(N5处理除外)的关系。 -->Fig. 3Relationships between net photosynthetic rate (Pn) and leaf nitrogen (N) content of Leymus chinensis across treatments (A) or across all nitrogen with an exception of the highest N addition (B). -->
2.3 叶片色素含量对N添加的响应
N添加对羊草叶绿素(Chl)有极显著影响(图4A; p < 0.01)。Chl随着施N量先增加后降低。CK、N1、N2差异不显著(p > 0.05)。而N3、N4、N5比CK分别显著增加了28.2%、55.0%和46.2% (p < 0.05)。Chl a/b和类胡萝卜素(Car)二者都是随着N添加的增加出现了微弱的增加然后急剧下降(图4B, C; p < 0.01)。与CK相比, N2和N3使二者微弱地增加, 但是差异性不显著(p > 0.05); 而N3、N4、N5使二者极显著下降(p < 0.01), 与CK相比, Chl a/b分别降低了26.4%、40.2%和36.4%, Car分别降低了35.0%、65.8%和52.9%。 显示原图|下载原图ZIP|生成PPT 图4不同施氮水平羊草叶片色素参数(平均值±标准误差)。不同小写字母表示处理间差异显著(p < 0.05)。处理同表2。 -->Fig. 4The leaf pigment parameters of Leymus chinensis under different nitrogen addition levels (mean ± SE). Different lowercase letters indicate significant differences among treatments (p < 0.05). Chl, chlorophyll; Car, carotenoids. Treatment see Table 2. -->
2.4 叶绿素荧光对N添加的响应
N添加对叶绿素荧光参数有极显著影响(表3; p < 0.01)。Fv/Fm在0.80-0.82范围内极显著波动(p = 0.006)。ФPSII、ETR和qP都随着N添加量的增加而轻微地增加, 然后急剧下降; 而NPQ起初极显著增加, 然后又极显著下降(p < 0.01), N4和N5与N1和N2达到了极显著差异(p < 0.01), 表现为“大起大落”的态势。在N5和CK之间, ФPSII、ETR和qP呈现出极显著差异(p < 0.01), 而Fv/Fm和NPQ并没有呈现出显著差异(p > 0.05)。 Table 3 表3 表3不同施氮水平下羊草叶绿素荧光特性(平均值±标准误差) Table 3Chlorophyll a fluorescence characteristics of Leymus chinensis under different nitrogen addition levels (mean ± SE)
氮处理 N treatment
最大光化学量子产量 Fv/Fm
实际光化学量子产量 ФPSII
电子传递速率 ETR (μmol·m-2·s-1)
光化学淬灭系数 qP
非光化学淬灭系数 NPQ
CK
0.81 ± 0.002c
0.23 ± 0.001a
153.0 ± 0.82a
0.54 ± 0.002b
2.63 ± 0.05c
N1
0.80 ± 0.001c
0.23 ± 0.003a
156.9 ± 2.02a
0.55 ± 0.003ab
3.08 ± 0.05a
N2
0.82 ± 0.002a
0.23 ± 0.004a
157.0 ± 2.80a
0.59 ± 0.010a
3.11 ± 0.05a
N3
0.82 ± 0.002ab
0.24 ± 0.009a
161.9 ± 6.18a
0.54 ± 0.008b
2.82 ± 0.05b
N4
0.81 ± 0.004c
0.19 ± 0.010b
127.3 ± 6.77b
0.46 ± 0.016c
2.74 ± 0.09bc
N5
0.81 ± 0.003abc
0.17 ± 0.003b
117.0 ± 1.62b
0.44 ± 0.004c
2.79 ± 0.05bc
不同小写字母表示处理间差异显著(p < 0.05)。处理同表2。ETR, electron transport rate; Fv/Fm, maximum photochemical efficiency; ФPSII, actual quantum yield; NPQ, non-photochemical quenching coefficient; qP, photochemical quenching coefficient. Different lower case letters indicate significant differences among treatments (p < 0.05). Treatment see Table 2. 新窗口打开
2.5 CO2响应曲线
N添加对所有CO2响应参数有极显著的影响(表4; p < 0.01)。不同处理的CE之间差异显著(p < 0.05), 并在N3处理出现了高峰; N1-N5处理的Vcmax比CK分别增加了55.3%、36.5%、225.5%、138.7%和183.6%, 差异显著(p < 0.05), 也在N3处理达到了高峰; Jmax变化趋势和Vcmax大体相同, N1-N5处理的Vcmax比CK分别增加了62.6%、50.4%、190.0%、92.3%和123.3%; Jmax/Vcmax变化较为缓慢, N1、N2处理轻微增加, 而N3-N5处理逐渐降低, 并且比CK降低了10.9%、19.4%和21.2%。 Table 4 表4 表4不同施氮水平下羊草叶片光合能力特性(平均值±标准误差) Table 4Photosynthetic capacity characteristics of Leymus chinensis under different nitrogen addition levels (mean ± SE)
氮处理 N treatment
初始羧化效率 CE (mol·m-2·s-1)
最大羧化速率 Vcmax (μmol·m-2·s-1)
最大电子传递速率 Jmax (μmol·m-2·s-1)
Jmax/Vcmax
CK
0.005 ± 0.000 2f
16.83 ± 0.49e
16.48 ± 0.48e
0.98 ± 0.03b
N1
0.015 ± 0.000 4d
26.14 ± 0.75d
26.79 ± 0.77d
1.02 ± 0.03ab
N2
0.018 ± 0.000 5c
22.98 ± 0.66d
24.79 ± 0.72d
1.08 ± 0.03a
N3
0.045 ± 0.001 3a
54.79 ± 1.58a
47.80 ± 1.38a
0.87 ± 0.03c
N4
0.030 ± 0.000 9b
40.18 ± 1.16c
31.69 ± 0.91c
0.79 ± 0.02d
N5
0.010 ± 0.000 3e
47.73 ± 1.38b
36.80 ± 1.06b
0.77 ± 0.02d
CE, carboxylation efficiency; Jmax, maximum electron transport rate; Vcmax, maximum carboxylation rate. Different lowercase letters indicate significant differences among treatments (p < 0.05). Treatment see Table 2.不同小写字母表示处理间差异显著(p < 0.05)。处理同表2。 新窗口打开
AberJD, GoodaleCL, OllingerSV, SmithML, MagillAH, MartinME (2003). Is nitrogen deposition altering the nitrogen status of northern forests? Revista Me?dica De Chile, 53, 158-167. [本文引用: 2]
[2]
AndersonTM, DongY, McnaughtonSJ (2006). Nutrient acquisition and physiological responses of dominant Serengeti grasses to variation in soil texture and grazing. Journal of Ecology, 94, 1164-1175. [本文引用: 1]
[3]
BaiWM, WangZW, ChenQS, ZhangWH, LiLH (2008). Spatial and temporal effects of nitrogen addition on root life span of Leymus chinensis in a typical steppe of Inner Mongolia. Functional Ecology, 22, 583-591. [本文引用: 1]
[4]
BaiX, ChengJH, ZhengSX, ZhanSX, BaiYF (2014). Ecophysiological responses of Leymus chinensis to nitrogen and phosphorus additions in a typical steppe. Chinese Journal of Plant Ecology, 38, 103-115. (in Chinese with English abstract)[白雪, 程军回, 郑淑霞, 詹书侠, 白永飞 (2014). 典型草原建群种羊草对氮磷添加的生理生态响应 . 植物生态学报, 38, 103-115.] [本文引用: 3]
[5]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2009). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Global Change Biology, 16, 358-372. [本文引用: 1]
[6]
BoussadiaO, MariemFB, MechriB, BrahamW, BrahamM, HadjSBE (2008). Response to drought of two olive tree cultivars (cv Koroneki and Meski). Scientia Horticulturae, 116, 388-393. [本文引用: 2]
[7]
BubierJL, SmithR, JuutinenS, MooreTR, MinochaR, LongS, MinochaS (2011). Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia, 167, 355-368. [本文引用: 1]
[8]
CaoCL, LISX, MiaoF (1999). The Research situation about effects of nitrogen on certain physiological and biochemical process in plants. The Journal of Northwestern Agricultural University, 20(4), 99-104. (in Chinese with English abstract)[曹翠玲, 李生秀, 苗芳 (1999). 氮素对植物某些生理生化过程影响的研究进展 . 西北农业大学学报, 20(4), 99-104.] [本文引用: 1]
[9]
ChenH, LiDJ, GurmesaGA, YuGR, LiLH, ZhangW, FangHJ, MoJM (2015). Effects of nitrogen deposition on car- bon cycle in terrestrial ecosystems of China: A meta- analysis. Environmental Pollution, 206, 352-360. [本文引用: 1]
[10]
ChenSP, BaiYF, ZhangL, HanX (2005). Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China. Environmental & Experimental Botany, 53, 65-75. [本文引用: 3]
[11]
ChenZZ, WangSP (2000). The Typical Grassland Ecosystem in China. Science Press, Beijing. (in Chinese)[陈佐忠, 汪诗平 (2000). 中国典型草原生态系统. 科学出版社, 北京.] [本文引用: 1]
[12]
ClarkCM, TilmanD (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 451, 712-715. [本文引用: 1]
[13]
Demmig-AdamsB, WilliamW (1996). The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends in Plant Science, 1, 21-26. [本文引用: 1]
[14]
DingXH, LuoSZ, LiuJW, LiK, LiuGH (2012). Longitude gradient changes on plant community and soil stoichiometry characteristics of grassland in Hulunbeir. Acta Ecologica Sinica, 11, 3467-3476. (in Chinese with English abstract)[丁小慧, 罗淑政, 刘金巍, 李魁, 刘国华 (2012). 呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化 . 生态学报, 11, 3467-3476.] [本文引用: 1]
[15]
DuE, LiuX, FangJ (2014). Effects of nitrogen additions on biomass, stoichiometry and nutrient pools of moss Rhytidium rugosum in a boreal forest in Northeast China. Environmental Pollution, 188, 166-171. [本文引用: 2]
[16]
DuJ, ShuS, ShaoQ, AnY, ZhouH, GuoS, SunJ (2016). Mitigative effects of spermidine on photosynthesis and carbon-nitrogen balance of Cucumber seedlings under Ca(NO3)2 stress. Journal of Plant Research, 129, 1-13. [本文引用: 1]
[17]
GonzálezJA, GallardoMG, BoeroC, CruzML, PradoFE (2007). Altitudinal and seasonal variation of protective and photosynthetic pigments in leaves of the world’s highest elevation trees Polylepis tarapacana (Rosaceae) . Acta Oecologica, 32, 36-41. [本文引用: 2]
[18]
GrassiG, MeirP, CromerR, TompkinsD, JarvisPG (2002). Photosynthetic parameters in seedlings of Eucalyptus grandis as affected by rate of nitrogen supply. Plant, Cell & Environment, 25, 1677-1688. [本文引用: 3]
[19]
GuoCA, LiuF, XuXM (2006). Chlorophyll-b deficient and photosynthesis in plants. Plant Physiology Communications, 42, 967-973. (in Chinese with English abstract)[郭春爱, 刘芳, 许晓明 (2006). 叶绿素b缺失与植物的光合作用 . 植物生理学通讯, 42, 967-973.] [本文引用: 1]
[20]
GuoEH, HuD, TianCY, HuY, WangCY, YuYY (2008). Study on the effects of soil nitrogen and moisture on plant photosynthetic physiological ecology. Journal of Anhui Agriculture Sciences, 26, 11211-11213. (in Chinese with English abstract)[郭二辉, 胡聃, 田朝阳, 胡颖, 王从彦, 于盈盈 (2008). 土壤氮素与水分对植物光合生理生态的影响研究 . 安徽农业科学, 26, 11211-11213.] [本文引用: 1]
[21]
GuoTC, FengW, ZhaoHJ, XueGD, WangHC, WangYH, YaoZJ (2004). Photosynthetic characteristics of flag leaves and nitrogen effects in two winter wheat cultivars with different spike type. Acta Agronomica Sinica, 30(2), 115-121. (in Chinese with English abstract)[郭天财, 冯伟, 赵会杰, 薛国典, 王化岑, 王永华, 姚战军 (2004). 两种穗型冬小麦品种旗叶光合特性及氮素调控效应 . 作物学报, 30(2), 115-121.] [本文引用: 1]
[22]
GuoX, WangRQ, ChangRY, LiangXQ, WangCD, LuoY, GuoW (2014). Effects of nitrogen addition on growth and photosynthetic characteristics of Acer truncatum seedlings. Dendrobiology, 72, 151-161. [本文引用: 2]
[23]
KitaokaS, WatanabeY, KoikeT (2009). The effects of cleared larch canopy and nitrogen supply on gas exchange and leaf traits in deciduous broad-leaved tree seedlings. Tree Physiology, 29, 1503-1511. [本文引用: 1]
[24]
KrauseK, CherubiniP, BugmannH, SchleppiP (2012). Growth enhancement of Picea abies trees under long-term, low-dose N addition is due to morphological more than to physiological changes. Tree Physiology, 32, 1471-1481. [本文引用: 2]
[25]
LambersH, PoorterH (1992). Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. Advances in Ecological Research, 23, 187-261. [本文引用: 1]
[26]
LiDJ, MoJM, FangYT, CaiXA, XueJH, XuGL (2004). Effects of simulated nitrogen deposition on growth and photosynthesis of Schima superba, Castanopsis chinensis and Cryptocarya concinna seedlings. Acta Ecologica Sinica, 24, 876-882. (in Chinese with English abstract)[李德军, 莫江明, 方运霆, 蔡锡安, 薛璟花, 徐国良 (2004). 模拟氮沉降对三种南亚热带树苗生长和光合作用的影响 . 生态学报, 24, 876-882.] [本文引用: 1]
[27]
LiL, LiXY, LinLS, WangYJ, XueW (2011). Comparison of chlorophyll content and fluorescence parameters of six pasture species in two habitats in China. Chinese Journal of Plant Ecology, 35, 672-680. (in Chinese with English abstract)[李磊, 李向义, 林丽莎, 王迎菊, 薛伟 (2011). 两种生境条件下6种牧草叶绿素含量及荧光参数的比较 . 植物生态学报, 35, 672-680.] [本文引用: 1]
[28]
LiLJ, ZengDH, MaoR, YuZY (2012). Nitrogen and phosphorus resorption of Artemisia scoparia, Chenopodium acuminatum, Cannabis sativa, and Phragmites communis under nitrogen and phosphorus additions in a semiarid grassland, China. Plant, Soil & Environment, 58, 446-451. [本文引用: 1]
[29]
LiX, FengW, ZengXC (2006). Advances in chlorophyll fluorescence analysis and its uses. Acta Botanica Boreali- Occidentalia Sinica, 10, 2186-2196. (in Chinese with English abstract)[李晓, 冯伟, 曾晓春 (2006). 叶绿素荧光分析技术及应用进展 . 西北植物学报, 10, 2186-2196.] [本文引用: 2]
[30]
LiYY, LüXT, WangZW, ZhouC, HanXG (2014). Linking relative growth rates to biomass allocation: The responses of the grass Leymus chinensis to nitrogen addition. Phyton, 83, 283-289. [本文引用: 1]
LiuXJ, DuanL, MoJM, DuEZ, ShenJL, LuXK, ZhangY, ZhouXB, HeCN, ZhangFS (2011). Nitrogen deposition and its ecological impact in China: An overview. Environmental Pollution, 159, 2251-2264. [本文引用: 2]
[33]
LiuYS, PanQM, ZhengSX, BaiYF, HanXG (2012). Intra-seasonal precipitation amount and pattern differentially affect primary production of two dominant species of Inner Mongolia grassland. Acta Oecologica, 44, 2-10. [本文引用: 1]
[34]
LongSP, BernacchiCJ (2003). Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. Journal of Experimental Botany, 54, 2393-2401. [本文引用: 1]
[35]
MakhnevAK, MakhnevaNE (2010). Landscape-ecological and population aspects of the strategy of restoration of disturbed lands. Contemporary Problems of Ecology, 3, 318-322. [本文引用: 1]
[36]
MariotteP, ButtlerA, JohnsonD, ThébaultA, VandenbergheC (2012). Exclusion of root competition increases competitive abilities of subordinate plant species through root-shoot interactions. Journal of Vegetation Science, 23, 1148-1158. [本文引用: 1]
[37]
MoonM, KangKS, ParkIK, KimT, KimHS (2015). Effects of leaf nitrogen allocation on the photosynthetic nitrogen-use efficiency of seedlings of three tropical species in Indonesia. Journal of the Korean Society for Applied Biological Chemistry, 58, 1-9. [本文引用: 2]
[38]
NicodemusMA, SalifuFK, JacobsDF (2008). Growth, nutrition, and photosynthetic response of black walnut to varying nitrogen sources and rates. Journal of Plant Nutrition, 31, 1917-1936. [本文引用: 1]
[39]
OnodaY, HikosakaK, HiroseT (2004). Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Functional Ecology, 18, 419-425. [本文引用: 1]
[40]
PanQM, BaiYF, HanXG, YangJC (2005). Effects of nitrogen addition on a Leymus chinensis population in typical steppe of Inner Mongolia. Acta Phytoecologica Sinica, 29, 311-317. (in Chinese with English abstract)[潘庆民, 白永飞, 韩兴国, 杨景成 (2005). 氮素对内蒙古典型草原羊草种群的影响 . 植物生态学报, 29, 311-317.] [本文引用: 1]
[41]
PengQ, QiYC, DongYS, HeYT, XiaoSS (2014). Litter decomposition and the C and N dynamics as affected by N additions in a semi-arid temperate steppe, Inner Mongolia of China. Journal of Arid Land, 6, 432-444. [本文引用: 1]
[42]
SchreiberU, BilgerW, NeubauerC (1995). Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. Ecological Studies Analysis & Synthesis, 100, 49-70. [本文引用: 1]
[43]
SunL, DongYS, QiYC, HeYT, PengQ, LiuXC, JiaJQ, GuoSF, CaoCC (2014). Intra- and inter-annual variation of soil microbial and enzymatic response to water and nitrogen addition in a Chinese semi-arid steppe. Journal of Pure and Applied Microbiology, 8, 1339-1351. [本文引用: 1]
[44]
ThomasRQ, CanhamCD, WeathersKC, GoodaleCL (2010). Increased tree carbon storage in response to nitrogen deposition in the US. Nature Geoscience, 3, 13-17. [本文引用: 1]
[45]
ÜloN, KullK (2003). Leaf structure vs. nutrient relationships vary with soil conditions in temperate shrubs and trees. Acta Oecologica, 24, 209-219. [本文引用: 1]
[46]
WallaceZP, LovettGM, HartJE, MachonaB (2007). Effects of nitrogen saturation on tree growth and death in a mixed-oak forest. Forest Ecology & Management, 243, 210-218. [本文引用: 1]
[47]
WangG, LiuF (2014). Carbon allocation of Chinese pine seedlings along a nitrogen addition gradient. Forest Ecology & Management, 334, 114-121. [本文引用: 2]
[48]
WangHZ, HanL, XuYL, NiuJL (2014). Photosynthetic responses of the heteromorphic leaves in Populus euphratica to light intensity and CO2 concentration. Chinese Journal of Plant Ecology, 38, 1099-1109. (in Chinese with English abstract)[王海珍, 韩路, 徐雅丽, 牛建龙 (2014). 胡杨异形叶光合作用对光强与CO2浓度的响应 . 植物生态学报, 38, 1099-1109.] [本文引用: 3]
[49]
WangRZ (1997). The niche breadths and niche overlaps of main plant populations in Leymus chinensis grassland for grazing. Acta Phytoecologica Sinica, 21, 9-16. (in Chinese with English abstract)[王仁忠 (1997). 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究 . 植物生态学报, 21, 9-16.] [本文引用: 1]
[50]
WangXK (2006). Principle and Technology of Plant Physiological and Biochemical Experiments. Higher Education Press, Beijing. 134-136. (in Chinese)[王学奎 (2006).植物生理生化实验原理和技术. 高等教育出版社, 北京. 134-136.] [本文引用: 1]
[51]
WangYH, HeXY, ZhouGS (2002). Study on the responses of Leymus chinensis steppe to grazing in Songnen Plain. Acta Agrestia Sinica, 10, 45-49. (in Chinese with English abstract)[王玉辉, 何兴元, 周广胜 (2002). 放牧强度对羊草草原的影响 . 草地学报, 10, 45-49.] [本文引用: 1]
[52]
WarrenCR, AdamsMA, ChenZL (2000). Is photosynthesis related to concentrations of nitrogen and Rubisco in leaves of Australian native plants? Functional Plant Biology, 27, 407-416. [本文引用: 1]
[53]
WarrenCR, DreyerE, AdamsMA (2003). Photosynthesis- Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Tree, 17, 359-366. [本文引用: 1]
[54]
WullschlegerSD (1993). Biochemical limitations to carbon assimilation in C3 plants—A retrospective analysis of the A/Ci curves from 109 species. Journal of Experimental Botany, 44, 907-920. [本文引用: 2]
[55]
XiaoSS, DongYS, QiYC, PengQ, HeYT, LiuXC (2010). Effects of mineral fertilizer addition on leaf functional traits and photosynthetic characteristics of Leymus chinensis from a temperate grassland in Inner Mongolia in China. Acta Scientiae Circumstantiae, 30, 2535-2543. (in Chinese with English abstract)[肖胜生, 董云社, 齐玉春, 彭琴, 何亚婷, 刘欣超 (2010). 内蒙古温带草原羊草叶片功能特性与光合特征对外源氮输入的响应 . 环境科学学报, 30, 2535-2543.] [本文引用: 3]
[56]
XiaoYG, ChenQ, ShanL, BrueckH, DittertK, TaubeF, SchnyderH (2011). Tradeoffs between nitrogen- and water-use efficiency in dominant species of the semiarid steppe of Inner Mongolia. Plant & Soil, 340, 227-238. [本文引用: 1]
[57]
XuZZ, ZhouGS (2006). Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis. Planta, 224, 1080-1090. [本文引用: 1]
[58]
YaoM, RuiJ, LiJ, DaiY, BaiY, HeděnecP, WangJM, ZhangSH, PeiKQ, LiuC, WangYF, HeZL, FrouzJ, LiXZ (2014). Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. Soil Biology & Biochemistry, 79, 81-90. [本文引用: 1]
[59]
YeZP, YuQ, KangHJ (2012). Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions. Photosynthetica, 50, 472-476. [本文引用: 1]
[60]
YuanYH, FanHB, HuangQR, LiaoYC, HuangRZ (2009). Effects of long-term fertilization on rice photosynthetic traits and water use efficiency. Chinese Journal of Ecology, 28, 2239-2244. (in Chinese with English abstract)[袁颖红, 樊后保, 黄欠如, 廖迎春, 黄荣珍 (2009). 长期施肥对水稻光合特性及水分利用效率的影响 . 生态学杂志, 28, 2239-2244.] [本文引用: 1]
[61]
ZhanSX, ZhengSX, WangY, BaiYF (2016). Response and correlation of above- and below-ground functional traits of Leymus chinensis to nitrogen and phosphorus additions. Chinese Journal of Plant Ecology, 40, 36-47. (in Chinese with English abstract)[詹书侠, 郑淑霞, 王扬, 白永飞 (2016). 羊草的地上-地下功能性状对氮磷施肥梯度的响应及关联 . 植物生态学报, 40, 36-47.] [本文引用: 2]
[62]
ZhangJ, HanX (2008). N2O emission from the semi-arid ecosystem under mineral fertilizer (urea and superphosphate) and increased precipitation in northern China. Atmospheric Environment, 42, 291-302. [本文引用: 1]
[63]
ZhangL, YangYX, ZhanXY, ZhangCJ, ZhouSX, WuDX (2010). Responses of a dominant temperate grassland plant (Leymus chinensis) to elevated carbon dioxide and nitrogen addition in China. Journal of Environmental Quality, 39, 251-259. [本文引用: 1]
[64]
ZhangT, YangS, GuoR, GuoJ (2016). Warming and nitrogen addition alter photosynthetic pigments, sugars and nutrients in a temperate meadow ecosystem. PLOS ONE, 11, e0155375. doi:10.1371/journal.pone.0155375. [本文引用: 1]
[65]
ZhangXC, YuXF, GaoSM (2010). Effects of nitrogen application rates on photosynthetic energy utilization in wheat leaves under elevated atmospheric CO2 concentration. Chinese Journal of Plant Ecology, 34, 1196-1203. (in Chinese with English abstract)[张绪成, 于显枫, 高世铭 (2010). 高大气CO2浓度下氮素对小麦叶片光能利用的影响 . 植物生态学报, 34, 1196-1203.] [本文引用: 1]
[66]
ZhangYH, HeNP, ZhangGM, HuangJH, HanXG (2013). Nitrogen deposition and Leymus chinensis leaf chlorophyll content in Inner Mongolian grassland. Acta Ecologica Sinica, 33, 6786-6794. (in Chinese with English abstract)[张云海, 何念鹏, 张光明, 黄建辉, 韩兴国 (2013). 氮沉降强度和频率对羊草叶绿素含量的影响 . 生态学报, 33, 6786-6794.] [本文引用: 1]
[67]
ZhangYM, ZhouGS (2012). Advances in leaf maximum carboxylation rate and its response to environmental factors. Acta Ecologica Sinica, 32, 5907-5917. (in Chinese with English abstract)[张彦敏, 周广胜 (2012). 植物叶片最大羧化速率及其对环境因子响应的研究进展 . 生态学报, 32, 5907-5917.] [本文引用: 1]
[68]
ZhangZY, GongJR, LiuM, HuangYM, YanX, QiY, WangYH (2013). Dominant species and ecosystem gas exchange in temperate grassland under different land use patterns. Chinese Journal of Plant Ecology, 37, 718-727. (in Chinese with English abstract)[张梓瑜, 龚吉蕊, 刘敏, 黄永梅, 晏欣, 祁瑜, 王忆慧 (2013). 温带草原不同土地利用方式下优势种植物和生态系统的气体交换 . 植物生态学报, 37, 718-727.] [本文引用: 1]
[69]
ZhaoC, LiuQ (2009). Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization. Canadian Journal of Forest Research, 39, 1-11. [本文引用: 4]
[70]
ZhuJT, LiXY, ZhangXM, LinLS, YangSG (2010). Nitrogen allocation and partitioning within a leguminous and two non-leguminous plant species growing at the southern fringe of China’s Taklamakan Desert. Chinese Journal of Plant Ecology, 34, 1025-1032. (in Chinese with English abstract)[朱军涛, 李向义, 张希明, 林丽莎, 杨尚功 (2010). 塔克拉玛干沙漠南缘豆科与非豆科植物的氮分配 . 植物生态学报, 34, 1025-1032.] [本文引用: 1]
Is nitrogen deposition altering the nitrogen status of northern forests? 2 2003
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
... 叶绿素在绿色植物光合作用中发挥着重要作用, 实现光能到化学能的转化, 为碳同化积累同化力(张云海等, 2013).叶绿素荧光对研究光系统对光能的吸收、传递、耗散、分配等具有独特的作用, 可以反映植物光合作用与环境之间相互关系的内在特性(李晓等, 2006).N元素是光合器官的组成成分, 叶片中有30%-40%的N参与到羧化反应中, 其中分配给羧化系统的N元素比例决定了最终的光合效率, 而叶片缺N将导致植物光合速率、光合量子效率和与碳代谢相关的酶类活性降低, 影响着植物叶片最大羧化速率(Vcmax) (Warren et al., 2000; 郭二辉等, 2008; Nicodemus et al., 2008; 朱军涛等, 2010).然而, 关于解释N沉降影响植物生长的光合生理机制仍有争论.土壤N含量主要受N沉降的速率和持续时间的影响, 而一些地区正从自然状态下的N限制转向富营养化甚至饱和(Aber et al., 2003; Krause et al., 2012).而N沉降对植物生长的促进和抑制作用均有报道, 可能主要依赖于N沉降是否引起了“N饱和” (Wallace et al., 2007; Zhao & Liu, 2009; Thomas et al., 2010).因此, 有必要确定当地是否出现了“N饱和”以及羊草最适合的施N量. ...
Nutrient acquisition and physiological responses of dominant Serengeti grasses to variation in soil texture and grazing. 1 2006
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Spatial and temporal effects of nitrogen addition on root life span of Leymus chinensis in a typical steppe of Inner Mongolia. 1 2008
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
典型草原建群种羊草对氮磷添加的生理生态响应 3 2014
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. 1 2009
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Response to drought of two olive tree cultivars (cv Koroneki and Meski). 2 2008
... N添加能够影响植物光合作用的过程以及相关的气体交换参数(Grassi et al., 2002; Chen et al., 2005).N元素是光合酶、可溶性蛋白质和光合色素的组成成分, 施N能够增加叶片的Gs, 促进CO2向叶肉细胞的扩散和供应, 从而增加植被的Pn, 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
Effects of nitrogen deposition on car- bon cycle in terrestrial ecosystems of China: A meta- analysis. 1 2015
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China. 3 2005
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
... N添加能够影响植物光合作用的过程以及相关的气体交换参数(Grassi et al., 2002; Chen et al., 2005).N元素是光合酶、可溶性蛋白质和光合色素的组成成分, 施N能够增加叶片的Gs, 促进CO2向叶肉细胞的扩散和供应, 从而增加植被的Pn, 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
... , 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
1 2000
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. 1 2008
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
The role of xanthophyll cycle carotenoids in the protection of photosynthesis. 1 1996
Mitigative effects of spermidine on photosynthesis and carbon-nitrogen balance of Cucumber seedlings under Ca(NO3)2 stress. 1 2016
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Altitudinal and seasonal variation of protective and photosynthetic pigments in leaves of the world’s highest elevation trees Polylepis tarapacana (Rosaceae) 2 2007
Nitrogen and phosphorus resorption of Artemisia scoparia, Chenopodium acuminatum, Cannabis sativa, and Phragmites communis under nitrogen and phosphorus additions in a semiarid grassland, China. 1 2012
Linking relative growth rates to biomass allocation: The responses of the grass Leymus chinensis to nitrogen addition. 1 2014
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Enhanced nitrogen deposition over China. 1 2013
... N添加试验设置6个水平, 分别为对照(CK)、N1 (0 g N·m-2·a?1)、N2 (2 g N·m-2·a?1)、N3 (5 g N·m-2·a?1)、N4 (10 g N·m-2·a?1)和N5 (25 g N·m-2·a?1), 其设置依据文献Liu等(2013).2010年我国北方草原生态系统的N沉降量为22.6 kg N·hm-2·a-1, 平均每年的增加量为0.42 kg N·hm-2 (Liu et al., 2013).N2基本上代表了2010年的N沉降量, 通过施加N肥来模拟未来几十年的N沉降.每个处理3组重复, 共计18个处理实验小区(面积为6 m × 6 m).N肥为硝酸铵.为确保不受磷限制, 除了CK外, 各处理小区添加一定量的磷(1 g P2O5·m-2).相邻小区之间设置1 m的缓冲带.从2014年开始, 每年6月一次性均匀喷洒. ...
Nitrogen deposition and its ecological impact in China: An overview. 2 2011
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Intra-seasonal precipitation amount and pattern differentially affect primary production of two dominant species of Inner Mongolia grassland. 1 2012
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. 1 2003
Landscape-ecological and population aspects of the strategy of restoration of disturbed lands. 1 2010
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Exclusion of root competition increases competitive abilities of subordinate plant species through root-shoot interactions. 1 2012
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Effects of leaf nitrogen allocation on the photosynthetic nitrogen-use efficiency of seedlings of three tropical species in Indonesia. 2 2015
... N添加能够影响植物光合作用的过程以及相关的气体交换参数(Grassi et al., 2002; Chen et al., 2005).N元素是光合酶、可溶性蛋白质和光合色素的组成成分, 施N能够增加叶片的Gs, 促进CO2向叶肉细胞的扩散和供应, 从而增加植被的Pn, 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
Growth, nutrition, and photosynthetic response of black walnut to varying nitrogen sources and rates. 1 2008
... 叶绿素在绿色植物光合作用中发挥着重要作用, 实现光能到化学能的转化, 为碳同化积累同化力(张云海等, 2013).叶绿素荧光对研究光系统对光能的吸收、传递、耗散、分配等具有独特的作用, 可以反映植物光合作用与环境之间相互关系的内在特性(李晓等, 2006).N元素是光合器官的组成成分, 叶片中有30%-40%的N参与到羧化反应中, 其中分配给羧化系统的N元素比例决定了最终的光合效率, 而叶片缺N将导致植物光合速率、光合量子效率和与碳代谢相关的酶类活性降低, 影响着植物叶片最大羧化速率(Vcmax) (Warren et al., 2000; 郭二辉等, 2008; Nicodemus et al., 2008; 朱军涛等, 2010).然而, 关于解释N沉降影响植物生长的光合生理机制仍有争论.土壤N含量主要受N沉降的速率和持续时间的影响, 而一些地区正从自然状态下的N限制转向富营养化甚至饱和(Aber et al., 2003; Krause et al., 2012).而N沉降对植物生长的促进和抑制作用均有报道, 可能主要依赖于N沉降是否引起了“N饱和” (Wallace et al., 2007; Zhao & Liu, 2009; Thomas et al., 2010).因此, 有必要确定当地是否出现了“N饱和”以及羊草最适合的施N量. ...
Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. 1 2004
... N添加能够影响植物光合作用的过程以及相关的气体交换参数(Grassi et al., 2002; Chen et al., 2005).N元素是光合酶、可溶性蛋白质和光合色素的组成成分, 施N能够增加叶片的Gs, 促进CO2向叶肉细胞的扩散和供应, 从而增加植被的Pn, 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
氮素对内蒙古典型草原羊草种群的影响 1 2005
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Litter decomposition and the C and N dynamics as affected by N additions in a semi-arid temperate steppe, Inner Mongolia of China. 1 2014
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. 1 1995
Intra- and inter-annual variation of soil microbial and enzymatic response to water and nitrogen addition in a Chinese semi-arid steppe. 1 2014
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Increased tree carbon storage in response to nitrogen deposition in the US. 1 2010
... 叶绿素在绿色植物光合作用中发挥着重要作用, 实现光能到化学能的转化, 为碳同化积累同化力(张云海等, 2013).叶绿素荧光对研究光系统对光能的吸收、传递、耗散、分配等具有独特的作用, 可以反映植物光合作用与环境之间相互关系的内在特性(李晓等, 2006).N元素是光合器官的组成成分, 叶片中有30%-40%的N参与到羧化反应中, 其中分配给羧化系统的N元素比例决定了最终的光合效率, 而叶片缺N将导致植物光合速率、光合量子效率和与碳代谢相关的酶类活性降低, 影响着植物叶片最大羧化速率(Vcmax) (Warren et al., 2000; 郭二辉等, 2008; Nicodemus et al., 2008; 朱军涛等, 2010).然而, 关于解释N沉降影响植物生长的光合生理机制仍有争论.土壤N含量主要受N沉降的速率和持续时间的影响, 而一些地区正从自然状态下的N限制转向富营养化甚至饱和(Aber et al., 2003; Krause et al., 2012).而N沉降对植物生长的促进和抑制作用均有报道, 可能主要依赖于N沉降是否引起了“N饱和” (Wallace et al., 2007; Zhao & Liu, 2009; Thomas et al., 2010).因此, 有必要确定当地是否出现了“N饱和”以及羊草最适合的施N量. ...
Leaf structure vs. nutrient relationships vary with soil conditions in temperate shrubs and trees. 1 2003
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Is photosynthesis related to concentrations of nitrogen and Rubisco in leaves of Australian native plants? 1 2000
... 叶绿素在绿色植物光合作用中发挥着重要作用, 实现光能到化学能的转化, 为碳同化积累同化力(张云海等, 2013).叶绿素荧光对研究光系统对光能的吸收、传递、耗散、分配等具有独特的作用, 可以反映植物光合作用与环境之间相互关系的内在特性(李晓等, 2006).N元素是光合器官的组成成分, 叶片中有30%-40%的N参与到羧化反应中, 其中分配给羧化系统的N元素比例决定了最终的光合效率, 而叶片缺N将导致植物光合速率、光合量子效率和与碳代谢相关的酶类活性降低, 影响着植物叶片最大羧化速率(Vcmax) (Warren et al., 2000; 郭二辉等, 2008; Nicodemus et al., 2008; 朱军涛等, 2010).然而, 关于解释N沉降影响植物生长的光合生理机制仍有争论.土壤N含量主要受N沉降的速率和持续时间的影响, 而一些地区正从自然状态下的N限制转向富营养化甚至饱和(Aber et al., 2003; Krause et al., 2012).而N沉降对植物生长的促进和抑制作用均有报道, 可能主要依赖于N沉降是否引起了“N饱和” (Wallace et al., 2007; Zhao & Liu, 2009; Thomas et al., 2010).因此, 有必要确定当地是否出现了“N饱和”以及羊草最适合的施N量. ...
Photosynthesis- Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. 1 2003
... 差异不显著(CK、N1、N2) (Wullschleger et al., 1993).超过N2之后, Jmax/Vcmax开始逐渐降低, 使更多的N分配给羧化系统, 增加羧化效率, 利于Pn的增加.N5处理时, 超过植物正常需求的N分配给Rubisco以蛋白质的形式储存起来, 并不表现出强烈的羧化能力, 最终使Pn显著降低(Grassi et al., 2002). ...
内蒙古温带草原羊草叶片功能特性与光合特征对外源氮输入的响应 3 2010
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
... N添加能够影响植物光合作用的过程以及相关的气体交换参数(Grassi et al., 2002; Chen et al., 2005).N元素是光合酶、可溶性蛋白质和光合色素的组成成分, 施N能够增加叶片的Gs, 促进CO2向叶肉细胞的扩散和供应, 从而增加植被的Pn, 而N添加引起光合速率增加的原因有气孔因素和非气孔因素(Chen et al., 2005; 肖胜生等, 2010).本研究中, 适量的N添加使Gs增加, 而Ci和CE也相应增加, 说明适量的N添加导致Pn的增加可能是因为Gs和羧化能力增加的共同作用.N5处理时, Pn随着Gs的降低而降低, 但是Ci并没有显著下降, CE却显著降低, 说明Pn的降低并不是Gs降低而产生的结果, 应该是叶肉细胞羧化能力降低引起的结果.植物的光合能力和叶片N含量存在显著的正相关关系(Moon et al., 2015).本文中二者相关性随着施N水平的增加而变化, 其中CK到N4处理下二者表现为显著的正相关关系(R2 = 0.43, p < 0.05), 而CK到N5处理下二者的关系不显著(R2 = 0.10, p > 0.05).这说明随着施N的增加(CK到N4), N较多地分配给光合器官, 进而提高了光合能力, 而大量施N (N5)后N可能更多地分配到细胞壁而非光合器官, 从而降低了光合酶的活性, 不利于光合能力的增加(Onoda et al., 2004; Moon et al., 2015).同时, 过量供N导致植株体内营养失衡, 也会导致N代谢加强, 与碳同化竞争光合作用中光反应产生的同化力(ATP和NADPH), 这会导致光合碳同化速率降低, 进而引起地上生物量的减少(曹翠玲等, 1999; 李德军等, 2004).高N时(N5), Gs、Tr和叶片含水量显著降低, 而WUE显著增加, 说明羊草的叶片在大量施N后可以优先提高WUE而不是把有限的水分用于碳固定, 这可能是羊草在适应高N施肥下的一种权衡机制(Guo et al., 2014).另外, 土壤含水量在高N时有下降的趋势, 这可能是因为施N后地上生物量显著增加, 叶片生物量也随之增加, 植物需要更多的水分来满足其生长的需要, 降低了土壤含水量, 从而表现出叶片含水量的降低. ...
Tradeoffs between nitrogen- and water-use efficiency in dominant species of the semiarid steppe of Inner Mongolia. 1 2011
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis. 1 2006
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. 1 2014
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions. 1 2012
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
N2O emission from the semi-arid ecosystem under mineral fertilizer (urea and superphosphate) and increased precipitation in northern China. 1 2008
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
Responses of a dominant temperate grassland plant (Leymus chinensis) to elevated carbon dioxide and nitrogen addition in China. 1 2010
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Warming and nitrogen addition alter photosynthetic pigments, sugars and nutrients in a temperate meadow ecosystem. 1 2016
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
... 近年来我国草地陆续开展了较多N沉降生态环境效应的试验研究, 并取得了显著的研究成果, 但相关研究基本上还局限在植物生长、生物多样性、生态系统碳水交换、土壤呼吸、N素转化、凋落物分解以及土壤微生物及养分变化等方面(潘庆民等, 2005; Liu et al., 2011; Peng et al., 2014; Sun et al., 2014), 针对N沉降对生态系统不同组织水平的影响开展的研究成果还少有报道, 例如主要优势种和建群种光合及生理生态特性对N沉降变化响应的试验更为鲜见, 并且相关机制尚不清楚(白雪等, 2014).优势种是草地生态群落的重要组成部分, 虽然物种数较少, 但在群落生物量上的分配比例较大, 在一定程度上决定了草地生态系统的属性(Mariotte et al., 2012).群落结构对环境做出的响应是优势种变化的集中体现, 而优势种的生理生态反应机制影响着群落结构和生态系统对环境变化的响应(Liu et al., 2012; 张梓瑜等, 2013).因此, 了解优势种对N添加的生理生态响应是理解草地生态群落和生态系统过程的关键, 能真实地反映不同N添加浓度下优势种对气候环境因子变化的响应(Anderson et al., 2006; Makhnev & Makhneva, 2010).羊草(Leymus chinensis)是我国温带草原生态系统的优势种, 为多年生根茎型的C3植物, 具有适口性好、营养价值高的特点(Xu & Zhou, 2006).目前, 关于N添加对羊草影响的研究主要集中在化学计量关系、N水利用效率权衡机制、地上地下生物量分配、微生物群落结构和功能、根系存活和寿命以及营养再吸收(Bai et al., 2008; Zhang et al., 2010; Xiao et al., 2011; Li et al., 2014; Yao et al., 2014; Zhang et al., 2016).羊草对N添加的生理生态响应机制也因施肥剂量、实验地点以及时间长短而异(Chen et al., 2005; 肖胜生等, 2010; 白雪等, 2014; 詹书侠等, 2016).因此, 有必要进一步探讨羊草光合特性对N添加的响应机制. ...
Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization. 4 2009
... 在陆地生态系统中, 氮(N)元素不仅是植物需求量最大的营养元素, 也是植物生长和光合作用的主要限制因子(Zhao & Liu, 2009; Krause et al., 2012; Wang & Liu, 2014; Du et al., 2016).近年来, 受人类活动和气候变化的影响, 我国北方草地生态系统的结构和功能受到长期过度放牧的影响而严重退化, 并且表现为土壤中可利用性N严重不足, 其生态系统不同组织水平对于N输入的变化较其他生态系统更为敏感(王仁忠, 1997; 陈佐忠和汪诗平, 2000; 王玉辉等, 2002; Zhang & Han, 2008; Liu et al., 2011; 白雪等, 2014; Chen et al., 2015).而大气N沉降日益加剧, 使得土壤中可利用性N显著增加(Clark & Tilman, 2008; Du et al., 2014).N沉降可以缓解生态系统的N限制, 促进植物光合作用, 提高植物的生产力和碳储存(Aber et al., 2003; Bai et al., 2009).N添加是恢复退化草地生态系统的一种有效手段(詹书侠等, 2016).因此, 研究草原植被对养分添加的响应和适应机制对草地生态系统的管理和恢复具有重要意义. ...
... 叶绿素在绿色植物光合作用中发挥着重要作用, 实现光能到化学能的转化, 为碳同化积累同化力(张云海等, 2013).叶绿素荧光对研究光系统对光能的吸收、传递、耗散、分配等具有独特的作用, 可以反映植物光合作用与环境之间相互关系的内在特性(李晓等, 2006).N元素是光合器官的组成成分, 叶片中有30%-40%的N参与到羧化反应中, 其中分配给羧化系统的N元素比例决定了最终的光合效率, 而叶片缺N将导致植物光合速率、光合量子效率和与碳代谢相关的酶类活性降低, 影响着植物叶片最大羧化速率(Vcmax) (Warren et al., 2000; 郭二辉等, 2008; Nicodemus et al., 2008; 朱军涛等, 2010).然而, 关于解释N沉降影响植物生长的光合生理机制仍有争论.土壤N含量主要受N沉降的速率和持续时间的影响, 而一些地区正从自然状态下的N限制转向富营养化甚至饱和(Aber et al., 2003; Krause et al., 2012).而N沉降对植物生长的促进和抑制作用均有报道, 可能主要依赖于N沉降是否引起了“N饱和” (Wallace et al., 2007; Zhao & Liu, 2009; Thomas et al., 2010).因此, 有必要确定当地是否出现了“N饱和”以及羊草最适合的施N量. ...
 , 罗亲普
, 罗亲普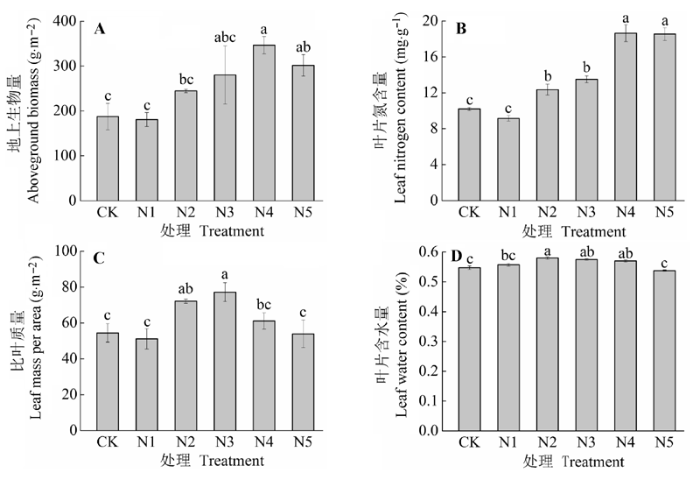 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT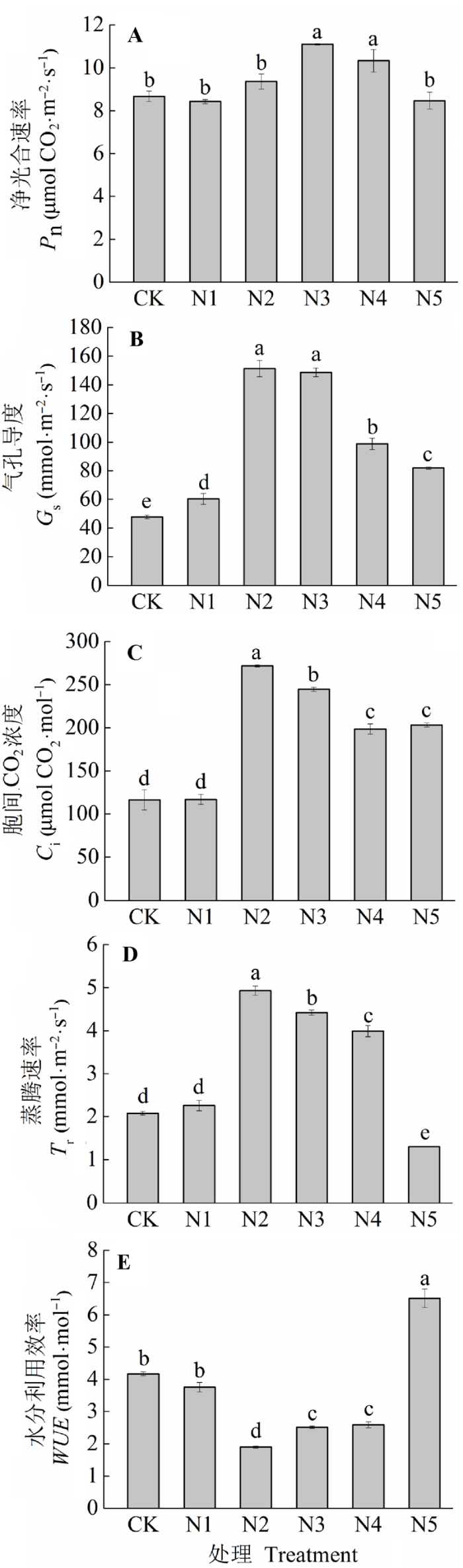 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT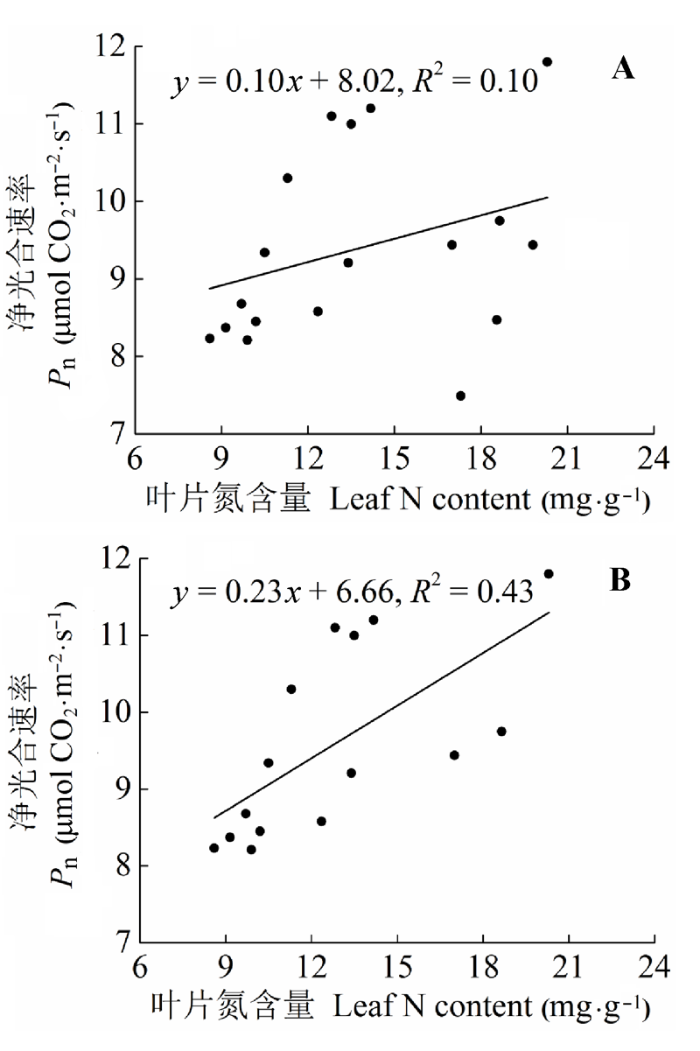 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT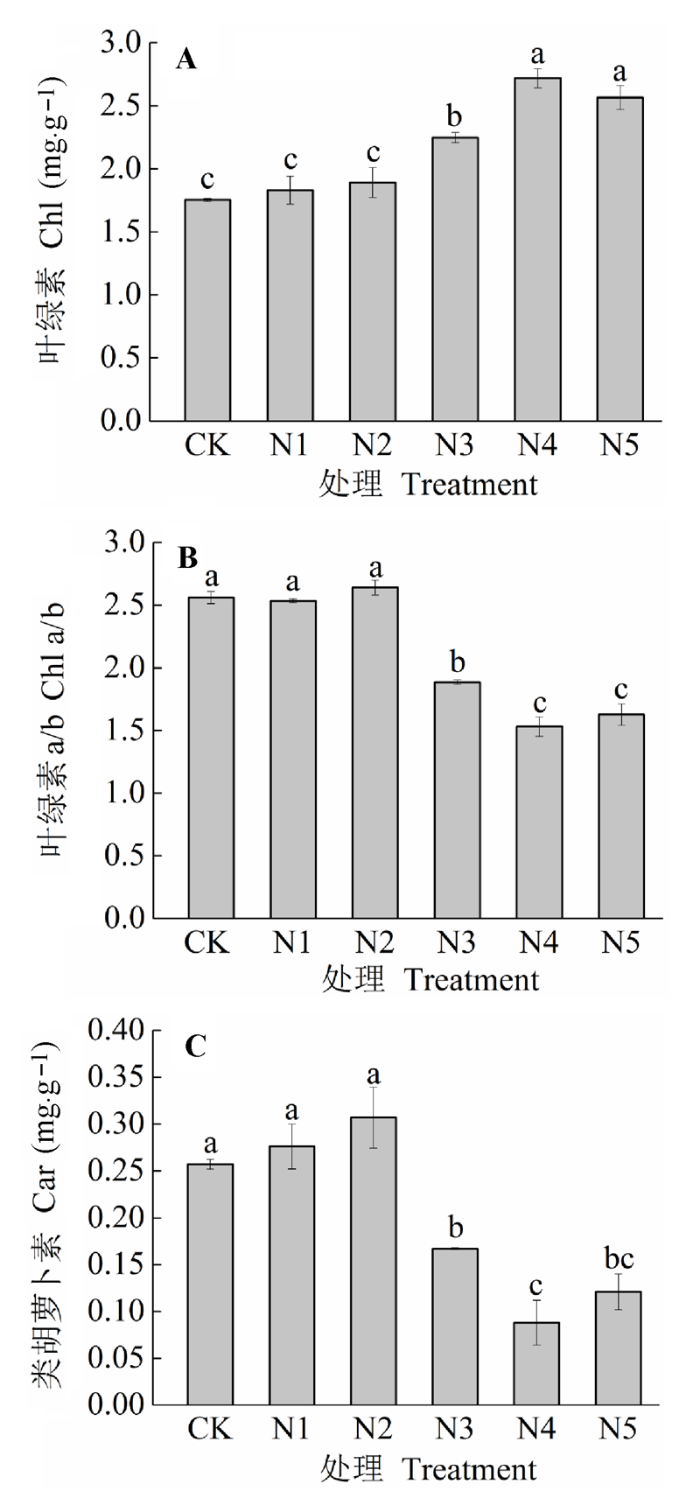 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}