0 引言
【研究意义】株高作为冬小麦的一个重要农艺性状,其变化与抗倒伏性、收获指数、产量和生物量等密切关联[1]。冬小麦在分蘖[2]、越冬[3]或拔节期[4]受到霜冻胁迫后,株高均可能出现降低现象。拔节后霜冻通常称为晚霜冻,在我国频繁发生于黄淮麦区,此时冬小麦抗冻性相比前期明显变差[5],因而产量损失风险更大。但由于冻害发生日期距小麦成熟仍有一段时间,其自身会通过后期生长恢复、以及提高穗数(新增分蘖)和粒重等补偿方式,在一定程度上弥补产量损失,从而对晚霜冻害影响评估及预测造成困难[4, 6-7]。相比之下,株高是一个稳定的农艺性状指标,不仅易于在全生育期进行原位观察和测定,而且有特定的节间伸长规律,并在开花后定长[8],这为晚霜冻害评估指标和方法研究开辟了一条可行的途径。【前人研究进展】晚霜冻发生在冬小麦拔节至成熟的各个阶段。WOODRUFF等[9]认为抽穗前冠层气温低于-6℃即可使幼穗和茎发生明显伤害症状。此时,幼穗因其不同部位冻害而形成各种残穗,致使穗粒数和产量降低[4, 10-11]。茎的冻害症状则往往出现在接近于节的位置,其组织首先变水渍状,随后皱缩和变干,甚至直接因破裂而发生倒伏[12],并最终枯死。对于没有出现明显冻害症状的茎而言,仍能继续生长并为籽粒形成输送营养物质[13]。尽管如此,继续生长的茎仍可能受到一定程度的隐性伤害,主要表现在两个方面:一是营养传导能力下降,导致穗部发育受限,并影响粒重[14];二是节间缩短,株高相应降低[4, 15]。WHALEY等[4]通过分析大田自然霜冻条件下小麦冻害与节间关系,认为雌雄蕊原基分化至药隔形成期冻害阻碍了小麦穗下节间、倒二节间和倒三节间的伸长,因此使其长度缩短。对于大田降温环境而言,直接导致茎在短时内破裂倒伏的现象较少出现,后期群体株高降低特征明显。【本研究切入点】株高降低与其构成因素(即节间和穗的长度)的贡献密不可分,并对产量变化具有重要指示意义。针对黄淮麦区晚霜冻害主要发生在拔节至抽穗期这一特点[16,17],前人研究并未系统考虑不同发育进程下、各种降温程度引起的株高及其构成因素降低规律,也较少涉及冻害胁迫下株高与籽粒产量的定量分析,因而对其内在关系仍不甚清楚。【拟解决的关键问题】本研究利用低温室和田间可移动式霜箱2种霜冻模拟手段,以发育进程和胁迫温度为2个梯度来设计控制试验,研究株高降低特性及与其构成因素、籽粒产量的关系,以期为晚霜冻害动态监测与评估提供思路和依据。1 材料与方法
1.1 试验设计
基于低温室模拟霜冻的试验地点位于商丘市双巴境内小麦试验基地,基于移动霜箱模拟霜冻的试验地点位于商丘市农林科学院南侧小麦试验田。两者管理方式均为大田常规管理,土壤类型为潮土,土壤肥力水平可确保冬小麦正常生长获得所需的营养。相关要素如表1所示。Table 1
表1
表1晚霜冻模拟试验相关要素
Table 1Relevant elements of late frost simulation test
| 晚霜冻模拟手段 Simulating method of late frost | 试验年份 Experimental year | 冬小麦品种 Winter wheat varieties | 种植方式 Planting pattern | 处理温度 Treatment temperature |
|---|---|---|---|---|
| 低温室模拟 Cold Climate Chamber simulating | 2015-2016 | 周麦22 ZM22 | 盆栽种植 Pot planting | -1℃、-3℃、-5℃、-7℃、-9℃、-11℃ |
| 田间可移动式霜箱模拟 Field Movable Climate Chamber simulating | 2011-2012 2013-2014 | 豫麦18、周麦18和偃展4110 YM18, ZM18 and YZ4110 周麦22 ZM22 | 小区种植 Plot planting | -1℃、-3℃、-5℃、-7℃、-9℃ |
新窗口打开
1.1.1 基于低温室的晚霜冻模拟试验 供试小麦品种为周麦22(ZM22),半冬性,中熟。试验用盆为直径25 cm、高度35 cm、两头开口的圆柱型空心管,播种前先按序排列埋于大田之中,使其顶部与地表基本持平,这样做的目的是尽量保证盆栽冬小麦生长环境等同于大田。土壤过筛后自然风干,施肥后搅拌均匀,再分别装盆、浇水。于2015年10月5日播种,每盆均匀定株11粒。
每次降温处理前,随机拔取3株盆栽小麦,在电子显微镜下观察幼穗所处分化阶段,并记录。将参与试验的小麦盆搬至低温室内,做好冻前准备工作。低温室的长、宽和高分别为7 m、5 m和2.6 m,在其顶部安装2条尼龙布形成的通风管道,分别与低温室外的两台制冷压缩机相连。在管道两侧壁钻出直径为5 cm且均匀分布的若干圆形通风口,冷风通过圆形通风口在1.9 m以上的水平方向上进行流动并形成制冷空间,冷空气均匀下沉,从而形成强辐射降温[18]。降温处理设计如表2所示。一个降温过程持续6 h,采用温控系统来模拟自然降温过程,以确保冻害影响机制类似于大田环境[18]。降温处理完成之后,再将小麦盆搬回大田原处,恢复原来生长环境。试验1至试验4的每个降温处理有3盆重复,1盆对照,试验5和试验6的每个降温处理有4盆重复,1盆对照。
Table 2
表2
表2基于低温室的降温处理设计
Table 2Design of decreasing-temperature treatment based on Cold Climate Chamber
| 试验 Experiment | 处理日期 Treatment date | 处理温度及时间间隔 Treatment temperature and time interval | 主茎幼穗分化阶段 Differentiation stage of young ear in the main stem |
|---|---|---|---|
| 试验1 Exp.1 | 03-20-03-22, 2016 | -11℃: 12:00-18:00; -9℃: 18:30-00:30; -7℃: 01:20-07:20; -5℃: 09:00-15:00; -3℃: 15:30-21:30; -1℃: 22:00-04:00 | 小花原基分化期 Floret primordia differentiation phase |
| 试验2 Exp.2 | 03-25-03-27, 2016 | -11℃: 12:30-18:30; -9℃: 18:30-00:30; -7℃: 01:00-07:00; -5℃: 08:30-14:30; -3℃: 14:30-20:30; -1℃: 21:00-03:00 | 雌雄蕊原基分化期 Pistil and stamen primordia differentiation phase |
| 试验3 Exp.3 | 03-30-04-01, 2016 | -11℃: 21:30-03:30; -9℃: 15:30-21:30; -7℃: 09:00-15:00; -5℃: 16:00-22:00; -3℃: 22:30-04:30; -1℃: 10:00-16:00 | 药隔形成前期 Early anther connective tissue formation phase |
| 试验4 Exp.4 | 04-05-04-07, 2016 | -11℃: 12:30-18:30; -9℃: 19:30-01:30; -7℃: 02:00-08:00; -5℃: 09:00-15:00; -3℃: 19:00-01:00; -1℃: 01:00-07:00 | 药隔形成后期 Late anther connective tissue formation phase |
| 试验5 Exp.5 | 04-10-04-12, 2016 | -9℃: 19:00-01:00; -7℃: 01:00-07:00; -5℃: 12:00-18:00; -3℃: 19:00-01:00; -1℃: 01:00-07:00 | 四分体时期 Tetrad formation phase |
| 试验6 Exp.6 | 04-15-04-16, 2016 | -9℃: 09:00-15:00; -7℃: 17:00-23:00; -5℃: 23:00-05:00; -3℃: 10:30-16:30; -1℃: 16:30-22:30 | 抽穗期 Heading phase |
新窗口打开
1.1.2 基于田间可移动式霜箱的晚霜冻模拟试验 2011—2012年度供试小麦品种为豫麦18(YM18)、周麦18(ZM18)和偃展4110(YZ4110),其中豫麦18和偃展4110为弱春性早熟品种,周麦18为半冬性中熟品种;于2011年10月8日种植5行,最外行(第1和第5行)作为保护行,第2、第3和第4行分别对应为豫麦18、周麦18和偃展4110。2013—2014年度供试小麦品种为周麦22(ZM22),于2013年10月5日种植5行。所有小麦均以手工点播,行宽14 cm,株距4 cm。
每次降温处理前,随机拔取3株小麦,在电子显微镜下观察幼穗所处分化阶段,并记录。各试验均需去掉最外行,并沿种植行方向划分出边长不超过60 cm且间隔1米的6个样区。其中,5个样区进行降温处理,第6个样区不作任何处理(对照,CK)。田间可移动式霜箱内径为60 cm×60 cm、水平温差小于0.5℃。将该霜箱完全覆盖样区,四周作密封处理,降温处理过程依据温控系统设定的降温曲线来完成[6, 18]。降温处理设计如表3所示。试验7和试验9的每次降温过程持续6 h,3 h内降至最低设定温度并保持2 h,之后1 h内升至周围环境温度;试验8的每次降温过程持续8 h,4 h内降至最低设定温度并保持3 h,之后1 h内升至周围环境温度。一次降温过程结束之后,移开田间可移动式霜箱,恢复大田生长环境条件。
Table 3
表3
表3基于田间可移动式霜箱的降温处理设计
Table 3Design of decreasing-temperature treatment based on Field Movable Climate Chamber
| 试验 Experiment | 处理日期 Treatment date | 处理温度及时间间隔 Treatment temperature and time interval | 主茎幼穗分化阶段 Differentiation stage of young ear in the main stem |
|---|---|---|---|
| 试验7 Exp.7 | 03-30-03-31, 2012 | -1℃: 6:30 - 12:30; -3℃: 12:30 - 18:30; -5℃: 18:30 - 0:30; -7℃: 0:30 - 6:30; -9℃: 6:30 - 12:30 | 药隔形成后期 Late anther connective tissue formation phase |
| 试验8 Exp.8 | 03-20-03-22, 2014 | -9℃: 17:20 - 01:20; -7℃: 01:20 - 09:20; -5℃: 09:20 - 17:20; -3℃: 17:20 - 01:20; -1℃: 01:20 - 09:20 | 雌雄蕊原基分化期 Pistil and stamen primordia differentiation phase |
| 试验9 Exp.9 | 04-01-04-02, 2014 | -9℃: 17:20 - 23:20; -7℃: 23:20 - 05:20; -5℃: 05:20 - 11:20; -3℃: 11:20 - 17:20; -1℃: 17:20 - 23:20 | 四分体时期 Tetrad formation phase |
新窗口打开
1.2 植株茎部冻害考察与株高、产量测定
在降温处理结束后一周内考察叶片和茎节冻害症状,若出现因变色、水渍或破裂等导致的枯死现象,则进行记录。从试验结束直至小麦成熟,植株未再受各类自然灾害胁迫影响。针对田间样区的每个小麦行,均随机选取10株,分别测定最高茎秆的高度作为株高(Plant height,PH),接着测其穗长(ear length,EL)、穗下节间长(peduncle length,PL)和倒二节间长(penultimate internode length,PIL);针对所有盆栽小麦,处理盆均选取8株,对照盆均选取11株,采取与样区小麦相同测定方法,分别测定株高、穗长、穗下节间长、倒二节间长、倒三节间长(antepenultimate internode length,AIL)和倒四节间长(fourth internode length from the top,FIL)。针对田间样区和盆栽小麦的各个处理,分别统计所有茎蘖数、粒数和粒重,进一步计算单株穗数(ear number per plant,EN)、穗粒数(kernel number per ear,KN)、千粒重(1 000-kernel weight,KW)和单株产量(yield per plant,YP)。1.3 数据处理及分析方法
1.3.1 数据处理 为避免数据记录过程中因人为原因导致的异常值(过大或过小)错误,在此采用平均值加减3倍标准差($\bar{X}$±3δ)方法,对同一处理温度下株高及其构成因素的异常值分别进行识别,将此区间以外数据定为异常值并剔除[19]。重新整理后的各试验中所有单茎数量如下表4所示。Table 4
表4
表4株高及其构成因素数据的样本量
Table 4Sample size of plant height and its components data
| 试验 Experiment | 品种 Cultivar | 处理 Treatment | 对照 CK | 试验 Experiment | 品种 Cultivar | 处理 Treatment | 对照 CK | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 试验1 Exp.1 | 周麦22 ZM22 | 136 | 11 | 试验7 Exp.7 | 豫麦18 YM18 | 50 | 10 | ||||
| 试验2 Exp.2 | 128 | 10 | 周麦18 ZM18 | 49 | 9 | ||||||
| 试验3 Exp.3 | 124 | 11 | 偃展4110 YZ4110 | 48 | 10 | ||||||
| 试验4 Exp.4 | 119 | 11 | 试验8 Exp.8 | 周麦22 ZM22 | 142 | 29 | |||||
| 试验5 Exp.5 | 132 | 11 | 试验9 Exp.9 | 周麦22 ZM22 | 141 | 29 | |||||
| 试验6 Exp.6 | 135 | 9 | |||||||||
新窗口打开
1.3.2 统计分析(1)利用一元线性函数对株高及其构成因素随处理温度降低的变化趋势进行线性拟合,若拟合精度达到0.05、0.01或0.001的显著性水平,则认为呈显著、强显著或极强显著变化。(2)利用方差分析方法(ANOVA)检测各处理温度间株高及其构成因素差异的显著性。一旦方差分析通过显著性检验(P<0.05),则采用Tukey方法(方差齐时)和Dunnett’s T3方法(方差不齐时)进行多重比较。(3)计算单期试验中每个处理的株高及其构成因素的变异系数(coefficient of variation,CV),以比较各处理间离散程度的大小。变异系数的优点在于可以消除株高与其构成因素值相差过大所造成的影响,其计算公式为原始数据标准差与平均值的比值。(4)利用突变检验方法检测株高降低是否存在突变现象,利用逐步回归分析方法建立低温室试验中各构成因素(自变量)与株高(因变量)之间的多元线性回归模型,并依据直接通径系数来比较各因素对株高降低的贡献。(5)利用线性、二次多项式、指数和幂函数等对株高与产量要素间的定量关系进行拟合,并通过比较拟合精度的显著性值大小来确定最佳拟合模型。
1.3.3 突变检验 在确定株高随处理温度呈显著线性降低的前提下,进一步对其是否存在突变进行检验。在此,引入气候突变研究中常用的Mann-Kendall(M-K)检验法。该方法优点在于不需要样本遵从一定的分布,也不受个别异常值的干扰。其设计思路在于检测时间序列上参量的突变性,每个时间点对应一个样本,即“一对一”的关系。本研究若以处理温度作为参考序列,则在同一处理温度下包含多个株高样本,即“一对多”的关系。因此,仅利用M-K检验法来检测株高随处理温度变化的突变点是不可行的。为解决这一问题,我们采用M-K检验和高阶函数拟合相结合的办法。具体计算步骤如下:
首先,对于具有n个总样本量的处理温度序列,构造一秩序列:
${{S}_{k}}=\sum\nolimits_{i=1}^{k}{{{r}_{i}}{{r}_{i}}=\left\{ \begin{matrix} 1{{x}_{i}}>{{x}_{j}} \\ 0{{x}_{i}}\le {{x}_{j}} \\\end{matrix}1\le j\le i2\le k\le n \right.}$ (1)
$E[{{S}_{k}}]=\frac{k(k-1)}{4}\operatorname{var}[{{S}_{k}}]=\frac{k(k-1)(2k+5)}{72}$ (2)
式中,x为株高,Sk表示第i个样本xi > xj的累计值,E[Sk]为均值,var[Sk]为方差。
其次,计算统计量:
$U{{F}_{k}}=\frac{{{S}_{k}}-E[{{S}_{k}}]}{\sqrt{\operatorname{var}[{{S}_{k}}]}}$ (3)
$U{{B}_{k}}=-U{{{F}'}_{k}}(n-k)$ (4)
先利用式(3)计算原始处理温度序列下所有样本值的统计量UFk,接着将该序列的样本值逆序排列,并按照式(2,3)计算得到逆序列的统计量UFk′,再根据式(4)计算UBk。给定显著性水平P = 0.05,当统计量的绝对值大于1.96时,表示通过了显著性检验。最后,高阶函数拟合与突变检验:构建所有样本的UFk、UBk与处理温度间的散点关系图,利用高阶函数分别对其进行曲线拟合。经对拟合精度进行0.05显著性水平的检验后,最终得到两条统计量曲线。如果UFk和UBk曲线出现一个交点且在统计量为±1.96的两条临界直线之间(信度区间),则该交点处表示突变开始。如果交点超出信度区间,则采用Yamamoto方法来计算信噪比(SNR),以判断交点前后2个子序列的均值是否有显著差异[20]。
$SNR=\frac{|{{{\bar{x}}}_{1}}-{{{\bar{x}}}_{2}}|}{{{S}_{1}}+{{S}_{2}}}$ (5)
式中,${{\bar{x}}_{1}}$、${{\bar{x}}_{2}}$、S1、S2分别代表交点前后n1、n2 2个子序列的平均值和标准差。本研究中n1、n2均大于10,若SNR>1.0,则认为有突变发生;SNR>2.0则为强突变。
2 结果
2.1 株高变化
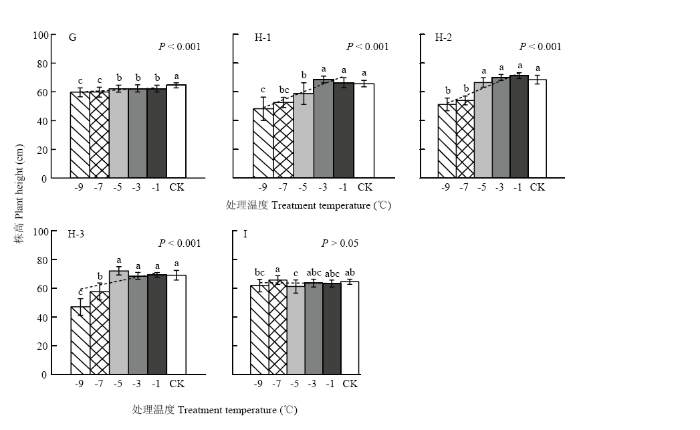
以各期试验开展时冬小麦所处发育进程为序,统计各低温处理下的平均株高,结果如图1和图2所示。可以看出,所有试验的平均株高均在最低处理温度(-11℃或-9℃)下达到最小值,且与对照间表现为显著差异(仅图1-E除外)。其中,试验2(图1-B)、试验3(图1-C)、试验4(图1-D)、试验7(图2-H)和试验8(图2-G)随着处理温度的降低,株高均呈极强显著线性降低趋势(P<0.001)。在试验2、试验3和试验4中,对照株高为73.4、70.7和66.9 cm,-9℃处理下的株高为72.5、68.2和62.8 cm,株高降幅分别为1.3%、3.6%和6.1%;在-11℃处理下的株高为70.2、65.8和56.1 cm,株高降幅分别达4.4%、7.0%和16.2%。在试验8和试验7中,株高在-9℃处理下的平均降幅分别达7.5%和27.9%。就不同品种来看,试验7中豫麦18、周麦18和偃展4110的对照株高为65.7、68.6和69.0 cm,-9℃处理下株高为48.2、51.3和47.0 cm,株高降幅分别达26.6%、25.2%和31.9%。多重比较结果表明(图1—2),试验2、试验3、试验4、试验7和试验8中株高的降低特性是在处理温度降至某一临界温度以下时,才显著开始的。从各项试验结果来看,临界处理温度多集中在-5℃(图1-C、图2-G、图2-H-2和图2-H-3)或-7℃(图1-B和图1-D),品种豫麦18甚至在低于-3℃时开始显著降低(图2-H-1)。与之相对应,株高在-9℃至临界处理温度区间的变异系数明显大于临界处理温度至-1℃区间的变异系数,二者之差在试验2、试验3和试验4中依次为0.026、0.028和0.079,在试验8及试验7的3个品种中依次为0.018、0.080、0.022和0.104。总之,不论是基于低温室还是移动霜箱的霜冻模拟试验,株高在幼穗发育进入雌雄蕊原基分化至药隔形成时期,均呈现出显著降低特性;在不同发育进程、且同一处理温度下,株高降幅随发育进程推移而明显增大,个体差异也越发明显;在相同发育进程、且同一处理温度下,基于移动霜箱的模拟冻害比低温室造成了更大的株高降幅;随着处理温度降低,株高会出现非渐变式(即突变性)降低特征,且不同植株个体、品种间的株高降低差异更加明显。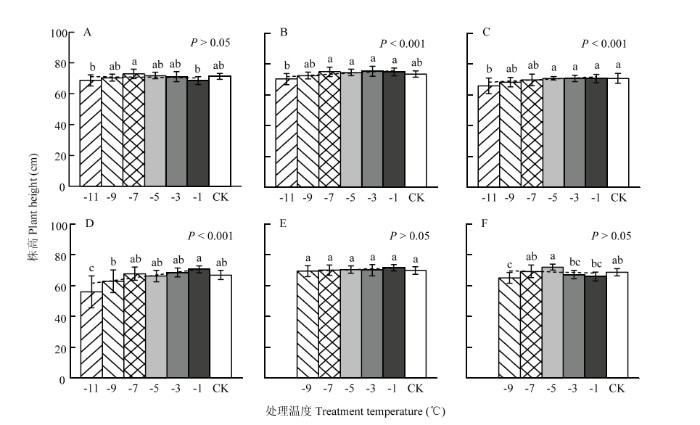 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1基于低温室模拟的晚霜冻胁迫对株高的影响
A―F表示试验1—试验6。误差线表示50倍的变异系数。短划线表示株高随处理温度降低的线性拟合直线,小写字母表示株高在各处理温度间的差异(α=0.05),
-->Fig. 1Effect of late frost stress on plant height based on the Cold Climate Chamber simulation
A-F represent Exp.1-Exp.6. Error bar represents the 50-time coefficient of variation. Dashed line indicates the linear fitting on plant height with the decreasing treatment temperature. Lowercase letter indicates the difference of plant height among treatment temperatures under the significance level of 0.05, the same as
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2基于移动霜箱模拟的晚霜冻胁迫对株高的影响
G、H和I分别表示试验8、试验7和试验9,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110。误差线表示50倍的变异系数
-->Fig. 2Effect of late frost stress on plant height based on the Field Movable Climate Chamber simulation
G, H and I represent Exp.8, Exp.7 and Exp.9, and H-1, H-2 and H-3 represent cultivars Yumai18, Zhoumai18 and Yanzhan4110, respectively. Error bar represents the 50-time coefficient of variation
-->
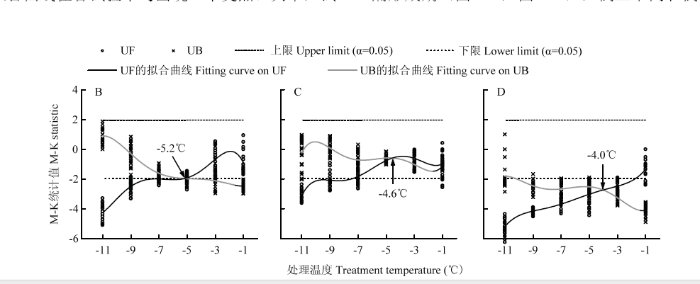
为进一步检验株高降低是否存在突变特性,在此利用M-K突变检验方法,分别计算试验2、试验3、试验4、试验7和试验8中株高的M-K统计量,并对统计量进行不同阶次的函数拟合。经比较,发现低温室试验和移动霜箱试验在函数阶次分别达到5阶和4阶时,其精度不仅达到0.001显著性水平,而且决定系数与显著性P值不再随阶次的升高而发生明显变化。据此,绘制M-K统计量随处理温度变化的拟合曲线,如图3—4所示。可以看出,UF和UB统计量的两条拟合曲线在各试验中均出现一个交点。其中,试验2(图3-B)、试验3(图3-C)、试验8(图4-G)、试验7豫麦18(图4-H-1)和偃展4110(图4-H-3)中的交点均处于信度区间,表明株高在该点处开始突变。试验4(图3-D)和试验7周麦18(图4-H-2)中的交点均位于信度区间之外,其SNR值分别为0.5(<1.0)和2.1(>2.0),据此认为试验4中株高没有发生突变现象,而试验7周麦18的株高则发生了强突变。如将两条统计量拟合曲线交点处的温度称作开始突变温度,则除试验7偃展4110为-7.1℃外,其他试验中株高的开始突变温度均处于-4.6℃—-5.4℃区间。比较移动霜箱试验中的3个小麦品种(图4-H),发现偃展4110的开始突变温度(-7.1℃)明显低于豫麦18(-5.4℃)和周麦18(-5.2℃),这可能表明各品种的抗冻性间存在一定差异。总体来看,不管是基于低温室还是移动霜箱的霜冻模拟试验,冬小麦株高的降低趋势均存在着突变特性。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3基于低温室模拟的晚霜冻胁迫下株高突变检验
B、C和D分别表示试验2、试验3和试验4
-->Fig. 3M-K test for sudden change of plant height under late frost stress simulated by Cold Climate Chamber
B, C and D represent Exp.2, Exp.3 and Exp.4, respectively
-->
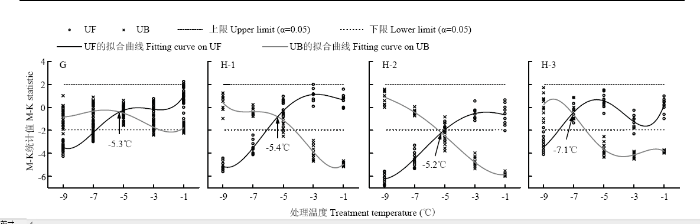 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4基于移动霜箱模拟的晚霜冻胁迫下株高突变检验
G和H分别表示试验8和试验7,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110
-->Fig. 4M-K test for sudden change of plant height under late frost stress simulated by Field Movable Climate Chamber
G and H represent Exp.8 and Exp.7, and H-1, H-2 and H-3 represent cultivars Yumai18, Zhoumai18 and Yanzhan4110, respectively
-->
2.2 株高构成因素变化
针对冬小麦不同发育进程开展的低温室霜冻模拟试验表明(图5),株高构成因素随处理温度降低而变化的趋势与其伸长规律联系密切。在小花原基分化期(图5-A),正值倒五节间(本研究未记录其长度)初伸长、倒四节间待伸长阶段,此时倒四节间长度因霜冻胁迫而强显著缩短,其他因素变化均不显著(P>0.05)。在雌雄蕊原基分化期(图5-B),为倒四节间初伸长、倒三节间待伸长阶段,此时倒三节间和穗长因霜冻胁迫而呈极强显著缩短趋势,倒二节间显著缩短,倒四节间和穗下节间长度变化不明显。进入药隔形成期(图5-C,图5-D),倒三节间和倒二节间先后初伸长,至药隔期末时倒四节间伸长结束,该时期倒四节间、倒三节间、倒二节间长和穗长均因霜冻胁迫而呈缩短趋势,其中除倒三节间长在药隔形成前期(图5-C)不显著外,其他均至少达到强显著水平(P<0.01);穗下节间长度变化不明显。在四分体形成期(图5-E),处于穗下节间初伸长阶段,至四分体期末时倒三节间和穗伸长结束,该时期仅倒三节间长在霜冻胁迫下显著缩短,穗下节间长则表现出强显著延长趋势。冬小麦抽穗时(图5-F),正值穗下节间快速伸长阶段,至开花前倒二节间伸长结束,此时霜冻胁迫仅造成穗下节间显著缩短,倒四节间和麦穗则出现延长趋势。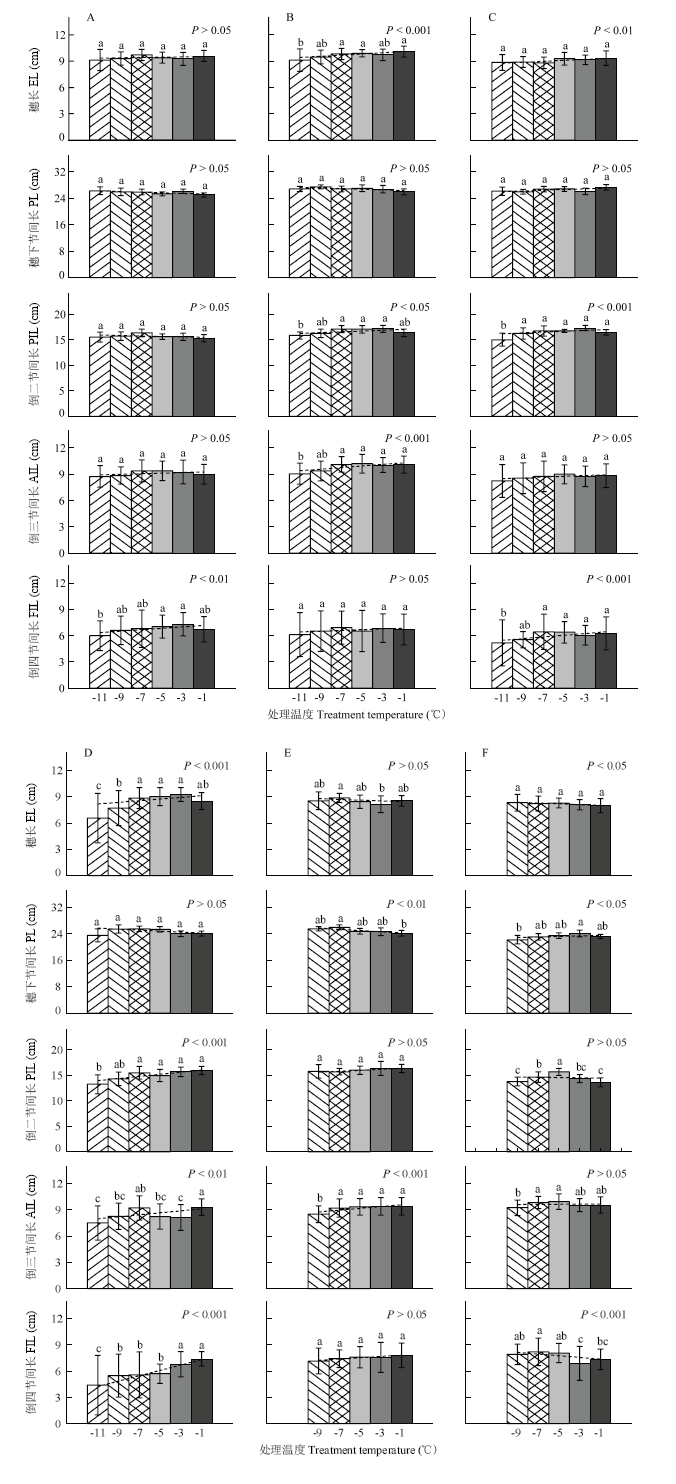 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5基于低温室模拟的晚霜冻胁迫对株高构成因素的影响
A―F表示试验1—试验6。误差线表示10倍的变异系数
-->Fig. 5Effect of late frost stress on plant height’s components based on the Cold Climate Chamber simulation
A-F represent Exp.1-Exp.6. Error bar represents the 10-time coefficient of variation
-->
移动霜箱试验仅考察了穗长、穗下节间长和倒二节间长,其随处理温度降低的变化趋势如图6所示。在雌雄蕊原基分化期(图6-G),穗长和倒二节间长因霜冻胁迫而强显著缩短,穗下节间长则极强显著延长。在药隔形成后期(图6-H),穗长、穗下节间长、以及其中2个品种(豫麦18除外)的倒二节间长均因霜冻胁迫而呈极强显著缩短趋势。在四分体时期(图6-I),除穗长强显著缩短外,穗下节间和倒二节间变化均不明显。总体来看,不论是低温室试验还是移动霜箱试验,植株节间长(穗下节间除外)、穗长在各发育进程霜冻胁迫下大体保持了一致的缩短规律,即显著缩短趋势仅发生在其伸长或者待伸长阶段,伸长结束后则无此现象。
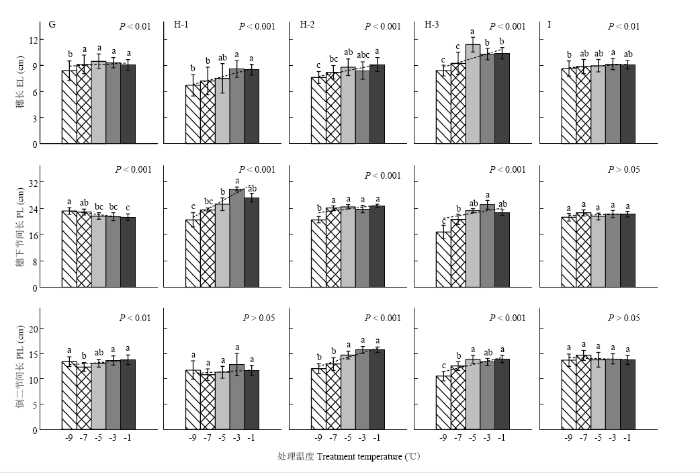 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6基于移动霜箱模拟的晚霜冻胁迫对株高构成因素的影响
G、H和I分别表示试验8、试验7和试验9,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110。误差线表示10倍的变异系数
-->Fig. 6Effect of late frost stress on plant height’s components based on the Field Movable Climate Chamber simulation
G, H and I represent Exp.8, Exp.7 and Exp.9, and H-1, H-2 and H-3 represent cultivars Yumai18, Zhoumai18 and Yanzhan4110, respectively. Error bar represents the 10-time coefficient of variation
-->
比较试验1—试验6中株高构成因素与处理温度的相关系数(表5),可以看出倒四节间长与处理温度间的正相关性在试验1和试验4中最为显著,在试验3中的正相关性仅次于倒二节间长;倒三节间长和处理温度间的正相关性则在试验2和试验5中最显著。这表明冬小麦抽穗前晚霜冻胁迫对倒四节间长和倒三节间长的总体影响是最大的。
Table 5
表5
表5株高构成因素与处理温度的相关系数
Table 5Correlation coefficient of plant height’s components and treatment temperature
| 试验 Experiment | 穗长 EL | 穗下节间长 PL | 倒二节间长 PIL | 倒三节间长 AIL | 倒四节间长 FIL |
|---|---|---|---|---|---|
| 试验1 Exp.1 | 0.080 | -0.175* | -0.112 | 0.100 | 0.261** |
| 试验2 Exp.2 | 0.339*** | -0.127 | 0.181* | 0.308*** | 0.114 |
| 试验3 Exp.3 | 0.270** | 0.132 | 0.323*** | 0.139 | 0.296*** |
| 试验4 Exp.4 | 0.472*** | -0.036 | 0.406*** | 0.259** | 0.626*** |
| 试验5 Exp.5 | -0.139 | -0.249** | 0.149 | 0.297*** | 0.201* |
| 试验6 Exp.6 | -0.186* | 0.175* | -0.073 | 0.047 | -0.294*** |
新窗口打开
2.3 株高与其构成因素的关系
为探究各构成因素在不同低温处理和发育进程下的缩短特性与株高是否存在密切联系,对其相关性大小、方向及显著性水平进行了统计,如表6所示。可以看出,倒四节间、倒三节间、倒二节间长度以及穗长与株高的相关系数在各试验中均至少达到显著正相关(P<0.05);除试验4和试验7(这两个试验均是在药隔形成后期开展的)外,其他试验中穗长与株高的相关系数均明显低于各节间和株高的相关系数;穗下节间长和株高的相关系数除在试验4、试验6、试验7和试验9达到显著正相关外,在其她试验中均无显著正相关性。可见,不论株高在晚霜冻后是否出现显著降低特性,穗下节间除外的其他各构成因素均与株高保持了显著正相关关系。Table 6
表6
表6晚霜冻胁迫下株高与其构成因素的相关系数
Table 6Correlation coefficient of plant height and its components under late frost stress
| 株高构成因素 PH’s component | 试验1 Exp.1 | 试验2 Exp.2 | 试验3 Exp.3 | 试验4 Exp.4 | 试验5 Exp.5 | 试验6 Exp.6 | 试验7 Exp.7 | 试验8 Exp.8 | 试验9 Exp.9 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 豫麦18 YM 18 | 周麦18 ZM18 | 偃展4110 YZ4110 | |||||||||
| 穗长 EL | 0.315*** | 0.422*** | 0.381*** | 0.806*** | 0.433*** | 0.392*** | 0.790*** | 0.525*** | 0.767*** | 0.404*** | 0.189* |
| 穗下节间长 PL | -0.225** | -0.321*** | 0.116 | 0.416*** | 0.160 | 0.183* | 0.841*** | 0.560*** | 0.744*** | 0.009 | 0.404*** |
| 倒二节间长 PIL | 0.724*** | 0.729*** | 0.820*** | 0.886*** | 0.770*** | 0.831*** | 0.441** | 0.845*** | 0.864*** | 0.521*** | 0.783*** |
| 倒三节间长 AIL | 0.748*** | 0.804*** | 0.737*** | 0.727*** | 0.555*** | 0.639*** | |||||
| 倒四节间长 FIL | 0.597*** | 0.786*** | 0.679*** | 0.742*** | 0.562*** | 0.504*** | |||||
新窗口打开
进一步基于低温室试验数据,利用逐步回归建模方法将株高构成因素(穗长、穗下节间长、倒二节间长、倒三节间长和倒四节间长)作为自变量,以构建其与株高间的逐步回归模型,所有模型均通过0.001显著性精度检验。接着计算所有参量因子的直接通径系数,结果如表7所示。为保持各期低温室试验中处理温度范围的一致性,均使用-1℃—-9℃处理的数据。结果表明,除试验1仅有倒二节间长和倒三节间长入选回归模型外,其他试验中各构成因素均入选模型;所有入选因子对株高的影响均呈正效应。如果在试验1中加入-11℃处理,则全部构成因素均能选入模型,这也表明该时期植株的抗冻性明显高于其他各期试验。即使处理温度降低至-11℃,模型中直接通径系数若按大小排序,排前两位的仍为倒二节间长(0.440)和倒三节间长(0.348)。可见,该时期这两个节间长度对株高的影响最大。在试验2—试验4中,直接通径系数最大的依次为倒四节间长(0.529)、倒三节间长(0.488)和倒二节间长(0.426),对株高影响最大的节间刚好与其初伸长时间相对应。试验5—试验6中直接通径系数最大的均为倒二节间长(分别达0.534和0.571),接下来依次是倒四节间长、穗下节间长、倒三节间长和穗长。除了试验4中穗长对株高的影响超过穗下节间长外,在其他试验中穗长的影响均为最小。进一步分析可知,试验1中仅倒四节间显著缩短,此时对株高起影响作用的主要是倒三节间和倒二节间,而这两个节间长度的变化趋势均不显著,因此该时期株高变化不显著;在试验2—试验4中,倒四节间、倒三节间和倒二节间的长度均表现为不同程度的显著缩短趋势,这3个阶段对株高的影响排前两位的分别为倒四节间和倒三节间、倒三节间和倒二节间、倒二节间和倒四节间,可见该时期对株高贡献最大的节间同时也是长度显著缩短的节间,因此株高呈显著降低特性;在试验5和试验6中,仅倒三节间和穗下节间分别表现为显著缩短趋势,这两个节间对株高的影响均明显低于倒二节间和倒四节间,因此该时期株高变化不明显。
Table 7
表7
表7逐步回归模型中各株高构成因素的直接通径系数
Table 7Direct path coefficients of plant height’s components based on stepwise regression model
| 试验 Experiment | 入选自变量的直接通径系数 Direct path coefficient of entered independent variable | 显著性水平 Significant level (P) | ||||
|---|---|---|---|---|---|---|
| 穗长 EL | 穗下节间长 PL | 倒二节间长 PIL | 倒三节间长 AIL | 倒四节间长 FIL | ||
| 试验1 Exp.1 | — | — | 0.459 | 0.483 | — | < 0.001 |
| 试验2 Exp.2 | 0.116 | 0.297 | 0.252 | 0.401 | 0.529 | < 0.001 |
| 试验3 Exp.3 | 0.116 | 0.202 | 0.424 | 0.488 | 0.172 | < 0.001 |
| 试验4 Exp.4 | 0.245 | 0.132 | 0.426 | 0.251 | 0.304 | < 0.001 |
| 试验5 Exp.5 | 0.176 | 0.270 | 0.534 | 0.183 | 0.362 | < 0.001 |
| 试验6 Exp.6 | 0.079 | 0.261 | 0.571 | 0.186 | 0.329 | < 0.001 |
新窗口打开
2.4 株高与籽粒产量的关系
为揭示晚霜冻胁迫下株高降低特性对产量变化的指示意义,本研究统计了试验2、试验3、试验4、试验7和试验8中株高与穗粒数、千粒重、单株产量的相关系数,并且发现株高与穗粒数、千粒重、单株产量间的相关性在所有试验中均至少达到0.01显著水平;相比较而言,株高与单株穗数间的相关性较差,其中试验7中豫麦18和周麦18、试验8中周麦22这3个品种均未达到显著相关(P>0.05)。分别合并低温室模拟(试验2、试验3和试验4)和移动霜箱模拟(试验7和试验8)的霜冻试验数据,构建株高与穗粒数、千粒重、单株产量间的散点关系,并利用线性、二次多项式、指数和幂函数等对其散点关系进行拟合,发现幂函数拟合精度的置信水平都最高,因此作为最终优选函数,其拟合曲线如图7所示。在低温室霜冻模拟条件下,株高与穗粒数、单株产量、千粒重间的幂函数拟合精度均达到0.001显著性(图7-A);在移动霜箱霜冻模拟条件下,株高与穗粒数(图7-B1)、单株产量(图7-B2)的幂函数拟合精度达到0.001显著性,株高与千粒重(图7-B3)的拟合精度达到0.01显著性。对基于低温室模拟和移动霜箱模拟的两类霜冻试验进行比较(图7-A和图7-B),发现其既有相同之处,又有不同之处。相同之处在于:(1)株高与千粒重间的拟合精度均明显低于株高与穗粒数、单株产量间的拟合精度;(2)随着株高降低,穗粒数、单株产量和千粒重均呈降低趋势,其中单株产量的降幅由大变小,直至变化不甚明显;(3)与药隔形成期相比,雌雄蕊原基分化期霜冻胁迫后的株高、穗粒数、单株产量和千粒重的值相对更高,分布也更为集中(如图7-A中绿色正方形符号和图7-B中粉色倒三角形符号所示)。不同之处在于:(1)在移动霜箱霜冻模拟试验中,株高与穗粒数、单株产量、千粒重间的幂函数拟合精度比低温室霜冻模拟试验相对较低;(2)在低温室霜冻模拟试验中,穗粒数降幅随株高降低亦呈现由大变小的态势,但在移动霜箱霜冻模拟试验中无此规律。
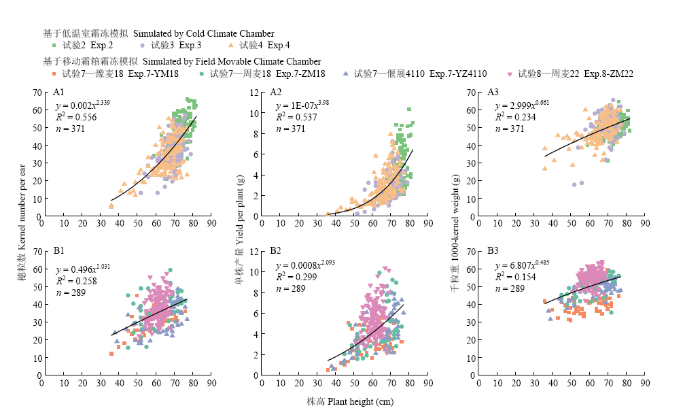 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7株高与穗粒数、单株产量、千粒重间的幂函数关系
黑色实线为基于幂函数的拟合曲线,n为样本量
-->Fig. 7Power function relationship between plant height and kernel number per ear, yield per plant, and 1000-kernel weight
Black solid line presents a fitting curve based on the power function, and n is the sample size
-->
3 讨论
3.1 株高降低特性及其限定条件
关于晚霜冻胁迫下冬小麦株高的降低特性,在已有研究中取得了一定共识。这些研究从发育进程、降温强度、品种与熟性等限定条件入手,认为不同小麦品种的株高在设定低温处理(如-2℃或-3℃)下,随着胁迫时间(如24、48或72 h)延长而显著降低,且多以药隔期之后研究为主[21,22]。针对大田自然晚霜冻的考察结果也表明,株高随冻害程度加深而明显降低[4, 23]。尽管模拟霜冻研究与自然霜冻研究结论相一致,但是利用超过20 h的低温胁迫时长作为冻害程度控制指标,显然不符合单次自然霜冻过程中的实际降温时长规律。为此,本研究从遵循我国黄淮麦区晚霜冻实际发生时间、降温幅度与时长以及当地主栽品种等的现实性出发,在开展霜冻模拟试验时充分考虑了5点设计原则。一是将试验地点放在晚霜冻频发的河南省商丘地区,以确保冬小麦整个生育期处在当地农田生长环境中。二是试验日期涵盖了拔节后幼穗分化的各个阶段。其中,基于低温室模拟的霜冻试验共开展了6期(试验1—试验6),从小花原基分化开始直至抽穗时结束,历时28 d;基于移动霜箱模拟的霜冻试验共开展了3期(试验7—试验9),从雌雄蕊原基分化至四分体形成时结束。此处必须说明的是,尽管试验7和试验9的时间仅相差1 d,但试验7开展时幼穗分化正处于药隔形成后期,而试验9时已进入四分体形成期,这可能是由于试验9冬小麦播种时间更早,且2014年3月至4月间平均气温比2012年同期高出1.5℃,其发育进程更快。三是设定包括-1℃、-3℃、-5℃、-7℃、-9℃和-11℃在内的处理温度序列,并利用温控系统模拟自然降温过程(如前述1.1节所示),使得每次试验过程均能造成植株由未见受冻到明显死亡的各种程度冻害。四是从当地主栽品种中挑选了2个半冬性中熟品种和2个弱春性早熟品种,以期探究品种间差异。五是利用田间可移动式霜箱和低温室2种手段,分别在农田原位(小区种植)和室内(盆栽种植)进行晚霜冻模拟试验,试验结果用于相互佐证株高的降低特性。由于移动霜箱试验的小麦田块位于城区边缘,热岛效应明显,而低温室试验的小麦田块位于野外,因此移动霜箱试验小麦的发育进程要早于低温室试验小麦。此外,在同一处理温度下,移动霜箱会比低温室造成更大的小麦冻害损伤,这可能与其降温空间狭小有关。基于以上限定条件开展晚霜冻模拟试验的结果表明,在雌雄蕊原基分化至药隔形成期,两类品种的株高均随处理温度降低而呈显著降低趋势,并且在低于-4.6℃—-5.4℃区间时出现明显突变现象。突变现象表明冬小麦植株的抗冻能力存在一定阈值区间。AL-ISSAWI等[24]和FULLER等[25]研究认为在二棱期/小花原基分化之后,小麦植株可以抵抗-5℃以上冠层低温的侵袭而不受冻,WOODRUFF等[9]和LIVINGSTON等[26]则认为冠层温度低于-6℃即可使幼穗和茎秆发生明显伤害症状。不论是-5℃还是-6℃低温,都恰恰与本研究中株高降低的突变温度区间相接近。本研究对株高显著降低的试验2、试验3、试验4、试验7和试验8中植株茎部冻害进行考察的结果也表明,在-1℃—-5℃处理下,植株叶片和茎节均未见明显外在冻害症状;在-7℃处理下,茎、叶冻害症状出现;在-9℃—-11℃处理下,茎、叶冻害比重明显增加。
3.2 节间缩短对株高降低特性的贡献
株高构成因素在冬小麦发育的不同时期伸长,共同影响株高性状[27],进而关系到小麦产量变化[28]。因此,揭示晚霜冻胁迫下株高降低特性与规律,离不开对其构成因素变化的分析。刘立伟等[22]认为药隔前期低温胁迫下倒四节间、倒五节间和穗的长度缩短明显,倒三节间,倒二节间和穗下节间长度缩短相对较不明显;WHALEY等[4]的研究结论则表明,药隔后期冻害主要阻碍了穗下节间和倒二节间的伸长,其次是倒三节间。可见,晚霜冻害主要对伸长期或者待伸长期的节间与穗的长度构成影响,并导致其显著缩短,本研究结论也清楚地表明了这一点。但是从试验1—试验9结果来看,节间因冻害缩短现象并非总与株高降低保持同步。试验1、试验5和试验6开展时对应伸长的倒四节间、倒三节间和穗下节间均出现缩短现象,但是株高并未显著降低。因此,株高降低与否还需考虑各构成因素对其综合影响。正如图5和表7结果所示,只有因冻害缩短的节间与对株高起决定性贡献的节间保持一致时,株高才表现为显著降低特性。另外,经比较发现,低温室试验和移动霜箱试验中穗下节间长的变化差异较大。在低温室试验中,穗下节间长在药隔期变化不明显(图5-F),与刘立伟等[22]研究结论一致;在移动霜箱试验中,穗下节间长在药隔期表现为极强显著缩短趋势(图6-H),与WHALEY等[4]研究结论一致。在两类试验中,穗下节间长在四分体时期变化均不显著,而该时期正是穗下节间初伸长阶段,似乎应该出现缩短现象。究其原因,就目前试验结果还无法作出进一步合理解释,因此有必要开展更多类似试验以完善相关研究结论。3.3 节间缩短与幼穗冻害对产量的共同影响
株高与穗粒数、千粒重、单株产量之间的显著相关性表明,利用株高降低幅度作为评判指标,可提早预测雌雄蕊原基分化至药隔形成期霜冻胁迫下的冬小麦产量损失风险。从株高与穗粒数、千粒重、单株产量间的幂函数拟合曲线(图7)来看,最大亮点在于单株产量降幅随株高降低而呈现出先大后小态势。当单株产量降低至约1.5 g以下时,不会再随株高继续降低而明显降低,主要原因可能在于此时的穗粒数变化已不大。总体上看,基于低温室试验数据拟合的精度要明显高于移动霜箱试验数据,这可能是由于低温室试验仅使用了1个品种,而移动霜箱试验中涉及4个品种,其株高范围和抗冻能力有所不同。若要探明节间缩短与幼穗冻害对产量的影响机制,需先对晚霜冻胁迫下冬小麦茎、叶和幼穗冻害规律有一个清晰的认识。我们在前面探讨了雌雄蕊原基分化至药隔形成期株高降低特性的突变温度区间,冠层气温低于该温度区间时茎和幼穗均因结冰而发生显著冻害。LIVINGSTON Ⅲ等[29]研究认为植株结冰总是先从茎的基部开始,并逐渐向上扩展至叶片,进而至幼穗。发生冻害的植株会迅速死亡,以致无法形成结实穗。但并非冠层气温低于该温度区间时植株就会结冰并死亡,此时仍有相当一部分植株会因处于过冷却状态而恢复生长,并最终形成不完全结实的穗。FREDERIKS等[14]则认为,即使冠层气温高于该温度区间(此时茎部看上去完好无损),包在叶鞘内部的幼穗也仍有可能发生冻害,并会形成不完全结实的穗。可见,在高于突变温度区间的晚霜冻胁迫下,茎节通常不会受冻,因而株高降低不明显(图1,图2),此时籽粒产量损失原因主要表现为穗粒数的下降;在低于突变温度区间的晚霜冻胁迫下,不仅穗粒数下降,群体成穗数也会因部分植株的死亡而显著降低[4, 30],这从试验2、试验3和试验4的结果也可以看出来,与对照(总株数均为48)相比,茎蘖死亡率在-7℃处理下分别为0、39.6%和41.7%,在-9℃处理下分别为27.1%、35.9%和43.8%,在-11℃处理下分别为31.3%、43.8%和45.8%。过冷却的植株恢复生长后,株高显著降低,原因在于对其起关键影响作用的节间发生了显著缩短现象。节间缩短的同时也会造成籽粒发育受损,进而影响产量。这主要表现在两个方面:一是茎的营养传导功能受限[12],导致籽粒皱缩、粒重降低[14];二是受冻茎秆相比正常而言变得更为纤弱,因而容易在生长后期发生倒伏[31]。研究表明[32,33],小麦倒伏一般发生在茎的基部第二节间(倒四节间)和第三节间(倒三节间)。这一结论恰好与表5所示结果相对应,即受晚霜冻影响最大的节间亦是最易发生倒伏的节间。可见,利用倒四节间长或倒三节间长的缩短规律来评估/预测晚霜冻胁迫下冬小麦倒伏风险与产量损失,具有潜在应用价值。
4 结论
在模拟晚霜冻条件下,正在伸长或待伸长的冬小麦节间长度与穗长随处理温度降低而显著缩短;当缩短节间与影响株高的关键节间相一致时,株高则降低。在雌雄蕊原基分化至药隔形成期,倒四节间长、倒三节间长和倒二节间长均显著缩短,并且对株高的总体贡献最大,因此株高表现出显著降低趋势;当处理温度低于-5℃左右时,部分植株的茎部因出现明显冻害症状而死亡,存活植株的整体高度急剧降低,株高在该温度值附近存在突变特性,且不同品种间有一定差别。株高与穗粒数、千粒重、单株产量之间存在幂函数曲线关系,当单株产量因冻害胁迫而降低至1.5 g以下时,随株高继续降低而不再明显减少,此时穗粒数变化也已不大。The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . 【目的】小麦越冬期低温冻害时有发生,影响产量形成,而前人关于越冬期冻害的补救措施报道较少,因此研究分蘖期受冻小麦施用恢复肥挽回产量的效应,探明增施尿素恢复产量的生理原因,可为小麦抗逆栽培提供理论依据。【方法】试验选用小麦品种扬麦16,利用人工智能控温室,设置昼夜温度-2℃/-6℃(2012年)和-2℃/-8℃(2013年),分别处理24、48、72 h,处理结束后5 d调查植株冻害指数。随后一次性增施恢复肥尿素(N 46%)75、150 kg·hm-2(2012)和75、120、180 kg·hm-2(2013)补救,增施尿素后10、20、30 d取主茎展二叶,测定可溶性糖、游离脯氨酸和内源激素含量变化,成熟期测量株高并收获籽粒测产。【结果】分蘖期低温胁迫后扬麦16受冻指数随着胁迫时间的延长由0.2上升到0.5左右。施肥补救后10 d,所有处理植株可溶性糖、游离脯氨酸以及脱落酸(ABA)和玉米素核甘(ZR)含量随胁迫时间延长呈增加趋势,不施肥处理显著高于施恢复肥处理,相同处理时段,随恢复肥施用量增加上述指标呈下降趋势,赤霉素(GA3)含量呈相反变化趋势;增施尿素后20 d,施肥处理可溶性糖和游离脯氨酸含量、ABA和ZR含量较不施肥处理快速下降,GA3含量上升;施肥后30 d,各低温处理的渗透调节物质和内源激素含量已恢复至接近自然对照水平。随低温处理时间延长,小麦产量、基部Ⅰ、Ⅱ节间长度及株高降低,相同处理时段内,基部节间长度、株高及产量恢复和挽回效应随恢复肥施用量增加而提高。【结论】分蘖期受冻小麦根据受冻指数及时适量施用恢复肥能明显缓解低温伤害,表现为植株渗透调节物质含量下降以及激素更趋于平衡,促进新生分蘖发生和基部Ⅰ、Ⅱ节间伸长,一定程度上挽回了产量的损失。研究兼顾产量挽回效应和氮肥偏生产力,提出分蘖期受冻指数为0.2左右的轻度冻害推17 . 【目的】小麦越冬期低温冻害时有发生,影响产量形成,而前人关于越冬期冻害的补救措施报道较少,因此研究分蘖期受冻小麦施用恢复肥挽回产量的效应,探明增施尿素恢复产量的生理原因,可为小麦抗逆栽培提供理论依据。【方法】试验选用小麦品种扬麦16,利用人工智能控温室,设置昼夜温度-2℃/-6℃(2012年)和-2℃/-8℃(2013年),分别处理24、48、72 h,处理结束后5 d调查植株冻害指数。随后一次性增施恢复肥尿素(N 46%)75、150 kg·hm-2(2012)和75、120、180 kg·hm-2(2013)补救,增施尿素后10、20、30 d取主茎展二叶,测定可溶性糖、游离脯氨酸和内源激素含量变化,成熟期测量株高并收获籽粒测产。【结果】分蘖期低温胁迫后扬麦16受冻指数随着胁迫时间的延长由0.2上升到0.5左右。施肥补救后10 d,所有处理植株可溶性糖、游离脯氨酸以及脱落酸(ABA)和玉米素核甘(ZR)含量随胁迫时间延长呈增加趋势,不施肥处理显著高于施恢复肥处理,相同处理时段,随恢复肥施用量增加上述指标呈下降趋势,赤霉素(GA3)含量呈相反变化趋势;增施尿素后20 d,施肥处理可溶性糖和游离脯氨酸含量、ABA和ZR含量较不施肥处理快速下降,GA3含量上升;施肥后30 d,各低温处理的渗透调节物质和内源激素含量已恢复至接近自然对照水平。随低温处理时间延长,小麦产量、基部Ⅰ、Ⅱ节间长度及株高降低,相同处理时段内,基部节间长度、株高及产量恢复和挽回效应随恢复肥施用量增加而提高。【结论】分蘖期受冻小麦根据受冻指数及时适量施用恢复肥能明显缓解低温伤害,表现为植株渗透调节物质含量下降以及激素更趋于平衡,促进新生分蘖发生和基部Ⅰ、Ⅱ节间伸长,一定程度上挽回了产量的损失。研究兼顾产量挽回效应和氮肥偏生产力,提出分蘖期受冻指数为0.2左右的轻度冻害推17 |
| [3] | . |
| [4] | . One winter wheat variety, Soissons, suffered extensive frost damage in a plant population density (PPD) and sowing date (SD) experiment in 1997/1998. Another variety, Cadenza, did not incur any frost damage, even though it was at a similar developmental stage to Soissons when the damaging frosts (minimum grass temperatures of 618 and 619 °C) occurred. Therefore, in addition to rapid development, other physiological characteristics of Soissons, such as the presence of awns and weak glaucosity must be important in determining its susceptibility to frost damage. When sown on 23 September, the low PPD crops suffered the most frost damage but when sown on 25 October the high PPD crops suffered the most. This challenges the assumption that low PPDs are more resistant to frost than high PPDs. No frost damage was observed in the crop sown on 16 December. Frosts killed spikelets, restricted internode extension (stem growth) and reduced yield. Frosts in April and May, after growth stage (GS) 33, appeared responsible for the damage symptoms observed in the crop. Minimum temperatures in April and May were similar to those recorded in the previous season when no frost damage was observed. There had been a cooler winter in 1996/1997, allowing slower crop development and more cold hardening, therefore the frosts in the spring of 1997 did not inflict obvious damage. |
| [5] | . Low temperature (LT) stress is one of the primary environmental factors limiting the expansion of winter wheat ( Triticum aestivum L.) production in northern temperate climates. The objective of this study was to clarify the interrelationship between developmental stage and freezing resistance (FR) in winter and spring habit cultivars acclimated under field conditions in regions with long mild winters. Four cultivars with different vernalization and photoperiod requirements were planted in the field at the recommended autumn seeding dates in 2002–2003 and 2003–2004 at Maragheh in north-west Iran. Freezing resistance, as measured by LT50, and stage of phenological development, as estimated from final leaf number (FLN) and shoot apex developmental morphology, were determined during the autumn and winter seasons. ‘Kohdasht’ spring wheat, which does not have a vernalization or day length requirement, quickly entered the reproductive stage and had a limited ability to LT acclimate. In contrast, a significant decrease in FLN associated with LT treatment and delayed double ridge formation under field conditions indicated that ‘Norstar’, ‘Sardari’ and ‘Azar2’ had vernalization responses. Sardari increased its FLN from 9 to 12 when grown at 16 h compared to 12 h day indicating that it was also day length sensitive. Azar2 had little day length sensitivity, but it had a higher unvernalized FLN with the result that both Sardari and Azar2 achieved their vegetative/reproductive transition and maximum FR (656115 °C) by early December. Norstar had a longer vernalization requirement and reached vernalization saturation and maximum FR (656128 °C) near the end December. The delay in the phenological development associated with a vernalization requirement, photoperiod sensitivity and/or increased leaf numbers was accompanied by increased expression of LT tolerance confirming that the length of the vegetative phase determines a plants ability to maintain a high level of LT tolerance gene expression. These observations suggest that more detailed studies should be initiated to establish the importance of interactions among factors that influence the time to vegetative/reproductive transition with the objective of identifying genetic combinations and management systems that extend the vegetative stage and provide longer term protection from LT stress in regions with long mild winters like those normally experienced in the north-west Iran. |
| [6] | . The influence of frost stress during the jointing stage in winter wheat was examined for original tillers (OT), regenerated tillers (RT) and the yield components of the cultivars Yumai 18, Zhoumai 18 and Yanzhan 4110. Ear number per plant, kernel number per ear, 1000-kernel weight and yield per plant were recorded under a range of subfreezing temperatures (611, 613, 615, 617 and 619°C). The results were modelled by linear and quadratic regression. In the quadratic model, ear number per plant and yield per plant of the three cultivars were all significant (P < 0.001) and their fitted curves showed a concave-downward parabolic trend as the subfreezing temperature decreased. By analysing the change characteristics of ear number and yield per plant of OT and RT following frost treatment, it was found that OT decreased and RT increased resulting in the concave-downward parabolic yield trend. Final ear number and yield were the result of these opposing behaviours with OT and RT partly offsetting one another and thus tending to stabilize yield loss. The resulting compensatory behaviour of the three cultivars showed obvious differences with the RT of Yumai 18 making the greatest contribution to grain yield. The work offers the potential for creating a rational basis for selecting cultivars having improved frost resistance based on recording their tillering behaviour under low-temperature conditions. |
| [7] | . 以国审矮抗58小麦品种为研究对象,考察其100个田间样点在2013年4月7、10和21日发生的3次自然霜冻过程中的冻害状况。依据受冻穗的长短对冻害进行分级,构建残穗指数(DEI)作为总体冻害程度评价指标,比较每百穗中不同冻害级别的籽粒产量及其构成要素之间的差异,对每百穗中籽粒产量及其构成要素随冻害级别的变化以及1m2内穗数和实际产量随残穗指数的变化趋势进行拟合,以揭示冬小麦产量及其构成要素随晚霜冻害变化的规律。结果表明,每百穗中各级冻害穗数、穗粒数和实际籽粒重均显著低于未受冻穗(P<0.01)。冻害级别越高,穗粒数和实际籽粒重越低,穗数呈先显著降低后小幅回升的变化态势,千粒重先显著增加而后降低(P<0.01)。随着冻害级别提高,每百穗中穗数、穗粒数、千粒重和实际籽粒重分别呈上开口抛物线型、线性函数型、下开口抛物线型和幂函数型降低趋势;残穗指数越大,每平方米内穗数和实际产量越低,二者分别呈幂函数型和线性函数型特点。研究结果对揭示拔节至孕穗期冬小麦霜冻害的影响机理具有指导作用。 . 以国审矮抗58小麦品种为研究对象,考察其100个田间样点在2013年4月7、10和21日发生的3次自然霜冻过程中的冻害状况。依据受冻穗的长短对冻害进行分级,构建残穗指数(DEI)作为总体冻害程度评价指标,比较每百穗中不同冻害级别的籽粒产量及其构成要素之间的差异,对每百穗中籽粒产量及其构成要素随冻害级别的变化以及1m2内穗数和实际产量随残穗指数的变化趋势进行拟合,以揭示冬小麦产量及其构成要素随晚霜冻害变化的规律。结果表明,每百穗中各级冻害穗数、穗粒数和实际籽粒重均显著低于未受冻穗(P<0.01)。冻害级别越高,穗粒数和实际籽粒重越低,穗数呈先显著降低后小幅回升的变化态势,千粒重先显著增加而后降低(P<0.01)。随着冻害级别提高,每百穗中穗数、穗粒数、千粒重和实际籽粒重分别呈上开口抛物线型、线性函数型、下开口抛物线型和幂函数型降低趋势;残穗指数越大,每平方米内穗数和实际产量越低,二者分别呈幂函数型和线性函数型特点。研究结果对揭示拔节至孕穗期冬小麦霜冻害的影响机理具有指导作用。 |
| [8] | . Drought as a major abiotic stress often occurs from stem elongation to the grain filling stage of wheat in northern China. Plant height (PH) is a suitable trait to model the dissection of drought tolerance. The purposes of the present study were to validate molecular markers for PH developmental behavior and identify elite alleles of molecular markers. After the phenotyping of 154 accessions for PH dynamic development under well-watered (WW) and drought stressed (DS) conditions, and the genotyping of 60 SSR markers from six candidate chromosome regions related to PH found in our previous linkage mapping studies, both parameters PH and drought tolerance coefficient (DTC) calculated by the conditional analysis were used for association mapping. A total of 46 significant association signals (P < 0.01) were identified in 23 markers, and phenotypic variation ranged from 7 to 50%. Among them, four markers Xgwm261-2D, Xgwm495-4B, Xbarc109-4B and Xcfd23-4D were detected under both water regimes. Furthermore, 10 markers were associated with DTC, and four with both parameters PH and DTC at the same plant development stage. The results revealed different allelic effects of associated markers; for example, the 155 bp Xgwm495-4B allele was associated with a reduced height of -11.2 cm under DS and -15.3 cm under WW, whereas the 167 bp allele exhibited increased height effects of 3.9 and 8.1 cm, respectively. This study demonstrates a strong power of joint association analysis and linkage mapping for the identification of important genes in wheat. |
| [9] | . |
| [10] | . A late frost occurred in the Southland region of New Zealand in November 1995 when many winter wheat (Triticum aestivum L.) crops were at early grain fill, and resulted in a large number of insurance claims for frost damage. In this study, frost damage was characterised. Spikes in affected crops were sharply tapered from a point half to two thirds from their bases. Grains were as much as 80% lighter in upper parts of affected spikes. Overall yield losses as a result of frost of between 13 and 33% were calculated in affected crops. Grains were sectioned and examined with a scanning electron microscope. Whereas in healthy grains, the layers making up the pericarp and testa were compressed, in frosted grains these layers comprised loosely compressed and unstructured networks of cells. The aleurone layer was less ordered in frosted than in normal grains, and was not always readily distinguished from the starchy endosperm. Rehydration of frosted grains was accompanied by expansion of pericarp and testa cells, whereas little cell expansion occurred in normal grains. Blisters were common in frosted grains, where the outer layers were detached from the remainder of the pericarp. |
| [11] | . . |
| [12] | . |
| [13] | . |
| [14] | . Abstract Radiant frost is a significant production constraint to wheat (Triticum aestivum) and barley (Hordeum vulgare), particularly in regions where spring-habit cereals are grown through winter, maturing in spring. However, damage to winter-habit cereals in reproductive stages is also reported. Crops are particularly susceptible to frost once awns or spikes emerge from the protection of the flag leaf sheath. Post-head-emergence frost (PHEF) is a problem distinct from other cold-mediated production constraints. To date, useful increased PHEF resistance in cereals has not been identified. Given the renewed interest in reproductive frost damage in cereals, it is timely to review the problem. Here we update the extent and impacts of PHEF and document current management options to combat this challenge. We clarify terminology useful for discussing PHEF in relation to chilling and other freezing stresses. We discuss problems characterizing radiant frost, the environmental conditions leading to PHEF damage, and the effects of frost at different growth stages. PHEF resistant cultivars would be highly desirable, to both reduce the incidence of direct frost damage and to allow the timing of crop maturity to be managed to maximize yield potential. A framework of potential adaptation mechanisms is outlined. Clarification of these critical issues will sharpen research focus, improving opportunities to identify genetic sources for improved PHEF resistance. The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com. |
| [15] | . . |
| [16] | . 根据霜冻害的观测和调查资料,进行气候学研究,阐述了我国冬小麦发生霜冻害的生长发育时段、最低气温范围、地区分布规律和年际变化趋势,讨论了防御霜冻害中值得注意的几个间题。 . 根据霜冻害的观测和调查资料,进行气候学研究,阐述了我国冬小麦发生霜冻害的生长发育时段、最低气温范围、地区分布规律和年际变化趋势,讨论了防御霜冻害中值得注意的几个间题。 |
| [17] | . 冬小麦晚霜冻害主要发生在中国的黄淮区域,河南占黄淮区域面积较大,是中国小麦的主要产地,又是晚霜冻害的重发区,因此文章以河南为例,从构成晚霜冻害的最低温度和小麦发育期两个因素出发,提出了晚霜冻害指数构建方法,使晚霜冻害指标定量化,并从多年数据库中计算出逐年晚霜冻害发生强度和发生天数。由于小麦发育期观测资料序列较短,影响了大尺度小麦晚霜冻害变化规律的分析,因而提出了冬小麦发育期资料推算方法;对计算出的近50年晚霜冻害资料进行EOF分析和Morlet小波分析,以发现晚霜冻害的时空分布与多时间尺度变化规律。结果表明:冬小麦晚霜冻害发生频率较高,高发区频率超过40%,且在20世纪70—90年代有增多的趋势;发生强度和天数变化趋势基本相似,具有发生强度较重区对应着发生天数较多区的趋势,且前者对小麦产量的影响大于后者;晚霜冻害发生强度的重发区分布在豫东南和豫西南,轻发区主要分布在淮河以南及豫西部分地区,其中第1模态分布型存在16年、第2模态分布型存在准2年的多时间尺度变化规律;发生天数的多发区主要分布在豫北北部和豫西南边界,少发区在豫东北及豫西南中部,其中第1模态分布型存在着准4年、第2模态分布型存在准3年和准6—7年的周期。 . 冬小麦晚霜冻害主要发生在中国的黄淮区域,河南占黄淮区域面积较大,是中国小麦的主要产地,又是晚霜冻害的重发区,因此文章以河南为例,从构成晚霜冻害的最低温度和小麦发育期两个因素出发,提出了晚霜冻害指数构建方法,使晚霜冻害指标定量化,并从多年数据库中计算出逐年晚霜冻害发生强度和发生天数。由于小麦发育期观测资料序列较短,影响了大尺度小麦晚霜冻害变化规律的分析,因而提出了冬小麦发育期资料推算方法;对计算出的近50年晚霜冻害资料进行EOF分析和Morlet小波分析,以发现晚霜冻害的时空分布与多时间尺度变化规律。结果表明:冬小麦晚霜冻害发生频率较高,高发区频率超过40%,且在20世纪70—90年代有增多的趋势;发生强度和天数变化趋势基本相似,具有发生强度较重区对应着发生天数较多区的趋势,且前者对小麦产量的影响大于后者;晚霜冻害发生强度的重发区分布在豫东南和豫西南,轻发区主要分布在淮河以南及豫西部分地区,其中第1模态分布型存在16年、第2模态分布型存在准2年的多时间尺度变化规律;发生天数的多发区主要分布在豫北北部和豫西南边界,少发区在豫东北及豫西南中部,其中第1模态分布型存在着准4年、第2模态分布型存在准3年和准6—7年的周期。 |
| [18] | . 冬小麦受晚霜冻影响的外部症状短期内不明显的特点, 为红边光谱的应用提供了契机。利用田间移动式霜箱和低温室系统分别开展不同温度梯度的霜冻试验, 基于冻后第1天测定的冬小麦冠层光谱数据, 使用最大一阶导数(FD)、 四点线性插值(FPI)、 多项式拟合(POLY)、 倒高斯拟合(IG)和线性外推(LE)等方法分别提取红边参数。通过相关分析、 线性回归建模以及波动分析, 从早期性、 敏感性和稳定性方面对红边参数检测冬小麦晚霜冻的能力进行定量研究。结果表明, LE方法提取的红边位置(REP)与霜冻温度的相关性为极显著(显著性水平达到0.01), 二者相关系数高于其他方法; REP随霜冻温度的降低而呈蓝移现象, 温度越低, 蓝移趋势越明显。FD方法提取的最小振幅(Drmin)和红边振幅与最小振幅比值(Dr/Drmin)对霜冻敏感性的表现最佳, 其次是FD和IG方法提取的红边振幅与红边面积比值(Dr/SDr), 敏感性最差的是LE方法提取的REP。总的来看, FD方法提取的Drmin和Dr/Drmin检测冬小麦晚霜冻的综合能力最强。以上结论可为基于冠层红边特征的冬小麦晚霜冻害早期诊断研究提供依据。 . 冬小麦受晚霜冻影响的外部症状短期内不明显的特点, 为红边光谱的应用提供了契机。利用田间移动式霜箱和低温室系统分别开展不同温度梯度的霜冻试验, 基于冻后第1天测定的冬小麦冠层光谱数据, 使用最大一阶导数(FD)、 四点线性插值(FPI)、 多项式拟合(POLY)、 倒高斯拟合(IG)和线性外推(LE)等方法分别提取红边参数。通过相关分析、 线性回归建模以及波动分析, 从早期性、 敏感性和稳定性方面对红边参数检测冬小麦晚霜冻的能力进行定量研究。结果表明, LE方法提取的红边位置(REP)与霜冻温度的相关性为极显著(显著性水平达到0.01), 二者相关系数高于其他方法; REP随霜冻温度的降低而呈蓝移现象, 温度越低, 蓝移趋势越明显。FD方法提取的最小振幅(Drmin)和红边振幅与最小振幅比值(Dr/Drmin)对霜冻敏感性的表现最佳, 其次是FD和IG方法提取的红边振幅与红边面积比值(Dr/SDr), 敏感性最差的是LE方法提取的REP。总的来看, FD方法提取的Drmin和Dr/Drmin检测冬小麦晚霜冻的综合能力最强。以上结论可为基于冠层红边特征的冬小麦晚霜冻害早期诊断研究提供依据。 |
| [19] | . 采用长江水系约260个背景样点沉积物原样和小于63μm细粒沉积物中11种重金属元素(铜、铅、锌、镉、汞、钴、镍、砷、铬、锰和铁)含量的数据,研究了数据的频率分布特征和变异系数,比较了算术均值、几何均值和中位数与由Box-Cox变换而得到的对称均值间的大小关系。结果表明,大多数重金属元素含量服从对数正态分布;采用算术均值表示均值会有较大的偏差,且这种偏差随变异系数的增大而增大;采用中位数偏差较小,但效率低;采用几何均值偏差也较小,在数据服从对数正态分布条件下可以使用,但不是最优;而采用对称均值来表示均值,由于分布的对称性,而最具有代表性,本研究采用稳健统计与Box-Cox变换相结合的方法,得到稳健对称均值,并采用此法表示长江水系河流沉积物中重金属元素含量的均值。 . 采用长江水系约260个背景样点沉积物原样和小于63μm细粒沉积物中11种重金属元素(铜、铅、锌、镉、汞、钴、镍、砷、铬、锰和铁)含量的数据,研究了数据的频率分布特征和变异系数,比较了算术均值、几何均值和中位数与由Box-Cox变换而得到的对称均值间的大小关系。结果表明,大多数重金属元素含量服从对数正态分布;采用算术均值表示均值会有较大的偏差,且这种偏差随变异系数的增大而增大;采用中位数偏差较小,但效率低;采用几何均值偏差也较小,在数据服从对数正态分布条件下可以使用,但不是最优;而采用对称均值来表示均值,由于分布的对称性,而最具有代表性,本研究采用稳健统计与Box-Cox变换相结合的方法,得到稳健对称均值,并采用此法表示长江水系河流沉积物中重金属元素含量的均值。 |
| [20] | . . |
| [21] | . . |
| [22] | [D]. [D]. |
| [23] | . 为有效抵御春季低温对小麦冻害的不利影响,提高小麦的抗冻能力,在山西临汾采用大田随机区组 方法研究了春季低温对不同播种期小麦叶片、节间、穗部冻害的影响。结果表明:单株叶片冻害率随着播期的推迟而提高,变幅为3.0%~55.4%,单株分蘖 叶片冻害率高于单株主茎叶片冻害率,二者相差16.6%,单株鲜重、单株绿叶与单株冻叶率相关性高于其他生物学性状;节间冻害呈现随着播期的推迟而趋重, 表现出第1节间轻于第2节间;冻穗率在试验播期范围内则呈现两端轻中间重的现象,以播期10月22日最轻,10月12日最重,变幅为11.58%;同时对 形成处理间冻害差异的原因进行了研究,提出培育冬前壮苗是有效缓解和防御春季低温冻害的关键措施。 . 为有效抵御春季低温对小麦冻害的不利影响,提高小麦的抗冻能力,在山西临汾采用大田随机区组 方法研究了春季低温对不同播种期小麦叶片、节间、穗部冻害的影响。结果表明:单株叶片冻害率随着播期的推迟而提高,变幅为3.0%~55.4%,单株分蘖 叶片冻害率高于单株主茎叶片冻害率,二者相差16.6%,单株鲜重、单株绿叶与单株冻叶率相关性高于其他生物学性状;节间冻害呈现随着播期的推迟而趋重, 表现出第1节间轻于第2节间;冻穗率在试验播期范围内则呈现两端轻中间重的现象,以播期10月22日最轻,10月12日最重,变幅为11.58%;同时对 形成处理间冻害差异的原因进行了研究,提出培育冬前壮苗是有效缓解和防御春季低温冻害的关键措施。 |
| [24] | . Occasionally, wheat can be exposed to overnight freezing temperatures whilst it is in the early reproductive stage and considerable frost damage can occur. In the vegetative stage, full expression of frost tolerance genes can be gained when plants are exposed to acclimation temperatures (4 C). Paradoxically, wheat during flowering has limited or no ability to effectively acclimate or re-acclimate and it demonstrates frost sensitivity during this stage. Using a combined approach involving infrared thermography and molecular analysis, the causes and effects of frost-induced damage to wheat during ear emergence were investigated. It was demonstrated that frost damage only occurs to wheat ears if the temperature falls below 5 C but some ears, and some spikelets on frozen ears do not freeze but supercool and escape freezing. In whole plant freezing experiments, spikelets showed both complete and partial sterility upon maturation and this was linked to parallel infrared observation of detached ears showing full or partial freezing of spikelets. It was hypothesised that wherever spikelet freezing occurred the spikelet was sterile and grain production per ear was drastically reduced, and in contrary, spikelet survival was as a result of supercooling. Molecular analysis of the upregulation of Cbf14 showed that as ear emergence progressed the ability to induce Cbf14 decreased but at early stages of ear emergence Cbf14 was able to be induced by exposure to acclimating temperatures (4 C). The hypothesis that the lack of the ear to acclimate is because of the incapability to induce the CBF transcription factor was only partially upheld in this investigation. Further work is needed to elucidate more precisely the molecular incompetency for the acclimation of wheat during ear emergence. |
| [25] | . Wheat is occasionally exposed to freezing temperatures during ear emergence and can suffer severe frost damage. Few studies have attempted to understand the characteristics of freezing and frost damage to wheat during late development stages.It was clearly shown that wheat appears to have an inherent frost resistance to temperatures down to 61502°C but is extensively damaged below this temperature. Acclimation, whilst increasing the frost resistance of winter wheat in a vegetative state was incapable of increasing frost resistance of plants at ear emergence. It is proposed that the ability to upregulate frost resistance is lost once vernalisation requirement is fulfilled.Culms and ears of wheat were able to escape frost damage at temperatures below 61502°C by supercooling even to as low as 611502°C and evidence collected by infrared thermography suggested that individual culms on a plant froze as independent units during freezing with little or no cross ice-nucleation strategies to protect wheat from frost damage in the field appear to revolve around avoiding ice nucleation. |
| [26] | . |
| [27] | . <p class="MsoNormal"><span >株高是影响小麦产量和控制倒伏的重要因素,研究小麦株高相关性状的遗传机制对高产育种具有指导意义。</span><span >以</span><span lang="EN-US">205</span><span >份</span><span >中国冬麦区小麦</span><span >品种</span><span lang="EN-US">(</span><span >系</span><span lang="EN-US">)</span><span >为材料,利用分布于小麦全基因组的</span><span lang="EN-US">24 355</span><span >个单核苷酸多态性</span><span lang="EN-US">(SNP)</span><span >标记对株高相关性状进行关联分析。共发现</span><span lang="EN-US">38</span><span >个</span><span lang="EN-US">SNP</span><span >与株高相关性状显著关联</span><span lang="EN-US">(<em >P</em> < 0.0001)</span><span >的,</span><span >分布在</span><span lang="EN-US">1B</span><span >、</span><span lang="EN-US">2A</span><span >、</span><span lang="EN-US">2B</span><span >、</span><span lang="EN-US">3A</span><span >、</span><span lang="EN-US">3B</span><span >、</span><span lang="EN-US">3D</span><span >、</span><span lang="EN-US">4A</span><span >、</span><span lang="EN-US">4B</span><span >、</span><span lang="EN-US">5A</span><span >和</span><span lang="EN-US">6D</span><span >染色体上。其中,</span><span lang="EN-US">11</span><span >个位点在至少</span><span lang="EN-US">2</span><span >个环境中稳定表达,<span >可用于开发</span></span><span lang="EN-US">CAPS</span><span >标记。同时,发掘了一批株高性状相关基因的优异等位变异,如降低株高的等位变异</span><span lang="EN-US">BobWhite_c48009_52</span><span >,平均降低株高</span><span lang="EN-US">12.9 cm</span><span >;降低</span><span >穗下节间长的等位变异</span><span lang="EN-US">BS00039422_51-C</span><span >和</span><span lang="EN-US">IAAV1698-A</span><span >,分别降低穗下节间长</span><span lang="EN-US">5.9 cm</span><span >和</span><span lang="EN-US">6.6 cm</span><span >。<span >本研究发掘的控制小麦株高基因位点,为</span></span><span >在分子水平上研究小麦株高复杂性状提供了有价值的参考</span><span >。</span><span lang="EN-US"><o:p></o:p></span></p> . <p class="MsoNormal"><span >株高是影响小麦产量和控制倒伏的重要因素,研究小麦株高相关性状的遗传机制对高产育种具有指导意义。</span><span >以</span><span lang="EN-US">205</span><span >份</span><span >中国冬麦区小麦</span><span >品种</span><span lang="EN-US">(</span><span >系</span><span lang="EN-US">)</span><span >为材料,利用分布于小麦全基因组的</span><span lang="EN-US">24 355</span><span >个单核苷酸多态性</span><span lang="EN-US">(SNP)</span><span >标记对株高相关性状进行关联分析。共发现</span><span lang="EN-US">38</span><span >个</span><span lang="EN-US">SNP</span><span >与株高相关性状显著关联</span><span lang="EN-US">(<em >P</em> < 0.0001)</span><span >的,</span><span >分布在</span><span lang="EN-US">1B</span><span >、</span><span lang="EN-US">2A</span><span >、</span><span lang="EN-US">2B</span><span >、</span><span lang="EN-US">3A</span><span >、</span><span lang="EN-US">3B</span><span >、</span><span lang="EN-US">3D</span><span >、</span><span lang="EN-US">4A</span><span >、</span><span lang="EN-US">4B</span><span >、</span><span lang="EN-US">5A</span><span >和</span><span lang="EN-US">6D</span><span >染色体上。其中,</span><span lang="EN-US">11</span><span >个位点在至少</span><span lang="EN-US">2</span><span >个环境中稳定表达,<span >可用于开发</span></span><span lang="EN-US">CAPS</span><span >标记。同时,发掘了一批株高性状相关基因的优异等位变异,如降低株高的等位变异</span><span lang="EN-US">BobWhite_c48009_52</span><span >,平均降低株高</span><span lang="EN-US">12.9 cm</span><span >;降低</span><span >穗下节间长的等位变异</span><span lang="EN-US">BS00039422_51-C</span><span >和</span><span lang="EN-US">IAAV1698-A</span><span >,分别降低穗下节间长</span><span lang="EN-US">5.9 cm</span><span >和</span><span lang="EN-US">6.6 cm</span><span >。<span >本研究发掘的控制小麦株高基因位点,为</span></span><span >在分子水平上研究小麦株高复杂性状提供了有价值的参考</span><span >。</span><span lang="EN-US"><o:p></o:p></span></p> |
| [28] | . 为了给小麦栽培和品质育种提供参考,以15个小麦品种为材料,分析了不同基因型小麦品种株高及各节间长度、株高构成指数与产量及产量构成因素、蛋白质、湿面筋、淀粉含量的关系。结果表明,株高及各节间长度除穗下节间外与产量均呈负相关,除穗下节间外与蛋白质、湿面筋含量均呈正相关;穗下节间长度/节间总长、(穗下节间长+穗长)/株高与产量呈正相关,与穗粒数、单穗重呈极显著正相关,与蛋白质、湿面筋含量呈负相关;千粒重与株高、各节间长、穗下节间长度/节间总长、(穗下节间长度+穗长)/株高均呈负相关;株高构成指数与结实粒数、粒重、产量均呈正相关,与蛋白质、湿面筋含量呈二次曲线关系,节间构成指数与产量、蛋白质、湿面筋含量呈负相关,株高及节间构成指数与穗粒数、单穗重均呈正相关。穗下节间长度/节间总长、(穗下节间长+穗长)/株高、株高构成指数可以作为产量和品质协调的育种选择指标,强筋、中筋小麦应选择株高适中、抗倒性好、穗下节间不宜过长、株高构成指数IL<0.54的植株;弱筋小麦应选择株高偏矮、穗下节间较长、株高构成指数IL为0.56~0.58的植株。 . 为了给小麦栽培和品质育种提供参考,以15个小麦品种为材料,分析了不同基因型小麦品种株高及各节间长度、株高构成指数与产量及产量构成因素、蛋白质、湿面筋、淀粉含量的关系。结果表明,株高及各节间长度除穗下节间外与产量均呈负相关,除穗下节间外与蛋白质、湿面筋含量均呈正相关;穗下节间长度/节间总长、(穗下节间长+穗长)/株高与产量呈正相关,与穗粒数、单穗重呈极显著正相关,与蛋白质、湿面筋含量呈负相关;千粒重与株高、各节间长、穗下节间长度/节间总长、(穗下节间长度+穗长)/株高均呈负相关;株高构成指数与结实粒数、粒重、产量均呈正相关,与蛋白质、湿面筋含量呈二次曲线关系,节间构成指数与产量、蛋白质、湿面筋含量呈负相关,株高及节间构成指数与穗粒数、单穗重均呈正相关。穗下节间长度/节间总长、(穗下节间长+穗长)/株高、株高构成指数可以作为产量和品质协调的育种选择指标,强筋、中筋小麦应选择株高适中、抗倒性好、穗下节间不宜过长、株高构成指数IL<0.54的植株;弱筋小麦应选择株高偏矮、穗下节间较长、株高构成指数IL为0.56~0.58的植株。 |
| [29] | . Main conclusion An extremely high resolution infrared camera demonstrated various freezing events in wheat under natural conditions. Many of those events shed light on years of misunderstanding... |
| [30] | . Climate change has brought more low temperature events and posed an increasing risk to the global wheat production. In order to evaluate the effects of low temperature at jointing and booting stages on wheat grain yield and its components, two years of environment-controlled phytotron experiments were carried out with two wheat cultivars under different low temperature levels and durations. Low temperature level and its interaction with low temperature duration had negative effects on the observed grain yield in two cultivars. Moreover, wheat yield was more sensitive to low temperature at booting than at jointing stages. Compared with the control treatment (T min /T max /T mean of 602°C/1602°C/1102°C, T1), 4.6%–56.4% and 3.1%–44.6% decreases of grain yield per plant (YPP) were observed under low temperature at jointing in Yangmai16 (spring wheat) and Xumai30 (semi-winter wheat), and 13.9%–85.2% and 3.2%–85.9% decreases under low temperature at booting in Yangmai16 and Xumai30, respectively. The spike number per plant (SNPP) and grain number per spike (GNPS) were more sensitive to low temperature at jointing and booting stages than 1000-grain weight (TGW). Furthermore, significant negative linear relationships were observed between the accumulated cold degree days (ACDD) and YPP, SNPP, GNPS and TGW in both cultivars. The contribution of GNPS to the variation of YPP was greater than SNPP and TGW at the mild low temperature level (T min /T max /T mean of 61202°C/802°C/302°C, T3) in both cultivars. However, at the extreme low temperature level (T min /T max /T mean of 61602°C/402°C/61102°C, T5), the major variation of YPP was caused by SNPP of Yangmai16 and GNPS of Xumai30. In general, the decreased YPP under low temperature condition was mainly from the decreased grain number per plant (GNPP02=02SNPP02×02GNPS) in both cultivars and treatment stages, thus maintaining a high GNPP is very important for compensating the yield losses caused by low temperature at jointing and booting stages. |
| [31] | . Wheat plants which had suffered frost injury to stems in the field were studied to determine the effects on grain production. Damage was also induced artificially by freezing sections of stems of plants growing outdoors and in the glasshouse. Stem injury caused lodging or death of some tillers, and reduced the number of grains set in others. However, on many occasions, even in the field, there appeared to be no association between damage to stems and grain production. In no case was the size or composition of the grains significantly affected. |
| [32] | . . |
| [33] | . . |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}