0 引言
【研究意义】作物产量性状与植株器官大小存在密切关系[1],植物器官大小除受光照、温度、营养和激素等外界环境条件影响外,主要受基因表达和信号转导等遗传因子调控。谷子具有耐旱耐瘠性强、基因组小、二倍体、生育期短等特点,以谷子为模式作物开展功能基因研究,能为解决多年来难以解析的抗旱和C4光合作用遗传分子机理提供崭新的机遇[2-3]。因此,发掘控制谷子器官大小的重要功能基因并用于遗传改良,对于加快谷子育种进程和提高谷子单产具有重要作用。【前人研究进展】研究认为,细胞总数目的增加[4]和单细胞体积增大[5]2个连续并相关的过程控制植物器官的大小,而植物器官大小在不同物种中差异显著但在物种内个体之间却相对一致,说明器官发育过程中,细胞分裂受到遗传物质的严格控制[6]。许多控制植物器官大小的相关基因已被研究,如ANT、ARF2、ARGOS、AtTOR、AtEBP1、BB和KLUH等[7],其中,拟南芥ARGOS基因家族包括ARGOS、ARL、ORS1和ORS2等4个基因。2003年,在拟南芥中首次发现了ARGOS,该基因受生长素上调表达并通过“auxin→AXR1→ARGOS→ANT”信号通路实现细胞增殖和器官生长的调节[9];ARL由油菜素内酯诱导,这种诱导依赖于油菜素内酯受体BRI1[10];此外,OSR1[11]和OSR2[12]通过乙烯诱导上调表达,但受ABA和油菜素内酯抑制。该基因家族的共同特点是含有一个保守的ORS结构域,包含2个跨膜螺旋结构,在不同植物激素诱导表达下,最终通过调控细胞数目或细胞体积来调节植物器官大小,对植株生长起到促进作用[8]。玉米ARGOS基因家族目前发现8个基因,其中ZmARGOS1受乙烯下调表达并参与器官大小的调控,拟南芥转基因植株抗旱性提高;ZmARGOS8超表达可使转基因玉米产量大幅提高[13]。水稻OsARGOS也受生长素诱导上调表达,转基因植株侧生器官的花和叶片变大,角果数目和种子数量增多[14];TaARGOSs受干旱、盐以及外源ABA、MeJA、NAA、ACC、GA3、BR等多种激素上调表达,但不同小麦基因组TaARGOS-A、TaARGOS-B和TaARGOS-D的表达模式存在差异,转TaARGOS-D拟南芥植株发芽率增加,叶和花环直径增大、单株角果数目增多[15];另外,ARGOS基因在大白菜[16-18]、萝卜[19]、紫花苜蓿[20]中也有报道,均参与植物器官大小的调节。【本研究切入点】谷子ARGOS基因家族成员的特性和受植物激素诱导后的表达模式以及其与产量性状的关系尚无报道。【拟解决的关键问题】本研究通过生物信息学方法获得谷子中所有ARGOS家族成员并分析其蛋白序列,采用同源克隆方法获得该基因家族的一个成员SiARGOS1的编码区及其启动子序列,用生物信息学方法分析启动子的顺式作用元件,采用实时荧光定量PCR分析SiARGOS1在谷子各组织器官以及不同植物激素诱导后的表达模式,通过基因编码区及其启动子序列进行SNP分析及标记开发,同时利用85份谷子品种的产量性状进行验证,开发用于检测该基因与谷子产量性状相关优异等位变异的功能标记。1 材料与方法
1.1 植物材料与性状测定
用于SiARGOS1克隆的谷子品种:晋29A和K186,种植方式:每材料种植1行,行长2.5 m,行距33 cm,取样时期:拔节期幼叶;用于基因器官表达的谷子品种:豫谷1号,种植方式同上,取样时期:抽穗后第7天,分别取其根、茎、叶及穗;用于植物激素IAA和ETH响应表达的谷子品种:豫谷1号,种植方式:将种子种植于混合等量营养土和适量水的钵中,放在25℃左右的光照培养箱中培养10 d。然后,用5 μmol·L-1 IAA和0.05% ETH分别处理幼苗,在处理后1、3、6和12 h分别取其叶片;用于SiARGOS1的SNP分析和标记开发的谷子品种共19份(电子附表1,用下划线标注的品种):种植方式与取样时期同晋29A和K186;用于SiARGOS1标记验证的谷子品种:85份谷子品种(电子附表1),其中63份分别来源于中国东北、华北和西北地区,6份来源于美国,4份来源于日本,另外12份来源于印度。于2015和2016年种植于山西省农业科学院谷子研究所试验田,行长3 m,行距0.33 m,取拔节期叶片用于DNA提取,并于植株籽粒成熟时随机选取10穗,称量其穗重和穗粒重,并随机取1 000粒种子称重,测定千粒重,利用SPSS软件IBM SPSS Statistics 19进行统计分析。1.2 SiARGOS家族成员的生物信息学分析
在NCBI网站(https://www.ncbi.nlm.nih.gov/ genome/?term=foxtail+millet)对已有植物ARGOS蛋白保守结构域进行BLAST,获得谷子中所有ARGOS家族成员,采用DNAMAN软件,结合拟南芥、水稻、玉米、小麦、短柄草和高粱的ARGOS家族成员进行蛋白比对,并采用MEGA5生成无根进化树,用邻接法进行同源进化分析。1.3 SiARGOS1全长cDNA克隆
通过拟南芥ARGOS基因家族中4个基因的mRNA序列,在phtozome(http://www.phytozome.net/ search.php)网站进行BLAST,获得一种谷子ARGOS基因家族序列,编号Seita.8G077300,根据此序列,采用软件Primer Premier5.0设计包含该基因ORF的引物SiARGOS1(SiARGOS1-1F:5′-ACAAATCCCCAC CCTTGTCA-3′,SiARGOS1-1R:5′-ACTCCTGAAAA GATGCTTCACA-3′),以晋29A和K186的cDNA为模板,扩增得到PCR产物,经克隆测序后得到目标序列。PCR扩增体系为模板2 μL、2×GC Buffer 10 μL、10 mmol·L-1 dNTPs 0.4 μL、2 μmol·L-1特异引物4 μL、rTaq DNA聚合酶0.2 μL,ddH2O补充至20 μL。PCR扩增程序为95℃ 3 min;95℃ 30 s,58℃ 30 s,72℃ 45 s,35个循环;72℃ 10 min,4℃保存。取PCR产物在1%琼脂糖凝胶上电泳检测。1.4 SiARGOS1启动子克隆
根据phtozome(http://www.phytozome.net/search. php)网站序号Seita.8G077300,将该基因向上游延伸约2 kb,用软件Primer Premier5.0设计启动子区PCR引物Argos-Pro,(Argos-Pro-F:5′-CTCTGTCGTCTG CAAGCAA-3′和Argos-Pro-R:5′-ACTGACAAGGG TGGGGATTT-3′),以晋29A和K186基因组DNA为模板,扩增体系和条件(除退火温度和延伸时间改为60℃ 30 s,72℃ 2 min)同上,经克隆测序获得该基因启动子区序列。将获得的启动子序列提交到PlantCARE(http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/),进行顺式作用元件的预测。1.5 RT-PCR和qRT-PCR分析
取豫谷1号抽穗后第7天的根、茎、叶和幼穗,分别提取总RNA并反转录为cDNA,另外,用5 μmol·L-1 IAA和0.05% ETH分别处理豫谷1号生长10 d的幼苗,在处理后1、3、6和12 h分别取样,提取总RNA并反转录为cDNA,半定量PCR和实时荧光定量PCR引物为SiARGOS1-RT-F:5′-TTGCGTCGA CTTACTTCAGC-3′,SiARGOS1-RT-R:5′-CATGCTC CTCACATCGGTTG-3′。以谷子ACTIN作为对照(SiActin-F:5′-TTCCCTGGTATTGCTGACCG-3′,SiActin-R:5′-CTCACCCTTCGAGATCCACA-3′)。半定量PCR反应程序为94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 30 s,28个循环;72℃ 5 min。反应产物经琼脂糖凝胶电泳分离;采用Bio-Rad C1000 cycler real time PCR system进行实时荧光定量PCR分析,反应体系为1/10 cDNA template(反转录第一链)1 μL、SYBR Premix Ex Taq (DRR041D)5 μL、2 μmol·L-1特异引物1 μL,补充ddH2O至10 μL。扩增程序为94℃ 5 min;94℃ 30 s,58℃ 30 s,72℃ 30 s,35个循环;72℃ 5 min。65℃—98℃绘制溶解曲线。采用比较阈值法进行定量分析,手工设定荧光阈值,确定循环数Ct值,根据Ct值,计算各样本的C值,C=2-ΔCt,ΔCt=Ct目标基因–Ct内标基因。试验设置3次生物学重复并采用双尾等方差t检验的方法进行显著性检验(P<0.05)。1.6 SNP和单倍型分析及功能标记开发
对19份谷子材料进行基因编码区及启动子全长测序,用引物SiARGOS1和Argos-Pro分别进行SiARGOS1编码区和启动子扩增,对PCR产物进行克隆测序,利用DNAstar软件系统的Sequman、Editseq和MegAlign软件包进行DNA序列的分析,包括拼接、整理与SNP和单倍型分析。引物SiARGOS1的扩增产物经限制性内切酶AccⅡ(CGˇCG)酶切后的片段差异,用于检测151 bp位点的碱基差异;根据启动子区两处插入和缺失设计SSR引物AP-1和AP-2,用于启动子区的单倍型分型,其中AP-1-F:5′-AGATGAC TCTAAAGGGCATCG-3′,AP-1-R:5′-CAAGGGCAG CAGTGTTTTC-3′;AP-2-F:5′-GTCTTTTGGGATTGT GTCATC-3′,AP-2-R:5′-TGACTAATGTGGTACGGG TC-3′,同时,开发的3处标记分别用85份谷子品种进行验证。2 结果
2.1 SiARGOS家族成员的生物信息学分析
在NCBI网站(https://www.ncbi.nlm.nih.gov/ genome/?term=foxtail+millet)对已有ARGOS蛋白保守结构域进行BLAST,共找到6个谷子ARGOS成员,将其分别命名为SiARGOS1、SiARGOS2、SiARGOS3、SiARGOS4、SiARGOS5和SiARGOS6。将得到的谷子ARGOS蛋白与拟南芥、玉米、水稻、小麦、高粱和短柄草的ARGOS蛋白进行了蛋白比对(图1-A)和进化树分析(图1-B),结果表明,与其他已知ARGOS一样,谷子ARGOS家族具有保守的OSR结构域,包含2个跨膜螺旋结构(图1-A)。SiARGOS1、SiARGOS3、SiARGOS5和SiARGOS6与拟南芥AtARGOS以及AtARL聚为一类,其中,SiARGOS1和SiARGOS3与小麦、短柄草和高粱的ARGOS同源性最高,SiARGOS5和SiARGOS6与玉米ZmARGOS1和水稻OsARGOS同源性最高,SiARGOS2和SiARGOS4与拟南芥AtOSR1聚为一类,与玉米ZmARGOS8同源性最高。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1谷子SiARGOSs和其他ARGOS蛋白的序列比对及进化树分析(A:谷子SiARGOSs和其他植物ARGOS家族的OSR保守结构域,加框的部分为2个跨膜螺旋,中间部分为一个由8个氨基酸(LPPLPPPP)组成的高度保守的富含亮氨酸区域;Si:谷子;At:拟南芥;Ta:小麦;Zm:玉米;Os:水稻;Bd:短柄草;Sb:高粱。B:ARGOS的系统进化树)
-->Fig. 1Sequence alignment and phylogenetic analysis of the SiARGOSs and other ARGOS proteins(A: The conserved OSR domain of SiARGOSs and ARGOS homologs in other plants. Two putative transmembrane helices are boxed, and a highly conserved proline-rich motif consisting of 8 amino acid residues (LPPLPPPP) is in the middle; Si: Setaria italica; At: Arabidopsis thaliana; Ta: Triticum aestivum; Zm: Zea mays; Os: Oryza sativa; Bd: Brachypodium distachyon; Sb: Sorghun bicolar. B: Phylogenetic tree of ARGOS)
-->
2.2 SiARGOS1编码区的克隆
获取拟南芥ARGOS基因家族中4个基因的mRNA序列,在phtozome(http://www.phytozome.net/ search.php)网站进行BLAST,获得该基因家族的一种谷子序列,编号Seita.8G077300,经比对分析为SiARGOS1,而该基因蛋白与拟南芥ARGOS基因家族中的AtARGOS归为一类,与已有小麦TaARGOSs、高粱SbARGOS和短柄草BdARGOS同源性最高。以晋29A和K186的cDNA为模板,用引物SiARGOS1进行扩增,获得两条特异性条带,长度为491 bp(图2-A),经克隆测序后发现ARGOS的ORF为342 bp,基因编码113个氨基酸,位于谷子第8染色体上,核苷酸序列比对结果显示该基因编码区序列在晋29A和K186中没有差异,同时,用相同引物在基因组DNA中的扩增产物也为491 bp,序列一致,说明该基因没有内含子,这与拟南芥,水稻和玉米中报道的结果一致[9,13-14]。2.3 SiARGOS1启动子的克隆和分析
为进一步了解SiARGOS1的表达调控机制,克隆得到SiARGOS1起始密码子前2 109 bp启动子序列(图2-B),采用PlantCARE对SiARGOS1起始密码子(ATG)上游的2 109 bp片段进行分析,发现该序列含有真核生物启动子的典型元件CAAT-box(19个)和TATA-box(6个),能够与起始转录的转录因子结合;该区域存在AE-box、Box II、G-box、GTGGC-motif和Sp1等5个与光反应有关的元件;AP-2-like、CGTCA-motif(3个)、GARE-motif和TGA-element是与乙烯、茉莉酸、赤霉素和生长素相关的转录因子的结合位点;GCN4-motif和Skn-1-motif(4个)是与胚乳表达相关的顺式作用元件;另外还存在其他转录因子结合的位点(见电子附表2)。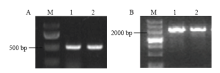 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2SiARGOS1及其启动子的扩增(A:SiARGOS1的扩增,M:100 bp DNA marker;1:晋29A;2:K186;B:SiARGOS1启动子的扩增,M:200 bp DNA marker;1:晋29A;2:K186)
-->Fig. 2Amplification profile of SiARGOS1 gene and promoter(A: Amplification of SiARGOS1 gene, M: 100 bp DNA marker; 1: Jin 29A; 2: K186; B: Amplification of SiARGOS1 promoter, M: 200 bp DNA marker; 1: Jin 29A; 2: K186)
-->
2.4 不同组织器官和不同激素条件下SiARGOS1的表达
在豫谷1号抽穗后7 d分别取其根、茎、叶和穗,提取总RNA、合成的cNDA经均一化、RT-PCR和实时定量PCR后,检测SiARGOS1在谷子抽穗期不同组织器官中的表达情况。结果表明,SiARGOS1的RT-PCR(图3-A)和实时定量PCR(图3-B)结果基本一致,该基因在谷子抽穗期的根、茎、叶和穗中均有表达,在根中的表达量最大,茎和叶次之,而穗中的表达量最小(图3-A)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3SiARGOS1在不同组织中的表达分析(A:通过半定量PCR检测SiARGOS1在不同组织中的表达;B:通过实时定量PCR检测SiARGOS1在不同组织中的表达)
-->Fig. 3Expression of SiARGOS1 in different tissues(A: Expression of SiARGOS1 in different tissues using semi-quantitative PCR; B: Expression of SiARGOS1 in different tissues using real-time PCR)
-->
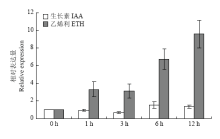
通过荧光定量PCR检测SiARGOS1对不同激素响应表达。结果显示,SiARGOS1对IAA不敏感,但受ETH上调表达,在ETH处理1和3 h后表达有所上升,且上升水平基本没有变化,随着处理时间的推移,表达量进一步升高,在12 h表达量最高(图4),说明该基因对生长素响应不敏感但受乙烯上调表达。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4SiARGOS1对IAA和ETH的响应表达
-->Fig. 4Expression of SiARGOS1 in response to IAA and ETH
-->
2.5 SiARGOS1的序列分析及标记开发
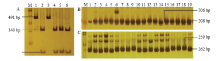
通过对19份谷子材料进行SiARGOS1编码区及其启动子测序,发现有2份材料在该基因编码区151 bp(起始密码子83 bp)处存在一个SNP(C/G),导致该基因第28个氨基酸发生突变(Ala/Gly),该碱基差异导致酶切位点AccⅡ(CGˇCG)发生变化(CGCG/CGGG,后者序列AccⅡ不能识别),据此设计了一个CAPS标记(图5)。用引物SiARGOS1对材料进行扩增,扩增长度为491 bp,扩增产物经AccⅡ酶切后,获得2种基因型,一种为(CGCG)基因型,将其命名为Hap-Ag-C,酶切不能被切开,酶切产物片段大小仍为491 bp,另一种为(CGGG)基因型,将其命名为Hap-Ag-G,经酶切产生340和151 bp片段(图6-A)。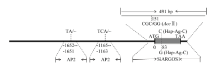 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5SiARGOS1的标记开发
-->Fig. 5Markers development of SiARGOS1
-->
序列分析表明SiARGOS1启动子区共存在19处SNP和2处插入和缺失(见电子附表3),根据-1 652—-1 651处(TA)2/3和-1 165—-1 163处(TCA)1/2的序列差异分别设计SSR引物AP-1和AP-2(图5),用于启动子区的分型检测。AP-1(图6-B)在不同品种的扩增条带分别为206和208 bp,AP-2(图6-C)在不同品种的扩增条带分别为259和262 bp。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6
-->Fig. 6Development of CAPS and SSR markers of SiARGOS1 gene and its promoter(A: CAPS marker of AccⅡabout SiARGOS1 SNP(C/G); M: DNA marker; 1-6: Mengxuan 5084, Changnong 35 hao, Shawanguzi, Shangdong-5, Jin29A, K186; B and C were SSR markers of AP-1 and AP-2 respectively, M: DNA marker; 1-19: Aininghuang, Shawanguzi, Miyou, Hebeishilixiang, Zhang 8311-13, ISE-4, Lengsheng-1, Pinzi 15, Qianchuanzi, Changnong 35 hao, Shangdong-5, Mengxuan 5084, PI614817, Heizhangu, Chadiangu, Tie 8503, Chao 108, Anchunwei)
-->
2.6 功能标记验证
用标记CPAs-AccⅡ、AP-1和AP-2对85份品种穗重(panicle weight,PW)、穗粒重(grain weight,GW)和千粒重(thousand-grain weight,TGW)等产量性状进行检测(表1)。用限制性酶AccⅡ开发的CPAS标记检测SiARGOS1编码区151 bp(起始密码子83 bp)SNP(C/G)位点,Hap-Ag-G基因型有9 份,75份为Hap-Ag-C基因型,2种基因型间穗重、穗粒重和千粒重的差异均不显著;用SSR标记AP-1检测SiARGOS1启动子区-1 652—-1 651处(TA)2/32 bp的缺失,~基因型有42份,43份为TA基因型,2种基因型间穗重、穗粒重和千粒重的差异均不显著;用SSR标记AP-2检测SiARGOS1启动子区-1 165—-1 163处(TCA)1/23 bp的缺失,~基因型有58份,27份为TCA基因型,2种基因型间千粒重的差异均不显著,但其穗重和穗粒重在2015和2016年两年间的差异均达到了显著水平。说明根据该处开发的SSR标记AP-2可作为功能标记用于SiARGOS1优异等位变异的检测,而位于该基因启动子区-1 165—-1 163处(TCA)1/23 bp缺失的等位变异可能对谷子产量性状有着重要的影响。Table 1
表1
表185份谷子品种中不同分子标记检测基因型与产量性状相关性
Table 1Association between allelic variants of SiARGOS1 and yield traits in 85 cultivars tested with different markers
| 年份 Year | 性状 Traits | CPAS-AccⅡ | AP-1 | AP-2 | |||
|---|---|---|---|---|---|---|---|
| Hap-Ag-G | Hap-Ag-C | ~ | TA | ~ | TCA | ||
| 2015 | 穗重 Panicle weight (g) | 16.23±3.01 | 17.28±1.25 | 17.28±1.58 | 17.01±1.67 | 15.51±1.39* | 20.61±1.88* |
| 穗粒重 Grain weight (g) | 12.34±2.44 | 13.89±1.06 | 14.15±1.36 | 13.33±1.40 | 12.31±1.20* | 16.74±1.54* | |
| 千粒重 Thousand-grain weight (g) | 2.81±0.26 | 2.63±0.08 | 2.71±0.54 | 2.60±0.84 | 2.59±0.10 | 2.78±0.10 | |
| 2016 | 穗重 Panicle weight (g) | 15.98±2.81 | 17.72±1.22 | 17.63±1.52 | 17.40±1.64 | 15.91±1.35* | 20.85±1.83* |
| 穗粒重 Grain weight (g) | 13.49±2.46 | 15.18±1.07 | 15.21±1.38 | 14.80±1.40 | 13.65±1.20* | 17.82±1.55* | |
| 千粒重 Thousand-grain weight (g) | 2.88±0.26 | 2.74±0.07 | 2.79±0.08 | 2.73±0.11 | 2.71±0.09 | 2.86±0.10 | |
新窗口打开
3 讨论
本研究通过对已有ARGOS蛋白保守结构域进行BLAST,共获得6个谷子ARGOS成员,均具有ARGOS家族典型的保守OSR结构域,包含2个跨膜螺旋结构。用拟南芥ARGOS基因家族中4个基因的mRNA序列在phtozome进行BLAST,获得谷子ARGOS基因家族的一种序列,经比对分析为SiARGOS1,该基因蛋白与拟南芥ARGOS基因家族中的AtARGOS归为一类,而AtARGOS的研究较该基因家族中另外3个成员最早也更为广泛和深入,与其归为一类的水稻OsARGOS[14]、玉米ZmARGOS1 [13]和小麦TaARGOS-D[15]等均已报道,它们均参与信号转导并对器官生长有促进作用[11],故对SiARGOS1进行了深入的研究。结果显示,该基因位于谷子第8染色体上,在基因组中仅有一个拷贝,并且编码区内不含有内含子。拟南芥[9]、玉米[13]、水稻[14]、小麦[15]和大白菜[16]ARGOS基因也仅有一个拷贝并且不含有内含子,说明该类基因在植物中保守性。SiARGOS1在豫谷1号抽穗后7 d的根、茎、叶和穗中均有表达,在根中的表达量最大,茎和叶次之,而穗中的表达量最小,这与谷子功能基因网站中(http://structuralbiology.cau.edu.cn/SIFGD/gene_detail. php?gene=Si027037m)查询的该基因组织表达特征基本一致,都是在根中表达量最高,穗部表达略有不同,这可能与不同生育期取样有关;另外,该基因在不同植物组织器官的表达也不尽相同,拟南芥AtARGOS在根、茎、花、幼嫩的花环叶和果荚呈现低水平表达,但在成熟的叶中基本检测不到[9];水稻OsARGOS在幼嫩的根、叶和授粉5 d的种子中表达量较高,而在茎、成熟叶和小穗中表达量较低[14]。小麦TaARGOSs主要在茎部表达[15]。
通过启动子预测发现SiARGOS1的启动子除了核心启动元件CAAT-box和TATA-box以外,还包括与乙烯、茉莉酸、赤霉素和生长素等多种植物激素相关的顺式作用元件调控元件AP-2-like[21]、CGTCA-motif[22]、GARE-motif[23]和TGA-element[24]。在拟南芥ARGOS基因家族中,ARGOS基因受生长素上调表达[9];ARL由油菜素内酯诱导,这种诱导依赖于油菜素内酯受体BRI1[10];OSR1和OSR2通过乙烯诱导上调表达[11-12]。另外,ARGOS在水稻[14]中受生长素上调表达,在玉米中却受乙烯下调表达[25-26],而在小麦中又受生长素、乙烯、茉莉酸和赤霉素等多种植物激素诱导表达[15],因此,分别选用生长素和乙烯利处理生长10 d谷子幼苗,结果显示,SiARGOS1对生长素不敏感,但受乙烯利上调表达。表明该基因家族中的不同基因受不同植物激素诱导表达,而且在不同物种中同一基因对不同激素的诱导表达也不同,或者在不同的物种中对植物激素存在一定的特异性,但最终都影响了植物器官大小的调控。另外,启动子区还存在与光反应和胚乳表达相关的元件,但目前还未见ARGOS与之相关的研究报道,有待于进一步的试验验证。
功能标记(functional marker,FM)是根据基因内部序列设计的,是一种区分和预测等位基因及相对性状的分子标记[27]。水稻qSH1的5′调控区的一个单核苷酸多态性T/G与其落粒性高度相关[28],玉米控制株高和开花期的tb1[29]和Dwarf8[30]、小麦中关于粒宽和光周期的TaGW2[31]和Ppd-D1[32]、控制番茄果实大小的fw2.2[33]等基因都成功用于功能标记的研究。目前,谷子中定位了多个与千粒重、穗长等产量相关性状QTL[3, 34],但对于产量性状相关的特定基因克隆和功能标记开发目前尚未报道。根据SiARGOS1及其启动子区的位点差异开发了1个CAPS标记和2个SSR标记,都具有位点特异性、共显性、操作简便等特点。其中,CPAS-AccⅡ标记是根据基因起始密码子83 bp处存在一个SNP(C/G)设计的,该处变异导致该基因第28个氨基酸发生突变(Ala/Gly),但该位点不在SiARGOS1的保守ORS结构域内,2种基因型间穗重、穗粒重和千粒重的差异均不显著;而根据启动子-1 165—-1 163处(TCA)1/2的序列差异设计的SSR标记AP-2在2种基因型(~/TCA)间穗重和穗粒重差异均达到显著水平,说明AP-2可作为功能标记,用于该基因与谷子产量性状相关优异等位变异的鉴定。这也进一步证实谷子ARGOS参与器官大小调控,最终影响谷子产量。
另外,用AP-2检测SiARGOS1基因启动子区-1 165—-1 163处(TCA)1/23 bp的缺失,我们选取的85份谷子材料,~基因型有58份,27份为TCA基因型,其中携带有TCA基因型材料的穗重和穗粒重均显著高于~基因型的材料,说明-1 165—-1 163处(TCA)1/23 bp的插入为优异等位变异。在中国的63份材料中,该变异在育成品种中出现的频率高于地方种,主要分布在河北(61.54%)、山西(44.44%)、辽宁(57.14%)和山东(44.44%)等4个省份;在来源于美国和印度的材料中均没有检测到,而在来源于日本的材料中该变异出现的频率为75%,说明SiARGOS1在谷子选育过程中受到了一定的选择,TCA基因型具有较高的谷子育种价值。
Table1
附表1
附表185份谷子材料及其来源
Table1Supplementary Table 1 The names and origins of 85 foxtail millet accessions
| 编号 Number | 名称 Name | 地理来源 Origin | 编号 Number | 名称 Name | 地理来源 Origin | |
|---|---|---|---|---|---|---|
| 1 | 白把子BAI BA ZI | 中国北京Beijing, China | 44 | 09KF6-449 | 中国山西Shanxi, China | |
| 2 | 白毛谷BAI MAO GU | 中国甘肃Gansu, China | 45 | 2009变异22 2009 BIANYI 22 | 中国山西Shanxi, China | |
| 3 | 茄头谷QIE TOU GU | 中国甘肃 Gansu, China | 46 | 2009变异27 2009 BIANYI 27 | 中国山西Shanxi, China | |
| 4 | 69-4 | 中国河北 Hebei, China | 47 | 99变异7 99 BIANYI 7 | 中国山西Shanxi, China | |
| 5 | 矮97 AI 97 | 中国河北 Hebei, China | 48 | 稗子 BAI ZI | 中国山西Shanxi, China | |
| 6 | 矮宁黄 AI NING HUANG | 中国河北 Hebei, China | 49 | 长0301 CHANG 0301 | 中国山西Shanxi, China | |
| 7 | 白根红粘BAI GEN HONG ZHAN | 中国河北 Hebei, China | 50 | 长农35号 CHANGNONG 35 HAO | 中国山西Shanxi, China | |
| 8 | 承168CHENG 168 | 中国河北 Hebei, China | 51 | 长农35变 CHANGNONG 35 BINA | 中国山西Shanxi, China | |
| 9 | 河北十里香HEBEI SHI LI XIANG | 中国河北 Hebei, China | 52 | 汾选8号 FENXUAN 8 HAO | 中国山西Shanxi, China | |
| 10 | 衡谷12 HENG GU 12 | 中国河北 Hebei, China | 53 | 晋汾6号 JINFEN 6 HAO | 中国山西Shanxi, China | |
| 11 | 衡研17 HENG YAN 17 | 中国河北 Hebei, China | 54 | 晋谷16 JINGU16 | 中国山西Shanxi, China | |
| 12 | 群育一号QUN YU YI HAO | 中国河北 Hebei, China | 55 | 冷生1 LENGSHENG 1 | 中国山西Shanxi, China | |
| 13 | 铁变16TIE BIAN 16 | 中国河北 Hebei, China | 56 | 品资15 PINZI15 | 中国山西Shanxi, China | |
| 14 | 小红谷XIAO HONG GU | 中国河北 Hebei, China | 57 | 品资39 PINZI39 | 中国山西Shanxi, China | |
| 15 | 张8311-13 ZHANG 8311-13 | 中国河北 Hebei, China | 58 | 小青谷XIAO QING GU | 中国山西Shanxi, China | |
| 16 | 刀把齐DAO BA QI | 中国黑龙江Heilongjiang, China | 59 | 小软谷XIAO RUAN GU | 中国山西Shanxi, China | |
| 17 | 大粒黄DA LI HUANG | 中国黑龙江Heilongjiang, China | 60 | 一把粘YI BA ZHAN | 中国山西Shanxi, China | |
| 18 | 大青谷DA QING GU | 中国黑龙江Heilongjiang, China | 61 | 竹叶青ZHU YE QING | 中国山西Shanxi, China | |
| 19 | 红谷子HONG GU ZI | 中国黑龙江Heilongjiang, China | 62 | 茶淀谷CHA DIAN GU | 中国天津Tianjin, China | |
| 20 | 红粘谷HONG ZHAN GU | 中国黑龙江Heilongjiang, China | 63 | 沙湾谷子SHA WAN GU ZI | 中国新疆Xinjiang, China | |
| 21 | 黄沙子二号HUANGSHA ZI ER HAO | 中国黑龙江Heilongjiang, China | 64 | PI296378 | 美国America | |
| 22 | 钱串子QIAN CHUAN ZI | 中国黑龙江Heilongjiang, China | 65 | PI614817 | 美国America | |
| 23 | 鹌鹑尾AN CHUN WEI | 中国吉林Jilin, China | 66 | PI649314 | 美国America | |
| 24 | 气死碱QI SI JIAN | 中国吉林Jilin, China | 67 | PI436631 | 美国America | |
| 25 | 朝66 CHAO 66 | 中国辽宁Liaoning, China | 68 | PI464312 | 美国America | |
| 26 | 白谷子BAI GU ZI | 中国辽宁Liaoning, China | 69 | PI363068 | 美国America | |
| 27 | 朝108 CHAO 108 | 中国辽宁Liaoning, China | 70 | 米优MI YOU | 日本Japan | |
| 28 | 黑粘谷HEI ZHAN GU | 中国辽宁Liaoning, China | 71 | 日本千斤谷RI BEN QIAN JIN GU | 日本Japan | |
| 29 | 红苗粘谷HONG MIAO ZHAN GU | 中国辽宁Liaoning, China | 72 | 沙湾谷子SHA WAN GU ZI | 日本Japan | |
| 30 | 铁8503 TIE 8503 | 中国辽宁Liaoning, China | 73 | 熊本国分2号 XIONG BEN GUO FEN 2 HAO | 日本Japan | |
| 31 | 铁7924 TIE 7924 | 中国辽宁Liaoning, China | 74 | SE-KEP-1 | 印度India | |
| 32 | 大青苗白米DA QING MIAO BAI MI | 中国内蒙古InnerMongolia, China | 75 | SE-KEP-7 | 印度India | |
| 33 | 蒙选5084 MENGXUAN 5084 | 中国内蒙古InnerMongolia, China | 76 | SE-KEP-14 | 印度India | |
| 34 | 拔谷子BA GU ZI | 中国山东Shandong, China | 77 | SE-KEP-19 | 印度India | |
| 35 | 大白糙谷DA BAI CAO GU | 中国山东Shandong, China | 78 | SE-KEP-20 | 印度India | |
| 36 | 大小穗红谷DA XIAO SUI HONG GU | 中国山东Shandong, China | 79 | SE-KEP-22 | 印度India | |
| 37 | 骡子尾LUO ZI WEI | 中国山东Shandong, China | 80 | ISE-4 | 印度India | |
| 38 | 蓬莱麦茬谷PENG LAI MAI CHA GU | 中国山东Shandong, China | 81 | ISE-8 | 印度India | |
| 39 | 山东-4 SHANDONG-4 | 中国山东Shandong, China | 82 | ISE-10 | 印度India | |
| 40 | 山东-5 SHANDONG-5 | 中国山东Shandong, China | 83 | ISE-11 | 印度India | |
| 41 | 小绳头XIAO SHENG TOU | 中国山东Shandong, China | 84 | ISE-13-1 | 印度India | |
| 42 | 猪屎谷子ZHU SHI GU ZI | 中国山东Shandong, China | 85 | ISE782 | 印度India | |
| 43 | 09K40-43-2 | 中国山西Shanxi, China |
新窗口打开
Table2
附表2
附表2SiARGOS1启动子的调控区域预测分析
Table2Supplementary Table 2 Predication analysis of the elements within the promoter of SiARGOS1
| 元件名称 Name of box | 序列 Sequence | 位置 Position (bp) | 功能 Function |
|---|---|---|---|
| AE-box | AGAAACAT | -222 | 光响应元件Part of a module for light response |
| AP-2-like | CGACCAGG | -2040 | 乙烯响应元件Ethylene-responsive transcription factor |
| ARB | TGGTTT | -426 | 厌氧诱导响应元件Essential for the anaerobic induction |
| Box II | ACACGTAGA | -1493 | 光响应元件Part of a light responsive element |
| Box-W1 | TTGACC | -842 | 真菌诱导响应元件Fungal elicitor responsive element |
| CAAT-box | gGCAAT | -119, -1480 | 启动子核心元件Common cis-acting element |
| CAATT | -68, -117, -1032, -1068, -1291, -1478 | ||
| CAAT | -467,-929,-1357,-1590 | ||
| CAAAT | -1223,-1265,-1760,-2012 | ||
| CCAAT | -1033 | ||
| CGTCA-motif | CGTCA | -368, -816, -1430,-1817 | 茉莉酸响应元件Involved in the MeJA-responsiveness |
| G-Box | CACGTA | -1492,-1690 | 光响应元件Involved in light responsiveness |
| GARE-motif | AAACAGA | -101 | 赤霉素响应元件Gibberellin-responsive element |
| GC-motif | CCCCCG | -778 | 厌氧诱导相关元件Involved in anoxic specific inducibility |
| GCN4-motif | TGTGTCA | -1365 | 胚乳表达相关元件Involved in endosperm expression |
| GTGGC-motif | GATTCTGTGGC | -271 | 光响应元件Part of a light responsive element |
| O2-site | GTTGACGTGA | -1188 | 蛋白代谢调节元件Zein metabolism regulation |
| Skn-1-motif | GTCAT | -32, -815, -1362,-1816 | 胚乳表达所需元件Required for endosperm expression |
| Sp1 | CC(G/A)CCC | -61, -779, -827, -990 | 光响应元件Light responsive element |
| TATA-box | TAATA | -169, -200, -1702 | 启动子核心元件Core promoter element |
| TTTTA | -276, -1147 | ||
| TATA | -1234,-1273, -1307 | ||
| TGA-element | AACGAC | -2085 | 生长素响应元件Auxin-responsive element |
| TGACG-motif | TGACG | -480 | 茉莉酸响应元件Involved in the MeJA-responsiveness |
新窗口打开
Table3
附表3
附表3SiARGOS1启动子的SNP及单倍型分型
Table3Supplementary Table 3 SNP and Haplotype in promoter of SiARGOS1
| 单倍型类型 Haplotype Type | Hap-AP1 | Hap-AP2 | Hap-AP3 | Hap-AP4 | |
|---|---|---|---|---|---|
| 材料数 Material number | 1 | 6 | 7 | 5 | |
| 单倍型频率 Frequency of Haplotype(%) | 5.26 | 31.58 | 36.84 | 26.32 | |
| ATG上游SNP的位点(bp) SNP of ATG upstream sites (bp) | -566 | C | T | C | C |
| -605 | A | G | G | A | |
| -1054 | T | C | C | C | |
| -1059 | G | A | G | G | |
| -1095 | T | A | A | A | |
| -1102 | G | A | A | A | |
| -1105 | T | C | C | C | |
| -1125 | G | A | A | A | |
| -1165—-1163 | TCA | ~ | ~ | TCA | |
| -1244 | G | T | T | T | |
| -1274 | A | A | A | G | |
| -1474 | T | T | C | T | |
| -1483 | G | A | G | G | |
| -1567 | T | C | T | T | |
| -1568 | T | T | C | T | |
| -1590 | T | T | T | C | |
| -1593 | G | G | A | G | |
| -1622 | G | G | A | G | |
| -1652—-1651 | TA | ~ | TA | ~ | |
| -1656 | T | C | C | C | |
| -2010 | T | T | T | A | |
新窗口打开
4 结论
获得6个谷子ARGOS成员,均具有ARGOS家族典型的保守OSR结构域,并克隆了与拟南芥AtARGOS归为一类的家族成员SiARGOS1的编码区及其启动子序列。其启动子区含有与植物激素、光周期及胚乳表达有关的元件,且在其编码区和启动子区分别开发了1个CAPS-AccⅡ标记和2个SSR标记,其中SSR标记AP-2可作为功能标记,用于该基因与谷子穗重和穗粒重等产量性状相关优异等位变异的鉴定。该基因在谷子根、茎、叶和穗等器官中均有表达,对生长素不敏感,但受乙烯利上调表达,与小麦、短柄草和高粱ARGOS蛋白同源性最高。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}