0 引言
【研究意义】干旱和盐碱是制约作物发展的重要环境因素,非生物逆境胁迫下作物生长缓慢[1]、黄化早衰[2-3]、产量降低[4],对农业生产造成严重的损害。植物自身可通过受体蛋白感知干旱、盐、温度等逆境信号[5],通过激素和信号转导基因传递,从而激活抗逆信号,实现对逆境的适应。目前,尽管大量的逆境相关基因被克隆和报道,然而可用于极端环境下作物遗传改良的报道仍然非常少。苏云金芽孢杆菌Bt基因在棉花抗棉铃虫中的成功应用[6],为利用外源微生物基因改良作物抗虫等农艺性状提供了重要的参考。然而,异源表达微生物基因提高植物抗非生物胁迫的报道仍然非常的少,是否存在类似Bt蛋白提高植物抗虫效果的优良基因有待深入研究。【前人研究进展】耐辐射异常球菌(Deinococcus radiodurans)是一类可生存于极端环境的微生物,对电离辐射、紫外线、干旱、丝裂霉素C等均具有抗性[7-8]。UDUPA等[9]从耐辐射异常球菌302菌株中分离得到了41株电离辐射敏感菌株,这些敏感菌株被划分成irrA至irrP的16个亚群,每个亚群均含有与电离辐射抗性相关基因的差异突变;EARL等[10]鉴定出irrE亚群中的唯一菌株IRS24,此菌株不仅对电离辐射抗性低而且对紫外辐射及丝裂霉素C都敏感,研究发现导致这些抗性减弱是由于irrE突变所致,irrE位于耐辐射异常球菌的1号染色体,编号为DR_0167,全长987 bp,编码328个氨基酸,大小为35 kD。IrrE蛋白是由3个结构域构成的独特组合:一个类锌肽结构域,一个HTH(螺旋-转角-螺旋)结构域和一个类GAF结构域[11]。LU等[12-13]通过分析irrE突变株与野生型菌株受到辐射前后的蛋白组差异,发现irrE存在时,31种蛋白表达量显著上调,除RecA与PprA参与DNA损伤修复[14-15],其他蛋白涉及了转录调控、能量代谢和应激反应等6个不同通路[12]。2014年,LUDANYI等[16]通过体内和体外实验证明IrrE是一种金属蛋白酶,可以与DdrO(DNA损伤诱导基因的负调节因子)相互作用[17],使其降解并促进DNA损伤诱导基因的表达,进而提高耐辐射异常球菌的辐射、干旱等抗性。此外,乐东海等[18]将irrE转入大肠杆菌,转化菌株的抗氧化性显著提高;在枯草芽胞杆菌中表达irrE,也能增强细胞的抗氧化与紫外辐射能力[19],PAN等[20]将此基因转入大肠杆菌,IrrE蛋白显著提高大肠杆菌细胞对盐、氧化、渗透和热等多种非生物胁迫的耐受性,同时验证了该基因提高油菜耐盐能力的作用。上述研究结果说明irrE对生物耐逆性提高具有显著的生物学功能。【本研究切入点】IrrE对耐辐射异常球菌的干旱和辐射等抗性具有重要作用,且相关研究表明irrE异源表达可显著提高宿主的抗逆性[21]。然而,按照植物密码子优化irrE后导入植物,对植物抗逆性的提高目前仍没有报道。研究优化的irrE转基因烟草的耐盐耐旱性,进一步了解irrE在植物中的抗逆功能。【拟解决的关键问题】将优化的irrE转入烟草,鉴定转基因烟草的耐盐耐旱性,并分析转基因植物的生理生化变化,以期获得改造后的耐辐射异常球菌irrE可显著提高转基因植物耐旱耐盐性,为植物耐逆基因工程提供优良基因源。1 材料与方法
试验于2016年在中国农业科学院生物技术研究所完成。1.1 试验材料
irrE序列来源于中国农业科学院生物技术研究所林敏研究员课题组,基因合成由上海生工生物工程有限公司完成,中间载体pUC-19、植物表达载体pBI-121、农杆菌GV3101感受态、烟草种子NC89均由生物技术研究所郭三堆研究员课题组保存。质粒小提试剂盒购于GenStar公司,Hind Ⅲ、EcoRⅠ等内切酶购于NEB公司,胶回收试剂盒购于Omega公司,植物DNA提取试剂盒购于天根公司,植物RNA提取试剂盒购于原平皓公司,反转录试剂盒和实时荧光定量PCR(qRT-PCR)试剂盒均购于全式金公司,高保真DNA聚合酶、In-Fusion HD Cloning Kit购于TaKaRa公司,常规试剂购于拜尔迪公司,引物(表1)由上海生工生物工程公司合成。Western blot用一抗由中国农业科学院生物技术研究所陆伟老师提供,其他试剂均购于康为试剂公司。Table 1
表1
表1引物信息
Table 1Primer information
| 引物名称 Primer name | 正向引物序列 Former primer sequences (5′-3′) | 反向引物序列 Later primer sequences (5′-3′) |
|---|---|---|
| Linker-02 | AGCTCCCCGAATTCCCCC | AGCTGGGGGAATTCGGGG |
| irrE PCR | CCCTATCAGCCTGTTCAGAGT | TGGTGTGAGAGGAGGTGGTAT |
| irrE qRT-PCR | GCTGCAAAGGCTAGGATGAG | GTGAACCTCTGCCTCTCAGG |
| NtActin9 | AGGACCTCAGGACAACGGAAACG | CTATTCTCCGCTTTGGACTTGGCA |
| NtABF2 | GATCCGGCGATCAATTCTAA | TTCTCCTCTGCCTCCTTTCA |
| NtzfP | ACTGGTCGTTCACACGTCTGTT | AGGCATGTTCAGGTCAAAGTCA |
| NtCDPK | CAGTGGCTACATTACCGCAGAC | GGTTTCCCTTTTGCATCATAGC |
| NtLEA5 | TCCAAACTCATCTCTGCTTTCG | AGCAGCGTCAATCTCTTTAGCA |
新窗口打开
1.2 植物表达载体的构建
为了转基因株系易于筛选,对pBI-121载体进行改造,利用GR79-EPSPS[22]替换NPTII抗性标记基因,获得植物表达载体GBI。irrE编码序列长987 bp,进行密码子优化并将基因内部的PstⅠ、XhoⅠ、Hind Ⅲ、EcoRⅠ和BamHⅠ酶切位点钝化,在ORF两端加上PstⅠ和XhoⅠ酶切位点,连接于中间载体上,获得中间载体pUC-irrE,载体表达盒的一端用Hind Ⅲ酶切后,插入linker-02,钝化Hind Ⅲ位点,插入EcoRⅠ位点,用EcoRⅠ单酶切的方式切下irrE表达盒片段,连接到GBI上,获得植物高效表达载体GBI-IE。1.3 转化烟草及阳性植株鉴定
将GBI-IE载体转化农杆菌GV3101感受态,叶盘法转化烟草,使用草甘膦筛选培养4—5周后,愈伤组织开始分化,待再生苗长至3—4片叶子时转接到生根培养基,幼苗长至10 cm左右开盖,PCR鉴定阳性转基因植株后,炼苗1—2 d,移栽至培养盆于温室培养。1.4 实时荧光定量PCR(qRT-PCR)分析
取幼苗自顶部向下第3叶,利用植物RNA提取试剂盒(TIANGEN,DP432)提取材料总RNA,反转录获得cDNA,利用KOD SYBR qPCR MIX(TOYOBO,QKD-201)进行实时荧光定量PCR检测,应用BIO-RAD Mini Opticon荧光定量分析仪进行qRT-PCR反应和分析。烟草NtActin9作为内参基因。按照2-ΔΔCt法计算目的基因的相对表达量[23]。1.5 Western blot检测
利用RIPA裂解液提取烟草叶片总蛋白,煮沸变性后,稀释5倍取30 µL进行SDS-PAGE电泳,浓缩胶电压为80 V,待溴酚蓝指示剂迁移至分离胶、分子量标准各条带略有分开后,换电压至120 V,待溴酚蓝迁移至距离凝胶底部0.5 cm时结束电泳;转膜,100 mA(约40 V)稳流转100 min;5% BSA封闭1 h;一抗按照1﹕5 000倍稀释,4℃孵育过夜,TBST洗膜3次;二抗按照1﹕5 000比例稀释,孵育1 h,TBST洗膜4次,曝光[24]。1.6 转基因烟草的耐盐耐旱性分析
取适量野生型和转基因烟草种子于无菌2 mL离心管中,75%酒精消毒30 s,无菌水洗一次,15% NaClO 25℃消毒30 min,无菌水洗5—6遍,点播于含有250 mmol·L-1 NaCl和300 mmol·L-1甘露醇的1/2 MS培养基上,黑暗1 d后,28℃光照培养,每隔24 h统计发芽率;正常萌发的种子移栽到250 mmol·L-1 NaCl和300 mmol·L-1甘露醇条件12 d后,统计根长、侧根数和鲜重,每个株系取10株;烟草于温室培养30 d后,250 mmol·L-1 NaCl和300 mmol·L-1甘露醇处理,24 h后取材料检测irrE和相关胁迫响应因子的表达量,一周后采集照片并测定SOD、CAT活性和MDA含量,每个株系取6株。2 结果
2.1 密码子优化
irrE来自于耐辐射异常球菌,基因全长987 bp,编码329个氨基酸,在转入植物之前,与植物基因组比对,其中有241个氨基酸的密码子使用的是非植物偏爱密码子,占比达到73.25%。理论上来说,若不对其进行密码子优化,直接导入植物,会造成基因表达量低甚至不表达[25]。因此,按植物密码子对其进行了优化,旨在让irrE在植物中正确翻译并高效表达。共优化290个碱基,涉及241个密码子,编码蛋白氨基酸序列和改造前完全相同(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1棉花偏好性密码子优化前后irrE编码区核苷酸(A)和编码蛋白氨基酸(B)序列比较
-->Fig. 1Comparison of irrE encoding nucleotide (A) and amino acid (B) sequences before and after the cotton preference codon optimized
-->
2.2 irrE植物表达载体构建和阳性转基因烟草分子鉴定
棉花遗传转化费时费力,为提高转基因苗阳性率,首先将载体pBI-121卡拉霉素抗性标记基因NPTII替换成为草甘膦抗性基因GR79-EPSPS。将irrE与Cozk、Kozak和ployA等元件连接后克隆到GBI载体上,构建植物表达载体GBI-IE(图2-A)。叶盘法转化烟草(图2-B),使用100 mg·L-1草甘膦筛选和PCR检测,共获得15个独立的转基因阳性株系(图2-C)。根据qRT-PCR检测结果,选择2个表达量最高的转基因株系,并命名为GO1和GO2进行耐盐耐旱试验(图2-D),为确定irrE在转基因烟草中蛋白正常翻译,进一步利用Western blot检测转基因烟草GO1和GO2中目标蛋白的表达(图2-E),转基因烟草在35 kD处清晰地检测到IrrE蛋白表达,而野生型非转基因对照完全没有条带,说明GO1和GO2中IrrE编码蛋白正确的翻译。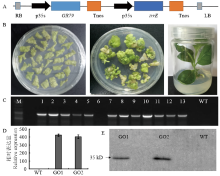 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2irrE植物表达载体改造、构建和转基因烟草获得与鉴定
A:载体图谱;B:转基因再生株系的获得;C:T0代阳性植株PCR鉴定,M:DNA Marker,CK:野生型对照,1—13:转基因株系;D:GO1和GO2中irrE的表达量检测;E:蛋白免疫印迹检测IrrE蛋白
-->Fig. 2Construction of plant expression vector, acquisition and identification of transgenic tobacco
A: Vector map; B: Obtainment of transgenic regeneration lines; C: Identification of T0 generation plants by PCR, M: DNA Marker, CK: WT, 1-13: Transgenic lines; D: Expression level of irrE in GO1 and GO2; E: Detection of IrrE protein by Western blot
-->
2.3 逆境条件下irrE转基因烟草萌发率
在正常1/2 MS培养基上,转基因株系GO1和GO2发芽率和野生型对照之间没有明显差异(图3-A和图3-B);而250 mmol·L-1 NaCl培养基中,GO1和GO2萌发率分别为78.8%和90.0%,野生型仅为10.3%(图3-C);300 mmol·L-1甘露醇条件下,与野生型对照相比,转基因株系GO1和GO2萌发率分别提高了42.9%和50.8%(图3-D)。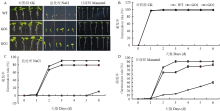 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3转irrE烟草种子在盐处理和甘露醇处理下的萌发率
A:野生型和GO1、GO2在不同培养基上第8天的萌发情况;B:野生型和GO1、GO2在1/2 MS培养基上的萌发率;C:野生型和GO1、GO2在含有250 mmol·L-1 NaCl培养基上的萌发率;D:野生型和GO1、GO2在含有300 mmol·L-1甘露醇培养基上的萌发率
-->Fig. 3The germination rate of transgenic tobacco seeds under salt and mannitol treatments
A: Germination of WT, GO1, and GO2 on different culture media on 8th day; B: Germination rates of WT, GO1, and GO2 on 1/2 MS medium; C: Germination rates of WT, GO1, and GO2 on 250 mmol·L-1 NaCl medium; D: Germination rates of WT, GO1, and GO2 on 300 mmol·L-1 mannitol medium
-->
2.4 irrE转基因烟草耐盐和耐旱性
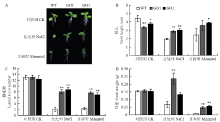
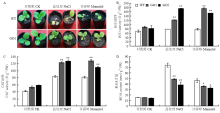
正常萌发的种子移栽到正常培养基12 d后,转基因GO1和GO2与对照组生长基本一致,根长(图4-A和图4-B)、侧根数(图4-C)和鲜重(图4-D)无显著差异;而移栽到含250 mmol·L-1 NaCl培养基上,与对照相比,转基因GO1和GO2的平均根长提高46.9%和53.3%,平均侧根数提高3和3.34倍,平均鲜重提高2.29和2.31倍;移栽到300 mmol·L-1甘露醇培养基上,转基因GO1和GO2的平均根长增加46.1%和58.4%,平均侧根数增加2.29和2倍,平均鲜重增加3和3.33倍。将正常萌发的GO1、GO2和野生型烟草移栽至温室中培养30 d后,进行盐和干旱处理。与野生型相比,irrE转基因烟草无论耐旱还是耐盐害能力均明显提高(图5-A)。
正常生长条件下,GO1、GO2与野生型SOD(图5-B)、CAT活性(图5-C)和MDA含量(图5-D)无明显差异;250 mmol·L-1 NaCl处理后,野生型烟草的SOD活性为79.08 U·g-1 FW,转基因GO1和GO2的SOD活性分别为122.07和175.84 U·g-1 FW,野生型烟草的CAT活性为84.00 U·g-1 FW,转基因烟草GO1和GO2的CAT活性分别为123.74和127.81 U·g-1 FW;野生型烟草的MDA含量为74.13 nmol·g-1 FW,转基因烟草GO1和GO2的MDA含量分别为28.70和28.16 nmol·g-1 FW;干旱处理后,转基因烟草GO1和GO2的SOD活性比野生型分别提升了56.27和74.58 U·g-1 FW,CAT活性提升了26.65和41.55 U·g-1 FW,MDA含量降低了24.73和27.83 nmol·g-1 FW。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4转基因株系和野生型根长、侧根数和鲜重比较
A:250 mmol·L-1 NaCl和300 mmol·L-1甘露醇条件12 d后野生型和转基因GO1和GO2的烟草;B:根长;C:侧根数;D:鲜重
“*”和“**”分别表示在P<0.05 和P<0.01水平上差异显著。下同
-->Fig. 4Comparison of root length, lateral roots number and fresh weight of transgenic lines and WT
A: After treatment of 250 mmol·L-1 NaCl and 300 mmol·L-1 mannitol for 12 days, WT and GO1, GO2; B: Root length; C: Lateral roots number; D: Fresh weight
“* ” and “**” indicate significant difference at P<0.05 and P<0.01 level, respectively. The same as below
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5irrE转基因烟草在温室中的抗旱耐盐性鉴定和酶活性检测
A:正常生长条件和胁迫处理的表型(野生型和GO1株系),盐处理:250 mmol·L-1 NaCl,干旱处理:300 mmol·L-1甘露醇;B:SOD活性;C:CAT活性;D:MDA含量
-->Fig. 5Identification of drought resistance and salt tolerance of transgenic tobacco in greenhouse and enzyme activity detection
A: Performance of control group and stress treatment (WT and GO1), Salt treatment: 250 mmol·L-1 NaCl, Drought treatment: 300 mmol·L-1 mannitol; B: SOD activity; C: CAT activity; D: MDA content
-->
2.5 转基因株系中逆境应答基因表达
qRT-PCR分析表明,250 mmol·L-1 NaCl和300 mmol·L-1甘露醇处理下,胁迫应答因子NtABF2(ABRE binding factor)[26]、Ntzfp(osmotic stress-induced zinc-finger protein)[27]、NtCDPK2(calcium dependent protein kinase)[28]以及水通道蛋白NtLEA5[29]等编码基因表达量均显著上调(图6)。NtABF2在正常生长条件下,野生型和转基因GO1、GO2表达量基本一致,然而盐处理后转基因烟草植株的表达量比野生型提高了2倍,干旱处理后提高了1倍;Ntzfp、NtCDPK2、NtLEA5无论正常生长条件下还是盐胁迫和甘露醇处理下,转基因植株的表达量均显著高于野生型。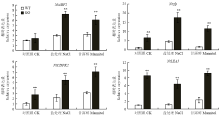 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6盐和干旱处理下WT和irrE转基因烟草胁迫响应相关基因的表达分析
-->Fig. 6Expression analyses of stress response genes in WT and irrE transgenic plants under salt and drought conditions
-->
3 讨论
植物对逆境的耐受性是一个复杂的性状,目前已经发现植物中有100多个基因参与逆境响应[30],为了提高作物对干旱、盐碱和低温等的耐受性,很多研究者将与逆境响应转录因子、渗透保护物质生物合成相关的基因等转入植物,一定程度上提高了植物的耐逆能力[31-33]。然而人为干预植物逆境响应或调节基因表达,往往对植物的生长发育产生不同程度的影响,进而影响其产量和品质。利用苏云金芽孢杆菌Bt提高棉花抗棉铃虫能力的成功,为植物耐逆基因工程提供了新的思路。利用改造的微生物耐逆关键基因改良植物耐逆性是否会取得同样的成功?目前,仅有零星利用微生物基因改良植物耐逆性的报道[34-35]。本研究中irrE来源于耐辐射异常球菌,其表达受到辐射和干旱的强烈诱导[13],推测其编码蛋白与DNA修复密切相关。PAN等[20]在大肠杆菌和油菜中表达irrE,盐耐受性均显著提高,临界盐浓度350 mmol·L-1处理6周,转基因油菜生长状况良好,而野生型在盐处理2周后死亡。本研究对irrE进行植物密码子优化,通过融合多个表达增强序列,利用叶盘法转化烟草,转录水平和翻译水平检测均证明irrE在转基因烟草中表达。正常条件下,转基因烟草与野生型长势基本一致,说明irrE导入不干扰转基因植株的生长。然而,250 mmol·L-1 NaCl和300 mmol·L-1甘露醇胁迫下,转基因烟草对高盐和干旱的耐受性均明显高于野生型,说明irrE可以提高植物耐盐和耐旱能力。此外,前期预实验证明,野生型烟草在临界盐浓度200 mmol·L-1 NaCl处理2周后即全部死亡,说明按照植物密码子优化的irrE导入烟草后,可使转基因烟草获得比临界盐浓度更高的耐盐性。植物在逆境胁迫下,体内会产生大量的超氧根阴离子和过氧化氢,SOD可以清除超氧根阴离子,CAT清除过氧化氢,减少对细胞的伤害;MDA是生物体内膜脂过氧化的产物,反映了膜脂过氧化的程度,其值越高,细胞膜受伤害程度越深;这3个指标是反应植物细胞受伤害程度的重要指标[36-37]。通过对转基因烟草SOD、CAT酶活性和MDA含量进行测定,发现正常生长条件三者含量(活性)水平和野生型之间没有显著差异,而干旱和盐逆境下SOD、CAT酶活性显著提高,MDA含量显著降低,这一结果说明转基因烟草的抗氧化能力显著高于野生型,膜脂过氧化程度低于野生型,从而提高转基因植物在逆境下的成活率,与奉斌等的研究结果一致[38]。其中,CAT活性比非转基因烟草平均提高48.80%,SOD活性比非转基因烟草平均提高88.55%,而转细菌irrE的油菜中CAT表达量比非转基因油菜平均提高约40%,SOD表达量比非转基因油菜平均提高约70%,说明转优化的irrE植物中的SOD和CAT表达量均高于转细菌irrE植物,优化的irrE可使转基因植物的清除活性氧能力更强。
耐逆基因的表达对植物耐逆性具有直接的指示作用。董仕波等[39]研究表明,irrE可以通过提高Na+/H+转运蛋白AtNHX1的表达,进而提高转基因拟南芥的盐胁迫耐受性。而在本研究中,实时荧光定量PCR结果证明irrE转基因烟草中,ABRE结合因子NtABF2、逆境诱导锌指蛋白Ntzfp、钙依赖蛋白激酶NtCDPK2、水通道蛋白NtLEA5表达量在盐和干旱胁迫的条件下均显著高于野生型,这些不同类型逆境响应基因的表达升高也暗示irrE可能通过激活植物多种逆境响应提高植物的抗旱和耐盐性。另外,本研究发现异源表达irrE转基因烟草种子萌发率在甘露醇和盐逆境下显著高于非转基因对照。萌发率和成苗率往往存在直接的正相关,对于很多作物而言,干旱和盐碱条件下,尤其是东部沿海盐碱地和西北次生盐碱地,种子萌发和成苗对于后期的产量具有决定性的作用。转基因烟草逆境下萌发率的提高暗示irrE对于盐碱地作物改良具有较好的应用前景。
4 结论
根据植物密码子优化耐辐射异常球菌irrE并转化模式植物烟草,转基因烟草无论耐旱还是耐盐能力均显著提高,表现为在逆境条件下种子萌发率升高、苗期存活率提高、清除活性氧能力增强和多类逆境响应基因表达激活。推测irrE可通过多种胁迫响应途径正调控植物的耐盐耐旱性。致谢:中国农业科学院生物技术研究所林敏研究员提供了irrE;周正富老师提供了相关资料并在研究过程中给予了帮助;陆伟老师提供了IrrE蛋白一抗。在此一并表示感谢!
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}