0 引言
【研究意义】新疆红肉苹果(Malus sieversii f. niedzwetzkyana (Dieck) Langenf)是新疆野苹果的变型,其富含花青苷等次生代谢产物,具有花、果、叶、枝均为红色的特点,是现代苹果品质育种的重要种质;但其高花青苷性状缺乏挖掘与利用,成为制约单一传统栽培品种向特色多样化发展的重要因素[1]。因此,以新疆红肉苹果与栽培苹果品种的杂交分离群体为试材,积极探讨MdMYB308响应外源分裂素调控花青苷合成代谢机制,完善苹果红色性状遗传与发育机理,对传统品种的性状改良及丰富栽培苹果遗传基因库具有重要意义[2-3]。【前人研究进展】花青苷生物合成是植物类黄酮合成途径的重要分支。利用模式植物同源克隆技术已从大多数植物中分离鉴定了花青苷合成酶及相关转运蛋白基因 [4-7]。然而,除了植物本身的种性(遗传性)外,花青苷的合成同时受到外界环境(光、温度、激素等)的调控[8-12]。细胞分裂素作为调控花青苷合成的重要激素之一,已在拟南芥、甘蓝、萝卜等植物中筛选与验证[13-15]。研究发现,6-BA可以诱导结构基因CHI、DFR、PAL1和CHS的表达,促进拟南芥叶片花青苷的累积,且过表达分裂素合成基因IPT的植株比野生型花青苷含量高[15]。在转录水平上,外源细胞分裂素有利于MYB类转录基因PAP1的表达从而间接促进植物花青苷的积累[16]。此外,AtMYBL2转录因子是具有EAR结构的MYB类抑制子,研究表明细胞分裂素可以抑制AtMYBL2的表达而增强拟南芥中花青苷的代谢合成[17]。【本研究切入点】笔者课题组于2006年率先建立了以新疆野苹果及其红肉变性为亲本的杂种分离群体[18]。针对杂交一代中果实质地、类黄酮含量、香气等性状遗传变异特点,对其不同株系间品质性状比较及相关基因表达分析,并深入探讨激素与氮对花青苷生物合成的影响[19-22],但外源分裂素调控苹果花青苷代谢机理及相关MYB转录因子的研究,尚未见报道。【拟解决的关键问题】采用不同浓度6-BA处理红肉愈伤组织,并进行花青苷相关基因的荧光定量分析,通过酵母双杂交与双分子荧光互补(BIFC)试验研究MdMYB308与MdbHLH3的关系,探讨外源分裂素对苹果花青苷MYB转录因子的调控机制,为研究红肉苹果花青苷代谢分子机理与高品质苹果品种选育提供参考。1 材料与方法
试验于2016年在山东农业大学园艺科学与工程学院进行。1.1 试验材料及处理
以新疆红肉苹果与富士杂交F1代分离群体中选出的‘紫红3号’优株幼嫩叶片诱导的红色愈伤组织为试材,将正常培养(0.3 mg∙L-1 NAA+0.6 mg∙L-1 6-BA)的愈伤组织转接到NAA浓度为0.3 mg∙L-1、6-BA浓度分别为0、0.2、0.4、0.6、0.8和1 mg∙L-1的MS培养基(Solarbio)上。每天进行16 h的光照培养,光强为2 000—2 500 lx,温度设置为(24±2)℃。处理20 d后取样,液氮速冻,-80℃保存。1.2 植物总RNA的提取与qRT-PCR
总RNA的提取方法参照TianGen RNA Plant Reagent 操作说明。反转录利用Fermentas公司生产的ReverAidTM First Strand cDNA Synthesis Kit试剂盒合成cDNA。荧光定量使用伯乐 CFX96实时定量PCR仪,总反应体系20 μL,包括10 μL 2.5×RealMasterMix/ 20×SYBR Solution,1 μL cDNA模板(50 ng·μL-1),上下游引物各1 μL (5 μmol·L-1),7 μL ddH2O。运行程序为:①95.0℃预变性 30 s;②95.0℃变性5 s;③58℃退火10 s;④72.0℃延伸30 s(②—④共45个循环);⑤65℃孵育 20 s;⑥溶解温度从55℃到95℃每升高0.5℃保持1 s;⑦停止反应。每个样品设置3个重复。以苹果肌动蛋白MdActin作为内参,数据的分析采用2-ΔΔCT方法。1.3 苹果MdMYB308的克隆、生物信息学分析
在苹果基因组中根据拟南芥中AtMYBL2蛋白序列进行Blast比对得到了一个基因序列号为MDP0000249611的基因,分别设计上下游引物F:5′-ATGGGAAGGTCTCCTTGC-3′,R:5′-TCATTTCA TCTCCAAGCTTCT-3′。具体反应体系:cDNA 2 μL,上下游引物各2.5 μL,5×HFbuffer 10 μL,10 mmol·L-1dNTP 1 μL,酶0.5 μL,ddH2O 31.5 μL。PCR程序为:98℃ 30 s;98℃ 10 s,56℃ 30 s,72℃ 45 s,30个循环;72℃ 10 min。PCR产物用1%的琼脂糖凝胶进行电泳并回收目的条带,连接PLB零背景载体(VT205)进行测序。MdMYB308蛋白通过Expasy网站(http://web. expasy.org/protparam/)进行理化性质分析,NCBI软件Blast比对蛋白质氨基酸序列,MEGA5.0软件构建系统进化树。
1.4 花青苷含量的测定
称取愈伤组织0.25 g,液氮研磨至粉末加入10 mL 1%HCL甲醇,4℃浸提24 h,12 500 r/min离心25 min,吸取上清液进行测定。以盐酸甲醇为空白,利用分光光度计测定样品提取液在530 nm下的吸光值,每g样品的吸光值为花青苷含量(Abs·g-1 FW)。1.5 酵母双杂试验
用引物5′-CCATATGATGGGAAGGTCTCCTTGC -3′和5′-CGGATCCTCATTTCATCTCCAAGCTTCT-3′扩增MdMYB308的编码框序列,用引物5′-GGAATTC ATGGCTGCACCGCC-3′和5′-GGATCCTTAAGAGTC AGATTGGGGTATAATTT-3′扩增MdbHLH3的编码框序列,将胶回收产物连接PLB零背景载体。酶切连接构建pGBKT7-MdbHLH3和pGADT7- MdMYB308重组载体。将重组载体质粒同时转化到酵母Y2H感受态细胞,分别培养在二缺-Leu/Trp与四缺-Ade/-His/ -Leu/-Trp选择性培养基中,并在四缺培养基中添加X-α-gal进行检测。1.6 双分子荧光互补试验
用引物5′-CGGGATCCATGGGAAGGTCTCCTT GC-3′和5′-GCGTCGACTTTCATCTCCAAGCTTCTG TA-3′扩增MdMYB308的编码框序列,用引物5′-GGA TCCATGGCTGCACCGCC-3′和5′-GGGGTACCAGA GTCAGATTGGGGTATAATTTG-3′扩增MdbHLH3的编码框序列,之后连接PLB零背景载体。用Bam H I和Sal I对MdMYB308-PLB和pSPYNE分别进行双酶切,用Bam H I和Kpn I对MdbHLH3-PLB和pSPYCE分别进行双酶切,构建NYFP-MdbHLH3和CYFP-MdMY308重组载体。将这两个重组载体分别转化农杆菌LBA4404感受态细胞,振荡培养至OD600约为0.8,洋葱内表皮浸入农杆菌菌液30 min,转入附加乙酰丁香酮(100 μmol·L-1)的MS固体培养基中,28℃暗培养1—2 d。在激光扫描共聚焦显微镜(laser scanning confocal microscope,Zeiss 510 Meta)下观察、扫描。2 结果
2.1 苹果MdMYB308的克隆与生物信息学分析
如图1所示,以‘紫红3号’诱导愈伤组织cDNA为模板,克隆得到一条大小约800 bp的条带。对克隆片段回收测序发现,该基因的开放阅读框长度为768 bp,编码255个氨基酸,蛋白质分子量为28.37 kD,理论等电点为8.94。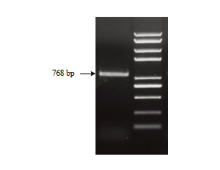 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1MdMYB308 编码区全长的RT-PCR扩增
-->Fig. 1Agarose gel electrophoresis of MdMYB308 RT-PCR products
-->
如图2所示,对来自不同物种的MYB家族基因构建系统进化树。其中,苹果MdMYB308与FaMYB1、AtMYBL2、AtMYB4在同一进化枝上,因此推测苹果中MdMYB308与FaMYB1、AtMYBL2、AtMYB4功能相似。利用NCBI蛋白结构分析发现,MdMYB308具有EAR抑制序列(图3)。
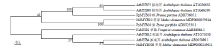 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2花青苷合成相关MYB转录因子进化树分析
-->Fig. 2Phylogenetic tree of relative MYB transcription factors in anthocyanin synthesis
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3花青苷合成MYB转录因子氨基酸序列比对
-->Fig. 3Multiple sequence alignment of MYB transcription factors from various plants
-->
2.2 愈伤组织花青苷积累对细胞分裂素浓度的响应试验
从图4、5可以看出,随着培养基6-BA浓度的升高,愈伤组织中的颜色逐渐变深,花青苷含量也呈上升趋势,6-BA浓度到达1 mg∙L-1时,花青苷含量最高。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4不同分裂素浓度下的苹果愈伤组织
-->Fig. 4The phenotype of red flesh apple callus on media added different concentration of cytokinin
-->
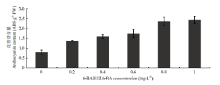 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5不同分裂素浓度对苹果愈伤组织花青苷含量的影响
-->Fig. 5Anthocyanin content in callus growth on different 6-BA concentration
-->
如图6所示,愈伤组织在含1 mg∙L-1 6-BA的培养基培养20 d后,与无分裂素培养基(对照)相比,花青苷合成相关基因表达量都有不同程度的上升,其中CHS、DFR、UFGT表达量约为对照的3—4倍,转录基因MYB10、bHLH3的表达量是对照的4倍左右,而MYB308表达明显被抑制。
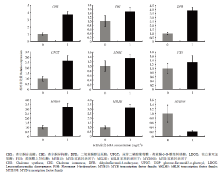 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6苹果花青苷合成相关基因对1 mg∙L-1 6-BA的响应
-->Fig. 6Dynamic change of anthocyanin synthesis-related genes expression to 1 mg∙L-1 6-BA
-->
2.3 MdMYB308 与 MdbHLH3 酵母双杂试验
由图7酵母双杂交试验可得,空载体pGADT7和MdbHLH3+pGBKT7、空载体pGBKT7和MdMYB308 +pGADT7共转Y2H均在二缺板上生长,在四缺及四缺+X-α-gal不生长;而MdbHLH3和MdMYB308共转Y2H在二缺、四缺及四缺+X-α-gal都能生长,因此推测其在酵母体内能够相互作用。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7MdMYB308和 MdbHLH3酵母双杂交
-->Fig. 7Yeast two-hybrid in MdMYB308 and MdbHLH3
-->
2.4 MdMYB308与MdbHLH3 双分子荧光互补试验
将MdMYB308-NYFP和MdbHLH3-CYFP共同转入洋葱表皮细胞,如图8所示,通过激光共焦显微镜扫描得到洋葱表皮核内产生黄色YFP荧光信号。此外,单独用空载CYFP与MdMYB308-NYFP共同转入洋葱表皮细胞中没有检测到YFP信号,因此推测其在植物体内能够相互作用。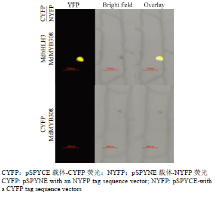 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8MdMYB308和MdbHLH3双分子荧光互补试验
-->Fig. 8BiFC assay to detect the interactions of MdMYB308 and MdbHLH3 protein
-->
3 讨论
外源激素作为环境因素之一,对植物花青苷代谢合成具有重要作用[20-22]。其中,细胞分裂素促进植物花青苷合成的研究已有诸多报道[23-24]。前人研究发现,6-BA浓度在10-9—10-6 μmol∙L-1有利于萝卜细胞中花青苷的累积,但超过10-6 μmol∙L-1将抑制花青苷的合成[13]。在拟南芥中,细胞分裂素可以激活花青苷结构基因PAL1、CHS、CHI、DFR以及转录因子PAP1的表达[15-16]。本研究发现,提高6-BA浓度促进红肉苹果愈伤组织的花青苷合成;花青苷结构基因MdCHS、MdDFR、MdUFGT与相关转录因子MdbHLH3、MdMYB10都有不同程度上升,而MdMYB308显著下降,说明细胞分裂素促进红肉苹果愈伤组织花青苷的合成与结构基因以及MYB类转录因子的差异表达有关。目前,EAR元件是广泛存在于植物转录因子中的一段保守抑制序列,在植物防御及非生物胁迫中具有重要作用[25-27]。MYB蛋白家族是植物中成员最多的转录因子家族之一。在拟南芥中,AtMYB4蛋白首先被报道其C端存在EAR序列并通过直接绑定下游结构基因启动子发挥抑制作用[28-29]。在高等植物花青苷合成代谢途径中,R2R3-MYB家族蛋白中的FaMYB1存在EAR元件,异源表达FaMYB1抑制烟草中花青苷的合成[30-31]。DUBOS等[17]发现,AtMYBL2作为一种存在EAR元件的新型抑制子与EGL3、GL3、bHLH转录蛋白结合调控花青苷的合成,而细胞分裂素可以降低AtMYBL2的表达。本研究发现,MdMYB308蛋白C端存在EAR元件并与MdbHLH3蛋白互作,说明MdMYB308可能与MdbHLH3结合抑制苹果花青苷的合成。
针对果树特色种质或新品种与传统栽培品种在品质性状形成与调控机理方面的差异特点,借鉴模式植物的最新研究进展与成果,结合现代分子生物技术与果树配套栽培方法,完善理论体系解决生产难题,是近年来果树研究领域理论联系实践的重要趋势。激素调控植物花青苷代谢合成已成为近年来的研究热点。本研究结果为苹果生产栽培技术提供理论支持,并为进一步选育具有高花青苷的苹果品种奠定了基础。
4 结论
在‘紫红3号’叶片诱导的苹果愈伤中,克隆了MdMYB308,进化树分析证明MdMYB308与AtMYB4、FaMYB1、AtMYBL2在同一进化枝上,氨基酸序列比对发现MdMYB308存在EAR抑制序列;MdMYB308的表达受细胞分裂素6-BA的抑制;MdMYB308可以与MdbHLH3结合。结果表明MdMYB308可能作为抑制子参与细胞分裂素调控花青苷的合成。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}