0 引言
【研究意义】小麦叶锈病是由小麦叶锈菌(Puccinia triticina)侵染引起的真菌性气传小麦病害,是危害中国小麦产量的重要因素之一,病害严重流行年份可导致产量损失达40%以上[1]。叶锈病发生范围广泛,20世纪70年代在墨西哥西北部发生叶锈病大流行,据估计造成产量损失高达70%[2]。中国北方麦区曾在1969、1973、1975和1979年发生过中度以上病害流行[3],2012年和2015年在多个小麦主产区暴发了小麦叶锈病,对小麦生产造成了严重危害[4-5]。因此不断发掘和定位小麦抗叶锈病基因对丰富中国小麦抗病基因库和培育抗病品种具有重要意义。【前人研究进展】小麦对叶锈病的抗性分为两种:垂直抗性和水平抗性。垂直抗性受单基因控制,表现为小种专化抗性,常因小种变异而丧失;水平抗性大多数受微效多基因控制,具有数量遗传特性,为非小种专化抗性。微效多基因一般具有加性效应和上位效应,不易引起抗病性丧失,表现为持久抗性,比如水平抗病品种Pavon76在生产上利用多年仍保持较好抗性,经分子检测品种Pavon76中携带有多个微效抗病基因[6]。目前国际上发现的100多个小麦抗叶锈病基因中大多数为苗期抗病基因,仅有Lr12、Lr13、Lr22(等位基因a和b)、Lr34、Lr35、Lr37、Lr46、Lr48、Lr49、Lr67和Lr68等12个为成株抗叶锈病基因,其中Lr34、Lr46、Lr67和Lr68为成株微效基因,具有持久抗病性[7]。抗病基因的研究方法主要有常规杂交法、基因推导法和分子标记等。基因推导法的应用原理是Flor提出的基因对基因假说[8],依据一套小麦抗叶锈病基因载体品种的表现型推导出供试材料中可能携带的抗病基因。1973年,BROWDER[9]首次应用13个近等基因系作为对照品种推导出了Lr1和 Lr10;胡长程等[10]在中国生产品种中推导出了Lr1、Lr2c、Lr10、Lr18和Lr26等抗病基因;袁军海等[11]在中国47个小麦新品种(系)中推导出Lr1、Lr3、Lr3bg、Lr9、Lr10、Lr13、Lr16、Lr23、Lr26和Lr34等10个抗叶锈病基因。由于基因推导法在小麦抗叶锈病遗传研究中普遍应用,有利于小麦抗叶锈病基因的发掘与利用,但基因推导容易受到小种鉴别力的影响,导致某些基因推导不出来,因此在对抗病基因鉴定时,常使用基因推导与分子标记法互相验证获得可靠结果。刘志勇等[12]在48份抗叶锈育种圃高代品系材料中鉴定出Lr9和Lr24;胡亚亚等[13]在14份小麦材料中鉴定出Lr1、Lr10、Lr26和Lr34等抗病基因;韩烨等[14]在来自CIMMYT品种鉴定出抗病基因Lr26、Lr34、Lr42和Lr47等;潘阳等[15]对来自104份新疆品种、高代品系中鉴定出Lr26、Lr34、Lr50、Lr3ka、Lr1和Lr14a等抗病基因;同时LI等[16]也应用基因推导法结合标记检测在中国小麦材料中发现了抗叶锈Lr1、Lr2a、Lr3bg、Lr3ka、Lr14a、Lr16、Lr17a、Lr18、Lr20、Lr23、Lr24、Lr26、Lr34和LrZH84[17]等14个抗病基因。【本研究切入点】选用中国30个小麦生产品种进行苗期基因推导、成株期抗叶锈性鉴定及分子标记检测。【拟解决的关键问题】确定这些材料可能含有的抗病基因并筛选慢锈品种,从而为作物育种和遗传研究提供有效遗传信息并为进一步抗病育种提供依据。1 材料与方法
1.1 供试品种及叶锈菌系
供试材料为陕西、贵州和湖北等3个小麦主产区的30个重要小麦生产品种,其名称、系谱及来源见表1。基因推导所用的36个已知抗叶锈病基因的载体品种和感病对照郑州5389及成株慢锈对照品种SAAR均由河北农业大学小麦叶锈病研究室提供[18];基因推导所用的18个中国小麦叶锈菌小种(PHGQ、THJT、PHJT、KHJS、PHJS、THTT①、KHHT、FHRT、FHJQ、PHTT、THTT②、PHTT、FHTR、FHHT①、FHHT②、TGGT、FHTT、FGMT)以及成株期所用的混合生理小种(FHRT、THTT、THJT)均采集于中国小麦主产区并进行单孢分离纯化,其小种命名参照LONG等[19]提出的三字母密码命名系统及后来扩展成的四字母命名法(http://wwwars.usda.gov/SP2UserFiles/ad_hoc/36400500Cerealrusts/ptnomen.pdf)。所有菌种均保存于河北农业大学小麦锈病研究室。1.2 苗期基因推导及侵染性鉴定
将对照感病品种郑州5389、36个携带已知抗叶锈病基因的载体品种和供试的30个小麦品种共67份材料,按顺序播种于温室中育苗盘内。待小麦第一片叶片完全展开,采用扫抹法将18个不同毒力的叶锈菌生理小种分别接于18套小麦叶片上。大约两周后待感病对照品种郑州5389充分发病时进行抗叶锈鉴定,参照ROELFS等[20]的6级侵染性标准,对每套小麦品种(系)进行抗叶锈病侵染型鉴定,并根据DUBIN等[21]提出的基因推导原则进行抗病基因推导。1.3 田间接种及成株期抗叶锈性鉴定
2014—2015年和2015—2016年度将32份小麦品种(包括慢锈对照品种SAAR和感病对照品种郑州5389)分别播种于河北保定河北农业大学小麦试验田和河南周口黄泛区农场试验田,种植方式采用完全随机区组设计、2次重复、行距25 cm、行长1.5 m,每10行种植一个高感对照品种郑州5389,并且将郑州5389垂直种植作为接种行。田间接种和成株抗叶锈菌鉴定可参照LI等[16]的方法。运用IBM SPSS Statistics 19.0软件进行方差分析(ANOVA),根据苗期与成株期的侵染型排除具有苗期抗性基因的品种,将田间最终严重度明显小于或与慢锈对照SAAR无显著差异的作为慢锈品种。1.4 已知抗叶锈病基因的分子检测
用CTAB法[22]提取小麦叶片基因组DNA,用TE稀释成50 μL备用,并利用分光光度计对DNA浓度和纯度进行检测。同时用ddH2O稀释DNA至终浓度40 ng·μL-1作为PCR工作液使用。采用与已知抗叶锈病Lr基因紧密连锁的11个分子标记进行分子检测。所有引物的PCR体系为20 μL,含10 μL 2×Taq PCR Mix、6 µL ddH2O、2 μL 4 mol·μL-1引物、2 μL 40 ng·μL-1 DNA模板,各引物标记扩增程序见表2,扩增后以1.5%琼脂糖凝胶电泳检测(Lr46除外);但Lr46的引物csLV46-G22对供试材料扩增后,需将其产物酶切后用12%非变性聚丙烯酰胺凝胶电泳检测。2 结果
2.1 苗期抗病性鉴定与基因推导分析
接种36个小麦抗叶锈病载体品种(系)、30个供试小麦生产品种及感病对照品种郑州5389的反应型(IT)列于表3。感病对照郑州5389对所测试的生理小种均表现高感(IT 4)。36个已知抗叶锈病载体品种(系)中,携带Lr9、Lr19、Lr24、Lr28、Lr47、Lr51和Lr53的载体品种对所有供试叶锈菌种均表现高抗(IT 0;),而携带Lr2c、Lr3、Lr16、LrB、Lr10、Lr3bg、Lr14b、Lr33和Lr44的载体品种对18个供试叶锈菌生理小种均表现为高感(IT 3或4),因此上述16个抗叶锈病基因无法通过苗期接种鉴定而推导出来,其余20个抗叶锈病基因均可以通过苗期鉴定推导出来(表1)。Lr2b只对PHGQ和FHRT两个小种表现抗病,对其余16个小种表现感病;Lr36只对PHTT和TGGT两个小种表现感病,对其余16个小种表现抗病;Lr29对4个小种表现感病,分别为PHJS、THTT①、FHHT②和FHTT号小种;另外,Lr21对PHGQ、FHRT、TGGT和FGMT这4个小种表现抗病,对其余小种表现感病;Lr45对小种FHRT和TGGT表现感病,而对其他小种表现抗病。30个小麦生产品种对18个小麦叶锈菌种表现出不同的抗性(表3)。推导结果表明鄂麦23、鄂麦15、襄麦55、鄂麦352、陕89150、鄂麦21、鄂麦11和陕旱8675等8个小麦品种对所有菌系均表现高感型,因此这些材料不含有已知抗叶锈病基因或携带未知抗病基因;Lr1对小种KHJS、KHHT、FHRT、FHJQ、FHTR、FHHT①、FHHT②、FHTT和FGMT表现低侵染型(;1),而小麦品种陕229、鄂麦14、鄂恩5号、西农979对所有Lr1无毒的小种均表现抗性,表明这些材料中携带Lr1;Lr13对编号PHTT、FHHT①、FHTT和FGMT 表现低侵染型(;1),陕229、贵农16和西农979等3个品种对此4个小种也表现相似的低侵染型,说明这些材料中可能含有Lr13;Lr26对小种TGGT和FGMT表现低侵染型(2),而小麦品种(系)鄂恩1号、鄂恩5号、鄂恩6号、贵农16、陕225、陕354、陕715、陕合6号、陕麦509和陕农7859对所有Lr26无毒的小种均表现抗性,表明这些材料可能携带有Lr26(表1)。Table 1
表1
表130个小麦品种来源、系谱及可能含有抗叶锈性基因的鉴定
Table 1Origins, pedigree and probable genes for leaf rust resistance in 30 wheat cultivars
| 品系 Cultivars (Lines) | 来源 Origin | 系谱 Pedigree | 基因推导结果 Lr gene based on gene postulation | 标记检测结果 Lr gene based on gene marker detection | 综合结果 Comprehensive result |
|---|---|---|---|---|---|
| 鄂恩1号 Een 1 | 湖北Hubei | (洛夫林10/761)F1/苏麦3号 (Luofulin 10/761) F1/Sumai 3 | Lr26,+ | Lr26 | Lr26,+ |
| 鄂恩5号 Een 5 | 湖北Hubei | 881-66/1097 | Lr1,Lr26,+ | Lr1,Lr26 | Lr1,Lr26,+ |
| 鄂恩6号 Een 6 | 湖北Hubei | “89-289”母本,“89-1012”父本杂交育成,组号为“91101” ♀(89-289) /(89-1012)♂ | Lr26,+ | Lr26 | Lr26,+ |
| 鄂麦11 Emai 11 | 湖北Hubei | 太谷显性核不育小麦作母本,进行多亲本聚合杂交经系谱法选育 TaiGu sterile lines | + | None | + |
| 鄂麦14 Emai 14 | 湖北Hubei | 繁6/偃大72-629 Fan 6/Yanda 72-629 | Lr1, + | Lr1 | Lr1,+ |
| 鄂麦15 Emai 15 | 湖北Hubei | [882-852/(鄂恩1号/Nppp-2)]/贵农11号 [882-852/(Een 1/Nppp-2)]/Guinong 11 | + | None | + |
| 鄂麦21 Emai 21 | 湖北Hubei | 由“黄冈150”系选育成 From ‘Huanggang 150’ breeding | + | None | + |
| 鄂麦23 Emai 23 | 湖北Hubei | 2078/川农8539//百农64 2078/Chuannong 8539//Bainong 64 | + | None | + |
| 鄂麦27 Emai 27 | 湖北Hubei | 扬00-123/鄂麦25 Yang 00-123/Emai 25 | + | None | + |
| 鄂麦352 Emai 352 | 湖北Hubei | 绵89-46/72103//华麦8号 Mian 89-46/72103//Huamai 8 | + | None | + |
| 鄂麦596 Emai 596 | 湖北Hubei | 郑麦9023/鄂麦12//丰优7号 Zhengmai 9023/Emai 12//Fengyou 7 | + | None | + |
| 贵农16 Guinong 16 | 贵州 Guizhou | 苏麦3号×外引C39作母本、外引P38父本 Sumai3×(♀C39/P38♂) | Lr13,Lr26,+ | Lr26 | Lr13,Lr26,+ |
| 陕225 Shaan 225 | 陕西Shaanxi | Lr26,+ | Lr26,Lr46 | Lr26,Lr46,+ | |
| 陕229 Shaan 229 | 陕西Shaanxi | 陕7853/80356 Shaan 7853/80356 | Lr1,Lr13,+ | Lr1 | Lr1,Lr13,+ |
| 陕354 Shaan 354 | 陕西Shaanxi | 陕213作母本、冬小麦167-6-4作父本杂交F6 (♀Shaan213×163-6-4♂) F6 | Lr26,+ | Lr26 | Lr26,+ |
| 陕715 Shaan 715 | 陕西Shaanxi | 88119-19-3-5-10/wx8911 | Lr26,+ | Lr26 | Lr26,+ |
| 陕89150 Shaan 89150 | 陕西Shaanxi | + | None | + | |
| 陕旱8675 Shaanhan 8675 | 陕西Shaanxi | 长武131×西植81206 Changwu 131×Xizhi 81206 | + | None | + |
| 陕合6号 Shaanhe 6 | 陕西Shaanxi | 郑州24变异株 Zhengzhou 24 variants | Lr26,+ | Lr26 | Lr26,+ |
| 陕麦509 Shaanmai 509 | 陕西Shaanxi | 母本Vp145、86585父本(小偃693×普通小麦) ♀Vp145/♂86585 (Xiaooyan 693×Common wheat) | Lr26,+ | Lr26 | Lr26,+ |
| 陕麦94 Shaanmai 94 | 陕西Shaanxi | 9229-2-2-3//中优杂交 9229-2-2-3//The optimal hybridization | + | None | + |
| 陕农7859 Shaannong 7859 | 陕西Shaanxi | 7576/327/68811(2)-2 | Lr26,+ | Lr26 | Lr26,+ |
| 陕优225 Shaanyou 225 | 陕西Shaanxi | 小偃6号/NS2761 Xiaoyan 6/NS2761 | + | None | + |
| 西农1376 Xinong 1376 | 陕西Shaanxi | 西农84G6作母本、比16作父本杂交 ♀Xinong84G6/♂Bi16 | + | None | + |
| 西农881 Xinong 881 | 陕西Shaanxi | 小偃6/西农65/83 (2)-3-3 Xiaoyan 6/Xinong 65/83(2)-3-3 | + | None | + |
| 西农979 Xinong 979 | 陕西Shaanxi | 西农2611作母本,(918×95选1)F1作父本 ♀Xinong 2611/(918×95) F1♂ | Lr1,Lr13,+ | Lr1 | Lr1,Lr13,+ |
| 襄麦55 Xiangmai 55 | 湖北Hubei | 8811/贵农24-7//鄂麦19 8811/Guinong 24-7//Emai 19 | + | None | + |
| 小偃6号 Xiaoyan 6 | 陕西Shaanxi | (ST2422/464)/小偃96 (ST2422/464)/Xiaoyan 96 | + | None | + |
| 小偃81 Xiaoyan 81 | 陕西Shaanxi | 小偃54×8602 Xiaoyan 54×8602 | + | Lr46 | Lr46,+ |
| 长武134 Changwu 134 | 陕西Shaanxi | 长武131/小黑麦代96/F1/长武131/F4/京花3号/NS2761/F1 Changwu 131/Triticale 96/F1/Changwu 131/F4/Jinghua 3/NS2761/F1 | + | None | + |
新窗口打开
Table 2
表2
表2分子标记的引物序列及PCR扩增程序
Table 2Primer sequences and PCR amplification programs for different primer combinations
| Lr | 引物Primer | 反应程序 Cycle condition | 参考文献 Reference | |
|---|---|---|---|---|
| 名称Name | 序列Sequence (5′-3′) | |||
| Lr1 | WR003F | GGGACAGAGACCTTGGTGGA | 94℃ 5 min; 35 cycles (94℃ 1 min; 55℃ 1 min; 72℃ 1 min); 72℃ 10 min;10℃ forever | QIU et al.[23] |
| WR003R | GACGATGATGATTTGCTGCTGG | |||
| Lr9 | J13/1 | TCCTTTTATTCCGCACGCCGG | 94℃ 6 min; 35 cycles (94℃ 1 min; 68.5℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | SCHACHERMAYR et al.[24] |
| J13/2 | CCACACTACCCCAAAGAGACG | |||
| Lr10 | Fl.2245 | GTGTAATGCATGCAGGTTCC | 94℃ 3 min; 35 cycles (94℃ 45 s; 60℃ 45 s; 72℃ 30 s); 72℃ 3 min; 10℃ forever | SCHACHERMAYR et al.[25] |
| Lr10-6/r2 | AGGTGTGAGTGAGTTATGTT | |||
| Lr19 | SCS265-F | GGCGGATAAGCAGAGCAGAG | 94℃ 5 min; 35 cycles (94℃ 1 min; 65℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | GUPTA et al.[26] |
| SCS265-R | GGCGGATAAGTGGGTTATGG | |||
| Lr19 | SCS253-F | GCTGGTTCCACAAAGCAAA | 94℃ 5 min; 35 cycles (94℃ 1 min; 60℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | GUPTA et al.[26] |
| SCS253-R | GGCTGGTTCCTTAGATAGGTG | |||
| Lr20 | STS638-L | ACAGCGATGAAGCAATGAAA | 94℃ 5 min; 35 cycles (94℃ 1 min; 60℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | NEU et al.[27] |
| STS638-R | GTCCAGTTGGTTGATGGAAT | |||
| Lr24 | J09/1 | TCTAGTCTGTACATGGGGGC | 94℃ 5 min; 35 cycles (94℃ 1 min; 60℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | SCHACHERMAYR et al.[28] |
| J09/2 | TGGCACATGAACTCCATACG | |||
| Lr26 | ω-secalinF | ACCTTCCTCATCTTTGTCCT | 94℃ 5 min; 35 cycles (94℃ 1 min; 65℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | CHAI et al.[29] |
| ω-secalinR | CCGATGCCTATACCACTACT | |||
| Lr26 | O11B5 | GGTACCAACAACAACAACCC | 94℃ 5 min; 35 cycles (94℃ 1 min; 65℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | FROIDMONT[30] |
| O11B3 | GTTGCTGCTGAGGTTGGTTC | |||
| Lr34 | csLv34F | GTTGGTTAAGACTGGTGATGG | 94℃ 5 min; 35 cycles (94℃ 1 min; 55℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ forever | LAGUDAH et al.[31] |
| csLv34R | TGCTTGCTATTGCTGAATAGT | |||
| Lr46 | csLV46G22-F | TCGACTTTGGAATGGAGTTGC | 94℃ 5 min; 35 cycles (94℃ 1 min; 60℃ 1 min; 72℃ 2 min); 72℃ 10 min; 10℃ Forever | SUENAGA et al.[32] |
| csLV46G22-R | GGCGAAGATGCCATCATCCACCAG | |||
新窗口打开
Table 3
表3
表330个已知抗叶锈病基因对18个叶锈菌生理小种的苗期侵染型
Table 3Seedling infection types on 30 wheat lines with known leaf rust resistance genes when tested with 18 pathotypes of P. triticina
| 品系(基因) Line (gene) | 对叶锈菌生理小种的侵染型Infection types to P. triticina race | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PHGQ | THJT | PHJT | KHJS | PHJS | THTT? | KHHT | FHRT | FHJQ | PHTT | THTT? | PHTT | FHTR | FHHT? | FHHT? | TGGT | FHTT | FGMT | |
| RL6003 (Lr1) | 4 | 3+ | 3+ | ;,1 | 4 | 4 | 1 | 1 | 1 | 4 | 4 | 4 | 1 | 1 | 1 | 4 | 1 | 1 |
| RL6016 (Lr2a) | 1 | 3+ | 2+ | 3 | 1 | 4 | 4 | 1 | 1 | 1 | 4 | 1 | 0,; | 1 | 1 | 4 | ; | 1 |
| RL6047 (Lr2C) | 4 | 4 | 4 | 3+ | 4 | 4 | 3+ | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 3+ | 3+ | 4 |
| RL6002 (Lr3) | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 4 | 4 |
| RL6010 (Lr9) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| RL6005 (Lr16) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 3 | 3 | 3 | 3 | 3 | 3+ | 3+ |
| RL6064 (Lr24) | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | ; | 0 | 0 |
| RL6078 (Lr26) | 3+ | 3+ | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 3 | 3 | 3 | 3 | 3+ | 2 | 3+ | 2 |
| RL6007 (Lr3ka) | 1 | 1 | 1 | 2 | ;,1 | 3 | 1 | 4 | 1 | 3 | 3 | 3c | 3+ | 2 | 2 | 1 | 3 | 3 |
| RL6053 (Lr11) | 4 | 4 | 3+ | 3 | 3 | 4 | 3 | 4 | 4 | 4 | 4 | 3c | 4 | 3+ | 3 | 3+ | 3+ | 2+ |
| RL6008 (Lr17) | 2 | 3c | 2+ | 1 | 3 | 4 | 1 | 1 | 1 | 3+ | 4 | 3 | 3 | 2+ | 1 | 2+ | 3 | 2 |
| RL6049 (Lr30) | 1 | 1 | 2+ | 1 | 1 | 3+ | 3c | 3 | 1 | 3+ | 4 | 4 | 4 | 3+ | 4 | 1 | 3 | 4 |
| RL6051 (LrB) | 4 | 3+ | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 4 | 3 | 4 | 4 | 3+ | 4 | 3+ | 4 |
| RL6004 (Lr10) | 3+ | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 3c | 4 | 3 | 3 | 3 | 4 | 4 | 4 |
| RL6013 (Lr14a) | x | 4 | 3+ | 4 | 3+ | 4 | 4 | 3+ | x | 4 | 3+ | 4 | X | 3+ | 3 | 3+ | 3+ | 3+ |
| RL6009 (Lr18) | 1 | 3+ | 3 | 1 | 2 | 3 | 3+ | 3 | 2 | 4 | 4 | 4 | 3 | 4 | 4 | 3+ | 4 | 3+ |
| RL6019 (Lr2b) | 2 | 4 | 3+ | 4 | 3+ | 3+ | 3+ | 2+ | 3 | 3+ | 4 | 3 | 3+ | 3+ | 3 | 3+ | 4 | 3+ |
| RL6042 (Lr3bg) | 4 | 4 | 3+ | 4 | 4 | 4 | 3+ | 3+ | 3+ | 4 | 4 | 3+ | 3 | 3+ | 3 | 3+ | 4 | 4 |
| RL4031 (Lr13) | 3 | 3 | 3 | 3 | 4 | 3 | 3+ | 3 | 3 | 3 | 4 | 2 | 3 | 2 | 3 | 3+ | 2 | 2 |
| RL6006 (Lr14b) | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 4 | 3 | 4 | 4 | 3 | 4 | 4 | 4 |
| RL6052 (Lr15) | 4 | 1 | 1 | 1 | 3+ | 4 | 1 | 1 | 1 | 1 | 4 | 1 | 3 | 1 | 1 | 1 | 2 | 1,2 |
| RL6040 (Lr19) | 0 | ; | 0 | 0 | 0 | 0 | 0 | ; | 0 | 0,; | 0 | ; | 0 | ; | 0 | 0 | 0 | 0 |
| RL6043 (Lr21) | 2 | 4 | 3 | 3 | 4 | 3 | 3 | ;,1 | 4 | 3+ | 3 | 3 | 3 | 4 | 3 | 2 | 3+ | 2+ |
| 续表3 Continued table 3 | ||||||||||||||||||
| 品系(基因) Line (gene) | 对叶锈菌生理小种的侵染型Infection types to P. triticina race | |||||||||||||||||
| PHGQ | THJT | PHJT | KHJS | PHJS | THTT? | KHHT | FHRT | FHJQ | PHTT | THTT? | PHTT | FHTR | FHHT? | FHHT? | TGGT | FHTT | FGMT | |
| RL6012 (Lr23) | 1,2 | 1 | 3 | 1 | 1 | 3 | 3 | 2+ | 3 | 3 | 1 | 1 | 1 | 1 | 1 | 3+ | 1 | 1 |
| RL6079 (Lr28) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ; | 0 | ; | 0 | ;,0 | 0 | 0 | ; | ; |
| RL6080 ( Lr29) | 1 | 1 | 1 | 2 | 3 | 1 | 2 | 2 | 2 | 1 | 2 | ; | 2 | 3 | 3 | 1 | 3 | 2 |
| RL6057 (Lr33) | 3+ | 3+ | 3+ | 3+ | 3+ | 4 | 3+ | 4 | 3+ | 4 | 4 | 4 | 3 | 3+ | 3+ | 3+ | 3+ | 3 |
| E84018 (Lr36) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 1 | 2 | 1 | 1 | 1 | 3 | 1 | 1 |
| KS86NGRC02 (Lr39)) | 3 | 2+ | 2+ | 2 | 3+ | 3 | 1 | 2 | 1 | 4 | 4 | 3 | 3 | 2 | 2 | 4 | 1 | 1 |
| KS91WGRC11 (Lr42)) | 3 | 3 | 3 | 2 | 1 | 2 | 3 | 3+ | 3 | 1 | 1 | 2 | 2 | 2 | 3 | 1 | 3+ | 3 |
| RL6147 (Lr44) | 4 | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 3 | 4 |
| RL6144 (Lr45) | 1 | 1 | 1 | ;,1 | 1 | 1 | 1 | 3 | ;,1 | 1 | 1 | 1 | 1 | 1 | 1 | 3+ | 1 | 1 |
| PAVON76 (Lr47) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ; | 0 | ; | ; | ; | ; | ; | 0 | ; |
| C78.5 (Lr51) | ; | ; | ; | 0 | ;, | ; | ; | ; | ; | 1 | 1 | ; | 1 | 1 | ; | 1 | 1 | ; |
| 98M71 (Lr53) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ; | 0 | ; | ; | ; | ; | ; | ; | 0,; |
| 1鄂麦23 (Emai 23) | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 4 | 4 | 4 | 3 | 4 | 4 | 3 | 3 | 3 | 3 | 4 |
| 2鄂麦15 (Emai 15) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 3 | 3+ | 4 | 4 | 3 | 3 | 3+ |
| 3鄂恩6号 (Een 6) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 2+ | 4 | 3+ | 3 | 3 | 3+ | 2+ | 4 | 2+ |
| 4襄麦55 (Xiangmai55) | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 4 | 4 | 3+ | 3+ | 3+ | 3+ |
| 5陕麦94 (Shaanmai 94) | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 2+ | 3 | 3 | 3 | 4 | 3 | 2+ |
| 6西农881 (Xinong 881) | 4 | 4 | 3 | 3 | 4 | 4 | 4 | 4 | 4 | 2+ | 2+ | 2+ | 2+ | 3 | 3+ | 3+ | 4 | 2 |
| 7鄂麦352 (Emai 352) | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 4 | 3+ | 4 | 3+ | 4 | 4 | 3+ |
| 8鄂麦27 (Emai 27) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 3+ | 2+ | 2+ | 3 | 3+ | 3+ | 4 | 2 |
| 9陕229 (Shaan 229) | 4 | 4 | 4 | ;,1 | 4 | 4 | 1 | 2 | 1 | 3 | 3 | 2+ | 2+ | 2+ | 2+ | 3 | 2 | 2+ |
| 10西农1376 (Xinong 1376) | 1 | ;,1 | 1 | ;,1 | 4 | ;,1 | 1 | 4 | 1 | 1 | 2+ | 2 | 3 | 2 | 1 | 3 | 3 | 2+ |
| 续表3 Continued table 3 | ||||||||||||||||||
| 品系(基因) Line (gene) | 对叶锈菌生理小种的侵染型Infection types to P. triticina race | |||||||||||||||||
| PHGQ | THJT | PHJT | KHJS | PHJS | THTT? | KHHT | FHRT | FHJQ | PHTT | THTT? | PHTT | FHTR | FHHT? | FHHT? | TGGT | FHTT | FGMT | |
| 11陕合6号 (Shaanhe 6) | 3+ | 3+ | 3+ | 3+ | 4 | 4 | 4 | 4 | 4 | 3 | 3+ | 3 | 3 | 3 | 3 | 2+ | 3 | 1 |
| 12小偃81 (Xiaoyan 81)6 | 3+ | 3+ | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 3 | 4 | 3 | 3 | 3 | 2+ |
| 13陕225 (Shaan 225) | 4 | 3 | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 2 | 4 | 2 | 2+ | 3 | 3 | 2+ | 3 | 2 |
| 14陕麦509 (Shaanmai 509) | 4 | 3+ | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 3+ | 4 | 3 | 3+ | x | 3 | 2 |
| 15贵农16 (Guinong 16) | 4 | 2+ | 1 | 1 | 4 | 2 | 1 | 3+ | 1 | 1 | 4 | 2+ | 3 | 1 | 1 | 2 | 1 | 2+ |
| 16鄂麦596 (Emai 596) | 4 | 3 | 3+ | 3+ | 4 | 4 | 3+ | 4 | 4 | 3+ | 3 | 3 | 3+ | 2+ | 3 | 4 | 2+ | 2+ |
| 17鄂麦14 (Emai 14) | 4 | 3+ | 3+ | 1 | 4 | 4 | 1 | 1 | 1 | 4 | 4 | 3 | 2+ | 1 | 1 | 4 | 1 | 2 |
| 18陕农7859 (Shaannong 7859) | 4 | 3+ | 4 | 4 | 4 | 4 | 3+ | 3 | 4 | 4 | 4 | 3 | 2+ | 3 | 3 | 2+ | 3 | 2+ |
| 19小偃6号(Xiaoyan 6) | 4 | 4 | 3+ | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 3+ | 3+ | 3 | 3 | 4 | 3 | 2+ |
| 20陕89150 (Shaan 89150) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 | 4 | 3 | 3 | 3+ | 3 | 4 | 3 | 3 |
| 21鄂恩5号 (Een 5) | 4 | 4 | 4 | 1 | 4 | 4 | 1 | 2 | x | 3+ | 4 | 3 | 2+ | 2+ | 2 | 2 | 2 | 2+ |
| 22鄂麦21 (Emai 21) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 4 | 3+ | 3 | 3+ | 3 | 3+ |
| 23陕354 (Shaan 354) | 2 | 2 | 1 | 1 | 3 | 1 | ; | 3 | 1 | 2 | 4 | 1 | 3+ | 1 | 3 | 2 | 3 | 2+ |
| 24鄂麦11 (Emai 11) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 3 | 3 | 3 | 4 |
| 25陕715 (Shaan 715) | 4 | 3+ | 4 | 4 | 4 | 4 | 3+ | 4 | 4 | 3 | 4 | 4 | 3+ | 3 | 3+ | 2+ | 3 | 2+ |
| 26陕旱8675 (Shaanhan 8675) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3+ | 3+ | 3 | 3 | 3 | 3+ | 3 | 3+ |
| 27陕优225 (Shaanyou 225) | 4 | 3 | 4 | 3+ | 4 | 3 | 4 | 4 | 2+ | 3 | 3 | 3+ | 3 | 3 | 3 | 3+ | 3 | 2+ |
| 28长武134 (Changwu 134) | 4 | 3+ | 4 | 3 | 4 | 3 | 4 | 4 | 4 | 3+ | 3 | 3+ | 3+ | 3+ | 3 | 4 | 3 | 2+ |
| 29鄂恩1号 (Een 1) | 4 | 4 | 4 | 3+ | 4 | 3 | 4 | 4 | 4 | 3 | 3 | 2+ | 3 | 3 | 3 | 2 | 3 | 2 |
| 30西农979 (Xinong 979) | 4 | 4 | 3+ | x | 4 | 3+ | 2+ | 1 | 2+ | 2 | 2+ | 2+ | 2+ | 2+ | 2+ | 3 | 2 | 2+ |
| 31郑州5389 (Zhengzhou 5389) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
新窗口打开
2.2 田间成株抗病性鉴定
2014—2015和2015—2016年度连续2年在河北保定和河南周口2个试验点对这些材料进行了成株抗叶锈鉴定,方差分析结果表明各品种间和地点间差异均极显著,年份间差异显著,品种与地点间和品种与年份间差异均极显著,而品种与重复间和重复间均不显著,这表明小麦叶锈病抗性的表达受基因型和环境互作共同影响(表4)。此外,根据2年2点田间鉴定,慢锈对照SAAR均表现明显的慢锈性(表5)。经IBM SPSS Statistics 19.0软件分析发现在30个小麦生产品种共有18个品种的FDS与慢锈对照的FDS差异不显著,且其侵染型在苗期对混合小种均表现感病,因此这些材料均表现为慢锈性(表5)。Table 4
表 4
表 430个小麦品种及慢锈SAAR和感病对照在2014—2015和2015—2016年分别在保定与周口最终病害严重度的方差分析
Table 4Analysis of variance of relative area final disease severity (FDS) in 30 wheat cultivars including slow rusting cultivar SAAR and susceptible checks tested in the 2014-2015 and 2015-2016 growing seasons
| 变异来源Source of variation | 平方SS | 自由度df | 方差MS | F值F value | P值P value |
|---|---|---|---|---|---|
| 品种 Cultivar | 63178.500 | 31 | 2038.016 | 18.823** | <0.001 |
| 地点 Location | 11489.160 | 1 | 11489.160 | 106.115** | <0.001 |
| 年份 Season | 475.785 | 1 | 475.785 | 4.394* | 0.038 |
| 重复 Replicate | 4.000 | 1 | 4.000 | 0.037 | 0.848 |
| 品种×地点 Cultivar×Location | 17750.527 | 31 | 572.598 | 5.289** | <0.001 |
| 品种×年份 Cultivar×Season | 11587.777 | 31 | 373.799 | 3.452** | <0.001 |
| 品种×重复 Cultivar×Replicate | 922.062 | 31 | 29.744 | 0.275 | 1.000 |
| 误差 Error | 13858.625 | 128 | 108.271 | ||
| 总计 Total | 293364.000 | 256 | |||
| 校正总计 Corrected total | 119266.438 | 255 |
新窗口打开
2.3 分子检测
30个小麦品种进一步利用11个分子标记进行检测(表2),共检测到Lr1、Lr26和Lr46的特异目的片段(图1、图2),而未检测到Lr9、Lr10、Lr19、Lr20、Lr24和Lr34相应的特异条带(图1)。鄂麦14、陕229、西农979和鄂恩5号等品种中检测出Lr1,鄂恩1号、鄂恩5号、鄂恩6号、陕合6号、贵农16、陕225、陕354、陕715、陕麦509和陕农7859等品种中检测到与Lr26相同的带型,陕225和小偃81两个品种检测出与Lr46基因相同的带型,其余的品种均未检测出Lr46(图2)。分子标记检测结果见表1,另外结果显示分子标记与基因推导鉴定结果表现一致。Table 5
表5
表532个小麦品种、慢锈品种及对照品种苗期对混合小种的反应型和成株期在2014—2015年和2015—2016年的最终病害严重度
Table 5Infection types (IT) in the seedling test with P. triticina pathotype mixed species and mean final disease severity (FDS) in the field experiments with the same pathotype in the 2014-2015 and 2015-2016 growing seasons for 32 wheat genotypes with slow rusting resistance to leaf rust
| 编号Code | 品种名 Cultivar | 苗期对混合小种抗性 Seedling IT to mix species | 2014—2015 保定严重度 FDS of Baoding (%) | 2015—2016 保定严重度 FDS of Baoding (%) | 2014—2015 周口严重度 FDS of Zhoukou (%) | 2015—2016 周口严重度 FDS of Zhoukou (%) | 严重度平均值 Average FDS (%) |
|---|---|---|---|---|---|---|---|
| 1 | SAARa | 4 | 3.00 | 5.00 | 1.00 | 1.00 | 2.50 |
| 2 | 陕354d Shaan 354d | 4 | 6.25 | 17.50 | 12.50 | 15.00 | 12.81 |
| 3 | 鄂麦15d Emai 15d | 4 | 3.75 | 20.00 | 8.75 | 20.00 | 13.13 |
| 4 | 鄂恩5号d Een 5d | 4 | 3.00 | 25.00 | 13.75 | 16.25 | 14.50 |
| 5 | 陕麦94d Shaanmai 94d | 4 | 10.00 | 27.50 | 12.50 | 8.75 | 14.69 |
| 6 | 陕225d Shaan 225d | 4 | 12.50 | 15.00 | 15.00 | 17.50 | 15.00 |
| 7 | 襄麦55d Xiangmai 55d | 4 | 20.00 | 18.75 | 11.25 | 12.50 | 15.63 |
| 8 | 鄂恩6号d Een 6d | 4 | 17.50 | 16.25 | 15.00 | 15.00 | 15.94 |
| 9 | 陕麦509d Shaanmai 509d | 4 | 15.00 | 16.25 | 22.50 | 16.25 | 17.50 |
| 10 | 陕715d Shaan 715d | 4 | 8.75 | 37.50 | 12.50 | 12.50 | 17.81 |
| 11 | 陕优225d Shaanyou 225d | 4 | 22.50 | 16.25 | 16.25 | 17.50 | 18.13 |
| 12 | 鄂麦352d Emai 352d | 4 | 16.25 | 27.50 | 12.50 | 17.50 | 18.44 |
| 13 | 鄂麦11d Emai 11d | 4 | 23.75 | 18.75 | 22.50 | 10.00 | 18.75 |
| 14 | 陕229d Shaan 229d | 4 | 11.25 | 28.75 | 15.00 | 22.50 | 19.38 |
| 15 | 西农979d Xinong 979d | 4 | 10.00 | 40.00 | 12.50 | 15.00 | 19.38 |
| 16 | 贵农16d Guinong 16d | 4 | 10.00 | 13.75 | 22.50 | 32.50 | 19.69 |
| 17 | 鄂麦27d Emai 27d | 4 | 11.25 | 25.00 | 20.00 | 25.00 | 20.31 |
| 18 | 西农881d Xinong 881d | 4 | 22.50 | 27.50 | 12.50 | 20.00 | 20.63 |
| 19 | 鄂麦596d Emai 596d | 4 | 15.00 | 28.75 | 18.75 | 25.00 | 21.88 |
| 20 | 鄂麦14 Emai 14 | 4 | 15.00 | 57.50 | 16.25 | 5.00 | 23.44 |
| 21 | 陕89150 Shaan 89150 | 4 | 20.00 | 40.00 | 20.00 | 15.00 | 23.75 |
| 22 | 鄂麦21 Emai 21 | 4 | 12.50 | 50.00 | 22.50 | 17.50 | 25.63 |
| 23 | 西农1376 Xinong 1376 | 4 | 35.00 | 47.50 | 8.75 | 13.75 | 26.25 |
| 24 | 鄂麦23 Emai 23 | 4 | 7.50 | 57.50 | 25.00 | 16.25 | 26.56 |
| 25 | 小偃6号 Xiaoyan 6 | 3 | 10.00 | 80.00 | 17.50 | 18.75 | 31.56 |
| 26 | 鄂恩1号 Een 1 | 4 | 55.00 | 40.00 | 17.50 | 30.00 | 35.63 |
| 27 | 陕农7859 Shaannong 7859 | 4 | 13.75 | 40.00 | 22.50 | 80.00 | 39.06 |
| 28 | 陕合6号 Shaanhe 6 | 3 | 6.25 | 70.00 | 3.00 | 90.00 | 42.31 |
| 29 | 长武134 Changwu 134 | 4 | 20.00 | 60.00 | 52.50 | 77.50 | 52.50 |
| 30 | 陕旱8675 Shaanhan 8675 | 4 | 27.50 | 60.00 | 67.50 | 60.00 | 53.75 |
| 31 | 小偃81 Xiaoyan 81 | 4 | 17.50 | 80.00 | 50.00 | 80.00 | 56.88 |
| 32 | 郑州5389b Zhengzhou 5389b | 4 | 75.00 | 90.00 | 80.00 | 75.00 | 80.00 |
| LSD | 20.60 | ||||||
新窗口打开
3 讨论
本研究结合基因推导和标记检测,共鉴定出Lr1、Lr13、Lr26和Lr46等少数几个抗病基因,说明中国小麦材料中抗病基因单一,而且缺乏有效的抗病基因。目前只有少数几个抗病基因如Lr9、Lr19、Lr24、Lr28、Lr47、Lr51和Lr53等对所有供试小种表现高抗,但这些基因在生产品种中尚未广泛应用,因此今后应进一步加强抗病育种工作,在生产品种中转化更多的有效抗病基因来抑制小麦叶锈病流行。本研究中陕225和小偃81经标记检测携带有Lr46,但其田间严重度分别为15%和57%,表现中度抗病及高度感病,表明Lr46单独存在时,效应比较微弱,需聚合更多的微效基因,才能达到高水平的成株抗病性。鄂恩1号由(洛夫林10/761)F1与苏麦3号杂交选育而来,亲本洛夫林10含有抗病基因Lr26[33],说明鄂麦1号中携带的Lr26可能来自于洛夫林10;鄂麦14由繁6/偃大72-629杂交选育而成,其亲本繁6的系谱为I413B01828/NP824/3/五一麦/成都光头分枝麦/中农483/4/中农2813分枝/阿夫,其中阿夫含有Lr1[34],说明鄂麦14中的抗病基因Lr1可能来自于阿夫;西农979的亲本西农2611是以陕229作母本、(84<14>43×83<2>3)×(西农65×小偃6号)作父本杂交选育而成,虽然陕229中的Lr1无法经系谱推出,但很明显西农979的Lr1来自于陕229。庄巧生[35]在2003年指出洛夫林、毕加索、山前草和阿芙乐尔等骨干品种均含有抗叶锈病基因Lr26,本研究中携带Lr26的4个品种陕农7859、陕麦509、陕354和陕715,其亲本均有携带Lr26的骨干亲本。经基因推导可知西农979、陕229和贵农16可能携带有Lr13,此3个品种在田间严重度为19%,表现为慢锈品种;Lr13的载体品系RL4031在田间的严重度为5%,西农979、陕229和贵农16是否携带Lr13有待进一步试验验证。本研究中还鉴定到18个表现成株慢锈的品种,这些材料中可能携带未知的成株慢锈基因,今后应利用遗传分析结合分子标记明确其成株抗性的抗病机理,同时这些材料也可将来用于持久抗性品种的选育。
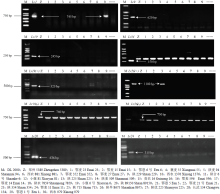 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1用Lr1、Lr9、Lr10、Lr19、Lr20、Lr24、Lr26和Lr34标记检测30个小麦品种的部分电泳结果
-->Fig. 1Part of PCR amplification of 30 Chinese wheat cultivars with the markers Lr1, Lr9, Lr10, Lr19, Lr20, Lr24, Lr26, and Lr34
-->
基因推导及标记检测各有优缺点:分子标记快速、准确且不受环境条件的限制,但易扩增出非特异性带型,出现假阳性,影响结果检测的准确性;基因推导法周期短、不受生长季节限制、在短期内可对大量品种进行分析且结果准确,但易受小种鉴别力不足或遗传背景等因素影响,使鉴定结果受到限制。在本研究中所选的分子标记特异性强,能准确地检测出抗病基因,而且苗期基因推导与标记检测结果一致,更加验证了结果的可靠性。部分材料由于含有未知抗病基因没有推导出已知抗叶锈病基因,用其与感病品种配置杂交组合,进一步利用遗传分析和分子定位对未知基因进行鉴定有望发现一些新的抗病基因,目前本课题组中共发现了LrZH84[17]、LrG98[36]、LrXi[37]、LrBi16[38]、LrNJ97[4]、LrFun[39]、LrZH22[40]等7个新的抗叶锈病基因,都是通过此方法进行鉴定和定位的。另外,本研究采用苗期基因推导和成株抗性鉴定相结合,苗期鉴定能够检测出苗期主效抗病基因,而不能检测出成株微效抗病基因,而田间通过接种强毒性生理小种进行鉴定,能够检测出成株慢锈基因。在本研究中筛选出18个品种可能携带成株慢锈抗病基因,这些材料可应用于小麦生产培育持久抗病品种来防治小麦叶锈病。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2用cslv46-G22标记检测30个小麦品种的电泳结果
-->Fig. 2The PCR amplification of 30 Chinese wheat cultivars with the CAPS marker cslv46-G22
-->
4 结论
根据苗期基因推导和分子标记检测结果,在30个小麦品种(系)中有14个品种(系)中可能携带有4个已知抗叶锈病基因,即Lr1、Lr13、Lr26和Lr46。检测到10个品种(系)含有Lr26,其中鄂恩5号携带Lr26和Lr1,贵农16携带Lr26和Lr13,另外Lr26和Lr46共同存在于陕225中,检测到含有Lr1的共有4个品种(系),除了上述鄂恩5号外,Lr1和Lr13共同存在陕229和西农979中,品种鄂麦14仅含有Lr1;含有Lr46的有2个品种(系),除了陕225外,小偃81仅携带Lr46。通过田间抗性鉴定共检测出18个品种(系)可能携带成株慢锈基因。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}