0 引言
【研究意义】玉米象(Sitophilus zeamais)是一种世界性的储粮害虫[1],也是较难防治的仓储害虫之一[2]。目前防治玉米象主要使用化学合成熏蒸剂,如溴甲烷、硫酰氟以及磷化铝等。2015年,溴甲烷在全球范围内已被禁用,磷化氢也因长期使用引起严重的抗药性问题[3-4],因此寻找新型熏蒸剂已成为当务之急。异硫氰酸酯类化合物因其对储粮害虫具有极强的熏蒸活性已成为研究开发的热点,但其作用机理研究较为匮乏。针对辣根素(烯丙基异硫氰酸酯,简称AITC)可能的作用靶标玉米象细胞色素C氧化酶亚基Ⅲ基因进行克隆、序列分析以及原核表达,获得该化合物作用于玉米象致死的直接证据,可为异硫氰酸酯类生物熏蒸剂作用机制和作用靶标的研究打下基础,为后续新农药分子的生物合理设计提供思路。【前人研究进展】线粒体DNA(mtDNA)是动物界细胞核外唯一具有遗传密码和蛋白质翻译系统的遗传物质,除了参与能量转换外,还参与4种呼吸酶复合物(13种多肽)的形成[5-6],其中细胞色素C氧化酶(Cco)是真核生物线粒体内膜和需氧菌细胞膜电子传递链上的终端金属膜蛋白酶[7-8],催化氧分子还原为水,并与质子泵功能相偶联,具有不同的金属活性中心。同时承担着细胞色素C到氧分子的电子传递功能,是唯一能将电子传递给氧分子的细胞色素,是线粒体氧化能力的关键调节部位[9]。所有真核细胞的COX由13个亚基组成,其中构成级联反应核心的最大3个亚基(COXI、Ⅱ和Ⅲ)由mtDNA编码,为COX的核心亚基[10]。其余10个亚基(COⅣ、Ⅴa、Ⅴb、Ⅵa、Ⅵb、Ⅵc、Ⅶa、Ⅶb、Ⅶc和Ⅷ)均由细胞核DNA编码。体外实验动力学研究表明磷化氢是牛心细胞色素氧化酶的非竞争性抑制剂,磷化氢对细胞色素氧化酶的抑制也一直被认为是昆虫致死的主要原因[11-12]。尽管磷化氢的作用机理仍然有很多争议,但是线粒体细胞色素C氧化酶被认为是一个主要作用靶标[13]。玉米象成虫对辣根素与磷化氢有类似的中毒症状,其线粒体均有明显的破坏[14];蛋白组学研究发现,ITCs能够与线粒体复合物Ⅰ和Ⅳ结合,活性氧簇(reactive oxygen species,ROS)的产生是一个重要的下游效应[15]。【本研究切入点】辣根素在活体和离体下均对复合体Ⅳ有抑制作用,产生的ROS能够进一步破坏线粒体膜电位,导致线粒体功能缺陷[16-17];目前关于COX亚基的研究主要集中在脊椎动物和细菌上的亚基Ⅰ、亚基Ⅱ上[18-19],关于无脊椎动物玉米象细胞色素C氧化酶亚基Ⅲ进行原核表达和更进一步的研究尚未见报道。【拟解决的关键问题】利用RT-PCR和RACE技术扩增克隆玉米象COXⅢ序列全长;生物信息学软件对该基因及其编码蛋白的结构、性质与功能进行预测;利用不同融合表达策略进行原核表达,为进一步研究其功能及明确异硫氰酸酯类化合物的作用位点提供依据。1 材料与方法
试验于2015年7月至2016年6月在西北农林科技大学无公害农药研究服务中心完成。1.1 供试昆虫
玉米象成虫由西北农林科技大学无公害农药研究服务中心室内人工饲养。小麦置于烘箱(80℃)中消毒2 h,调整含水量至约14%,分装入罐头瓶内,接入玉米象成虫,置于(25±1)℃,相对湿度75%±5%,光周期12L﹕12D的养虫室内连续饲养7日后筛出成虫,挑取个头大小一致试虫备用。1.2 主要试剂及工具酶
Primer Script Reverse Transcriptase、RNaseH、胶回收试剂盒、质粒提取试剂盒购自北京天根生化科技有限公司;pMD19-T载体、DL2000 Marker、6×Loading Buffer、限制性内切酶EcoR I、Xho I购自宝生物工程(大连)有限公司;DAB显色试剂盒、T4 DNA连接酶、蛋白质分子量Marker购自北京索莱宝科技有限公司;大肠杆菌BL21(DE3)和Transetta(DE3),pEASY-Blunt E1购自全式金公司;原核表达载体pET28a、pET30a均为笔者实验室保存,引物由生工生物工程(上海)股份有限公司合成;测序由英潍捷基(上海)贸易有限公司完成。其他试剂均为国产或进口分析纯试剂。1.3 总RNA的提取
用改进后的CTAB法[20]提取玉米象成虫总RNA。(1)5 mL离心管中加入4 mL CTAB buffer和200 μL β-巯基乙醇于65℃预热;(2)液氮将成虫玉米象研磨成粉末,迅速将样品转移至预热好的装有CTAB buffer离心管中,65℃孵育10 min,每2 min涡旋一次;(3)取1 mL分装到2 mL离心管中,加等体积氯仿-异戊醇混合溶液(氯仿﹕异戊醇=24﹕1),涡旋混匀,冰浴10 min,12 000 r/min、4℃离心15 min;(4)吸取上清液加入新的2 mL离心管中,加等体积氯仿-异戊醇混合溶液,冰浴10 min,12 000 r/min、4℃离心10 min;(5)取上清600 μL,加入200 μL的8 mol·L-1 LiCl,置于-80℃冰箱沉淀RNA 1 h,12 000 r/min、4℃离心30 min;(6)弃上清,加1 mL 75%乙醇洗涤,7 500 r/min、4℃离心5 min;(7)弃上清,40 μL DEPC-water溶解,-80℃保存备用。酶标仪检测RNA溶液的纯度和浓度;1%的琼脂糖凝胶电泳检测提取玉米象总RNA的完整性。
1.4 玉米象COXⅢ全长基因的获得与验证
1.4.1 玉米象COXⅢ基因cDNA片段的克隆 根据反转录试剂盒HiFi Script Quick gDNA Removal cDNA Kit操作说明,以总RNA为模板反转录合成cDNA第一链。根据NCBI中登记的玉米象COXⅢ cDNA的部分已知序列,利用Primer Premier 5(PP5)软件设计引物,以上述第一链cDNA为模板,以cox3 1F、cox3 1R为引物(表1)进行PCR扩增。PCR反应体系:cDNA 1.0 µL,引物cox3 1F和cox3 1R各1.0 µL,2×Taq Master Mix 12.5 µL,灭菌ddH2O补足至25 µL。PCR程序:94℃预变性3 min;94℃ 30 s,52℃ 30 s,72℃ 1 min,共32个循环;72℃延伸5 min。
Table 1
表1
表1玉米象COXⅢ基因克隆所用引物
Table 1Primers used for cloning of S. zeamais COXⅢ
| 引物Primer | 序列Sequence (5′-3′) | 退火温度Annealing temperature (℃) |
|---|---|---|
| cox3 1F | TATTACCAAGCCCCATTCTCTATTAGA | 52 |
| cox3 1R | GGTTGAAAAATGATTGAAGTAAAGTCG | |
| qc COX3 1F | ATGTTAAATAATAAAAAAAATCACCCCT | 55 |
| qc COX3 1R | TTATCTTCCTCATCAGTAAATAGAAAGA | |
| gene cox3 1F | ATGTTAAATAATAAAAAAAATCACCCCTT | 55 |
| gene cox3 1R | TTAACTTCCCCACCAGTAAATAGAAAGA | |
| COX3 E F1 | GGAATTCATGTTAAATAATAAAAAAAATCACCCC | 55 |
| COX3 X R1 | CCGCTCGAGTTAACTTCCCCACCAGTAAATA | |
| T7 promoter primer | TAATACGACTCACTATAGGG | 55 |
| T7 Terminator Primer | GCTAGTTATTGCTCAGCGG | |
| M13F(-47) | CGCCAGGGTTTTCCCAGTCACGAC | 55 |
| M13R(-48) | AGCGGATAACAATTTCACACAGGA | |
| UPM-long | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| UPM-short | CTAATACGACTCACTATAGGGC | |
| NUP | AAGCAGTGGTATCAACGCAGAGT |
新窗口打开
PCR产物经1%的琼脂糖凝胶电泳检测,经切胶回收后,连接于pMD-19T克隆载体,并转化感受态DH5α中,蓝白斑筛选后挑取白色菌落进行菌落PCR,所用引物为M13F(-47)、M13R(-48),鉴定正确后,挑取阳性菌落于5 mL LB液体培养基(含氨苄),在200 r/min,37℃培养12 h条件下,提取质粒,送英潍捷基(上海)贸易有限公司测序。
1.4.2 细胞色素C氧化酶亚基Ⅲ 3′-RACE扩增 取3.75 µL总RNA为模板,以接头引物1 µL 3′-CDS PrimerA为反转录引物,72℃孵育3 min,42℃ 2 min,离心收集,之后按照SMARTerTM RACE cDNA Amplification Kit User Manual试剂盒说明书,合成3′-RACE的第一链cDNA。根据NCBI中亚基Ⅲ部分已知序列,利用软件Primer Primier 5.0设计3′-RACE扩增特异性引物cox3 1F,反应体系:2.5 μL 3′-RACE cDNA,5 μL下游引物UPM,1 μL上游引物cox3 1F,1 μL 50×Advantage 2 Polymerase Mix,1 μL dNTP Mix,1 μL10×Advantage 2 PCR Buffer,灭菌ddH2O补足至50 µL。PCR程序:94℃预变性3 min;94℃ 30 s,52℃ 45 s,72℃ 1 min,共32个循环;72℃延伸5 min。PCR产物的检测、纯化及测序方法同1.4.1。
1.4.3 细胞色素C氧化酶亚基Ⅲ 5′-RACE扩增 取2.75 µL总RNA为模板,以接头引物1.0 µL 5′-CDS PrimerA为反转录引物,72℃孵育3 min,42℃ 2 min,离心收集到管底,加入1 µL SMARTerⅡAoligo,之后按照SMARTerTM RACE cDNA Amplification Kit User Manual试剂盒说明书,合成5′-RACE的第一链cDNA。利用软件PP5设计5′-RACE扩增特异性引物cox3 1R(表1),反应体系:2.5 μL 5′-RACE cDNA,5 μL上游引物UPM,1 μL下游引物cox3 1R,1 μL 50×Advantage 2 Polymerase Mix,1 μL dNTP Mix,1 μL10×Advantage 2 PCR Buffer,灭菌ddH2O补足至50 µL。PCR程序:94℃预变性3 min;94℃ 30 s,55℃ 45 s,72℃ 1 min,共32个循环;72℃延伸5 min。PCR 产物的检测、纯化及测序方法同1.4.1。
1.4.4 细胞色素C氧化酶亚基Ⅲ全长验证 利用ContigExpress拼接测序所得的3′-RACE扩增片段和5′-RACE扩增片段以及中间片段;利用在线网站ORF Finder查找开放阅读框(ORF);利用PP5在其起始密码子近5′端和终止密码子的近3′端设计特异性上游引物qc COX3 1F和特异性下游引物qc COX3 1R(表1),反应体系:1 µL cDNA模板,上下游引物各1 µL,12.5 µL的2×Taq Mix,灭菌ddH2O补足至25 µL,PCR程序:94℃预变性3 min;94℃ 30 s,55℃ 30 s,72℃ 1 min,共28个循环;72℃延伸5 min。PCR 产物的检测、纯化及测序方法同1.4.1。
1.5 玉米象COXⅢ基因核苷酸序列及其编码氨基酸序列生物信息学分析
将克隆得到的玉米象COXⅢ序列在NCBI数据库中Blast验证,使用ContigExpress软件拼接得玉米象COXⅢ cDNA 全长序列,利用ORF finder在线软件查找其开放阅读框(ORF);使用DNAMAN对玉米象COXⅢ序列及编码蛋白进行生物信息学分析,并将其与赤拟谷盗(Tribolium castaneum)、黑盾胡蜂(Vespa bicolor)、黑腹果蝇(Drosophila melanogaster)、红棕象甲(Rhynchophorus ferrugineus)、家蚕(Bombyx mori)、米象(Sitophilus oryzae)6种昆虫的COXⅢ编码氨基酸序列进行多重比对。利用MEGA5.1软件,采取Neighbor-Joining法构建系统进化树;使用在线软件ExPASy-ProtParam tool(http://web.expasy.org/ protparam/)预测玉米象COXⅢ编码氨基酸的基本理化性质;利用SignalP(http://www.cbs.dtu.dk/services/ SignalP/)在线工具对玉米象COXⅢ基因编码蛋白N端信号锚定序列进行预测;TMHMM Server v.1.0(http://www.cbs.dtu.dk/services/TMHMM/)在线工具对玉米象COXⅢ基因编码蛋白跨膜区进行预测,蛋白质三级结构预测采用SWISS-MODEL。1.6 玉米象COXⅢ原核表达载体的构建与鉴定
1.6.1 原核表达载体pET系列构建与鉴定 根据玉米象COXⅢ ORF及原核表达载体多克隆位点序列设计含酶切位点的一对引物:COX3 E F1、COX3 E F1(表1)。以基因合成后的片段为模板,PCR扩增,引物为gene cox3 1F、gene cox3 1R(表1),扩增产物经切胶回收、纯化后连接到克隆载体pMD19-T。EcoRI和XhoI分别双酶切重组质粒pMD-19T-COXⅢ和表达载体pET28a、pET30a、pET32a、pET42a,16℃过夜连接。连接产物转化克隆感受态DH5α,菌落PCR鉴定,引物为T7 promoter primer、T7 Terminator Primer(表1),挑阳性克隆提质粒,再次双酶切鉴定、测序,获得玉米象的重组表达载体,分别命名为pET28a-COXⅢ、pET30a-COXⅢ、pET32a-COXⅢ、pET42a-COXⅢ。1.6.2 原核表达载体PEASY Blunt E1构建 试验中,针对目的片段COXⅢ分别进行了5个不同载体的构建:pEASY-Blunt E1、pET-28a、pET-30a、pET-32a、pET-42a。pEASY-Blunt E1和pET系列表达载体不同之处在于:pEASY-Blunt E1不含酶切位点,为平滑末端;pET系列表达载体含多克隆酶切位点。对pEASY-Blunt E1原核表达载体则设计不含相应酶切位点引物,无需酶切鉴定,其余方法参照pET系列载体构建过程,命名为pEASY-Blunt E1-COXⅢ。
1.7 原核表达
鉴定正确的上述重组质粒分别转化至表达感受态BL21(DE3),挑取含有PEASY Blunt E1-COXⅢ、pET32a-COXⅢ的BL21(DE3)阳性单克隆菌分别于5 mL LB液体培养基(AMP 100 mg·mL-1);挑取含有pET28a-COXⅢ、pET30a-COXⅢ、pET42a- COXⅢ的BL21(DE3)单克隆接种到5 mL LB液体培养基中(Kan 50 mg·mL-1),37℃振荡培养过夜。取适量菌液送生工测序,鉴定目的片段连接是否正确。根据测序结果,取连接正确的单克隆菌菌液以1﹕100(V/V)的比例接种于含有相应抗生素的LB液体培养基中,摇床200 r/min,37℃培养。待OD600为0.6—0.8时,不同温度、IPTG浓度、时间诱导表达。1.8 融合蛋白的可溶性及Western blot检测
诱导结束后,将菌液于4℃ 10 000 r/min离心10min,收集菌体,称重,按照1﹕10的比例加入25 mmol·L-1 Tris-HCl缓冲液(pH 8.0)进行重悬浮,之后再冰上进行超声破碎,功率300 W,工作8 s,间隔9 s,超声20 min,超声后在离心机4℃ 12 000 r/min离心30 min中,分别收集上清和沉淀,再加入4×聚丙烯酰胺凝胶电泳(SDS-PAGE)上样缓冲液,水浴煮沸10 min,12 000 r/min离心5 min。取上清用12%的SDS-PAGE凝胶电泳分离蛋白,并用考马斯亮蓝染色检测蛋白的表达情况。Western blot印迹杂交验证目的蛋白的表达。
2 结果
2.1 玉米象COXⅢ cDNA克隆及序列分析
克隆获得一个玉米象COXⅢ的cDNA全长序列,ORF Finder分析发现该基因含有一个为792 bp,编码263个氨基酸,起始密码子为ATG,终止密码子为TAA(图1)。在NCBI数据库Blastn发现该序列与米象COXⅢ序列的最高一致性为99%,与其他昆虫的最高一致性也在75%以上;进一步用ORF翻译所得的氨基酸序列进行Blastp发现其与其他昆虫的COXⅢ编码氨基酸的一致性在70%以上。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1玉米象COXⅢ ORF及其编码氨基酸序列
-->Fig. 1The ORF sequence and amino acid sequence of COXⅢ from S. zeamais
-->
2.2 玉米象COXⅢ编码氨基酸序列比对及分子进化关系分析
应用DANMAN 8.0生物学软件对玉米象COXⅢ氨基酸序列与其他6种昆虫的氨基酸序列进行多重序列比对(图2)。分析表明玉米象COXⅢ与所比对的昆虫高度同源,同源性达75.23%,其结果与COXⅢ是线粒体其中一个高度保守的核心亚基报道相一致[10]。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2玉米象与6种其他昆虫COXⅢ编码氨基酸序列多重比对
-->Fig.2Multiple alignment of the COXⅢ amino acid sequences of S. zeamais and six other insects
-->
根据玉米象COXⅢ在NCBI中blastp比对结果,采用MEGA5.0对同源性较高的昆虫构建玉米象COXⅢ的系统发育树[21-22](图3),系统发育树显示玉米象与鞘翅目象鼻虫科昆虫米象进化关系最近,相似性为100%;昆虫COXⅢ编码氨基酸以目单位各聚为支,其中双翅目的黑腹果蝇和鳞翅目的家蚕与鞘翅目的进化关系最近,膜翅目的黑盾胡蜂与玉米象COXⅢ氨基酸序列同源性不大,与昆虫的进化关系相一致。
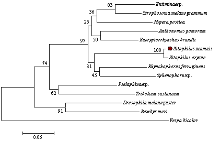 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3玉米象及其他昆虫COXⅢ氨基酸序列的分子系统进化树
-->Fig. 3Phylogenetic tree of COXⅢ from S. zeamais and other insects
-->
2.3 玉米象COXⅢ编码蛋白的基本理化性质预测
利用在线软件ExPASy-ProtParam tool(http://web. expasy.org/protparam/)分析玉米象COXⅢ基因编码蛋白的基本理化性质,发现该蛋白共包含263个氨基酸,预测分子式为C1492H2154N338O362S10,分子量为30 938.1 Da,理论等电点(pI)为6.51,半衰期为30 h,脂肪指数为106.01,负电荷残基(Asp+Glu)总电荷为13,正电荷残基(Arg+Lys)总电荷为10,该蛋白的亲水性为0.492,推测为疏水性蛋白。预测蛋白的不稳定参数为34.49,通常认为不稳定参数小于40的蛋白是稳定蛋白,所以此蛋白为稳定蛋白。利用SignalP(http://www.cbs.dtu.dk/services/SignalP/)在线工具对玉米象COXⅢ基因编码蛋白N端信号锚定序列进行预测,发现该蛋白不存在信号肽切割位点(图4),说明玉米象COXⅢ编码蛋白为膜结合蛋白。利用TMHMM Server v. 1.0(http://www.cbs. dtu.dk/services/ TMHMM/)在线工具对玉米象COXⅢ基因编码蛋白跨膜区进行预测,发现7个跨膜区(图5)。利用在线3D模型构建工具Automatic Modelling Mode(http://swiss model.eapasy.org/)构建三级结构(图6),三维结构模型显示玉米象COXⅢ有7个α-螺旋,与图6中7个跨膜区相对应。
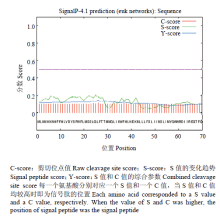 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4玉米象COXⅢ基因编码蛋白信号肽预测图
-->Fig. 4The predicted signal amino acids of COXⅢ from adult S. zeamais
-->
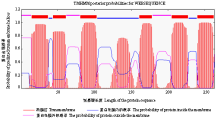 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5玉米象COXⅢ基因编码蛋白跨膜预测
-->Fig.5The prediction of transmembrane helices in COXⅢ of adult S. zeamais
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6玉米象COXⅢ蛋白的3-D结构预测模型
-->Fig. 6The predicted three-dimensional structure of COXⅢ of adult S. zeamais
-->
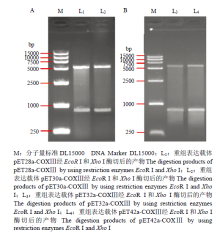 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7重组表达载体pET系列鉴定
-->Fig. 7Identification of pET expression vectors
-->
2.4 原核表达载体的构建和重组子鉴定
生物界中存在不同的编码系统,线粒体基因编码与核基因编码是两个不同的编码系统。通过生物信息学软件分析,按照密码子的简并性原则,对COXⅢ基因相应位置碱基置换,基因合成目的片段,构建表达载体,进行原核表达。利用EcoR I和Xho I对重组载体pET系列分别进行双酶切鉴定,酶切产物用1%的琼脂糖凝胶检测,图7为pET系列4个表达载体双酶切电泳图。此外重组子的测序结果与目的序列完全一致,表明玉米象的原核表达载体构建成功。同样,对重组子pEASY-Blunt E1-COXⅢ进行菌落PCR和测序鉴定,测序正确,无碱基的缺失和突变,表明玉米象COXⅢ原核表达载体pEASY-Blunt E1-COXⅢ构建成功。
2.5 融合蛋白的可溶性及Western blot检测
分别对玉米象COXⅢ基因构建的5个不同的表达载体进行融合表达。SDS-PAGE染色、脱色后,E. coli BL21(DE3)中的4个表达载体pEASY-Blunt E1-COXⅢ、pET28a-COXⅢ、pET30a-COXⅢ、pET32a-COXⅢ未能在预测分子量33、34、36、51 kD条带位置处观察到融合蛋白条带;进一步通过WB印记杂交验证、DAB显色,也未在上述蛋白分子量位置处显色。从上述SDS-PAGE和Western blot结果可以看出,pEASY- Blunt E1-COXⅢ-BL21(DE3)、pET28a-COXⅢ-BL21(DE3)、pET30a-COXⅢ-BL21(DE3)、pET32a-COXⅢ-BL21(DE3)4个表达体系未能成功表达出融合蛋白。图8为4个表达载体中PET30a-COXⅢ重组载体在16℃,转速200 r/min,诱导24 h后的SDS-PAGE电泳图。由图可以看出,重组载体pET30a-COXⅢ诱导后的上清和沉淀,与pET30a空载体诱导相比,并无明显条带差异。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8玉米象pET30-COXⅢ原核表达
-->Fig.8Prokaryotic expression of pET30-COXⅢ from S. zeamais
-->
在16℃,1 mmol·L-1 IPTG诱导24 h条件下,以抗His标签鼠单克隆抗体(HRP标记)为标签抗体。根据Western blot显色结果,5个表达载体中只有pET42a-COXⅢ在E. coli BL21(DE3)中成功表达的融合蛋白,蛋白分子量大小为62 kD,与理论预测值相一致,该蛋白主要以包涵体形式存在(图9)。
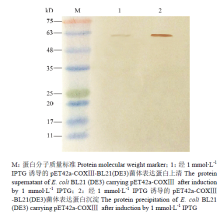 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图9玉米象重组蛋白COXⅢ的Western blot检测
-->Fig. 9Western blot analysis of recombinant COXⅢ
-->
3 讨论
细胞色素C氧化酶是需氧菌细胞膜电子传递链上和真核生物线粒体内膜的终端酶[7-8],同时也是磷化氢[11-12]、威百亩[23]、氢氰酸[24]可能的作用靶标。真核生物细胞COX由13个亚基组成,其中最大3个亚基(COXI、II和III)由mtDNA编码,存在于大多数含heme/copper的终端酶中,所以COXI、II和III通常被称为核心亚基。本试验得到玉米象COXⅢ开放阅读框,利用swiss model在线分析软件预测后,含有7个跨膜螺旋,与其他文献报道的细胞色素C氧化酶亚基Ⅲ含有7个跨螺旋一致[25]。尽管亚基Ⅲ存在于几乎所有的细胞色素氧化酶中,但其具体功能仍不清楚,它既不参与对辅因子的键合又非质子泵功能所必需[26]。有文献报道可能参与细胞色素C氧化酶的装配[27]或形成氧气的通道入口以引导到达双核活性位点[28]。本研究通过RT-PCR以及RACE技术得到细胞色素C氧化酶的开放阅读框为792 bp,编码263个氨基酸,由于线粒体基因组和核基因编码有两套不同的编码系统[29],例如在无脊椎动物线粒体中TGA编码色氨酸,在核基因中TGA不编码氨基酸,为终止子;ATA在无脊椎动物线粒体中编码甲硫氨酸,在核基因中编码异亮氨酸。按照目标序列编码蛋白一级序列的不变性,对获得细胞色素C亚基Ⅲ基因进行碱基置换,修改完成后,基因合成,构建表达载体,进行原核表达。
对玉米象COXⅢ序列同源相似性搜索结果发现,
该序列与其他昆虫COXⅢ氨基酸序列有较高的同源一致性,可能与COXⅢ是线粒体细胞色素C氧化酶中核心亚基有关,该亚基在长期的生物进化过程中,具有高度保守性[10]。所构建的系统发育树也出现相似的结果,在COXⅢ氨基酸整个系统发育过程中,同属鞘翅目的昆虫玉米象与米象进化关系最近,来自不同目的昆虫COXⅢ同源性不一,其中鞘翅目与双翅目、鳞翅目进化关系较近,与鞘翅目的进化关系较远,由此说明,COXⅢ除了具有相对保守性外,还有一定程度的差异性,从而使得它能在不同物种间行使其独特的生物学功能。
前期研究表明,玉米象细胞色素C氧化酶为辣根素主要作用靶标[17]。本研究针对玉米象COXⅢ基因构建5个不同表达载体,分别为pEASY-blunt EI-COXⅢ、pET28a-COXⅢ、pET30a-COXⅢ、pET32a-COXⅢ、PET42a-COXⅢ,经双酶切和测序验证,目的片段成功连接到表达载体并转化到表达菌株BL21(DE3)中,目的片段无碱基缺失或移码突变。对构建好的表达体系pEASY-Blunt E1-COXⅢ-BL21(DE3)、pET28a-COXⅢ-BL21(DE3)、pET30a-COXⅢ-BL21(DE3)、pET32a-COXⅢ-BL21(DE3)、pET42a-COXⅢ-BL21(DE3)分别进行原核表达。在pEASY-blunt EI-COXⅢ-BL21(DE3)诱导中,以试剂盒中750 bp长度的片段为阳性对照,在不同诱导温度、IPTG浓度、时间下,未看到目的片段的表达产物,阳性对照表达蛋白大约为30 kD;以相应的空载体表达菌株为阴性对照,pET28a-COXⅢ-BL21(DE3)、pET30a-COXⅢ-BL21(DE3)、pET32a-COXⅢ-BL21(DE3)、pET42a-COXⅢ-BL21(DE3),以上述的条件进行表达,只有pET42a-COXⅢ-BL21(DE3)成功表达出带有6×His标签和GST tag标签的融合蛋白,经Western blot印记验证后在相应的条带位置显色,融合蛋白主要以包涵体的形式存在;外源序列在表达感受态细胞中不能表达相应蛋白,影响因素很多,比如宿主菌、载体选择不合适,具体是何原因还需进一步研究。
4 结论
利用RT-PCR和RACE技术从玉米象体内获得了COX Ⅲ的全长cDNA序列。生物信息学分析表明COXⅢ具有该基因所编码蛋白典型生物特性;在pET-42a表达系统中可成功进行该基因所编码蛋白的原核表达。该研究工作可为后期蛋白质的活性研究以及AITC熏蒸剂对玉米象COX Ⅲ作用机理研究提供理论依据。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}