Overexpression of OsPR1A Enhanced Xa21-Mediated Resistance to Rice Bacterial Blight
LIU YuQing,1, YAN GaoWei1, ZHANG Tong2, LAN JinPing3, GUO YaLu4, LI LiYun1, LIU GuoZhen,1, DOU ShiJuan,11College of Life Sciences, Hebei Agricultural University, Baoding 071001, Hebei 2Beijing Institute of Biological Products Co., Ltd, Beijing 102600 3Life Science Research Center, Hebei North University, Zhangjiakou 075000, Hebei 4Agricultural Genomics Institute, Chinese Academy of Agricultural Sciences, Shenzhen 518116, Guangdong
Abstract 【Background】Previous studies revealed that the expression of pathogenesis-related protein OsPR1A was regulated by the upstream resistance gene Xa21. The rapid induction of OsPR1A protein at early stage after inoculation was crucial in Xa21-mediated rice bacterial blight resistance. The expression of OsPR1A was induced by Xanthomonas oryzae pv. oryzae (Xoo). OsPR1A was well known as a marker gene to demonstrate the reaction between rice and pathogen, however, no direct evidence was obtained for the biological function of OsPR1A. 【Objective】 In this paper, transgenic plants overexpression OsPR1A were obtained and the phenotype and agronomic traits were investigated. The relationship between OsPR1A expression and resistance were surveyed to further explore the function of OsPR1A in the process of rice resistance to bacterial blight.【Method】The construct of OsPR1a-OX was transferred to rice recipient 4021 via Agrobacterium-mediated protocol. Positive homozygous transgenic lines were identified by PCR and western blot (WB) respectively. At the mature stage, the phenotype and agronomic traits of OsPR1A overexpression rice plants were investigated, including plant height, spike length, tiller number, seed setting rate and grain size. Rice seedlings of TP309, 4021 and OsPR1A overexpression plants grown for two weeks were inoculated with Xoo at 31℃. The length of lesions was measured at 0, 2, 4, 6, 8, 10, and 12 days post-inoculation (dpi) respectively. At 0, 4 and 6 dpi, the rice leaves of TP309, 4021 and OsPR1A overexpression plants were collected to extract total protein, and the expression profiling of OsPR1A were surveyed by WB.【Result】The OsPR1a-OX transformation vector was constructed and transformed into recipient 4021. Two homozygous OsPR1A overexpression lines (#704 and #709) were identified. At the mature stage, the phenotype and agronomic traits of the OsPR1A overexpression plants were investigated. Compared with the control 4021, #704 and #709 lines showed lower plant height, shorter panicle length, fewer tiller number, lower seed-setting rate. The grain size in transgenic rice plants were larger, which might be related to the lower seed-setting rate. At 31℃, the lesion length of OsPR1A overexpression plants was significantly shorter than that of the control 4021 (P<0.05). At 0, 4, and 6 dpi, the abundance of OsPR1A expression of overexpression plants was higher than that of 4021 and TP309, and the high level of OsPR1A protein might contribute to the resistance of Xoo.【Conclusion】OsPR1A overexpression transgenic plants were obtained by the Agrobacterium-mediated method. Overexpression of OsPR1A affected the normal development of rice plants and also enhanced the resistance to bacterial blight mediated by Xa21. Keywords:rice;Xa21;bacterial blight;OsPR1A;vector construction;western blot
PDF (1495KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 刘玉晴, 燕高伟, 张彤, 兰金苹, 郭亚璐, 李莉云, 刘国振, 窦世娟. 超表达OsPR1A增强了Xa21介导的水稻对白叶枯病的抗性反应. 中国农业科学, 2021, 54(23): 4933-4942 doi:10.3864/j.issn.0578-1752.2021.23.001 LIU YuQing, YAN GaoWei, ZHANG Tong, LAN JinPing, GUO YaLu, LI LiYun, LIU GuoZhen, DOU ShiJuan. Overexpression of OsPR1A Enhanced Xa21-Mediated Resistance to Rice Bacterial Blight. Scientia Acricultura Sinica, 2021, 54(23): 4933-4942 doi:10.3864/j.issn.0578-1752.2021.23.001
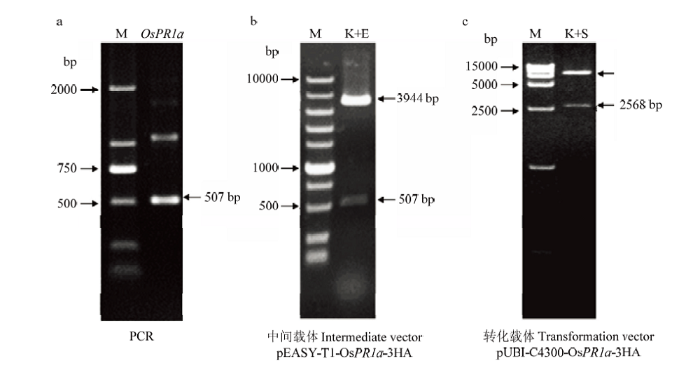
含有水稻目的片段OsPR1a(LOC_Os07g03710)全长的cDNA序列的质粒购自日本农业生物资源研究所水稻基因组资源中心(Rice Genome Resource Center,National Institute of Agrobiological Sciences)。扩增目的片段使用的引物分别为OsPR1a-OX-F:5’-GCGGTACCATGGCGAGTTCGTCGAGCAGG-3’(下划线为KpnⅠ酶切位点),OsPR1a-OX-R:5’-GCGAATTCTCAGTAGGGAGATTGGCCGAC-3’(下划线为EcoRⅠ酶切位点)。
a:水稻成熟期表型图,bar=100 cm;b:水稻穗部表型图,bar=10 cm;c:水稻籽粒表型图,bar=1cm;d:株高统计学分析;e:穗长统计学分析;f:分蘖数统计学分析;g:结实率统计学分析;*表示P<0.05,**表示P<0.01。下同 Fig. 3The phenotype and agronomic traits of OsPR1a overexpression transgenic plants
a: Phenotype of rice at mature stage, bar=100 cm; b: Phenotype of rice panicle, bar=10 cm; c: Phenotype of rice grain, bar=1 cm; d: Statistical analysis of plant height; e: Statistical analysis of panicle length; f: Statistical analysis of tiller number; g: Statistical analysis of seed setting rate; * means P <0.05, ** means P<0.01. The same as below
a:接种12 d内不同材料的叶片病斑生长曲线,平均值±标准误差(n=4);b:水稻接种Xoo第12天时不同抗感材料和OsPR1a-OX转基因材料的叶片表型图,bar=1 cm;c:接种第12天时不同材料的叶片病斑长度统计学分析,平均值±标准误差(n=4) Fig. 4The lesion growth curve and phenotype of OsPR1a overexpression transgenic plants after inoculation.
a: The growth curve of leaf lesions of different materials within 12 days of inoculation, average ± standard error (n=4); b: Leaf phenotype of TP309, 4021 and OsPR1a-OX transgenic materials at 12 dpi, bar=1 cm; c: Statistical analysis of leaf lesion length of different materials at 12 dpi, average ± standard error (n=4)
JIANGN, YANJ, LIANGY, SHIY, HEZ, WUY, ZENGQ, LIUX, PENGJ. Resistance genes and their interactions with bacterial blight/leaf streak pathogens (Xanthomonas oryzae) in rice (Oryza sativa L.)-An updated review Rice, 2020, 13(1):3-14. DOI:10.1186/s12284-019-0358-yURL [本文引用: 1]
LIUW, LIUJ, TRIPLETTL, LEACH JE, WANG GL. Novel insights into rice innate immunity against bacterial and fungal pathogens Annual Review of Phytopathology, 2014, 52:213-241. DOI:10.1146/phyto.2014.52.issue-1URL [本文引用: 1]
AGRAWAL GK, JWA NS, RAKWALR. A novel rice (Oryza sativa L.) acidic PR1 gene highly responsive to cut, phytohormones, and protein phosphatase inhibitors Biochemical and Biophysical Research Communications, 2000, 274(1):157-165. DOI:10.1006/bbrc.2000.3114URL [本文引用: 4]
NEELAMK, MAHAJANR, GUPTAV, BHATIAD, GILL BK, KOMALR, LORE JS, MANGAT GS, SINGHK. High-resolution genetic mapping of a novel bacterial blight resistance gene xa-45(t) identified from Oryza glaberrima and transferred to Oryza sativa Theoretical and Applied Genetics, 2020, 133(3):689-705. DOI:10.1007/s00122-019-03501-2URL [本文引用: 1]
JIZ, WANGC, ZHAOK. Rice routes of countering Xanthomonas oryzae. International Journal of Molecular Sciences, 2018, 19(10):3008-3022. [本文引用: 1]
LUOD, HUGUET-TAPIAJ C, RABORNR T, WHITEF F, BRENDELV P, YANGB. The Xa7 resistance gene guards the rice susceptibility gene SWEET14 against exploitation by the bacterial blight pathogen Plant Communications, 2021, 2(3):100164-100188. DOI:10.1016/j.xplc.2021.100164URL [本文引用: 1]
SONG WY, WANG GL, CHEN LL, KIM HS, PI LY, HOLSTENT, GARDNERJ, WANGB, ZHAI WX, ZHU LH, FAUQUETC, RONALDP. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science, 1995, 270(5243):1804-1806. DOI:10.1126/science.270.5243.1804URL [本文引用: 1]
SONG WY, PI LY, WANG GL, GARDNERJ, HOLSTENT, RONALD PC. Evolution of the rice Xa21 disease resistance gene family The Plant Cell, 1997, 9(8):1279-1287. [本文引用: 1]
CHENX, CHERNM, CANLAS PE, RUAND, JIANGC, RONALD PC. An ATPase promotes autophosphorylation of the pattern recognition receptor XA21 and inhibits XA21-mediated immunity Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(17):8029-8034. [本文引用: 2]
VO K TX, KIM CY, HOANG TV, LEE SK, SHIRSEKARG, SEO YS, LEE SW, WANG GL, JEON JS. OsWRKY67 plays a positive role in basal and XA21-mediated resistance in rice Frontiers in Plant Science, 2018, 8:2220-2233. DOI:10.3389/fpls.2017.02220URL [本文引用: 2]
LUU DD, JOEA, CHENY, PARYSK, BAHARO, PRUITTR, CHANL J G, PETZOLDC J, LONGK, ADAMCHAKC, STEWARTV, BELKHADIRY, RONALDP C. Biosynthesis and secretion of the microbial sulfated peptide RaxX and binding to the rice XA21 immune receptor Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(17):8525-8534. [本文引用: 1]
PENGY, BARTLEY LE, CHENX, DARDICKC, CHERNM, RUANR, CANLAS PE, RONALD PC. OsWRKY62 is a negative regulator of basal and Xa21-mediated defense against Xanthomonas oryzae pv. oryzae in rice Molecular Plant, 2008, 1(3):446-458. DOI:10.1093/mp/ssn024URL [本文引用: 2]
PARK CJ, PENGY, CHENX, DARDICKC, RUAND, BARTR, CANLAS PE, RONALD PC. Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity PLoS Biology, 2008, 6(9):e231. DOI:10.1371/journal.pbio.0060231URL [本文引用: 1]
PARK CJ, BARTR, CHERNM, CANLAS PE, BAIW, RONALD PC. Overexpression of the endoplasmic reticulum chaperone BiP3 regulates XA21-mediated innate immunity in rice PLoS ONE, 2010, 5(2):e9262. DOI:10.1371/journal.pone.0009262URL [本文引用: 1]
WANG YS, PI LY, CHENX, CHAKRABARTY PK, JIANGJ, DELEON A L, LIUG Z, LIL, BENNYU, OARDJ, RONALDP C, SONGW Y. Rice XA21 binding protein 3 is a ubiquitin ligase required for full Xa21-mediated disease resistance The Plant Cell, 2006, 18(12):3635-3646. DOI:10.1105/tpc.106.046730URL [本文引用: 1]
PARK CJ, WEIT, SHARMAR, RONALD PC. Overexpression of rice auxilin-like protein, XB21, induces necrotic lesions, up-regulates endocytosis-related genes, and confers enhanced resistance to Xanthomonas oryzae pv. oryzae. Rice, 2017, 10(1):27-38. DOI:10.1186/s12284-017-0166-1URL [本文引用: 1]
JIANGY, CHENX, DINGX, WANGY, CHENQ, SONG WY. The XA21 binding protein XB25 is required for maintaining XA21- mediated disease resistance The Plant Journal, 2013, 73(5):814-823. DOI:10.1111/tpj.2013.73.issue-5URL [本文引用: 1]
HUH, WANGJ, SHIC, YUANC, PENGC, YINJ, LIW, HEM, WANGJ, MAB, WANGY, LIS, CHENX. A receptor like kinase gene with expressional responsiveness on Xanthomonas oryzae pv. oryzae is essential for Xa21-mediated disease resistance Rice, 2015, 8(1):34-42. DOI:10.1186/s12284-015-0069-yURL [本文引用: 1]
CHENX, ZUPS, SCHWESSINGERB, CHERNM, CANLAS PE, RUAND, ZHOUX, WANGJ, DAUDIA, PETZOLD CJ, HEAZLEWOOD JL, RONALD PC. An XA21-associated kinase (OsSERK2) regulates immunity mediated by the XA21 and XA3 immune receptors Molecular Plant, 2014, 7(5):874-892. DOI:10.1093/mp/ssu003URL [本文引用: 1]
CHOIC, HWANG SH, FANG IR, KWON SI, PARK SR, AHNI, KIM JB, HWANG DJ. Molecular characterization of Oryza sativa WRKY6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR)10a promoter and confers reduced susceptibility to pathogens The New Phytologist, 2015, 208(3):846-859. DOI:10.1111/nph.2015.208.issue-3URL [本文引用: 1]
SONS, AN HK, SEOL YJ, PARK SR, IM JH. Rice transcription factor WRKY114 directly regulates the expression of OsPR1a and chitinase to enhance resistance against Xanthomonas oryzae pv. oryzae Biochemical and Biophysical Research Communications, 2020, 533(4):1262-1268. DOI:10.1016/j.bbrc.2020.09.141URL [本文引用: 1]
WANGG, DINGX, YUANM, QIUD, LIX, XUC, WANGS. Dual function of rice OsDR8 gene in disease resistance and thiamine accumulation Plant Molecular Biology, 2006, 60(3):437-449. DOI:10.1007/s11103-005-4770-xURL [本文引用: 1]
WANGH, MENGJ, PENGX, TANGX, ZHOUP, XIANGJ, DENGX. Rice WRKY4 acts as a transcriptional activator mediating defense responses toward Rhizoctonia solani, the causing agent of rice sheath blight Plant Molecular Biology, 2015, 89(1/2):157-171. DOI:10.1007/s11103-015-0360-8URL [本文引用: 2]
MEIC, QIM, SHENGG, YANGY. Inducible overexpression of a rice allene oxide synthase gene increases the endogenous jasmonic acid level, PR gene expression, and host resistance to fungal infection Molecular Plant-Microbe Interactions, 2006, 19(10):1127-1137. DOI:10.1094/MPMI-19-1127URL [本文引用: 2]
WUQ, HOU MM, LI LY, LIU LJ, HOU YX, LIU GZ. Induction of pathogenesis-related proteins in rice bacterial blight resistant gene XA21-mediated interactions with Xanthomonas oryzae pv. oryzae Journal of Plant Pathology, 2011, 93(2):455-459. [本文引用: 2]
CHENQ, HUANGX, CHENX, SHAMSUNNAHER, SONGW Y. Reversible activation of XA21-mediated resistance by temperature European Journal of Plant Pathology, 2018, 153(4):1177-1184. DOI:10.1007/s10658-018-01634-6URL [本文引用: 2]
YAN GW. The functional analysis of rice pathogenesis-related protein OsPR1A in bacterial leaf blight resistance response [D]. Baoding: Hebei Agricultural University, 2020. (in Chinese) [本文引用: 4]
CHENY, WANGT X Z, YANGS, ZHANGT, MAJ J, YANG W, LIUY Q, ZHOUY, SHIJ N, LANJ P, WEIJ, DOUS J, LIUL J, YANGM, LIL Y, LIUG Z. Expression profiling and functional characterization of rice transcription factor OsWRKY68 Scientia Agricultura Sinica, 2019, 52(12):2021-2032. (in Chinese) [本文引用: 1]
LIX, HUIB, WANGX, LIL, CAOY, JIANW, LIUY, LIUL, GONGX, LINW. Identification and validation of rice reference proteins for western blotting Journal of Experimental Botany, 2011, 62(14):4763-4772. DOI:10.1093/jxb/err084URL [本文引用: 1]
ALIS, GANAI BA, KAMILI AN, BHAT AA, MIR ZA, BHAT JA, TYAGIA, ISLAM ST, MUSHTAQM, YADAVP, RAWATS, GROVERA. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance Microbiological Research, 2018(212/213):29-37. [本文引用: 1]
LOON L CV, KAMMEN AV. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. “Samsun” and “Samsun NN”: II. Changes in protein constitution after infection with tobacco mosaic virus Virology, 1970, 40(2):190-211. [本文引用: 1]
BOL JF, LINTHORSTH J M, CORNELISSENB J C. Plant pathogenesis-related proteins induced by virus infection Annual Review of Phytopathology, 1990, 28(1):113-138. DOI:10.1146/phyto.1990.28.issue-1URL
VAN LOON LC, REPM, PIETERSE CM. Significance of inducible defense related proteins in infected plants Annual Review of Phytopathology, 2006, 44:135-162. DOI:10.1146/phyto.2006.44.issue-1URL [本文引用: 1]
SELSJ, MATHYSJ, DECONINCK B M, CAMMUEB P, DEBOLLE M F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides Plant Physiology and Biochemistry, 2008, 46(11):941-950. DOI:10.1016/j.plaphy.2008.06.011URL [本文引用: 1]
DOU SJ, GUAN ML, LI LY, LIU GZ. Pathogenesis-related genes in rice Chinese Science Bulletin, 2014, 59(3):245-258. (in Chinese) [本文引用: 1]
AGRAWAL GK, RAKWALR, JWA NS, AGRAWAL VP. Signalling molecules and blast pathogen attack activates rice OsPR1a and OsPR1b genes: A model illustrating components participating during defence/stress response Plant Physiology and Biochemistry, 2001, 39(12):1095-1103. DOI:10.1016/S0981-9428(01)01333-XURL [本文引用: 1]
DAL DEGANF, ROCHERA, CAMERON-MILLSV, VON WETTSTEIND. The expression of serine carboxypeptidases during maturation and germination of the barley grain Proceedings of the National Academy of Sciences of the United States of America, 1994, 91(17):8209-8213. [本文引用: 1]
LIY, FANC, XINGY, JIANGY, LUOL, SUNL, SHAOD, XUC, LIX, XIAOJ, HEY, ZHANGQ. Natural variation in GS5 plays an important role in regulating grain size and yield in rice Nature Genetics, 2011, 43(12):1266-1269. DOI:10.1038/ng.977URL [本文引用: 2]
XUP, JIANGL, WUJ, LIW, ZHANGS. Isolation and characterization of a novel pathogenesis-related protein gene (GmPRP) with induced expression in soybean (Glycine max) during infection with Phytophthora sojae Molecular Biology Reports, 2015, 10(6):4899-4909. [本文引用: 1]
LIJ, LEASE KA, TAX FE, WALKER JC. BRS1, a serine carboxypeptidase, regulates BRI1 signaling in Arabidopsis thaliana Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(10):5916-5921. [本文引用: 1]
MUGFORD ST, QIX, BAKHTS, HILLL, WEGELE, HUGHES RK, PAPADOPOULOUK, MELTONR, PHILOM, SAINSBURYF, LOMONOSSOFF GP, ROY AD, GOSS RJ, OSBOURNA. A serine carboxypeptidase-like acyltransferase is required for synthesis of antimicrobial compounds and disease resistance in oats The Plant Cell, 2009, 21(8):2473-2484. DOI:10.1105/tpc.109.065870URL [本文引用: 1]
BONTPARTT, FERREROM, KHATERF, MARLINT, VIAlETS, VALLVERDU-QUERALTA, PINASSEAUL, AGEORGESA, CHEYNIERV, TERRIERN. Focus on putative serine carboxypeptidase- like acyltransferases in grapevine Plant Physiology and Biochemistry, 2018, 130:356-366. DOI:10.1016/j.plaphy.2018.07.023URL [本文引用: 1]
WOLF AE, DIETZ KJ, SCHRODERP. Degradation of glutathione S-conjugates by a carboxypeptidase in the plant vacuole FEBS Letters, 1996, 384(1):31-34. DOI:10.1016/0014-5793(96)00272-4URL [本文引用: 1]
LIZ, TANGL, QIUJ, ZHANGW, WANGY, TONGX, WEIX, HOUY, ZHANGJ. Serine carboxypeptidase 46 regulates grain filling and seed germination in rice (Oryza sativa L.) PLoS ONE, 2016, 11(7):e0159737. DOI:10.1371/journal.pone.0159737URL [本文引用: 1]
CENTURY KS, LAGMAN RA, ADKISSONM, MORLANJ, TOBIASR, SCHWARTZK, SMITHA, LOVEJ, RONALD PC, WHALEN MC. Short communication: Developmental control of Xa21-mediated disease resistance in rice The Plant Journal, 1999, 20(2):231-236. DOI:10.1046/j.1365-313x.1999.00589.xURL [本文引用: 1]
PRUITT RN, SCHWESSINGERB, JOEA, THOMASN, LIUF, ALBERTM, ROBINSON MR, CHAN LJ, LUU DD, CHENH, BAHARO, DAUDIA, DE VLEESSCHAUWERD, CADDELLD, ZHANGW, ZHAOX, LIX, HEAZLEWOOD JL, RUAND, MAJUMDERD, CHERNM, KALBACHERH, MIDHAS, PATIL PB, SONTI RV, PETZOLD CJ, LIU CC, BRODBELT JS, FELIXG, RONALD PC. The rice immune receptor XA21 recognizes a tyrosine-sulfated protein from a Gram-negative bacterium Science Advances, 2015, 1(6):e1500245. DOI:10.1126/sciadv.1500245URL [本文引用: 1]
 ,1, 燕高伟1, 张彤2, 兰金苹3, 郭亚璐4, 李莉云1, 刘国振
,1, 燕高伟1, 张彤2, 兰金苹3, 郭亚璐4, 李莉云1, 刘国振

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT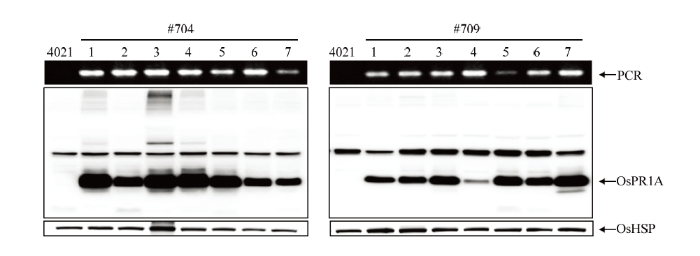 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT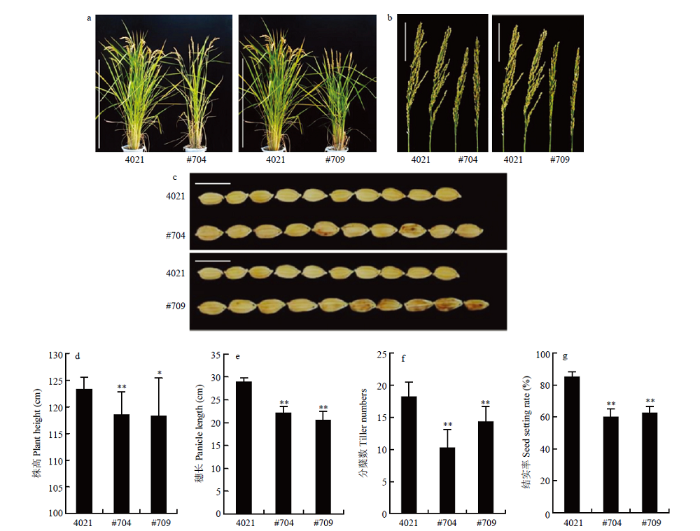 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT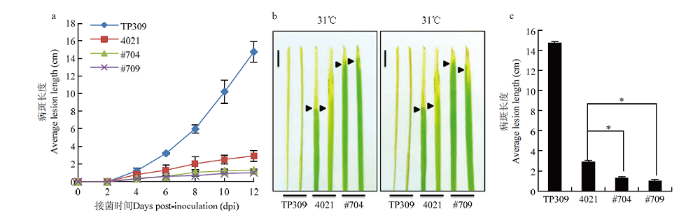 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT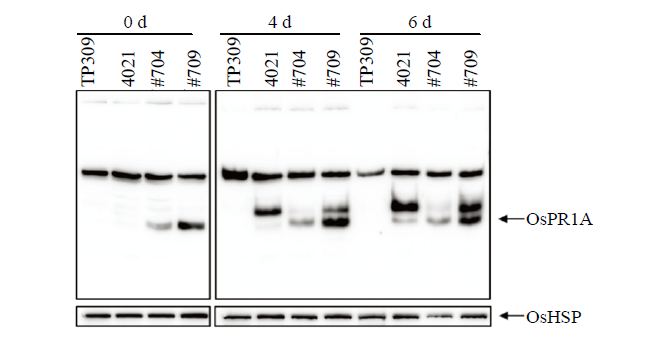 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}