 ,, 李紫腾,, 张静怡, 张静娜, 胡同乐, 王树桐, 王亚南,, 曹克强,河北农业大学植物保护学院,河北保定 071001
,, 李紫腾,, 张静怡, 张静娜, 胡同乐, 王树桐, 王亚南,, 曹克强,河北农业大学植物保护学院,河北保定 071001Analysis of dsRNA Carried by Alternaria alternata f. sp. mali in China and Identification of a dsRNA Virus
CAO YuHan,, LI ZiTeng,, ZHANG JingYi, ZHANG JingNa, HU TongLe, WANG ShuTong, WANG YaNan,, CAO KeQiang,College of Plant Protection, Hebei Agricultural University, Baoding 071001, Hebei通讯作者:
责任编辑: 岳梅
收稿日期:2021-04-6接受日期:2021-05-10
| 基金资助: |
Received:2021-04-6Accepted:2021-05-10
作者简介 About authors
曹钰晗,E-mail:
李紫腾,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (3794KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
曹钰晗, 李紫腾, 张静怡, 张静娜, 胡同乐, 王树桐, 王亚南, 曹克强. 我国苹果斑点落叶病菌携带dsRNA分析及一种dsRNA病毒的鉴定. 中国农业科学, 2021, 54(22): 4787-4799 doi:10.3864/j.issn.0578-1752.2021.22.007
CAO YuHan, LI ZiTeng, ZHANG JingYi, ZHANG JingNa, HU TongLe, WANG ShuTong, WANG YaNan, CAO KeQiang.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】苹果斑点落叶病(apple Alternaria blotch)又称褐纹病,是由链格孢苹果专化型(Alternaria alternata f. sp. mali)侵染而引起的一种世界性分布气传病害,该病危害苹果叶片、新梢和果实,严重影响苹果生产[1,2],现已成为亚洲国家苹果产区的主要病害[3,4]。目前,苹果斑点落叶病的防治途径主要包括种植抗性品种、化学防治和生物防治,生产上一直以使用化学药剂防治为主[5,6,7],但随着经济的发展,人们开始关注食品安全和环境问题,一些化学杀菌剂的使用已在发达国家和地区受到了严重限制[8,9],生物防治日益受到人们的关注,利用携带病毒的弱毒株引起病原菌群体致病力衰退,将成为真菌病害生物防治的新途径[10,11]。【前人研究进展】真菌病毒是指能够侵染真菌并在其中完成自我复制的病毒。真菌病毒的分类依据主要有两个,首先依据真菌病毒的核酸类型将其分为DNA病毒和RNA病毒,就目前所发现的真菌病毒而言,大多数为RNA病毒,包括双链RNA(dsRNA)和单链RNA(ssRNA)[12,13]。从具有弱毒特性的病毒入手寻求新的生防因子,对深入了解病毒及其对寄主真菌的影响具有重要意义。链格孢属真菌中也发现了很多真菌病毒,1988年,SHEPHERD[14]从棉花种子中分离出链格孢菌株共21个,其中6株携带真菌病毒,且这6株均不产生色素;1997年,ZABALGOGEAZCOA等[15]从茄链格孢(A. solani)中检测到未经包被的两条大小分别为8.3和5.5 kb的dsRNA;1988年,HAYASHI等[16]研究的12个链格孢日本梨致病型(A. alternata Japanese pear pathotype)菌株中,有7个菌株含有dsRNA;2009年,AOKI等[17]从链格孢菌株EGS 35-193中分离到Alternaria alternata virus 1(AaV1),该病毒使菌株的菌丝生长变得异常,并且出现大量的囊泡结构;2015年,LIN等[18]从长柄链格孢(A. longipes)HN28中发现Alternaria longipes dsRNA virus 1(AlRV1),其分类地位尚未明确;2016年,KOMATSU等[19]从乔木链格孢(A. arborescens) 中发现一个新的维多利亚病毒Alternaria arborescens victorivirus 1(AaVV1);ZHONG等[20]从芸薹生链格孢(A. brassicicola)中发现病毒 Alternaria brassicicola fusarivirus 1(AbFV1);2017年,向均[21]从链格孢日本梨致病型中分离出病毒Alternaria botybirna virus 1(ABRV1);2018年,日本****OKADA等[22]从链格孢日本梨致病型中分离出病毒Alternaria alternata chrysovirus 1(AaCV1)。AaCV1损害宿主真菌的生长,同时影响宿主AK毒素的产生。【本研究切入点】目前,已报道了多种作物链格孢属真菌病毒,但关于苹果斑点落叶病菌真菌病毒还未见报道,有待进一步解析。【拟解决的关键问题】在我国苹果产区8个省份采集苹果斑点落叶病样本,通过dsRNA提取,明确我国苹果斑点落叶病菌携带dsRNA的多样性;通过生物学测定揭示携带dsRNA菌株的生物学特征,分析dsRNA多样性与寄主病原菌生物学性状的关系;最后,鉴定弱毒力菌株携带dsRNA病毒的种类。为寻找对苹果斑点落叶病具有生防潜力的弱毒菌株打下基础。1 材料与方法
1.1 供试材料
102株苹果斑点落叶病菌菌株:2018年5—8月自我国河北、河南、山东、甘肃、辽宁、黑龙江、陕西及云南8个省份采集有典型斑点落叶症状的叶片样本,采用组织分离法和单孢子稀释法进行病原菌的分离纯化[23]。在叶片和果实上利用柯赫氏法则进行验证。具体样本信息见表1。Table 1
表1
表1本研究中苹果斑点落叶病样本信息
Table 1
| 序号 Number | 菌株编号 Strain number | 采集地点 Collection location | 采集时间 Collection time | 品种 Cultivar | 树龄 Tree age (a) |
|---|---|---|---|---|---|
| 1 | ND | 河北保定Baoding, Hebei | 05-10 | 中秋王Mid-autumn King | 10 |
| 2-7 | WA-3, WA-8, WA-17, WA-20, WA-25, WA-28 | 河北武安Wuan, Hebei | 05-12 | 富士Fuji | 10 |
| 8-13 | QY-2, QY-3, QY-4, QY-5, QY-6, QY-8 | 河北保定Baoding, Hebei | 05-22 | 藤木一号Vine Wood One | 10 |
| 14-16 | SMX-1-1, SMX-1-3, SMX-1-9 | 河南三门峡Sanmeixia, Henan | 06-22 | 金冠Golden Delicious | 6 |
| 17 | SMX-2-1 | 河南三门峡Sanmeixia, Henan | 06-22 | 嘎啦Gala | 6 |
| 18 | SMX-3-1 | 河南三门峡Sanmeixia, Henan | 06-22 | 红尾Red Tail | 6 |
| 19-24 | WH-5, WH-8, WH-12, WH-15, WH-17, WH-3-1 | 山东威海Weihai, Shandong | 06-21 | 威海金Weihai Gold | 3 |
| 25-31 | SQ-1-1, SQ-1-2, SQ-1-5, SQ-1-6, SQ-1-7, SQ- 1-8, SQ-1-9 | 河南商丘Shangqiu, Henan | 06-21 | 礼泉Liquan | 9 |
| 32-33 | SQ-2-2, SQ-2-3 | 河南商丘Shangqiu, Henan | 06-21 | 新红星New Red Star | 24 |
| 34-40 | SQ-3-1, SQ-3-6, SQ-3-7, SQ-3-8, SQ-3-11, SQ- 3-13, SQ-3-14 | 河南商丘Shangqiu, Henan | 06-21 | 长富2号Changfu 2 | 24 |
| 41-44 | CL-2-6, CL-2-3, CL-2-9, CL-2-12 | 河北昌黎Changli, Hebei | 06-21 | 太平洋玫瑰Pacific Rose | 4 |
| 45-49 | CL-3-1-1, CL-3-2-1, CL-3-2-2, CL-3-2-3, CL-3-2-4 | 河北昌黎Changli, Hebei | 06-21 | 秋香Qiuxiang | 3 |
| 50-55 | CL-4-1, CL-4-2, CL-4-9, CL-4-7, CL-4-6, CL-4-13 | 河北昌黎Changli, Hebei | 06-21 | 北斗Beidou | 4 |
| 56 | YA | 陕西延安Yan’an, Shaanxi | 06-23 | 延丰Yanfeng | 8 |
| 57-62 | LN-2, LN-6, LN-7, LN-8, LN-9, LN-10 | 辽宁Liaoning | 06-29 | 绿帅Lvshuai | 13 |
| 63 | YT-2-1 | 山东烟台Yantai, Shandong | 07-03 | 富士Fuji | 5 |
| 64-66 | YT-3-1, YT-3-7, YT-3-9 | 山东烟台Yantai, Shandong | 07-03 | 岳红Yuehong | 13 |
| 67-69 | YT-4-1, YT-4-2, YT-4-3 | 山东烟台Yantai, Shandong | 07-03 | 粉红女士Pink Lady | 13 |
| 70-77 | HLJ-1-3, HLJ-1-4, HLJ-1-6, HLJ-1-7, HLJ-1-8, HLJ-1-9, HLJ-1-10, HLJ-1-11 | 黑龙江牡丹江Mudanjiang, Heilongjiang | 07-14 | QRZ-1 | 6 |
| 78-85 | HLJ-2-1, HLJ-2-2, HLJ-2-3, HLJ2-5, HLJ-2-6, HLJ2-7, HLJ2-8, HLJ-2-9 | 黑龙江牡丹江Mudanjiang, Heilongjiang | 07-14 | QRZ-2 | 5 |
| 86-90 | GS-1-1, GS-1-2, GS-1-8, GS-1-14, GS-1-17 | 甘肃天水Tianshui, Gansu | 07-17 | 富士Fuji | 未知Unknown |
| 91-95 | GS-3-2, GS-3-3, GS-3-7, GS-3-11, GS-3-12 | 甘肃天水Tianshui, Gansu | 07-17 | 金冠Golden Delicious | 未知Unknown |
| 96-100 | SJZ-1, SJZ-2, SJZ-3, SJZ-4, SJZ-8 | 河北石家庄Shijiazhuang, Hebei | 07-20 | 嘎啦Gala | 8 |
| 101 | YN-1-2 | 云南石林Shilin, Yunnan | 08-20 | 红露Honglu | 4 |
| 102 | YN-2-3 | 云南弥勒Mile, Yunnan | 08-20 | 富士Fuji | 3 |
新窗口打开|下载CSV
1.2 主要试剂、仪器和引物
1.2.1 主要试剂和仪器 氯仿,北京酷莱博科技有限公司;纤维素粉,阿拉丁试剂(上海)有限公司;3M乙酸钠、氨苄,北京索莱宝科技有限公司;Taq Mix、载体PMD18-T,宝生物工程有限公司;感受态细胞Tans1-T、TRlzol、反转录试剂盒,北京全式金生物科技有限公司;DNA胶回收试剂盒,生工生物工程(上海)股份有限公司;RNeasy Mini Kit总RNA提取试剂盒,杭州沃森生物技术有限公司。组织研磨破碎仪,德国QIAGEN公司;台式冷冻高速离心机,默克生命科学(上海)有限公司;超低温冰箱、PCR仪,赛默飞世尔科技;DYY-12型电泳仪、DYY-31DN型琼脂糖水平电泳槽,北京六一仪器厂;BioRad凝胶成像系统,美国SYNGENE公司;超净工作台,江苏苏净集团公司。1.2.2 引物 本研究所用引物具体信息见表2。
Table 2
表2
表2本试验所用引物
Table 2
| 引物名称 Primer name | 引物序列 Primer sequence (5′-3′) | 条带大小 Product size (bp) | 用途 Purpose | 来源 Source |
|---|---|---|---|---|
| CRV1-5F | GCAAAAAAGAACTAAAGGAC | 800 | AaCV2 ORF5的扩增 Amplification of AaCV2 ORF5 | 自行设计 Designed in this study |
| CRV1-5R | ACACACAAATGGGATACC | |||
| CRV1-1F | ATCTCAAGGTACCGGAGTCA | 450 | AaCV2 RT-PCR验证 Identification of AaCV2 by RT-PCR | 自行设计 Designed in this study |
| CRV1-1R | GCTGCTTAAACATCACCTGCA |
新窗口打开|下载CSV
1.3 dsRNA的提取
参考李波[24]的报道,采用纤维素粉的方法对102株苹果斑点落叶病菌纯化菌株进行dsRNA提取,最后加入无水乙醇和乙酸钠(3 mol·L-1)在-80℃沉淀,12 000 r/min离心后,用1 mL的70%乙醇洗涤沉淀,20 μL DEPC H2O溶解,取8 μL在1%(W/V)的琼脂糖凝胶电泳中检测(电压80 V,50 min),紫外灯下观察dsRNA电泳条带。1.4 苹果斑点落叶病菌带毒菌株的生物学特性
1.4.1 培养特性及菌丝生长速率的测定 将5株带毒菌株及随机选取的20株不带毒菌株同时扩繁,25℃ PDA培养7 d后,观察菌落的形状、颜色,每个菌株设置3个重复。用十字交叉法测菌落直径,计算生长速率。1.4.2 在苹果叶片和果实上致病力的测定 采集海棠幼嫩无病叶片,清水洗净,放于铺有两层浸湿滤纸的瓷盘中,使用接种针将叶片的叶脉一侧刺伤;从25个苹果斑点落叶病菌菌株的PDA平板相同位置的边缘打取菌饼,将菌丝块倒置于叶片的伤口处,每个菌株设置3个重复;用保鲜膜将瓷盘包裹严实,置于25℃条件下培养7 d后,测量病斑大小。
取表观相近的同一批次健康富士苹果,先清水冲洗,再用75%的酒精擦洗干净,75%酒精淋湿,放于超净工作台中吹干。用接种针将果实表面刺一个针孔,从25个苹果斑点落叶病菌菌株的PDA平板相同位置边缘打取菌饼,有菌丝的一面贴于伤口处,保鲜膜包裹保湿,每个菌株至少3个重复。25℃培养7 d后测量病斑大小。
1.4.3 数据处理 数据采用SPSS Statistics 17.0单因素分析法中的均值比较法进行分析。
1.5 菌株QY-2携带病毒的高通量测序
1.5.1 RNA的抽提与质检 采用RNeasy Mini Kit进行菌株QY-2的total RNA抽提,然后通过Agilent Bioanalyzer 2100电泳分析仪进行RNA质量检测,合格后纯化total RNA。1.5.2 文库构建与测序 委托上海伯豪生物技术有限公司采用宏转录组测序的方法,对纯化后的total RNA进行rRNA去除、片段化、第一链cDNA合成、第二链cDNA合成、末端修复、3′末端加A、连接接头、富集等步骤,完成测序样本文库构建,采用paired-end程序,进行双端测序。测序过程由Illumina提供的data collection software进行实时数据分析。
1.5.3 QY-2携带病毒种类分析及基因组序列分析 通过高通量测序结果分析QY-2菌株携带的病毒种类及占比。将样本测序获得的reads进行reference mapping,获得病毒reads覆盖度,用RPKM(reads per kilo bases per million reads)量化标准对reads覆盖度进行表达量计算,计算公式:transcription reads /(transcription length×total assembly reads in run)其中,transcription reads为覆盖病毒整个UniGene的reads数目,transcription length为UniGene长度,total assembly reads in run为该样本所有参与拼接的总reads数目。通过Vector NTI 11.0软件去除载体序列和序列比对;NCBI上预测开放阅读框(ORF);运用MEGA 7.0中的邻接法进行序列系统发育分析,重复1 000次。
1.6 菌株QY-2携带病毒的RT-PCR鉴定
1.6.1 病毒特异性引物的设计 根据已报道的病毒序列,使用Premier5.0设计该病毒的特异性引物,委托生工生物工程(上海)股份有限公司进行合成。引物具体信息见表2。1.6.2 总RNA的提取 取适量纯化好的新鲜菌丝,液氮研磨成粉末状,转移至2 mL离心管中,加入1 mL的Trizol试剂,颠倒混匀,室温下放置5 min;继续加入200 μL的氯仿,剧烈振荡15 s,室温下放置3 min;4℃,14 000 r/min离心15 min;取上清700 μL到1.5 mL的离心管中,加入700 μL的异丙醇,颠倒混匀,室温下放置10 min;4℃,14 000 r/min离心10 min,弃上清,吸除残留;加入1 mL 75%的乙醇洗涤沉淀;4℃,9 000 r/min离心5 min,弃上清,短暂离心后吸除残留的乙醇;室温干燥5—10 min,加入50 μL的DEPC-H2O。
1.6.3 反转录合成cDNA 使用全式金反转录试剂盒合成cDNA,操作方法参考说明书。具体步骤如下:dsRNA样品2 μL;Random Primer(N9)(0.1 μg·μL-1)1 μL;2×ES Reaction Mix 10 μL;Easy Script RT Mix 1 μL;DEPC-H2O 6 μL。25℃反应10 min,42℃反应30 min,然后85℃反应5 min,即为cDNA。
PCR反应体系:cDNA样品,1 μL;2×Taq mix,10 μL;引物F 1 μL;引物R 1 μL;DEPC-H2O 7 μL。PCR反应程序:94℃预变性5 min,94℃变性30 s,58℃退火30 s,72℃延伸45 s,35个循环后,72℃完全延伸7 min。1%琼脂糖凝胶电泳检测目的片段。
1.6.4 目的片段的克隆测序 采用琼脂糖凝胶回收试剂盒回收DNA,按试剂盒说明书进行操作。按照pMD18-T载体试剂盒说明书进行载体连接,利用感受态细胞Tans1-T进行转化,挑取单菌落到含有氨苄抗生素的液体LB中,37℃,200 r/min振荡至液体浑浊。以菌液为模板,菌落PCR鉴定阳性克隆,阳性克隆委托华大基因测序公司进行测序。
2 结果
2.1 苹果斑点落叶病菌的dsRNA多态性分析
对102株苹果斑点落叶病菌菌株进行dsRNA的提取,使用DNase I及S1核酸酶消解,在102个菌株中有5个菌株携带dsRNA,如图1所示,菌株名称分别为CL-2-6、QY-2、SQ-1-1、SJZ-4、YT-3-7。dsRNA分为4种类型:菌株YT-3-7携带的dsRNA条带大小在8、2.5和1.5 kb左右;菌株SJZ-4携带的dsRNA条带在8 kb左右;菌株QY-2携带的dsRNA条带在3和0.8 kb左右;菌株CL-2-6和SQ-1-1携带的dsRNA条带在2.5和1.5 kb左右。图1
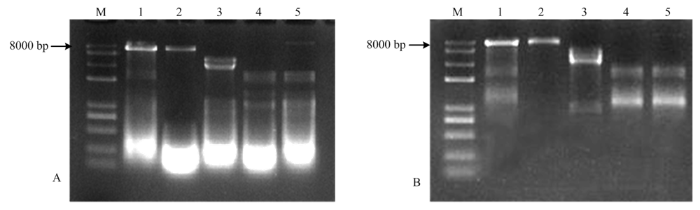 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1我国苹果斑点落叶病菌dsRNA的条带类型
A:酶处理前dsRNA Untreated dsRNA;B:酶处理后dsRNA Enzyme-treated dsRNA
Fig. 1DsRNA types of A. alternata f. sp. mali in China
M: DL8000; 1: YT-3-7; 2: SJZ-4; 3: QY-2; 4: CL-2-6; 5: SQ-1-1
2.2 苹果斑点落叶病菌带毒菌株的培养性状
将苹果斑点落叶病菌带毒菌株和不带毒菌株在PDA培养基上25℃黑暗培养7 d,观察菌落颜色、气生菌丝形态等特征,将5个带毒菌株划分为两类。第1类(图2):菌丝灰白色或灰绿色,个别菌落后期有黑色素产生。菌丝分布均匀,多数菌株的菌丝层较薄,个别菌株的气生菌丝发达呈棉絮状。具有这一类特征的菌株有12个,包括两个带毒菌株SJZ-4和CL-2-6以及10个不带毒菌株,分别为HLJ-2-1、HLJ-2-3、CL-4-1、QY-5、QY-4、YT-3-9、YT-4-3、HLJ-1-6、HLJ-2-9、SQ-1-2。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2苹果斑点落叶病菌菌株的培养类型 I
A、B:带毒菌株The strains taking virus (SJZ-4, CL-2-6);C—L:未带毒菌株The strains without virus (HLJ-2-1, HLJ-2-3, CL-4-1, QY-5, QY-4, YT-3-9, YT-4-3, HLJ-1-6, HLJ-2-9, SQ-1-2)
Fig. 2Cultivation type I of A. alternata f. sp. mali strains
第2类(图3):菌丝前期白色,后期呈绿色,多数菌株内部的绿色菌丝和外围的白色菌丝间形成明显的环状界限。具有这一类特征的菌株有13个,包括3个带毒菌株,分别为QY-2、YT-3-7、SQ-1-1,以及10个不带毒菌株,分别为SQ-3-14、QY-8、CL-4-2、SQ-3-6、CL-2-9、CL-4-9、SMX-1-3、SQ-1-6、YA、LN-9。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3苹果斑点落叶病菌菌株的培养类型 II
A—C:带毒菌株The strains taking virus (QY-2, YT-3-7, SQ-1-1);D—M:未带毒菌株The strains without virus (SQ-3-14, QY-8, CL-4-2, SQ-3-6, CL-2-9, CL-4-9, SMX-1-3, SQ-1-6, YA, LN-9)
Fig. 3Cultivation type II of A. alternata f. sp. mali strains
2.3 苹果斑点落叶病菌带毒菌株菌丝的生长速率
苹果斑点落叶病菌不同菌株的菌丝生长速率测定结果如图4所示,带毒菌株与非带毒菌株相关性差异不明显,各菌株间生长速度的快慢因菌株而异。图4
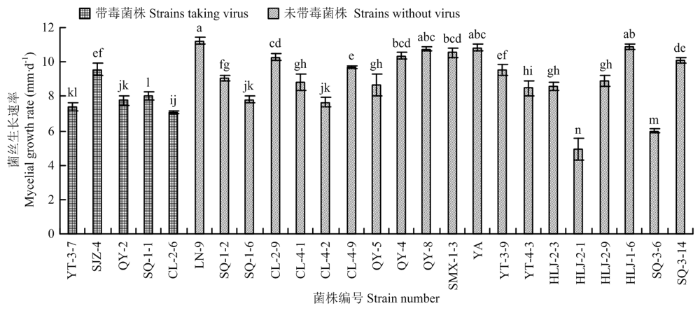 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4苹果斑点落叶病菌不同菌株的菌丝生长速率
Fig. 4The mycelial growth rate of different A. alternata f. sp. mali strains
2.4 苹果斑点落叶病菌带毒菌株的致病力
将带毒菌株和20个不带毒菌株接种海棠叶片,测定各菌株的致病力。测量接种第7天的病斑大小(图5),发现在5个带毒菌株中QY-2和YT-3-7的致病力较弱,其余菌株致病力中等。在20个不带毒的菌株中LN-9和CL-2-9的致病力较强,其他菌株的致病力中等(图6)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5苹果斑点落叶病菌不同菌株在海棠叶片及苹果果实上的病斑大小
A:CK;B—F:接种带毒菌株Inoculation of strains taking virus (YT-3-7, SJZ-4, QY-2, SQ-1-1, CL-2-6);G—Z:接种不带毒菌株Inoculation of strains without virus (LN-9, SQ-1-2, SQ-1-6, CL-2-9, CL-4-1, CL-4-2, CL-4-9, QY-5, QY-4, QY-8, SMX-1-3, YA, YT-3-9, YT-4-3, HLJ-2-3, HLJ-2-1, HLJ-2-9, HLJ-1-6, SQ-3-6, SQ-3-14)
Fig. 5Spots size of different strains of A. alternata f. sp. mali on cherry-apple leaves and apple fruits
图6
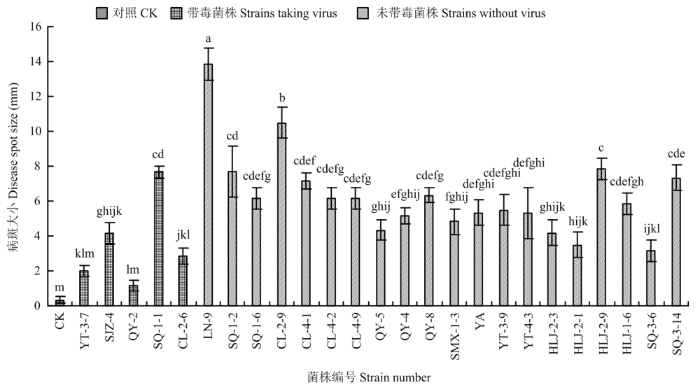 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6苹果斑点落叶病菌不同菌株在海棠叶片上的致病力
Fig. 6Pathogenicity of different strains of A. alternata f. sp. mali on cherry-apple leaves
将带毒菌株和20个不带毒菌株接种富士苹果,测量第7天的病斑大小,发现在5个带毒菌株中QY-2为弱致病力菌株,SJZ-4的致病力较强,其余菌株致病力中等。在20个不带毒的菌株中LN-9、CL-2-9、CL-4-9、YA、YT-3-9、HLJ-2-9、HLJ-1-6、SQ-3-14的致病力较强,其他菌株致病力中等(图7)。
图7
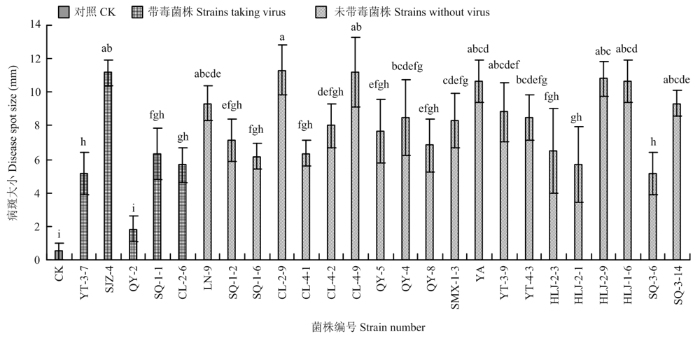 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7苹果斑点落叶病菌不同菌株在苹果果实上的致病力
Fig. 7Pathogenicity of different strains of A. alternata f. sp. mali on apple fruits
带毒菌株QY-2在叶片和果实上的致病力均较弱,从寻找具有弱毒特性的菌株方面考虑,该菌株可能具有潜在的应用价值,选为进一步研究的对象。
2.5 苹果斑点落叶病菌菌株QY-2中dsRNA病毒的鉴定
2.5.1 病毒的全基因组序列 通过分析QY-2菌株中dsRNA高通量测序结果,获得了4条与真菌病毒相关的序列,经过Blast表明4个Contig均与来源于链格孢日本梨致病型的AaCV1序列有较高的相似性,以AaCV1的4个dsRNA片段序列为参考序列,有1 451 570、2 331 542、1 167 609、1 268 863 个reads与AaCV1 dsRNA片段序列相符,经比对和拼接获得了Contig 10、Contig 41、Contig 38、Contig 24,对reads覆盖度进行表达量计算,4个Contig的RPKM值在所有基因中是最高的,达到了4 820—9 292。E-value数值很小,说明了结果的可靠性(表3)。Table 3
表3
表3高通量测序苹果斑点落叶病菌菌株QY-2携带病毒信息
Table 3
| 基因ID Unigene ID | 基因长度 Unigene length (nt) | 蛋白描述 Protein description | 登录号 Accession number | E-value | Reads数量 Number of Reads | RPKM |
|---|---|---|---|---|---|---|
| Contig 10 | 3665 | RNA-dependent RNA polymerase [AaCV1] | BBC27878.1 | 0.00E+00 | 1451570 | 4820 |
| Contig 41 | 3054 | Putative coat protein [AaCV1] | BBC27881.1 | 3.5E-176 | 2331542 | 9292 |
| Contig 38 | 2824 | Hypothetical protein [AaCV1] | BBC27880.1 | 2.9E-108 | 1167609 | 5032 |
| Contig 24 | 2819 | Hypothetical protein [AaCV1] | BBC27879.1 | 2.3E-137 | 1268863 | 5478 |
新窗口打开|下载CSV
已报道的AaCV1基因组具有5条dsRNA片段,而高通量测序结果只获得了病毒的4条dsRNA序列,QY-2 dsRNA琼脂糖凝胶电泳(图1)显示在800 bp左右处也存在一条dsRNA,根据已报道的AaCV1设计其第5个片段的特异性引物CRV1-5进行分子克隆测序,最终获得了菌株QY-2 dsRNA病毒的第5个片段序列。综上,菌株QY-2 dsRNA病毒包括5个dsRNA片段,分别为dsRNA1—dsRNA5,片段大小依次为3 665、3 054、2 824、2 819、831 nt。5条dsRNA序列已全部提交到GenBank,登录号分别为MK672910、MK672913、MK672912、MK672911、MK836314。
2.5.2 病毒的基因组结构分析 将dsRNA1—dsRNA5的序列在NCBI网站上进行ORF分析,结果显示dsRNA1—dsRNA5分别编码一个推定的ORF,依次为ORF1—ORF5。 ORF1—ORF5依次含有1 117、774、774、769、115 aa,编码蛋白的分子量依次为124、83、84、83、13 kD。dsRNA1—dsRNA5的5′端非翻译区(UTR)大小分别为205、573、357、401、102 nt;3′ UTR大小分别为107、157、133、109、382 nt(图8)。
图8
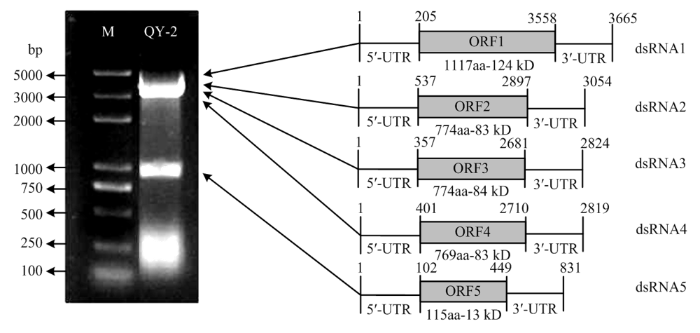 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8苹果斑点落叶病菌菌株QY-2 dsRNA病毒基因组模式图
Fig. 8A schematic diagram of the genomic organization of A. alternata f. sp. mali QY-2 dsRNA virus
2.5.3 病毒开放阅读框序列相似性分析 将QY-2菌株dsRNA病毒ORF1—ORF5编码的氨基酸序列在NCBI上进行blastp搜索,结果显示与其相似性相对较高的病毒有4个,分别是AaCV1、Botryosphaeria dothidea chrysovirus 1(BdCV1)、Asperqillus themomutatus chrysovirus 1(AtCV1)、Penicillium janczewskill chrysovirus 2(PjCV2)(表4)。与QY-2菌株dsRNA病毒ORF1(RdRP)氨基酸序列相似性分别为93%、56%、55%、52%,核苷酸序列相似性为77%、59%、57%、54%;ORF2氨基酸序列相似性为91%、47%、44%、40%,核苷酸序列相似性为75%、54%、50%、49%;ORF3氨基酸序列相似性为80%、35%、32%、32%,核苷酸序列相似性为70%、40%、42%、42%;ORF4氨基酸序列相似性为90%、47%、43%、43%,核苷酸序列相似性为76%、51%、50%、56%;与ORF5氨基酸序列相似性较高的只有AaCV1编码的假定蛋白,相似性为87%,核苷酸序列相似性为95%。根据国际病毒分类委员会(ICTV)2020年发布的Chrysoviridae内种的分类标准(
Table 4
表4
表4苹果斑点落叶病菌菌株QY-2的dsRNA与其他病毒的核苷酸及氨基酸序列的相似性
Table 4
| 名称 Name | ORF1 | ORF2 | ORF3 | ORF4 | ORF5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| nt (%) | aa (%) | nt (%) | aa (%) | nt (%) | aa (%) | nt (%) | aa (%) | nt (%) | aa (%) | |
| AaCV1 | 77 | 93 | 75 | 91 | 70 | 80 | 76 | 90 | 95 | 87 |
| BdCV1 | 59 | 56 | 54 | 47 | 40 | 35 | 51 | 47 | ||
| AtCV1 | 57 | 55 | 50 | 44 | 42 | 32 | 50 | 43 | ||
| PjCV2 | 54 | 52 | 49 | 40 | 42 | 32 | 56 | 43 | ||
新窗口打开|下载CSV
2.5.4 病毒系统进化关系分析 为分析病毒系统进化关系,将苹果斑点落叶病菌菌株QY-2携带的AaCV2 RdRP氨基酸序列与Chrysoviridae、Totiviridae以及一些未分类病毒的RdRP氨基酸序列构建系统发育树。结果表明,菌株QY-2携带的病毒与Chrysoviridae的Chrysovirus聚在一个分支,并且与病毒AaCV1遗传关系最近,其自展支撑值为100(图9)。因此将QY-2菌株携带的病毒暂定命名为Alternaria alternata chrysovirus 2(AaCV2)。
图9
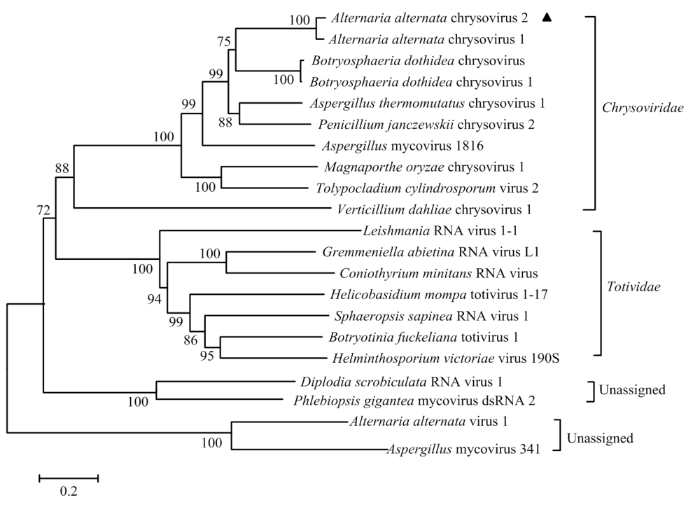 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9以真菌病毒的RdRP氨基酸序列为基础构建的系统发育树
Fig. 9Phylogenetic tree based on RNA-dependent RNA polymerase amino acid sequence of mycovirus
2.6 菌株QY-2携带病毒的RT-PCR鉴定
根据获得的QY-2 dsRNA病毒基因序列,设计特异性引物CRV1-1对其进行RT-PCR检测。琼脂糖凝胶电泳显示,菌株QY-2出现了453 bp的病毒特异性目的条带,对照菌株及清水对照均没有条带出现,进一步说明了菌株QY-2中AaCV2的存在(图10)。图10
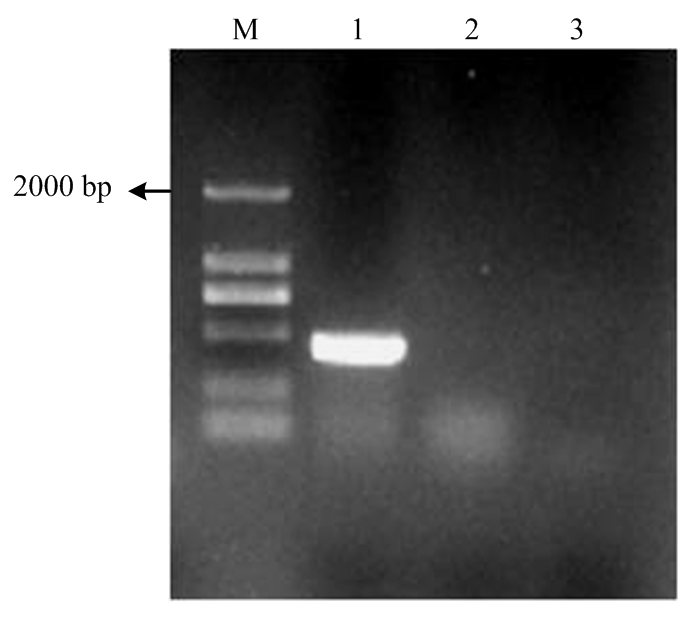 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10苹果斑点落叶病菌菌株QY-2中AaCV2 RT-PCR鉴定
M:DL2000;1:QY-2;2:对照菌株Control strain;3:清水对照Water control
Fig. 10Identification of AaCV2 in A. alternata f. sp. mali strain QY-2 by RT-PCR
3 讨论
目前,苹果斑点落叶病防治的主要方式为化学防治。为避免化学防治对人体和环境产生的不利影响,寻求安全有效的绿色生防措施颇为重要。部分真菌病毒可导致真菌致病力的衰退,因此可作为一种生防因子用于真菌病害的防治[25,26]。本研究对采自我国8个省份苹果主产区的102株苹果斑点落叶病菌菌株进行dsRNA检测,根据琼脂糖凝胶电泳结果,明确了其中5个菌株具有明显的dsRNA条带,可能携带真菌病毒,约占5%的菌比例,在苹果斑点落叶病菌中dsRNA具有一定的普遍性,且根据条带大小来看,病毒种类具有多样性。据报道链格孢属真菌目前发现的病毒包括:从棉花种子中分离出6个携带真菌病毒的链格孢菌株,dsRNA片段在1.0—5.1 kb [14];从茄链格孢中检测到两条dsRNA大小分别为8.3和5.5 kb[15];另外还有真菌病毒AaV1(dsRNA片段:3 617、2 794、2 576、1 420 bp)[17]、AlRV1(dsRNA片段:3 145 bp)[18]、AaVV1(dsRNA片段:5 203 bp)[19]、AbFV1(ssRNA片段:6 639 bp)[20]、ABRV1(dsRNA片段:6 188、5 903 bp)[21]、AaCV1(dsRNA片段:3 647、2 857、2 785、2 772、836 bp)[22]。不同的链格孢菌株携带dsRNA条带差异较大,且与本研究所分离到的dsRNA病毒条带有较大差异。因此,本研究发现的苹果斑点落叶病菌真菌病毒有可能是未揭示的新病毒。
随着真菌病毒种类及数量不断增多,人们开始研究病毒对真菌生物学功能方面的影响,发现大多数病毒在菌株中能够稳定遗传[27,28],但因病毒种类不同,对真菌致病力及其他方面会有不同的影响。真菌病毒与寄主间的相互作用成为现代病毒学研究的热点领域。本研究主要针对携带dsRNA病毒的5个菌株进行了形态学观察、生长速率测定及致病力鉴定,同时与不携带dsRNA病毒的其他菌株进行比较,以明确携带病毒是否引起了菌株的特异性或其是否具有生防意义。通过测定发现,带毒菌株间菌落形态、菌丝的生长速率和菌株的致病能力存在显著差异,不带毒菌株间这3个方面同样也存在显著差异。菌株的菌落特征、生长速率与dsRNA携带与否及类型没有明显的关系,致病力测定表明含有dsRNA病毒的供试菌株大多属于低毒力类型,其中的线性关系或多因素关系,还有待进一步研究。
在含有dsRNA病毒的低致病力菌株中,QY-2致病力最低,与清水对照无显著性差异。经鉴定该菌株中携带大量的产黄青霉病毒科、β产黄青霉病毒属的Alternaria alternata chrysovirus,这是苹果斑点落叶病菌中该病毒的首次报道。王利华等[29]明确了产黄青霉病毒科成员Botryosphaeria dothidea Chrysovirus 1(BdCV1)对梨轮纹病菌(B. dothidea)的菌落形态、生长速度有明显抑制作用,是引起梨轮纹病菌致病力衰退的主要因子;MA等[30]报道细链格孢(A. tenuissima)携带AaCV1-AT1,AaCV1-AT1的感染降低了宿主真菌的菌落生长速度和产孢能力;OKADA等[22]报道了AaCV1对链格孢日本梨致病型具有两个方面的作用,一是影响真菌宿主的生长速度,二是影响真菌宿主毒素的产生。因此,AaCV2也可能是引起QY-2菌株致病力衰退的主要原因。下一步将对QY-2携带的AaCV2进行深入研究,以开发对苹果斑点落叶病具有生防潜力的dsRNA病毒。
4 结论
我国苹果斑点落叶病菌菌株携带dsRNA的概率在5%左右,dsRNA群体多样,有4种类型:类型I的dsRNA在8、2.5和1.5 kb左右;类型II的dsRNA在8 kb左右;类型III的dsRNA在3和0.8 kb左右;类型IV的dsRNA在2.5和1.5 kb左右。dsRNA的有无及类型与寄主病原菌培养性状没有明显相关性,携带dsRNA的菌株多为弱致病力,致病力最低的QY-2菌株携带产黄青霉病毒科、β产黄青霉病毒属的Alternaria alternata chrysovirus 2。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/BF02931111URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1094/PHYTO-99-4-0369URL [本文引用: 1]
DOI:10.21273/JASHS.136.2.109URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.cropro.2006.04.007URL [本文引用: 1]
DOI:10.1126/science.215.4532.466URL [本文引用: 1]
DOI:10.1128/mr.56.4.561-576.1992URL [本文引用: 1]
DOI:10.1038/196962a0URL [本文引用: 1]
DOI:10.1007/s00705-008-0272-xURL [本文引用: 1]
DOI:10.1007/s00705-009-0507-5URL [本文引用: 1]
DOI:10.1128/jvi.62.10.3888-3891.1988URL [本文引用: 2]
DOI:10.1017/S0953756296003097URL [本文引用: 2]
DOI:10.3186/jjphytopath.54.250URL [本文引用: 1]
[本文引用: 2]
DOI:10.1007/s00705-014-2218-9URL [本文引用: 2]
DOI:10.1007/s00705-016-2796-9URL [本文引用: 2]
DOI:10.1016/j.virusres.2015.11.012URL [本文引用: 2]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
DOI:10.1016/j.virol.2018.03.027URL [本文引用: 3]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1128/JVI.01830-09URL [本文引用: 1]
[本文引用: 1]
DOI:10.1080/07060669709500576URL [本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.virusres.2020.197904URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}