Cloning, Expression and Anti-Virus Function Analysis of Solanum lycopersicum SlN-like
LIU ChangYun,1, LI XinYu1, TIAN ShaoRui1, WANG Jing1, PEI YueHong1, MA XiaoZhou1,2, FAN GuangJin1, WANG DaiBin,3, SUN XianChao,11College of Plant Protection, Southwest University, Chongqing 400715 2Key Laboratory of Horticulture Science for Southern Mountainous Regions of Ministry of Education, College of Horticulture and Landscape Architecture, Southwest University, Chongqing 400715 3Chongqing Tobacco Science Research Institute, Chongqing 400715
Abstract 【Objective】As an important vegetable crop, tomato (Solanum lycopersicum) is endangered by various biological factors including pests, fungi, bacteria and viruses. The objective of this study is to clarify the antiviral function and mechanism of S. lycopersicum resistance gene SlN-like, and to provide a theoretical basis for the genetic breeding of antiviral S. lycopersicum and the targeted development of the antiviral agents. 【Method】The full length of SlN-like was obtained from the Solanaceae Genomics Network database and was divided into four segments, fusion PCR was used to amplify the full length of sequence. Bioinformatics was used to analyze the evolutionary relationship, protein characteristics, conserved domains, subcellular location and interaction relationship of SlN-like. Real-time fluorescent quantitative PCR was used to analyze the SlN-like expression in S. lycopersicum roots, stems, leaves, flowers and fruits and its response after tobacco mosaic virus (TMV) infection. S. lycopersicum endogenous SlN-like was silenced using tobacco rattle virus (TRV)-mediated gene silencing technology, and the silent plants were inoculated with TMV-GFP to clarify the influence of SlN-like on virus infection. The expressions of abscisic acid (ABA), jasmonic acid (JA) and ethylene (ET) hormone-related genes in silenced plants, and the expression of SlN-like after application of ethephon (ETH) for 3, 6, 12 and 24 h were analyzed by real-time fluorescence quantitative PCR to investigate the mechanism of SlN-like regulatory hormone pathway in response to virus infection. 【Result】Through molecular cloning and fusion PCR technology, a 3 444 bp SlN-like was cloned from S. lycopersicum variety Micro-Tom, and uploaded to NCBI to obtain the sequence number MW792493. Through bioinformatics analysis, it was found that SlN-like contains TIR, NB-ARC and NACHT domains, and is closely related to Solanum tuberosum N-like (AAP44394.1). SlN-like expressed in all tissues of S. lycopersicum, with the highest expression in stems, followed by roots, flowers, leaves and fruits. After TMV-GFP infection S. lycopersicum at 5th and 7th day, the SlN-like expression level was higher than that of PBS treatment, and TMV-GFP infection would cause the expression of SlN-like to increase continuously. TRV vector induced silencing of SlN-like in S. lycopersicum, and it was found that silencing 78.3% of SlN-like did not affect tomato growth phenotype, but silencing SlN-like promoted the infection of TMV-GFP. Real-time fluorescent quantitative PCR analysis found that the expression of ERF1 in SlN-like-silent plants was significantly reduced, only 12.5% of that in the control group. The expression of SlN-like increased after 3 h of external application of ethephon, and reached the highest peak at 12 h, which was 2.71 times that of the control group, and returned to normal at 24 h. 【Conclusion】S. lycopersicum SlN-like belongs to the NBS-LRR disease-resistant protein family, its expression is induced by TMV infection. Silencing SlN-like can promote TMV-GFP infection and reduce the expression of ethylene-related gene ERF1, while external application of ethephon resulted in the differential expression of SlN-like, revealing that SlN-like participates in S. lycopersicum antiviral defense through the ethylene pathway. Keywords:Solanum lycopersicum;SlN-like;tobacco mosaic virus (TMV);gene expression;ethylene
PDF (1197KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 刘昌云, 李欣羽, 田绍锐, 王靖, 裴悦宏, 马小舟, 樊光进, 汪代斌, 孙现超. 番茄SlN-like的克隆、表达与抗病毒功能. 中国农业科学, 2021, 54(20): 4348-4357 doi:10.3864/j.issn.0578-1752.2021.20.009 LIU ChangYun, LI XinYu, TIAN ShaoRui, WANG Jing, PEI YueHong, MA XiaoZhou, FAN GuangJin, WANG DaiBin, SUN XianChao. Cloning, Expression and Anti-Virus Function Analysis of Solanum lycopersicum SlN-like. Scientia Acricultura Sinica, 2021, 54(20): 4348-4357 doi:10.3864/j.issn.0578-1752.2021.20.009
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】番茄(Solanum lycopersicum)是茄科番茄属的一种一年生或多年生草本植物,在农业种植方面有着重要的地位。生产过程中,番茄会遭受高温、干旱等非生物胁迫或病毒、真菌等生物胁迫,造成生物组织的损伤,进而引起减产。其中由烟草花叶病毒(tobacco mosaic virus,TMV)引起的番茄病毒病是番茄生产过程的重要病害之一。挖掘并研究番茄抗病毒基因及其抗病毒机制,对番茄抗病毒育种及利用药剂诱导调控抗病毒基因防控病毒病具有重要意义。【前人研究进展】N是最早发现的抗TMV基因,烟草N可对包括TMV、番茄花叶病毒(tomato mosaic virus,ToMV)在内的绝大多数烟草花叶病毒组成员产生抗性,属TIR-NBS-LRR类植物抗性基因家族中的一员[1,2]。早期研究发现,TMV复制酶126 kD能够引起N介导的超敏反应(hypersensitivity,HR)[3],而位于复制酶126 kD羧基端末端的约50 kD的解旋酶片段(p50)能够导致N转录产物的积累,并有效地引起N介导的HR反应[4,5],因此编码p50的核苷酸序列又被称为N对应的无毒基因(avirulence,Avr)。TMV侵染引起心叶烟(Nicotiana glutinosa)、三生烟(Nicotiana tabacum var. Samsun NN)等抗病品种坏死斑的形成是植物体自身防御病原微生物侵入所形成的细胞程序性死亡(programmed cell death,PCD)[6]。目前N介导的TMV识别过程及其对下游抗病基因的诱导已经研究得非常透彻[7]。NRIP1(N receptor- interacting protein 1)是一种同时与N蛋白的TIR结构域和p50产生相互作用的硫氰酸酶(硫转移酶),NRIP1定位于叶绿体中。SPL6(squamosa promoter binding protein-like 6)是一种与N蛋白在核小体内互作的转录因子。在TMV未侵染的细胞中,N蛋白处于活性被抑制的状态,其表达极低,并且分布在细胞核和细胞质中[8]。此时核内的N蛋白不与SPL6结合。当TMV通过机械损伤进入植物体内后,病毒在细胞质中脱壳、复制、转录和翻译,p50激发子引起了NRIP1的重新定位。NRIP1与p50结合后,被细胞质中的N蛋白识别,三者形成NRIP1-p50-N复合体[9]。随着三者的结合,p50引起了N蛋白构象上的变化,而该变化可能需要ATP结合或水解。结合ATP的N蛋白进入细胞核,与SPL6互作进而引起SPL6的转录激活功能,激活了下游抗性基因的表达[10]。此外,NRIP1-p50-N形成复合体的同时,N蛋白利用其LRR结构域与p50形成次级结合,释放出TIR-NBS区段以提高核酸结合能力,促成N蛋白的聚合。聚合后的N蛋白进入核内,与SPL6结合,激活并引起下游抗性基因的表达[11]。【本研究切入点】N-like是N的同源基因,同样含有TIR-NBS-LRR结构域,其TIR-NBS保守结构域区段同样可与TMV p50产生HR反应,而LRR并不能和p50产生HR反应[12],因此推测这可能与N-like与TMV的识别有关[13],并且可能行驶N的第一种调控模式。但番茄中N-like如何响应病毒侵染,如何调控寄主免疫防御知之甚少[14,15]。【拟解决的关键问题】通过克隆番茄抗性基因SlN-like的cDNA全长,并对核酸序列和蛋白质序列进行生物信息学分析,利用实时荧光定量PCR技术明确SlN-like的表达模式,通过病毒介导的基因沉默(virus-induced gene silencing,VIGS)分析SlN-like对TMV-GFP侵染的影响,确定SlN-like的抗病功能,并明确SlN-like可能影响的激素通路,为解析SlN-like在番茄抗病应答中的调控机制提供科学依据,同时为番茄抗病品种的选育和抗病毒药剂的靶向开发提供理论依据。
A:SlN-like的组织表达,统计分析采用ANOVA(LSD检测,P<0.05)Tissue expression of SlN-like, the statistical analyses were performed using One-way ANOVA (LSD’s test, P<0.05);B:TMV侵染后SlN-like的表达。统计分析采用Student’s t检验(**0.001<P<0.01,***P<0.001),每个处理进行3次生物学重复,每次生物学重复3株番茄,数值代表3次生物学重复的平均值±标准误SlN-like expression after TMV infection. The statistical analyses were performed using Student’s t-test (**0.001<P<0.01, ***P<0.001). The experiments were repeated three times with three plants each time. Values represent means±SE from three biological replications Fig. 2Expression analysis of SlN-like
A:SlN-like沉默后的番茄表型Tomato phenotype after SlN-like silenced;B:TRV:SlN-like的沉默效率检测。统计分析采用Student’s t检验 (***P<0.001),每个处理进行3次生物学重复,每次生物学重复10株番茄,数值代表3次生物学重复的平均值±标准误 Silencing efficiency detection in TRV:SlN-like. The statistical analyses were performed using Student’s t-test (***P<0.001). The experiments were repeated three times with ten plants each time. Values represent means±SE from three biological replications Fig. 3Phenotype and silencing efficiency detection in SlN-like silenced plant
A:TRV:SlN-like和TRV:00接种TMV-GFP后的症状图Symptoms after inoculation with TMV-GFP in TRV:SlN-like and TRV:00;B:TRV:SlN-like和TRV:00中TMV MP含量检测。统计分析采用Student’s t检验(**0.001<P<0.01,***P<0.001),每个处理进行3次生物学重复,每次生物学重复10株番茄,数值代表3次生物学重复的平均值±标准误 Detection of TMV MP content in TRV:SlN-like and TRV:00. The statistical analyses were performed using Student’s t-test (**0.001<P<0.01, ***P<0.001). The experiments were repeated three times with ten plants each time. Values represent means±SE from three biological replications Fig. 4Silencing of SlN-like promotes TMV-GFP infection
统计分析采用Student’s t检验(**0.001<P<0.01,***P<0.001),每个处理进行3次生物学重复,每次生物学重复3株番茄,数值代表3次生物学重复的平均值±标准误 The statistical analyses were performed using Student’s t-test (**0.001<P<0.01, ***P<0.001). The experiments were repeated three times with three plants each time. Values represent means±SE from three biological replications。图6同The same as Fig. 6 Fig. 5Hormone-related gene expression in SlN-like silenced plant
MARATHER, ANANDALAKSHMIR, LIUY, DINESH-KUMARS P. The tobacco mosaic virus resistance gene, N Molecular Plant Pathology, 2002, 3(3):167-172. DOI:10.1046/j.1364-3703.2002.00110.xURL [本文引用: 1]
PADGETTH S, BEACHYR N. Analysis of a tobacco mosaic virus strain capable of overcoming N gene-mediated resistance The Plant Cell, 1993, 5(5):577-586. [本文引用: 1]
WANGQ, LIUG S. The tobacco N gene and the signal transduction of N-mediated TMV resistance Chinese Tobacco Science, 2016, 37(3):93-99. (in Chinese) [本文引用: 1]
WHITHAMS, MCCORMICKS, BAKERB. The N gene tobacco confers resistance to tobacco mosaic virus in transgenic tomato Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(16):8776-8781. [本文引用: 1]
ERICKSONF L, HOLZBERGS, CALDERÓN-URREAA, HANDLEYV, AXTELLM, CORRC, BAKERB. The helicase domain of the TMV replicase proteins induces the N-mediated defence response in tobacco The Plant Journal, 1999, 18(1):67-75. DOI:10.1046/j.1365-313X.1999.00426.xURL [本文引用: 1]
GREENBERGJ T. Programmed cell death in plant-pathogen interactions Annual Review of Plant Physiology and Plant Molecular Biology, 1997, 48:525-545. DOI:10.1146/arplant.1997.48.issue-1URL [本文引用: 1]
DINESH-KUMARS P, THAMW H, BAKERB J. Structure-function analysis of the tobacco mosaic virus resistance gene N Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(26):14789-14794. [本文引用: 1]
WHITHAMS, DINESH-KUMARS P, CHOID, HEHLR, CORRC, BAKERB. The product of the tobacco mosaic virus resistance gene N: Similarity to Toll and the interleukin-1 receptor Cell, 1994, 78(6):1101-1115. DOI:10.1016/0092-8674(94)90283-6URL [本文引用: 2]
DINESH-KUMARS P, WHITHAMS, CHOID, HEHLR, CORRC, BAKERB. Transposon tagging of tobacco mosaic virus resistance gene N: Its possible role in the TMV-N-mediated signal transduction pathway Proceedings of the National Academy of Sciences of the United States of America, 1995, 92(10):4175-4180. [本文引用: 1]
ZHANGH M, CHENJ P, CHENGY. Study on the resistant gene N of tobacco to tobacco mosaic virus (TMV) Acta Agriculturae Zhejiangensis, 2001, 13(2):55-60. (in Chinese) [本文引用: 1]
PADMANABHANM S, MAS, BURCH-SMITHT M, CZYMMEKK, HUIJSERP, DINESH-KUMARS P. Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor- mediated plant innate immunity PLoS Pathogens, 2013, 9(3):e1003235. DOI:10.1371/journal.ppat.1003235URL [本文引用: 1]
GAOJ S, SASAKIN, KANEGAEH, KONAGAYAK I, TAKIZAWAK, HAYASHIN, OKANOY, KASAHARAM, MATSUSHITAY, NYUNOYAH. The TIR-NBS but not LRR domains of two novel N-like proteins are functionally competent to induce the elicitor p50-dependent hypersensitive response Physiological and Molecular Plant Pathology, 2007, 71:78-87. DOI:10.1016/j.pmpp.2007.11.002URL [本文引用: 2]
ZHANGG Y, CHENM, GUOJ M, XUT W, LIL C, XUZ S, MAY Z, CHENX P. Isolation and characteristics of the CN gene, a tobacco mosaic virus resistance N gene homolog, from tobacco Biochemical Genetics, 2009, 47(3/4):301-314. DOI:10.1007/s10528-009-9229-3URL [本文引用: 1]
XIEP. Studies on hypersensitive response mediated by TMV resistance gene N and cloning of homologous gene in Nicotiana tabacum [D]. Hefei: Anhui Agricultural University, 2013. (in Chinese) [本文引用: 1]
XIEP, HUL, CAIY P, LINY, GAOJ S. Studies on hypersensitive response mediated by TMV resistance gene N in Nicotiana tabacum Journal of Nuclear Agricultural Sciences, 2013, 27(12):1809-1816. (in Chinese) [本文引用: 1]
FERNANDEZ-POZON, MENDAN, EDWARDSJ D, SAHAS, TECLEI Y, STRICKLERS R, BOMBARELYA, FISHER-YORKT, PUJARA, FOERSTERH, YANA, MUELLERL A. The sol genomics network (SGN)—From genotype to phenotype to breeding Nucleic Acids Research, 2015, 43(Database issue):D1036-D1041. DOI:10.1093/nar/gku1195URL [本文引用: 1]
CHOUK C, SHENH B. Cell-PLoc 2.0: An improved package of web-servers for predicting subcellular localization of proteins in various organisms Natural Science, 2010, 2(10):1090-1103. DOI:10.4236/ns.2010.210136URL [本文引用: 1]
LVX, XIANGS Y, WANGX C, WUL, LIUC Y, YUANM T, GONGW W, WINH, HAOC, XUEY, MAL S, CHENGD Q, SUNX C. Synthetic chloroinconazide compound exhibits highly efficient antiviral activity against tobacco mosaic virus Pest Management Science, 2020, 76(11):3636-3648. DOI:10.1002/ps.v76.11URL [本文引用: 1]
LIUC Y, PUY D, PENGH R, LVX, TIANS R, WEIX F, ZHANGJ, ZOUA H, FANG J, SUNX C. Transcriptome sequencing reveals that photoinduced gene IP-L affects the expression of PsbO to response to virus infection in Nicotiana benthamiana Physiological and Molecular Plant Pathology, 2021, 114:101613. DOI:10.1016/j.pmpp.2021.101613URL [本文引用: 1]
PANQ, LIUX X, PENGH R, PUY D, ZHANGY Z, YES H, WUG T, QINGL, SUNX C. Cloning, expression analysis of Solanum lycopersicum SYTA Scientia Agricultura Sinica, 2017, 50(15):2936-2945. (in Chinese) [本文引用: 2]
KNAPPE, LEWANDOWSKID J. Tobacco mosaic virus, not just a single component virus anymore Molecular Plant Pathology, 2001, 2(3):117-123. DOI:10.1046/j.1364-3703.2001.00064.xURL [本文引用: 1]
SCHOLTHOFK B G, ADKINSS, CZOSNEKH, PALUKAITISP, JACQUOTE, HOHNT, HOHNB, SAUNDERSK, CANDRESSET, AHLQUISTP, HEMENWAYC, FOSTERG D. Top 10 plant viruses in molecular plant pathology Molecular Plant Pathology, 2011, 12(9):938-954. DOI:10.1111/j.1364-3703.2011.00752.xURL [本文引用: 1]
DANGLJ L, JONESJ D. Plant pathogens and integrated defence responses to infection Nature, 2001, 411(6839):826-833. DOI:10.1038/35081161URL [本文引用: 1]
VANDER BIEZEN E A, JONESJ D. Plant disease-resistance proteins and the gene-for-gene concept Trends in Biochemical Sciences, 1998, 23(12):454-456. DOI:10.1016/S0968-0004(98)01311-5URL [本文引用: 1]
JIANGG, LIUD, YIND, ZHOUZ, SHIY, LIC, ZHUL, ZHAIW. A rice NBS-ARC gene conferring quantitative resistance to bacterial blight is regulated by a pathogen effector-inducible miRNA Molecular Plant, 2020, 13(12):1752-1767. DOI:10.1016/j.molp.2020.09.015URL [本文引用: 1]
HULBERTS H, WEBBC A, SMITHS M, SUNQ. Resistance gene complexes: Evolution and utilization Annual Review of Phytopathology, 2001, 39:285-312. DOI:10.1146/phyto.2001.39.issue-1URL [本文引用: 1]
LIJ, LIC Y. Seventy-year major research progress in plant hormones by Chinese scholars Scientia Sinica Vitae, 2019, 49(10):1227-1281. (in Chinese) [本文引用: 1]
左建儒, 漆小泉, 林荣呈, 钱前, 顾红雅, 陈凡, 杨淑华, 陈之端, 白永飞, 王雷, 王小菁, 姜里文, 萧浪涛, 种康, 王台. 2019年中国植物科学若干领域重要研究进展 植物学报, 2020, 55(3):257-269. DOI:10.11983/CBB20108 [本文引用: 1] 2019年中国植物科学家在国际综合性学术期刊及植物科学主流期刊发表的论文数量大幅增加, 在光生物学、植物抗逆和分子进化等若干领域取得了重要成果。其中, 硅藻光合膜蛋白超分子结构和功能研究入选2019年度中国科学十大进展和中国生命科学十大进展; 植物抗病小体的结构与功能研究入选2019年度中国生命科学十大进展。该文评述了2019年中国科学家在植物科学若干领域取得的重要研究进展, 以期追踪和报道当前中国植物科学领域发展的前沿和热点及展示中国科学家所取得的辉煌成果。 ZUOJ R, QIX Q, LINR C, QIANQ, GUH Y, CHENF, YANGS H, CHENZ D, BAIY F, WANGL, WANGX J, JIANGL W, XIAOL T, ZHONGK, WANGT. Achievements and advance in Chinese plant sciences in 2019 Chinese Bulletin of Botany, 2020, 55(3):257-269. (in Chinese) DOI:10.11983/CBB20108 [本文引用: 1] 2019年中国植物科学家在国际综合性学术期刊及植物科学主流期刊发表的论文数量大幅增加, 在光生物学、植物抗逆和分子进化等若干领域取得了重要成果。其中, 硅藻光合膜蛋白超分子结构和功能研究入选2019年度中国科学十大进展和中国生命科学十大进展; 植物抗病小体的结构与功能研究入选2019年度中国生命科学十大进展。该文评述了2019年中国科学家在植物科学若干领域取得的重要研究进展, 以期追踪和报道当前中国植物科学领域发展的前沿和热点及展示中国科学家所取得的辉煌成果。
ZHAOH, YINC, MAB, CHENS Y, ZHANGJ S. Ethylene signaling in rice and Arabidopsis: New regulators and mechanisms Journal of Integrative Plant Biology, 2021, 63(1):102-125. DOI:10.1111/jipb.v63.1URL [本文引用: 1]
DINGY, MURPHYK M, PORETSKYE, MAFUS, YANGB, CHARS N, CHRISTENSENS A, SALDIVARE, WUM, WANGQ, et al. Multiple genes recruited from hormone pathways partition maize diterpenoid defences Nature Plants, 2019, 5(10):1043-1056. DOI:10.1038/s41477-019-0509-6URL [本文引用: 1]
YANGD L, YANGY, HEZ. Roles of plant hormones and their interplay in rice immunity Molecular Plant, 2013, 6(3):675-685. DOI:10.1093/mp/sst056URL [本文引用: 1]
AERTSN, PEREIRAMENDES M, VANWEES S C. Multiple levels of crosstalk in hormone networks regulating plant defense The Plant Journal, 2021, 105(2):489-504. DOI:10.1111/tpj.v105.2URL [本文引用: 1]
ZHAOZ X, FENGQ, LIUP Q, HEX R, ZHAOJ H, XUY J, ZHANGL L, HUANGY Y, ZHAOJ Q, FANJ, LIY, XIAOS Y, WANGW M. RPW8.1 enhances the ethylene-signaling pathway to feedback-attenuate its mediated cell death and disease resistance in Arabidopsis New Phytologist, 2021, 229(1):516-531. DOI:10.1111/nph.v229.1URL [本文引用: 1]
TANGD, CHRISTIANSENK M, INNESR W. Regulation of plant disease resistance, stress responses, cell death, and ethylene signaling in Arabidopsis by the EDR1 protein kinase Plant Physiology, 2005, 138(2):1018-1026. DOI:10.1104/pp.105.060400URL [本文引用: 1]
ALAZEMM, LINN S. Roles of plant hormones in the regulation of host-virus interactions Molecular Plant Pathology, 2015, 16(5):529-540. DOI:10.1111/mpp.2015.16.issue-5URL [本文引用: 1]
GERIC, LOVEA J, CECCHINIE, BARRETTS J, LAIRDJ, COVEYS N, MILNERJ J. Arabidopsis mutants that suppress the phenotype induced by transgene-mediated expression of cauliflower mosaic virus (CaMV) gene VI are less susceptible to CaMV-infection and show reduced ethylene sensitivity Plant Molecular Biology, 2004, 56(1):111-124. DOI:10.1007/s11103-004-2649-xURL [本文引用: 1]
WANGS, HANK, PENGJ, ZHAOJ, JIANGL, LUY, ZHENGH, LINL, CHENJ, YANF. NbALD1 mediates resistance to turnip mosaic virus by regulating the accumulation of salicylic acid and the ethylene pathway in Nicotiana benthamiana Molecular Plant Pathology, 2019, 20(7):990-1004. DOI:10.1111/mpp.2019.20.issue-7URL [本文引用: 1]
SANSREGRETR, DUFOURV, LANGLOISM, DAAYFF, DUNOYERP, VOINNETO, BOUARABK. Extreme resistance as a host counter-counter defense against viral suppression of RNA silencing PLoS Pathogens, 2013, 9(6):e1003435. DOI:10.1371/journal.ppat.1003435URL [本文引用: 1]
PENGH R, PUY D, ZHANGY Z, XUEY, WUG T, QINGL, SUNX C. Subcellular localization and expression analyses of IP-L protein interacting with ToMV coat protein Scientia Agricultura Sinica, 2017, 50(17):3344-3351. (in Chinese) [本文引用: 1]
 ,1, 李欣羽1, 田绍锐1, 王靖1, 裴悦宏1, 马小舟1,2, 樊光进1, 汪代斌
,1, 李欣羽1, 田绍锐1, 王靖1, 裴悦宏1, 马小舟1,2, 樊光进1, 汪代斌

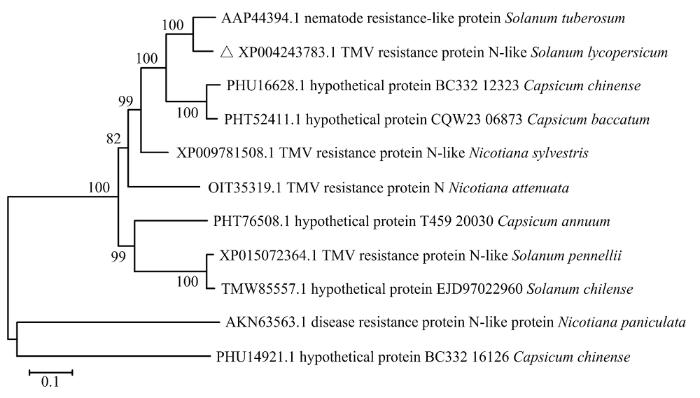 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT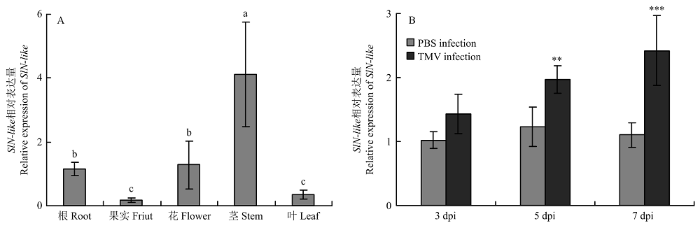 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}