 ,1,2, 时晓磊2, 张金波2, 耿洪伟3, 肖菁2, 路子峰2, 倪中福1, 丛花,2
,1,2, 时晓磊2, 张金波2, 耿洪伟3, 肖菁2, 路子峰2, 倪中福1, 丛花,2Genome-Wide Association Study of Grain Quality Related Characteristics of Spring Wheat
YAN YongLiang,1,2, SHI XiaoLei2, ZHANG JinBo2, GENG HongWei3, XIAO Jing2, LU ZiFeng2, NI ZhongFu1, CONG Hua,2通讯作者:
责任编辑: 李莉
收稿日期:2021-02-1接受日期:2021-03-17
| 基金资助: |
Received:2021-02-1Accepted:2021-03-17
作者简介 About authors
严勇亮,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (702KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
严勇亮, 时晓磊, 张金波, 耿洪伟, 肖菁, 路子峰, 倪中福, 丛花. 春小麦籽粒主要品质性状的全基因组关联分析. 中国农业科学, 2021, 54(19): 4033-4047 doi:10.3864/j.issn.0578-1752.2021.19.001
YAN YongLiang, SHI XiaoLei, ZHANG JinBo, GENG HongWei, XIAO Jing, LU ZiFeng, NI ZhongFu, CONG Hua.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】小麦是全球种植面积最广、总产量和营养价值最高、加工食品种类最多的粮食作物[1,2,3,4]。随着中国经济的发展和人们生活水平的不断提高,消费者对面制品、保健食品和营养食品的需求不断增加,并且对小麦相关产品的品质要求也越来越高。在供给侧改革需求和产业内循环的新时代要求下,在加工品质研究的基础上,营养和健康品质已成为小麦品质改良的重要方向和重要育种目标[5]。蛋白质含量、湿面筋含量、淀粉含量和沉降值等都是重要的小麦品质性状。其中,蛋白质含量和湿面筋含量是小麦品质分级的重要指标;淀粉含量对面粉品质和加工品质具有重要意义[6,7,8],研究此类性状关键基因的挖掘及遗传机理对于提高小麦品质的遗传改良具有重要的意义。【前人研究进展】小麦籽粒品质性状是由多基因控制的复杂的数量性状,关二旗等[9]通过研究小麦籽粒品质与基因型及环境条件的关系,发现小麦籽粒品质主要受基因型控制,但也受环境条件的影响。在环境因素中,气温和降水量的变化对小麦籽粒品质影响尤为明显。信志红等[10]研究了小麦籽粒品质性状对气象因子的响应,结果表明,小麦籽粒品质主要受基因型控制,其中,淀粉含量和蛋白质含量受环境影响最小。全基因组关联分析(genome-wide association study,GWAS)和数量性状位点(quantitative trait locus,QTL)定位技术的不断发展为小麦籽粒品质性状的分子检测和相关数量性状的研究提供了有力支撑[6, 11]。较QTL定位来说,用于GWAS的群体构建耗时短,群体遗传背景较为丰富,更容易发现重要的数量性状基因位点,挖掘出更多的功能基因,在小麦数量性状研究方面的应用越来越广泛[12]。LI等[13]、ECHEVENY-SOLARTE等[14]、吴云鹏等[15]、郭利建等[6]、黄梦豪等[16]利用RIL群体结合SSR标记、DArT标记、SNP芯片等,在1A、1B、1D、2A、2B、2D、3A、3B、3D、4B、5A、5B、6A、6B、7B和7D染色体上共定位到31个与小麦籽粒蛋白质含量相关的QTL,表型贡献率为0.7%—16.9%。LI等[13]、郭利建等[6]、黄梦豪等[16]利用RIL群体分别结合SSR标记、SNP芯片等,在1A、1B、1D、2A、4A和6B染色体上共定位到20个与湿面筋含量相关的QTL,表型贡献率为0.6%—13.0%。ECHEVENY-SOLARTE等[14]以RIL群体结合包含939个DArT标记的图谱,在1A、1B、2B、3D、4A和6A染色体上共定位到6个与小麦籽粒出粉率相关的QTL,表型贡献率为4.9%—19.0%。吴云鹏等[15]、郭利建等[6]、黄梦豪等[16]利用RIL群体结合SSR标记、SNP芯片等,在1B、1D、2A、3A、3B、5A和7D染色体上共定位到11个与小麦籽粒沉降值相关的QTL,表型贡献率为2.7%—20.2%。郭利建等[6]、黄梦豪等[16]利用RIL群体结合SSR标记、SNP芯片等,在1A、2A、2D、4B、5A和6A染色体上共定位到10个与小麦籽粒淀粉含量相关的QTL,表型贡献率为2.9%—11.2%。【本研究切入点】SNP标记具有分布广、数量多、定位精度高等特点,是目前最具潜力的分子标记[17]。应用GWAS挖掘新疆春小麦品质性状相关基因的研究仍鲜见报道。【拟解决的关键问题】本研究以298份春小麦品种(系)构成的自然群体为材料,利用小麦55K SNP基因芯片分型数据结合蛋白质含量、湿面筋含量、沉降值、淀粉含量、籽粒硬度,出粉率和容重7个小麦品质相关性状表型鉴定结果,进行全基因组关联分析,发掘小麦品质相关性状的遗传位点,筛选相关候选基因,为相关基因克隆和分子标记辅助选择提供理论依据。1 材料与方法
1.1 试验材料与表型鉴定
以298份春小麦品种(系)为材料,其中包含国外引进品种(系)110份、新疆地方种141份、新疆育成种47份,该批材料由国家作物种质资源库——新疆分库提供。该群体一定程度上反映了不同时期适合新疆地区种植的小麦种质的特性。2018—2020年连续3个年度将参试材料种植于新疆乌鲁木齐市新疆农业科学院安宁渠基地,分别简称为E1、E2和E4;2019—2020年连续2个年度将参试材料种植于新疆喀什市新疆农业科学院泽普小麦育种家基地,分别简称为E3和E5。采用随机区组设计,每份材料种植3行,行长2 m,行距20 cm,3次重复,人工播种。在试验田四周设置保护行。试验田土壤肥力、灌溉等田间管理基本一致,播种前施基肥25 kg磷酸二铵和5 kg尿素,拔节期和抽穗期均使用滴灌追肥尿素10 kg和磷酸一铵5 kg。由于地方品种白粉病等发病率较高,在抽穗期人工喷施己唑醇进行防治,喷施2次,间隔10 d。抽穗期小麦植株会出现轻微倒伏现象,使用竹竿进行支撑固定以防止出现大面积倒伏。为保证测量数据的准确性,将小麦材料收获脱粒后,在适宜的环境保存2个月,采用瑞典波通仪器公司生产的DA-7250近红外分析仪对蛋白质含量、湿面筋含量、沉降值、淀粉含量、籽粒硬度,出粉率和容重7个品质相关性状进行测定,3次重复,取平均值;5个环境下各重复试验每份材料各品质性状结果取平均值。
1.2 表型数据的统计与分析
采用Excel 2016、SPSS 21.0软件对蛋白质含量、湿面筋含量、沉降值、淀粉含量、籽粒硬度,出粉率和容重7个品质相关性状进行统计分析,并计算各性状的平均值、标准差和变异系数。采用SAS软件进行描述性统计分析和方差分析[18]。广义遗传力用公式H2=σg2/(σg2+σge2+σƐ2/n)计算,其中,σg2为遗传方差,σge2为基因型与环境互作方差,σƐ2为环境方差,n为环境数。1.3 DNA提取及SNP分型
小麦生长至三叶期时取10株幼苗,将叶片混合后利用改良的CTAB法提取DNA[19]。用1.0%琼脂糖凝胶电泳检验DNA质量和浓度。利用小麦基因组55K SNP芯片对供试材料进行基因分型。小麦55K SNP芯片分型分析及样品质检工作由中玉金标记(北京)生物技术股份有限公司完成,共检测到覆盖小麦全基因组的53 063个SNP标记。采用Tassel 5.0软件对标记信息去冗余,剔除最小等位基因频率(minor allele frequency,MAF)小于5%和缺失率(missing rate)大于25%的SNP,最终得到23 632个多态性高、稳定性好的优质SNP标记用于后续关联分析。1.4 群体结构分析
应用Power Maker软件计算多态性信息量(polymorphic information content,PIC,PIC=1-ΣP2ij,P2ij表示第i个位点的第j个等位变异出现的频率)[20]。从筛选过的标记中,选取2 000个最小等位基因频率大于10%且在染色体上均匀分布的SNP标记,利用Structure 2.3.4软件进行群体结构分析。参数设置:Length of Burn-Period=10 000,MCMC Reps after Burn- in=100 000,选择Admixture Ancestry模型和Dependent Allele Frequencies模式,令K=2—12,每个K值重复运行5次。将不同K值下重复运行5次的结果上传至Structure Harvester(1.5 连锁不平衡分析
以位点间的相关系数平方(r2)作为衡量多态性位点两两之间的连锁不平衡(linkage disequilibrium,LD)参数。采用TASSEL5.0软件计算r2,以第95百分位的r2值作为阈值估测LD衰减距离。全基因组关联分析中,超过LD衰减距离的2个位点则认为是2个不同的位点,在LD衰减距离内的位点均视为同一位点。1.6 全基因组关联分析与候选基因筛选
使用TASSEL5.0软件中的Q+K混合线性模型对298份试验材料在5个不同环境下的8个籽粒品质性状的表型值进行性状与标记之间的GWAS,以P=1.0×10-3为阈值,判定SNP标记与目标性状关联的显著性[22],以LD衰减距离为依据将获得的SNP转化为与目标性状显著关联的遗传位点,将在2个及2个以上的环境中发现的位点视为稳定的位点。将多个环境中稳定出现的SNP标记的延伸序列在小麦TGACv1.0数据库(
2 结果
2.1 小麦籽粒品质性状表型数据的统计分析
通过分析298份供试材料在5个不同环境下的7个小麦籽粒品质性状的表型,发现国外引进品种(系)、新疆地方品种(系)和新疆育成品种(系)的品质性状有一定的差异(表1),3种类型小麦在5个环境下的变异系数分别为1.3%—13.4%、1.1%—18.6%和1.0%—13.9%。其中,容重的变异幅度最小;沉降值的变异幅度最大。不同环境中,国外引进品种(系)的蛋白质含量、湿面筋含量和沉降值平均值最高,分别为17.9%、40.4%和41.0%;新疆育成品种(系)的淀粉含量、籽粒硬度和出粉率最高,分别为67.4%、63.2%和71.3%;新疆地方品种(系)的上述6个品质性状在5个环境下均介于国外引进品种(系)和新疆育成品种(系)之间;容重在各环境中的表现不具有一致性,这个性状可能受环境的影响较大。供试材料7个品质性状的基因型×年份、环境×年份和基因型×环境×年份互作效应间差异均达极显著水平(表2),说明基因型×年份、环境×年份和基因型×环境×年份对于小麦品质性状均有极显著影响。所测品质性状的遗传力为0.61—0.95,其中,容重的遗传力最小;籽粒硬度的遗传力最大,说明籽容重受环境因素的影响较大。供试材料7个品质性状5个环境下的相关性分析显示(电子附表1),容重在5个环境两两之间相关系数平均值较小,分别为0.48;蛋白质含量、湿面筋含量、沉降值、淀粉含量、籽粒硬度和出粉率5个环境两两之间相关系数平均值为0.64—0.81,且各性状5个环境两两之间相关系数平均值与遗传力呈正相关。Table 1
表1
表1春小麦品质性状表型变异
Table 1
| 性状 Trait | 环境 Environment | 最大值 Max | 最小值 Min | 平均值 Mean | 标准差 SD | 变异系数 CV (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 外引种 Introduced varieties | 地方种 Xinjiang landrace | 育成种 Xinjiang bred varieties | 外引种 Introduced varieties | 地方种 Xinjiang landrace | 育成种 Xinjiang bred varieties | 外引种 Introduced varieties | 地方种 Xinjiang landrace | 育成种 Xinjiang bred varieties | 外引种 Introduced varieties | 地方种 Xinjiang landrace | 育成种 Xinjiang bred varieties | 外引种 Introduced varieties | 地方种 Xinjiang landrace | 育成种 Xinjiang bred varieties | ||
| 蛋白质 含量 PRC (%) | E1 | 22.60 | 22.03 | 19.79 | 13.96 | 13.40 | 14.22 | 18.67 | 16.53 | 16.81 | 1.70 | 1.71 | 1.51 | 9.13 | 10.36 | 8.98 |
| E2 | 21.40 | 23.24 | 19.97 | 14.72 | 13.18 | 12.93 | 17.95 | 16.16 | 15.80 | 1.42 | 1.87 | 1.15 | 7.89 | 11.60 | 7.27 | |
| E3 | 22.02 | 22.79 | 18.84 | 13.92 | 13.24 | 13.48 | 17.61 | 16.17 | 15.45 | 1.38 | 1.48 | 1.14 | 7.84 | 9.12 | 7.40 | |
| E4 | 20.79 | 21.83 | 19.71 | 13.99 | 12.37 | 13.18 | 16.60 | 16.18 | 15.85 | 1.42 | 1.58 | 1.15 | 8.54 | 9.74 | 7.28 | |
| E5 | 23.13 | 22.72 | 18.51 | 15.03 | 15.16 | 14.59 | 18.59 | 17.68 | 16.45 | 1.35 | 1.39 | 0.91 | 7.26 | 7.87 | 5.54 | |
| 湿面筋 含量 WGC (%) | E1 | 51.27 | 50.05 | 45.15 | 31.90 | 29.18 | 31.43 | 42.03 | 36.76 | 37.93 | 3.88 | 4.19 | 3.29 | 9.23 | 11.40 | 8.68 |
| E2 | 48.52 | 52.90 | 45.46 | 33.19 | 28.44 | 27.97 | 40.39 | 35.88 | 35.80 | 3.33 | 4.57 | 2.72 | 8.23 | 12.73 | 7.61 | |
| E3 | 50.05 | 52.44 | 43.09 | 31.76 | 28.97 | 29.88 | 39.90 | 36.09 | 35.35 | 3.21 | 3.58 | 2.66 | 8.05 | 9.93 | 7.52 | |
| E4 | 46.74 | 49.74 | 44.84 | 31.84 | 26.92 | 29.13 | 37.61 | 36.17 | 36.14 | 3.18 | 3.76 | 2.67 | 8.46 | 10.39 | 7.39 | |
| E5 | 52.26 | 51.98 | 40.80 | 34.31 | 33.33 | 32.40 | 41.84 | 39.54 | 37.67 | 3.11 | 3.41 | 2.07 | 7.42 | 8.61 | 5.51 | |
| 沉降值 SV (mL) | E1 | 62.27 | 63.12 | 51.69 | 31.06 | 26.64 | 30.23 | 46.69 | 39.78 | 39.17 | 6.00 | 6.40 | 4.82 | 12.86 | 16.08 | 12.31 |
| E2 | 58.03 | 60.47 | 45.38 | 30.57 | 23.30 | 25.38 | 42.61 | 35.93 | 34.05 | 4.74 | 6.68 | 3.63 | 11.13 | 18.58 | 10.67 | |
| E3 | 56.52 | 59.74 | 42.38 | 27.60 | 22.37 | 23.75 | 38.62 | 34.86 | 30.38 | 5.19 | 5.90 | 4.22 | 13.44 | 16.93 | 13.88 | |
| E4 | 50.56 | 57.78 | 43.56 | 25.21 | 18.66 | 25.09 | 38.54 | 35.72 | 32.19 | 4.83 | 6.40 | 3.98 | 12.52 | 17.93 | 12.37 | |
| E5 | 56.48 | 64.21 | 47.12 | 26.42 | 29.08 | 28.06 | 38.32 | 39.66 | 35.16 | 4.90 | 5.36 | 3.63 | 12.78 | 13.51 | 10.32 | |
| 淀粉 含量 STC (%) | E1 | 72.62 | 72.24 | 72.87 | 61.06 | 57.76 | 65.01 | 66.35 | 68.71 | 69.51 | 2.46 | 2.35 | 1.51 | 3.71 | 3.42 | 2.17 |
| E2 | 72.35 | 72.92 | 73.93 | 61.90 | 61.81 | 65.98 | 68.22 | 70.07 | 70.78 | 1.89 | 1.74 | 1.32 | 2.77 | 2.49 | 1.86 | |
| E3 | 71.73 | 71.50 | 72.66 | 60.33 | 62.54 | 66.05 | 67.45 | 68.27 | 70.24 | 1.77 | 1.63 | 1.28 | 2.62 | 2.39 | 1.82 | |
| E4 | 73.21 | 73.24 | 72.89 | 64.26 | 60.94 | 65.63 | 69.27 | 69.14 | 69.98 | 1.84 | 1.85 | 1.33 | 2.65 | 2.68 | 1.91 | |
| E5 | 70.07 | 70.25 | 70.54 | 59.80 | 61.63 | 66.97 | 65.90 | 66.75 | 68.69 | 1.75 | 1.75 | 0.93 | 2.65 | 2.62 | 1.36 | |
| 籽粒 硬度 GH (%) | E1 | 75.02 | 69.37 | 75.17 | 51.99 | 49.51 | 55.72 | 61.66 | 58.76 | 65.97 | 4.99 | 4.36 | 4.09 | 8.10 | 7.42 | 6.20 |
| E2 | 76.16 | 72.29 | 73.20 | 53.60 | 51.13 | 53.75 | 66.02 | 59.51 | 67.24 | 5.14 | 5.36 | 4.09 | 7.79 | 9.00 | 6.07 | |
| E3 | 69.44 | 67.07 | 70.07 | 53.35 | 49.54 | 54.14 | 61.56 | 56.27 | 64.42 | 3.92 | 3.97 | 3.34 | 6.37 | 7.05 | 5.18 | |
| E4 | 77.10 | 70.26 | 68.89 | 53.03 | 49.70 | 52.78 | 68.67 | 57.63 | 63.44 | 5.38 | 4.57 | 3.57 | 7.83 | 7.92 | 5.62 | |
| E5 | 70.30 | 73.12 | 70.65 | 45.98 | 48.64 | 53.29 | 57.97 | 55.55 | 63.51 | 5.56 | 4.28 | 3.66 | 9.59 | 7.70 | 5.76 | |
| 出粉率 FY (%) | E1 | 74.36 | 73.44 | 75.22 | 62.39 | 57.47 | 66.46 | 68.42 | 69.57 | 72.06 | 2.71 | 2.19 | 1.82 | 3.96 | 3.14 | 2.52 |
| E2 | 80.24 | 77.06 | 76.73 | 68.37 | 68.37 | 71.28 | 73.09 | 72.46 | 74.78 | 2.00 | 1.55 | 1.27 | 2.73 | 2.14 | 1.70 | |
| E3 | 77.41 | 74.34 | 74.16 | 67.10 | 66.82 | 69.34 | 71.12 | 69.89 | 72.51 | 1.59 | 1.41 | 1.22 | 2.24 | 2.02 | 1.68 | |
| E4 | 76.55 | 75.40 | 76.93 | 68.18 | 67.33 | 69.93 | 72.43 | 70.84 | 73.18 | 1.77 | 1.55 | 1.34 | 2.44 | 2.19 | 1.82 | |
| E5 | 75.38 | 74.65 | 73.72 | 67.38 | 64.45 | 67.53 | 71.20 | 68.55 | 70.51 | 1.68 | 1.56 | 1.34 | 2.36 | 2.28 | 1.90 | |
| 容重 TW (g·L-1) | E1 | 823.25 | 821.55 | 828.75 | 751.62 | 732.95 | 779.65 | 790.40 | 794.85 | 804.72 | 15.07 | 13.20 | 11.08 | 1.91 | 1.66 | 1.38 |
| E2 | 840.12 | 825.69 | 824.17 | 770.63 | 777.00 | 778.85 | 805.11 | 801.52 | 799.75 | 13.32 | 9.81 | 10.82 | 1.65 | 1.22 | 1.35 | |
| E3 | 818.22 | 817.23 | 825.58 | 756.64 | 771.87 | 788.60 | 796.16 | 795.10 | 804.40 | 10.67 | 9.08 | 8.39 | 1.34 | 1.14 | 1.04 | |
| E4 | 839.55 | 817.75 | 818.78 | 763.24 | 758.88 | 758.43 | 812.36 | 793.52 | 783.93 | 14.49 | 9.79 | 11.88 | 1.78 | 1.23 | 1.52 | |
| E5 | 824.60 | 821.85 | 822.48 | 774.59 | 764.13 | 771.23 | 796.36 | 796.72 | 797.77 | 11.27 | 9.84 | 9.69 | 1.42 | 1.24 | 1.21 | |
新窗口打开|下载CSV
Table 2
表2
表2春小麦品质性状方差分析
Table 2
| 性状 Trait | 基因型 Genotype | 环境 Environment | 基因型×环境 G×E | 重复 Replicate | 误差 Error | 遗传力 H2 |
|---|---|---|---|---|---|---|
| 蛋白质含量PRC | 30.50*** | 323.25*** | 2.71*** | 28.92*** | 0.94*** | 0.92 |
| 湿面筋含量WGC | 173.71*** | 1575.75*** | 14.49*** | 138.56*** | 5.07*** | 0.92 |
| 沉降值SV | 396.29*** | 5503.76*** | 38.53*** | 482.08*** | 12.76*** | 0.91 |
| 淀粉含量STC | 42.33*** | 1067.05*** | 4.58*** | 50.85*** | 1.38*** | 0.90 |
| 籽粒硬度GH | 406.19*** | 4375.56*** | 20.43*** | 20.52*** | 3.83*** | 0.95 |
| 出粉率FY | 41.49*** | 1793.73*** | 4.76*** | 5.17*** | 0.91*** | 0.89 |
| 容重TW | 1308.20*** | 7599.86*** | 267.46*** | 247.03*** | 54.04*** | 0.82 |
新窗口打开|下载CSV
2.2 SNP多态性及分布
从小麦55K SNP芯片中筛选出具有多态性的SNP标记23 632个,分布在A、B和D基因组染色体上的SNP位点数目分别为11 559(48.9%)、10 366(43.9%)和1 707(7.2%)个,可以看出,B基因组的多态性最高;D基因组的多态性明显低于A和B 2个基因组的(电子附表2)。在小麦21条染色体中,2A染色体上的标记数目最多(2 033个),4D染色体上的标记数目最少(117个)。3个染色体组的PIC表现为B(0.3119)>A(0.3036)>D(0.2319)。全基因组的PIC变异范围在0.0068—0.3750,平均值为0.3108。其中2A染色体的PIC最大(0.3423),4D染色体的最小(0.2204)。说明选用的标记在各染色体组上分布较均匀,且标记的等位变异间具有高度的多态性。2.3 群体结构分析和连锁不平衡分析
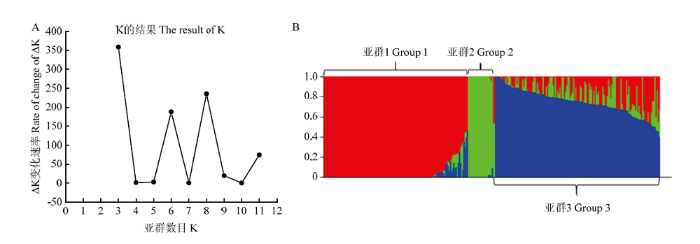
利用Structure2.3.4软件对群体遗传结构进行分析,对每个可能的K值模拟运算,用K值与ΔK值做图(图1-A),在K=3处ΔK值最大,曲线变化程度最大。由图1-B可以看出,供试材料被分为3个亚群,其中亚群1包含128(43.0%)份,主要来自新疆的地方品种(系);亚群2包含24(8.1%)份,主要包括外引品种(系)和新疆地方种(系);亚群3包含146(48.9%)份,主要是外引品种(系),还包括新疆育成品种(系)。因此将K=3时生成的Q矩阵,用于性状与标记的关联分析。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1298份小麦品种(系)群体结构分析
A:群体的∆K值;B:群体结构示意图
Fig. 1Population structure analysis of 298 wheat varieties (lines)
A: Estimation of ∆K value in population; B: Group structure diagram
考虑连锁群(linkage group)间连锁不平衡背景的影响,对连锁群间的r2值进行平方根转换,以大于此分布95%的参数值为阈值,用来截取同一连锁群内LD的衰减距离[24],经计算得到A、B和D基因组及全基因组的LD衰减距离分别为10、10、6和8 Mb,依据全基因组的LD衰减距离,将在物理图谱上前后8 Mb区间内的位点认定为一个候选位点(图2)。
图2
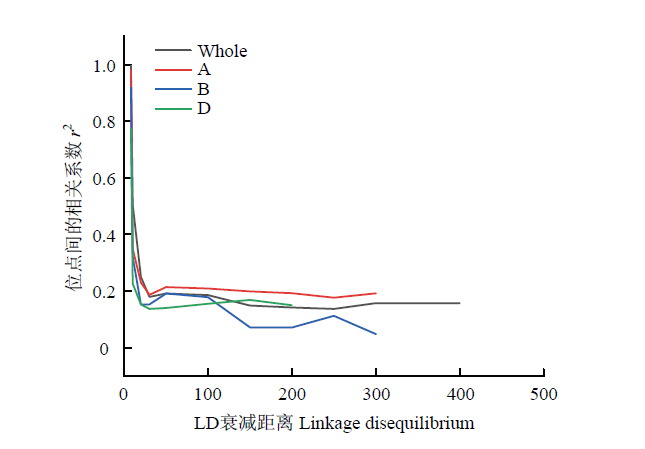 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2连锁不平衡衰减图
Fig. 2Linkage disequilibrium plot
2.4 全基因组关联分析
对298份供试材料在5个环境下的籽粒品质性状表型结合55K基因芯片分型的23 632个SNP标记进行GWAS,并采用MLM(mixed linear model,MLM)Q+K关联混合模型,共有85个位点同时在2个及2个以上环境中检测到,分布在除4D和6B染色体外的19条染色体上,是比较稳定的位点(表3)。在1B、1D、2D、3A、3D、4A、4B、5A、6A、6D、7A和7D染色体上检测到稳定的、且同时与多个性状关联的位点(表4)。其中,7A染色体上的8.91—16.64 Mb和26.47—32.70 Mb区段、7D染色体上的534.44—540.87 Mb和566.20—566.20 Mb区段检测到的稳定且同时与多个性状关联的位点可作为后续研究的重点。Table 3
表3
表3春小麦品质性状显著关联的稳定的位点信息
Table 3
| 性状 Trait | 位点 Marker | 染色体 Chr. | 物理位置 Position (Mb) | P值 P value | 表型贡献率 R2 (%) | 环境 Environment |
|---|---|---|---|---|---|---|
| 沉降值 SV (mL) | AX-109923777 | 1B | 335.51 | 1.25E-05—4.48E-05 | 6.96—7.83 | E4, E5 |
| AX-109389304 | 1B | 445.67 | 6.44E-05—4.57E-04 | 5.49—6.79 | E4, E5 | |
| AX-94935157 | 1D | 470.73—478.86 | 2.57E-05—3.85E-04 | 5.42—7.54 | E2, E5 | |
| AX-109486695 | 3A | 725.15 | 1.15E-04—5.59E-04 | 5.11—6.23 | E2, E5 | |
| AX-108986146 | 3B | 547.59 | 2.89E-05—3.18E-04 | 5.56—7.22 | E4, E5 | |
| AX-111583589 | 4A | 121.51 | 5.34E-05—7.95E-04 | 4.95—6.94 | E4, E5 | |
| AX-110417316 | 4A | 130.42 | 1.05E-05—6.22E-05 | 6.78—7.98 | E4, E5 | |
| AX-111264892 | 4A | 241.73 | 1.40E-04—2.24E-04 | 5.79—6.17 | E4, E5 | |
| AX-109608730 | 4A | 413.76 | 2.96E-05—3.05E-04 | 5.67—7.25 | E4, E5 | |
| AX-109285136 | 4A | 594.90—598.79 | 9.75E-05—8.96E-04 | 3.82—6.39 | E2, E3, E4 | |
| AX-109834040 | 4A | 609.82 | 2.7E-04—6.83E-04 | 4.98—5.65 | E4, E5 | |
| AX-110002719 | 4B | 43.56 | 2.69E-04—9.30E-04 | 3.79—4.69 | E2, E4 | |
| AX-94513518 | 4B | 556.06 | 2.13E-07—5.41E-04 | 5.24—10.89 | E2, E3 | |
| AX-110023236 | 5A | 30.66 | 7.47E-04—9.28E-04 | 4.80—4.95 | E2, E4 | |
| AX-109374128 | 5A | 600.17 | 7.45E-05—4.00E-04 | 5.43—6.61 | E4, E5 | |
| AX-110149809 | 5B | 554.32 | 2.12E-04—3.56E-04 | 5.45—5.92 | E3, E5 | |
| AX-109983932 | 5B | 604.61 | 5.60E-04—5.87E-04 | 5.15—5.18 | E2, E3 | |
| AX-109402563 | 5D | 28.88 | 3.63E-05—1.86E-04 | 6.05—7.11 | E4, E5 | |
| AX-110082918 | 5D | 529.22 | 5.79E-05—3.09E-04 | 5.57—6.72 | E4, E5 | |
| AX-109512342 | 6A | 25.80—28.33 | 3.91E-05—4.89E-04 | 5.25—7.11 | E2, E3, E4, E5 | |
| AX-94564307 | 6D | 21.98 | 4.64E-05—7.11E-04 | 4.98—6.93 | E2, E4 | |
| AX-111614875 | 6D | 70.62 | 3.43E-05—1.81E-04 | 5.96—7.10 | E4, E5 | |
| AX-109452823 | 7A | 26.47—32.70 | 5.93E-05—9.46E-04 | 3.72—6.34 | E1, E2, E4, E5 | |
| AX-109477546 | 7B | 155.48—159.71 | 1.99E-05—7.71E-04 | 5.03—10.36 | E2, E3, E4, E5 | |
| AX-94420739 | 7D | 540.87 | 9.04E-04—9.43E-04 | 4.83—5.08 | E2, E4 | |
| 出粉率 FY (%) | AX-109428041 | 1A | 22.48 | 4.14E-04—7.46E-04 | 5.02—5.24 | E3, E5 |
| AX-94802245 | 1D | 73.73—77.36 | 1.14E-04—1.00E-03 | 4.88—6.34 | E3, E4 | |
| AX-111099555 | 2B | 18.94—24.13 | 5.37E-05—3.64E-04 | 4.42—5.72 | E3, E4 | |
| AX-111040045 | 4A | 632.76-659.66 | 1.51E-04—9.74E-04 | 3.83—5.96 | E3, E5 | |
| AX-110671478 | 6A | 3.80—6.57 | 3.32E-06—9.73E-04 | 3.78—7.77 | E3, E4 | |
| AX-111534875 | 6A | 5.51—12.28 | 1.44E-05—3.98E-04 | 5.61—7.91 | E1, E3, E4 | |
| AX-111745518 | 6A | 17.54—24.41 | 1.51E-04—4.85E-04 | 6.14—6.15 | E1, E4 | |
| AX-111611885 | 6D | 2.49—5.82 | 1.57E-04—7.67E-04 | 5.00—6.14 | E3, E4 | |
| AX-111076418 | 7A | 8.91—12.44 | 1.70E-04—8.12E-04 | 3.75—5.14 | E4, E5 | |
| 蛋白质含量 PRC (%) | AX-94935157 | 1D | 470.73—478.18 | 8.52E-04—9.52E-04 | 3.85—4.77 | E2, E4 |
| AX-94694898 | 2B | 620.64 | 2.57E-04—8.16E-04 | 4.87—5.71 | E2, E4 | |
| AX-111218282 | 2D | 523.36 | 3.71E-04—8.94E-04 | 4.82—5.47 | E2, E4 | |
| AX-94589328 | 2D | 624.10 | 6.02E-04—8.85E-04 | 5.10—5.12 | E2, E4 | |
| AX-110442313 | 3A | 510.31—510.60 | 2.91E-04—8.05E-04 | 4.88—5.62 | E2, E4 | |
| AX-110002719 | 4B | 43.56 | 1.20E-04—2.83E-04 | 4.59—5.13 | E2, E4, E5 | |
| AX-111560874 | 5B | 10.50 | 1.38E-04—5.85E-04 | 5.11—6.15 | E2, E4 | |
| AX-109300733 | 6A | 12.28—17.54 | 5.03E-05—8.33E-04 | 4.87—6.98 | E2, E4 | |
| AX-95194586 | 6D | 77.52 | 3.12E-04—9.12E-04 | 4.97—5.68 | E2, E4 | |
| AX-111654743 | 7A | 32.00—32.70 | 8.79E-05—8.48E-04 | 3.83—5.30 | E1, E2, E3, E5 | |
| AX-108797850 | 7B | 453.77 | 2.79E-04—7.67E-04 | 4.93—5.66 | E2, E4 | |
| AX-108753131 | 7D | 534.44—540.87 | 3.72E-04—6.37E-04 | 5.17—5.70 | E2, E4 | |
| AX-108974357 | 7D | 566.20—566.23 | 4.25E-04—8.84E-04 | 4.99—5.36 | E2, E4 | |
| AX-110945986 | 7D | 609.24—610.23 | 2.75E-04—8.28E-04 | 4.97—5.68 | E2, E3 | |
| 淀粉含量 STC (%) | AX-95180100 | 1D | 359.67 | 2.12E-05—2.57E-04 | 5.75—7.55 | E4, E5 |
| AX-94589328 | 2D | 624.10—629.21 | 8.13E-05—8.67E-04 | 4.86—6.63 | E1,E2 | |
| AX-109934763 | 3D | 559.16—561.01 | 3.24E-05—9.76E-04 | 4.78—7.56 | E1, E2 | |
| AX-110417316 | 4A | 130.42—136.97 | 4.48E-04—8.12E-04 | 4.93—5.59 | E1, E2 | |
| AX-109332913 | 4A | 701.53 | 2.32E-04—7.15E-04 | 5.26—5.79 | E1, E2 | |
| AX-110169414 | 5A | 575.26 | 2.31E-04—5.72E-04 | 4.07—4.90 | E1, E2 | |
| AX-110046517 | 6A | 8.09—12.28 | 2.58E-05—8.16E-04 | 5.14—7.43 | E1, E3 | |
| AX-111745518 | 6A | 17.54 | 5.73E-04—8.12E-04 | 4.90—5.42 | E1, E2 | |
| AX-108888443 | 6D | 16.23—21.98 | 6.92E-04—9.04E-04 | 4.82—5.26 | E1, E2 | |
| AX-110545157 | 7A | 32.70 | 2.86E-04—7.03E-04 | 4.14—4.54 | E1, E2 | |
| AX-108974357 | 7D | 566.20—566.23 | 2.60E-04—7.07E-04 | 5.00—5.80 | E1, E2 | |
| 容重 TW (g·L-1) | AX-109301633 | 2A | 84.15 | 4.89E-04—9.53E-04 | 4.50—5.44 | E1, E3 |
| AX-110173580 | 2A | 697.04—698.01 | 2.69E-04—4.71E-04 | 4.23—6.47 | E1, E2 | |
| AX-86184511 | 2B | 158.15—164.43 | 2.03E-04—9.14E-04 | 4.53—4.79 | E1, E4 | |
| AX-110946610 | 4A | 608.03—616.89 | 7.28E-04—7.84E-04 | 3.93—4.71 | E1, E2 | |
| AX-109947431 | 5B | 672.48—678.51 | 2.29E-04—6.26E-04 | 5.26—5.84 | E3, E5 | |
| AX-110402876 | 7D | 626.92 | 3.38E-04—4.10E-04 | 5.43—5.56 | E2, E5 | |
| 湿面筋含量 WCG (%) | AX-110002719 | 4B | 43.56 | 1.27E-04—3.97E-04 | 4.32—5.06 | E2, E4, E5 |
| AX-111560874 | 5B | 10.50 | 2.83E-04—9.94E-04 | 4.71—5.61 | E2, E4 | |
| AX-111745518 | 6A | 17.54—25.80 | 3.01E-04—6.74E-04 | 4.97—5.75 | E2, E4 | |
| AX-111654743 | 7A | 32.00—32.70 | 7.64E-05—7.13E-04 | 4.72—5.35 | E1, E2 | |
| AX-94420739 | 7D | 540.87 | 6.47E-04—6.52E-04 | 5.04—-5.22 | E2, E4 | |
| 籽粒硬度 GH (%) | AX-111107679 | 1A | 476.08—483.94 | 1.32E-04—6.54E-04 | 4.92—5.01 | E4, E5 |
| AX-110918237 | 1A | 487.48—489.18 | 5.71E-05—8.41E-04 | 4.76—6.59 | E4, E5 | |
| AX-109499408 | 1A | 496.31—500.66 | 1.77E-04—6.78E-04 | 3.94—5.91 | E3, E5 | |
| AX-110369038 | 1B | 627.15—628.51 | 3.72E-04—9.45E-04 | 3.81—4.74 | E1, E2, E3 | |
| AX-94534666 | 1D | 457.15—457.95 | 3.16E-06—5.85E-04 | 4.01—7.42 | E2, E3, E5 | |
| AX-111636440 | 2A | 505.33—507.36 | 1.45E-04—9.12E-04 | 3.92—5.34 | E1, E3 | |
| AX-94447403 | 2D | 439.44 | 5.38E-04—7.59E-04 | 4.84—5.39 | E1, E3 | |
| AX-111640432 | 3A | 728.42—731.08 | 1.70E-04—5.00E-04 | 4.12—4.76 | E2, E3, E5 | |
| AX-110951756 | 3B | 7.08 | 7.16E-06—8.58E-04 | 4.73—8.11 | E2, E3 | |
| AX-111018209 | 3D | 553.59—560.75 | 3.97E-04—8.39E-04 | 4.98—5.28 | E1, E5 | |
| AX-111031457 | 5A | 571.71—571.94 | 9.82E-05—1.82E-04 | 4.72—5.12 | E3, E5 | |
| AX-108945948 | 6A | 9.14—13.06 | 1.00E-04—9.09E-04 | 5.00—6.02 | E1, E5 | |
| AX-108802871 | 7A | 10.09—16.64 | 2.96E-04—8.09E-04 | 4.32—5.85 | E1, E5 | |
| AX-111664463 | 7A | 123.20—127.55 | 8.90E-05—6.93E-04 | 3.86—6.29 | E4, E5 | |
| AX-110495496 | 7A | 484.39—489.95 | 2.30E-04—9.39E-04 | 3.72—5.65 | E3, E4, E5 |
新窗口打开|下载CSV
Table 4
表4
表4同时与多个品质性状显著关联的稳定位点信息
Table 4
| 性状 Trait | 位点 Marker | 染色体 Chr. | 物理位置 Position (Mb) | P值 P value | 表型贡献率 R2 (%) | 环境 Environment |
|---|---|---|---|---|---|---|
| 沉降值,蛋白质含量 SV, PRC | AX-94935157 | 1D | 470.73—478.86 | 2.57E-05—9.52E-04 | 3.85—7.54 | E2, E4, E5 |
| 淀粉含量,蛋白质含量 STC, PRC | AX-94589328 | 2D | 624.10—629.21 | 8.13E-05—8.85E-04 | 4.86—6.63 | E1, E2, E4 |
| 籽粒硬度,沉降值 GH, PRC | AX-111640432 | 3A | 725.15—731.08 | 1.15E-04—5.59E-04 | 4.12—6.23 | E2, E3, E5 |
| 籽粒硬度,淀粉含量 GH, STC | AX-111018209 | 3D | 553.59—561.01 | 3.24E-05—9.76E-04 | 4.78—7.56 | E1, E2, E5 |
| 淀粉含量,沉降值 STC, SV | AX-110417316 | 4A | 130.42—136.97 | 1.05E-05—8.12E-04 | 4.93—7.98 | E1, E2, E4, E5 |
| 容重,沉降值 TW, SV | AX-110946610 | 4A | 608.03—616.89 | 2.70E-04—7.84E-04 | 3.93—5.65 | E1, E2, E4, E5 |
| 沉降值,蛋白质含量,湿面筋含量 SV, PRC, WGC | AX-110002719 | 4B | 43.56 | 1.20E-04—9.30E-04 | 3.79—5.13 | E2, E4, E5 |
| 籽粒硬度,淀粉含量 GH, STC | AX-111031457 | 5A | 571.71—575.26 | 9.82E-05—5.72E-04 | 4.07—5.12 | E1, E2, E3, E5 |
| 蛋白质含量,湿面筋含量 PRC, WGC | AX-111560874 | 5B | 10.50 | 1.38E-04—9.94E-04 | 4.71—6.15 | E2, E4 |
| 出粉率,淀粉含量,籽粒硬度 FY, STC, GH | AX-111534875 | 6A | 5.51—12.28 | 1.44E-05—9.09E-04 | 5.00—7.91 | E1, E3, E4, E5 |
| 淀粉含量,蛋白质含量 STC, PRC | AX-111745518 | 6A | 12.28—17.54 | 5.03E-05—8.33E-04 | 4.87—6.98 | E1, E2, E4 |
| 出粉率,湿面筋含量 FY, WGC | AX-111745518 | 6A | 17.54—25.80 | 1.51E-04—6.74E-04 | 4.97—6.15 | E1, E2, E4 |
| 淀粉含量,沉降值 STC, SV | AX-108888443 | 6D | 16.23—21.98 | 4.64E-05—9.04E-04 | 4.82—5.69 | E1, E2, E4 |
| 沉降值,蛋白质含量 SV, PRC | AX-111614875 | 6D | 70.62—77.52 | 3.43E-05—9.12E-04 | 4.97—7.10 | E2, E4, E5 |
| 籽粒硬度,出粉率 GH, FY | AX-108802871 | 7A | 8.91—16.64 | 1.70E-04—8.12E-04 | 3.75—5.85 | E1, E4, E5 |
| 沉降值,淀粉含量,蛋白质含量,湿面筋含量 SV, STC, PRC, WGC | AX-109452823 | 7A | 26.47—32.70 | 5.93E-05—9.46E-04 | 3.72—6.34 | E1, E2, E3, E4, E5 |
| 沉降值,湿面筋含量,蛋白质含量 SV, WGC, PRC | AX-94420739 | 7D | 534.44—540.87 | 3.72E-04—9.43E-04 | 4.83—5.70 | E2, E4 |
| 蛋白质含量,淀粉含量 PRC, STC | AX-108974357 | 7D | 566.20—566.23 | 2.60E-04—8.84E-04 | 4.99—5.80 | E1, E2, E4 |
新窗口打开|下载CSV
位于1B染色体445.67 Mb检测到与沉降值相关的位点;3A染色体510.31—510.60 Mb与蛋白质含量相关的位点;6A染色体5.51—12.28 Mb同时与出粉率、淀粉含量和籽粒硬度相关的位点;6A染色体12.28—17.54 Mb同时与淀粉含量和蛋白质含量相关的位点;6A染色体17.54—25.80 Mb同时与出粉率和湿面筋含量相关的位点;7A染色体26.47—32.70 Mb同时与沉降值、淀粉含量、蛋白质含量和湿面筋含量相关的位点;以上6个位点可能为新的位点(表4)。
2.5 候选基因筛选
将与小麦籽粒品质性状显著关联的稳定位点SNP标记序列在普通小麦中国春基因组数据库搜索,获取基因序列,在NCBI数据库中进行BLASTx[25],共得到10个与小麦籽粒品质性状相关的基因(表5)。其中,TraesCS4A01G299800编码阳离子氨基酸转运蛋白(cationic amino acid transporter,putative);TraesCS6A01G054400编码F-box家族蛋白(F-box family protein);TraesCS7A01G059500编码色氨酸脱羧酶(tryptophan decarboxylase);TraesCS6A01G012800编码富含亮氨酸的重复受体样蛋白激酶家族蛋白(leucine-rich repeat receptor-like protein kinase family protein);TraesCS4B01G054200编码组蛋白H2B(Histone H2B);TraesCS1D01G382000编码锌指状蛋白(zinc finger protein-like);TraesCS3A01G507800编码含螺旋结构域的蛋白(coiled- coil domain-containing protein);TraesCS7A01G331200和TraesCS7D01G418700编码木葡聚糖内转葡糖基酶/水解酶(xyloglucan endotransglucosylase/hydrolase);TraesCS7D01G445400对应结构为ATP依赖性Clp蛋白酶ATP结合亚基(ATP-dependent Clp protease ATP-binding subunit)。Table 5
表5
表5稳定位点筛选获得候选基因信息
Table 5
| 性状 Trait | 位点 Marker | 染色体 Chr. | 物理位置 Position (Mb) | 基因 Gene | 基因注释或编码蛋白 Gene annotation or coding protein |
|---|---|---|---|---|---|
| 沉降值 SV | AX-109285136 | 4A | 594.90—598.79 | TraesCS4A01G299800 | 阳离子氨基酸转运蛋白,推定 Cationic amino acid transporter, putative |
| AX-109512342 | 6A | 25.80—28.33 | TraesCS6A01G054400 | box家族蛋白 F-box family protein | |
| 蛋白质含量,沉降值 PRC, SV | AX-109452823 | 7A | 26.47—32.70 | TraesCS7A01G059500 | 色氨酸脱羧酶 Tryptophan decarboxylase |
| 出粉率 FY | AX-111534875 | 6A | 5.51—12.28 | TraesCS6A01G012800 | 富含亮氨酸的重复受体样蛋白激酶家族蛋白 Leucine-rich repeat receptor-like protein kinase family protein |
| 湿面筋含量, 蛋白质含量 WGC, PRC | AX-110002719 | 4B | 43.56 | TraesCS4B01G054200 | 组蛋白H2B Histone H2B |
| 籽粒硬度 GH | AX-94534666 | 1D | 457.15—457.95 | TraesCS1D01G382000 | 锌指状蛋白 Zinc finger protein-like |
| AX-111640432 | 3A | 728.42—731.08 | TraesCS3A01G507800 | 含螺旋结构域的蛋白Coiled-coil domain-containing protein | |
| AX-110495496 | 7A | 484.39—489.95 | TraesCS7A01G331200 | 木葡聚糖内转葡糖基酶/水解酶 Xyloglucan endotransglucosylase/hydrolase | |
| 蛋白质含量 PRC | AX-108974357 | 7D | 566.20—566.23 | TraesCS7D01G445400 | ATP依赖性Clp蛋白酶ATP结合亚基 ATP-dependent Clp protease ATP-binding subunit |
| AX-108753131 | 7D | 534.44—540.87 | TraesCS7D01G418700 | 木葡聚糖内转葡糖基酶/水解酶 Xyloglucan endotransglucosylase/hydrolase |
新窗口打开|下载CSV
3 讨论
3.1 春小麦籽粒品质性状表型变异
小麦是全球主要粮食作物之一。自20世纪80年代以来,小麦品质遗传改良一直是育种家比较关注的领域[6]。本研究以298份春小麦为材料,通过近红外谷物分析仪测定5个环境下小麦籽粒品质性状。结果显示,不同环境下国外引进品种(系)的蛋白质含量、湿面筋含量和沉降值均最高;新疆育成品种(系)的淀粉含量、籽粒硬度和出粉率最高,这可能是由于新疆地区对于小麦品质的关注较晚一些,之前新疆小麦育种家在小麦品种选育时关注小麦的产量、出粉率等比较直观的性状,而忽略了肉眼不可见的品质性状。近些年,小麦品质逐渐成为新疆小麦育种的主要目标之一。本研究和张金波等[26]研究均表明,近十几年来,育种家以新疆地方种和国外引进品种(系)为亲本选育了一批适合在新疆种植的品质较优的春小麦品种,这些品种的蛋白质含量和湿面筋含量较之前主要种植的地方品种都有显著提高,这也说明了小麦育种家不再单一注重小麦的产量和出粉率,也逐渐关注小麦品质遗传改良的结果。本研究与金欣欣等[27]研究均表明,小麦籽粒品质性状主要由遗传因素控制,因此,可以通过挖掘与小麦品质性状相关的遗传位点和候选基因对春小麦进行遗传改良,为优质小麦新品种选育提供参考。3.2 小麦品质的生态适应性
本研究表明,7个小麦籽粒品质性状的基因型×年份、环境×年份和基因型×环境×年份互作效应间差异均达极显著水平(表2),说明基因型×年份、环境×年份和基因型×环境×年份对于小麦品质性状均有极显著影响,基因型和环境是决定小麦品质性状的重要因素;但是,7个品质性状在5个环境的相关性分析显示,容重在5个环境两两之间相关系数平均值较小(0.48),同时其遗传力也相对较小(0.82);蛋白质含量、湿面筋含量、沉降值、淀粉含量、籽粒硬度和出粉率在5个环境两两之间相关系数平均值为0.64—0.81,遗传力为0.89—0.95;且遗传力大的性状其5个环境两两之间相关系数平均值也较大,说明基因型是决定小麦品质性状的主要因素,这与燕丽等[3]的研究结果一致。与北疆地区种植的(E1、E2和E4)小麦品质性状相比,南疆地区种植的小麦籽粒蛋白质含量和湿面筋含量较高;而沉降值、淀粉含量、籽粒硬度、出粉率和容重相对较低,这是由于不同的品质性状对环境变化的响应有所差异所导致的。光照和温湿度对面筋含量和蛋白质含量影响显著,这与张影全等[28]的研究结果一致。信志红等[10]和吴新元等[29]的研究也表明,光照和温度与小麦籽粒蛋白质含量呈正相关。同一生育期南疆地区的日照和温度均高于北疆地区,这可能是导致南疆地区小麦籽粒蛋白质含量和湿面筋含量较高的主要原因。可以通过挖掘调控小麦品质的遗传位点和功能基因,选育出适宜南、北疆地区种植的春小麦品种,扩大优质小麦的种植面积,以期解决现阶段人们高质量的饮食需求。3.3 春小麦品质性状全基因组关联分析
全基因组关联分析在小麦遗传改良方面的应用越来越广泛[8]。LI等[13]在1B、1D、6B和7D染色体上检测到与湿面筋含量相关的位点,与本研究检测到的位点一致。本研究在7D染色体上的540.87 Mb处还检测到与湿面筋含量显著关联的环境稳定位点。黄梦豪等[16]、杨林等[30]也在7D染色体上检测到与小麦品质性状相关的位点,进一步验证了7D染色体对于调控小麦品质性状的重要作用。本研究与吴云鹏等[15]均在3A和3B染色体上检测到与蛋白质含量相关的位点,在1B、1D和3B染色体上检测到与沉降值相关的位点,表明这些染色体上可能存在调控小麦籽粒蛋白质含量和沉降值的基因。本研究和LOU等[31]在2B染色体上18.94—24.13 Mb这一区段内同时检测到与小麦籽粒品质性状相关的位点,说明2B染色体的这一区段在调控小麦籽粒品质性状方面起重要作用,可以作为下一步研究的重点。本研究与杨林等[30]、沈玮囡等[32]均在1B和3B染色体上检测到与小麦籽粒品质性状相关的位点。本研究结合前人的研究结果筛选出多个与小麦籽粒品质性状相关的相同位点和重要染色体区段,这些位点具有深入研究的价值。LOU等[31]在3A染色体上的484.64 Mb处检测到与蛋白质含量相关的稳定位点。本研究在3A染色体的510.31—510.60 Mb区段检测到与蛋白质含量相关的稳定位点。这两个位点在染色体上的距离为25.67—25.96 Mb,可以确定不是同一位点,本研究发现的这一位点可能是新的控制小麦籽粒品质性状的遗传位点。本研究在3B染色体上的547.59 Mb处检测到的位点与LOU等[31]在3B染色体554.03 Mb处检测到的位点在染色体上的距离为6.44 Mb,根据全基因组LD衰减距离(8 Mb)可以确定这两个位点为同一位点;但位于1B染色体上的445.67 Mb区段与沉降值相关的位点与LOU等[31]在1B染色体上的471.52 Mb处检测到的位点在染色体上的距离为25.85 Mb,也可以确定不是同一位点,同样也未见相关报道,可能是新的位点。本研究与郭利建等[6]、杨林等[30]在5A和6A染色体上检测到的与蛋白质含量、湿面筋含量等相关的位点。其中本研究在6A染色体上的3.80—25.80 Mb区间内发现多个稳定的位点,且未见有关于这一区段内控制品质性状位点报道。LOU等[31]在7A染色体的6.85 Mb处检测到与淀粉含量相关的位点与本研究在7A染色体上的8.91—12.44 Mb处检测到的与出粉率和籽粒硬度相关的位点在染色体上的距离为2.06—5.59 Mb,根据全基因组LD衰减距离(8 Mb)可以确定这两个位点为同一位点。本研究在7A染色体上的26.47—32.70 Mb区段检测到的与沉降值、淀粉含量、蛋白质含量和湿面筋含量相关的位点未见报道,这一位点可能是新的控制小麦品质性状的遗传位点。
3.4 候选基因预测
将全基因组关联分析检测到的与小麦籽粒品质性状显著关联、且稳定的位点及重要区段进行候选基因筛选,在7A染色体上的484.39—489.95 Mb区段和7D染色体上的534.44—540.87 Mb区段处筛选出与籽粒硬度相关的候选基因TraesCS7A01G331200和与蛋白质含量相关的候选基因TraesCS7D01G418700编码木葡聚糖内转葡糖基酶/水解酶(xyloglucan endotransglucosylase/hydrolase,XTH)。前人研究表明,XTH基因在植株体内表现出非常复杂的功能[33]。它参与了种子萌发[34]、根系建成[35]、花形成开放以及果实发育成熟[36]等重要生物学过程。关于XTH基因在小麦籽粒品质方面的作用还需进一步研究。本研究还筛选出编码F-box家族蛋白(TraesCS6A01G054400)、富含亮氨酸的重复受体样蛋白激酶家族蛋白(TraesCS6A01G012800)、锌指状蛋白(TraesCS1D01G382000)等结构的候选基因,这些基因在调控小麦抗逆性方面有重要作用[37,38,39]。关于这些基因在品质方面的作用,还需要进一步研究。小麦中含有多种人体必需氨基酸,同时,小麦籽粒中赖氨酸等氨基酸的含量对小麦的营养品质有重要作用[40],本研究在4A染色体上的594.90—598.79 Mb区段筛选到编码阳离子氨基酸转运蛋白(TraesCS4A01G299800)的候选基因;在7A染色体上的26.47—32.70 Mb区段内筛选到编码色氨酸脱羧酶(TraesCS7A01G059500)的候选基因;在6A染色体上的5.51—12.28 Mb区段内筛选到编码富含亮氨酸的重复受体样蛋白激酶家族蛋白(TraesCS6A01G012800)的候选基因;这些候选基因可能与小麦籽粒中氨基酸含量有关,需要进一步对其进行深入研究,为小麦品质改良和新品种选育奠定基础。4 结论
通过对298份春小麦品种(系)3年5点的7个籽粒品质性状结合55K SNP基因芯片进行全基因组关联分析,发现85个稳定的显著关联位点,包括18个与多个性状关联的多效性位点;在1B(445.67 Mb)、3A(510.31—510.60 Mb)、6A(5.51—12.28 Mb、12.28—17.54 Mb和17.54—25.80 Mb)和7A(26.47—32.70 Mb)染色体上的位点可能是新的品质性状位点。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 8]
[本文引用: 8]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s10681-005-9062-7URL [本文引用: 1]
DOI:10.1186/s12870-017-0999-1URL [本文引用: 1]
DOI:10.4238/2013.May.21.5URL [本文引用: 3]
DOI:10.1007/s00122-015-2478-0URL [本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 5]
[本文引用: 5]
DOI:10.1007/s00122-013-2065-1URL [本文引用: 1]
DOI:10.1016/j.cj.2015.01.001URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/ng1702URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1534/genetics.105.044586URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1007/s00122-020-03704-yURL [本文引用: 5]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.jplph.2010.06.029URL [本文引用: 1]
DOI:10.1093/jxb/eru202URL [本文引用: 1]
DOI:10.1007/s00299-017-2105-4URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.11983/CBB16001 [本文引用: 1]

质膜作为原生质体与外界环境的屏障, 除了维持正常的细胞内稳态和营养状况, 还参与感知和应答各种环境胁迫。近年来, 植物质膜蛋白质组学研究为深入分析植物应答不同生物和非生物胁迫的分子机制提供了重要信息, 已经报道了模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)等10种植物质膜应对生物胁迫(白叶枯病菌(Xanthomonas oryzae pv. oryzae)感染)与非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、氮素、脱落酸、壳聚糖和壳寡糖)过程的蛋白质丰度模式变化。通过整合分析植物质膜响应逆境的蛋白质组学研究结果, 揭示了质膜在植物应答逆境胁迫过程中的重要作用。植物通过调节转运蛋白、通道蛋白及膜泡运输相关蛋白的丰度变化促进细胞内外的信号传递、物质交换与运输; 同时利用膜相关的G蛋白、Ca<sup>2+</sup>信号、磷酸肌醇信号途径及BR信号途径等多种信号通路, 通过蛋白质可逆磷酸化作用感知和传递胁迫信号, 调节植物抵御胁迫。研究结果为从蛋白质水平认识质膜逆境应答分子调控机制提供了新线索。
DOI:10.11983/CBB16001 [本文引用: 1]
质膜作为原生质体与外界环境的屏障, 除了维持正常的细胞内稳态和营养状况, 还参与感知和应答各种环境胁迫。近年来, 植物质膜蛋白质组学研究为深入分析植物应答不同生物和非生物胁迫的分子机制提供了重要信息, 已经报道了模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)等10种植物质膜应对生物胁迫(白叶枯病菌(Xanthomonas oryzae pv. oryzae)感染)与非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、氮素、脱落酸、壳聚糖和壳寡糖)过程的蛋白质丰度模式变化。通过整合分析植物质膜响应逆境的蛋白质组学研究结果, 揭示了质膜在植物应答逆境胁迫过程中的重要作用。植物通过调节转运蛋白、通道蛋白及膜泡运输相关蛋白的丰度变化促进细胞内外的信号传递、物质交换与运输; 同时利用膜相关的G蛋白、Ca<sup>2+</sup>信号、磷酸肌醇信号途径及BR信号途径等多种信号通路, 通过蛋白质可逆磷酸化作用感知和传递胁迫信号, 调节植物抵御胁迫。研究结果为从蛋白质水平认识质膜逆境应答分子调控机制提供了新线索。
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}