Interference in TP53INP2 Gene Inhibits the Differentiation of Bovine Myoblasts
DU JiaWei,1, DU XinZe1, YANG XinRan1, SONG GuiBing1, ZHAO Hui1, ZAN LinSen1,2, WANG HongBao,1,2,*1College of Animal Science and Technology, Northwest A&F University, Yangling 712100, Shaanxi; 2National Beef Cattle Improvement Center, Yangling 712100, Shaanxi
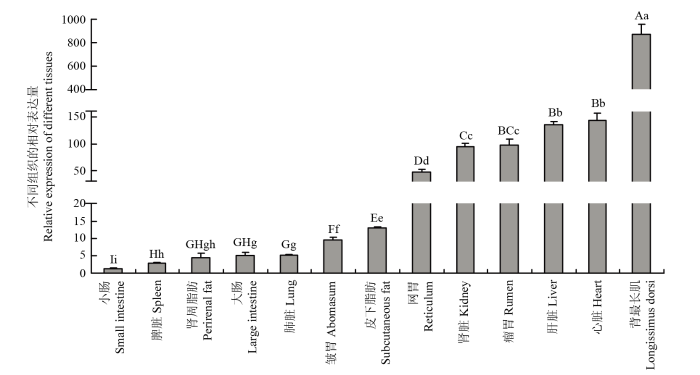
Abstract 【Background】Muscle can maintain the motor function of mammals and regulate body metabolism. Its quantity and distribution have an important influence on meat quality. The growth and development of skeletal muscle and its genetic characteristics influence and even determine the meat production and meat quality to a large extent. It is of great significance to study the growth and development of skeletal muscle. The regulatory effect of TP53INP2 on autophagy and the regulation mechanism on the differentiation of preadipocytes have been studied, but whether it affects the differentiation of bovine myoblasts has not been reported. 【Objective】 This study aims to explore the effect of TP53INP2 on the differentiation of Qinchuan bovine myoblasts in order to provide a theoretical basis for the molecular breeding of beef cattle meat traits. 【Method】 The real-time fluorescence quantitative PCR (RT-qPCR) technology was used to detect the expression characteristics of TP53INP2 in different tissues of Qinchuan cattle at 24 months of age. At the same time, the expression patterns of bovine skeletal myoblasts cultured in vitro at different stages of differentiation were analyzed. TP53INP2 gene siRNA was synthesized and transfected to Qinchuan bovine myoblasts, cells were induced to differentiation 12 hours after transfection, phenotypic changes of myoblasts were observed at different time pionts, and RT-qPCR and Western Blot technologies were performed respectively to detect the expression of differentiation marker genes and proteins on the fourth day of induced differentiation. 【Result】 1. RT-qPCR results showed that the expression level of TP53INP2 was the highest in adult Qinchuan cattle Longissimus dorsi muscle tissue and the lowest in the small intestine tissue. It is higher in adult bovine heart, liver, kidney, reticulum and rumen, and lower in other tissues (compared with longissimus dorsi). 2. With the differentiation of myoblasts, the expression of this gene increased from 0 to 4 days and reached a peak on the 4th day, and then decreased. 3. After interfering with TP53INP2 siRNA in myoblasts, the number and length of myotubes in the test group were significantly lower than those in the control group. 4. RT-qPCR results showed that the expression of myoblast differentiation marker genes myogenin (MYOG) and myosin heavy chain protein 3 (MYH3) was significantly lower than that of the control group. Western Blot results showed that the protein expressions of MYOG, MYH3 and MYOD in the test group were reduced and the differences were extremely significant compared with the control group.【Conclusion】Interfering of TP53INP2 has an inhibitory effect on the differentiation of bovine myoblasts, suggesting that this gene may have an important regulatory effect on the growth and development of Qinchuan cattle muscle tissue, and it can be used for in-depth functional research for molecular breeding of beef cattle practice. Keywords:TP53INP2;differentiation;myoblasts;Qinchuan cattle
PDF (2258KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 杜嘉伟, 杜鑫泽, 杨昕冉, 宋贵兵, 赵慧, 昝林森, 王洪宝. 干扰TP53INP2抑制牛成肌细胞分化. 中国农业科学, 2021, 54(21): 4685-4693 doi:10.3864/j.issn.0578-1752.2021.21.017 DU JiaWei, DU XinZe, YANG XinRan, SONG GuiBing, ZHAO Hui, ZAN LinSen, WANG HongBao. Interference in TP53INP2 Gene Inhibits the Differentiation of Bovine Myoblasts. Scientia Agricultura Sinica, 2021, 54(21): 4685-4693 doi:10.3864/j.issn.0578-1752.2021.21.017
标不同小写字母表示差异显著(P<0.05),标不同大写字母表示差异极显著(P<0.01),标相同字母表示差异不显著 Fig. 1The expression of TP53INP2 gene in different tissues of adult Qinchuan cattle
Different lowercase letters indicate significant differences(P<0.05), different uppercase letters indicate extremely significant differences(P<0.01), and the same letters indicate that the difference is not significant
D0:成肌细胞诱导分化第0天;D2:成肌细胞诱导分化第2天;D4:成肌细胞诱导分化第4天;D6:成肌细胞诱导分化第6天;D8:成肌细胞诱导分化第8天;D10:成肌细胞诱导分化第10天 Fig. 2Expression of TP53INP2 in muscle cells on different days
D0: day 0 of induced differentiation of myoblasts; D2: day 2 of induced differentiation of myoblasts; D4: day 4 of induced differentiation of myoblasts; D6: day 6 of induced differentiation of myoblasts; D8: induction of myoblasts Day 8 of differentiation; D10: Day 10 of differentiation of myoblasts
CHEN LJ, CHENZ, LIN, SUN YW, LI HB, SONG WW, ZHANGY, YAOG. Comparison of the carcass and beef quality traits with the expression of the lipid metabolism related genes between Xinjiang brown cattle and Angus beef cattle Scientia Agricultura Sinica, 2020(22):4700-4709. (in Chinese) [本文引用: 1]
WU YQ, CHEN SK, SHENG XH, QI XL, WANG XG, NI HM, GUOY, WANG CD, XINGK. Differential expression of mRNA and lncRNA in longissimus dorsi muscle of Songliao black pig and Landrace pig based on high-throughput sequencing technique Scientia Agricultura Sinica, 2020(4):836-847. (in Chinese) [本文引用: 1]
XU YF, WANW, SHOUX, HUANGR, YOU ZY, SHOU YH, WANG LL, ZHOU TH, LIUW. TP53INP2/DOR, a mediator of cell autophagy, promotes rDNA transcription via facilitating the assembly of the POLR1/RNA polymerase I preinitiation complex at rDNA promoters Autophagy, 2016, 12(7):1118-1128. doi: https://www.chinaagrisci.com/article/2021/0578-1752/10.1080/15548627.2016.1175693. URL [本文引用: 1]
KOVSANJ, BLÜHERM, TARNOVSCKIT, KLÖTINGN, KIRSHTEINB, MADARL, SHAII, GOLANR, HARMAN-BOEHM I, SCHÖNM R, GREENBERGA S, ELAZARZ, BASHANN, RUDICHA. Altered autophagy in human adipose tissues in obesity The Journal of Clinical Endocrinology and Metabolism, 2011, 96(2):E268-E277. doi: https://www.chinaagrisci.com/article/2021/0578-1752/10.1210/jc.2010-1681. URL [本文引用: 1]
FROMM-DORNIEDENC, LYTOVCHENKOO, VONDER HEYDE S, BEHNKEN, HOGLS, BERGHOFFJ, KÖPPERF, OPITZL, RENNEU, HOEFLICHA, BEISSBARTHT, BRENIGB, BAUMGARTNERB G. Extrinsic and intrinsic regulation of DOR/TP53INP2 expression in mice: effects of dietary fat content, tissue type and sex in adipose and muscle tissues Nutrition & Metabolism, 2012, 9(1):86. doi: https://www.chinaagrisci.com/article/2021/0578-1752/10.1186/1743-7075-9-86. [本文引用: 1]
ZHAO YF. The effect of Rybp gene on the differentiation of Qinchuan bovine myoblasts [D]. Yangling: Northwest A & F university.2017. (in Chinese) [本文引用: 1]
LI PW, WUS, WANG YN, WANG HB, ZAN LS. Expression patterns of PLIN2 gene in subcutaneous adipose tissue and association analysis of single nucleotide polymorphisms with meat quality traits in Qinchuan cattle China Animal Husbandry & Veterinary Medicine, 2018(6):1580-1589. (in Chinese) [本文引用: 1]
MA YL. Genetic variation of cattle IGF1R gene and its effect on the proliferation and differentiation of myoblasts [D]. Yangling: Northwest A & F university.2019. (in Chinese) [本文引用: 1]
WANG XY, SHI TP, ZHAO ZD, HU WP, SHANG MY, ZHANGL. The analysis of PI3K-AKT signal pathway based on the proteomic results of sheep embryonic skeletal muscle Scientia Agricultura Sinica, 2020(14):2956-2963. (in Chinese) [本文引用: 1]
KIM JH, CHOI TG, PARKS, YUN HR, NGUYENN N Y, JOY H, JANGM, KIMJ, KIMJ, KANGI, HAJ, MURPHYM P, TANGD G, KIMS S. Mitochondrial ROS-derived PTEN oxidation activates PI3K pathway for mTOR-induced myogenic autophagy Cell Death and Differentiation, 2018, 25(11):1921-1937. doi: https://www.chinaagrisci.com/article/2021/0578-1752/10.1038/s41418-018-0165-9. URL [本文引用: 1]
 ,1, 杜鑫泽1, 杨昕冉1, 宋贵兵1, 赵慧1, 昝林森1,2, 王洪宝
,1, 杜鑫泽1, 杨昕冉1, 宋贵兵1, 赵慧1, 昝林森1,2, 王洪宝

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT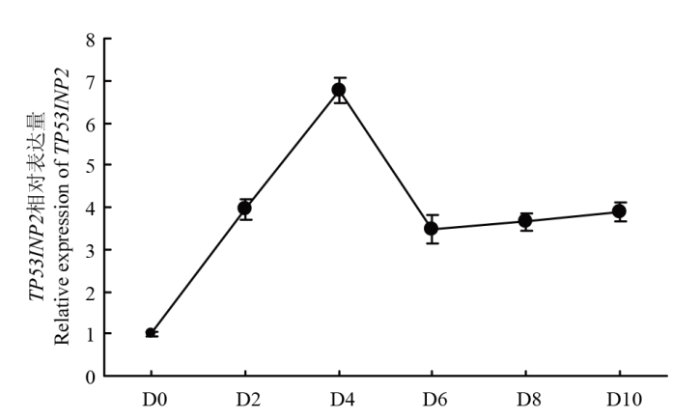 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT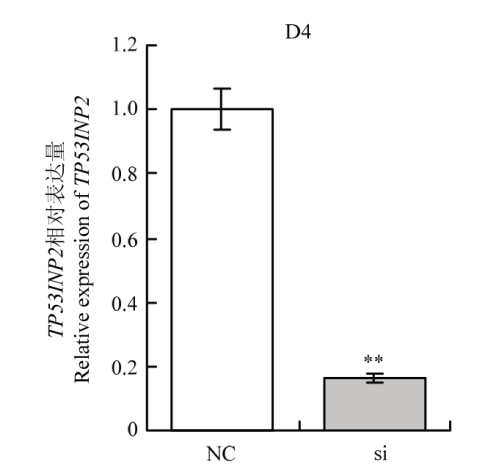 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT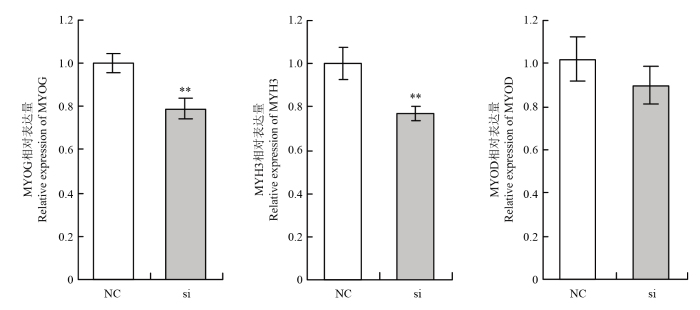 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT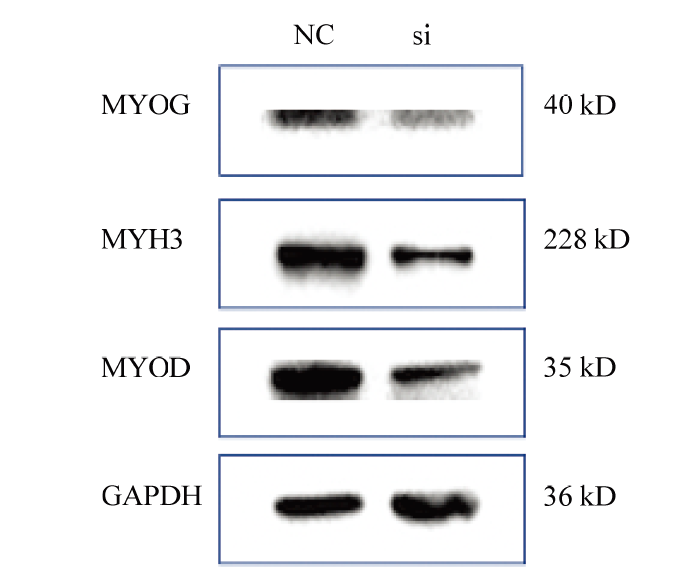 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT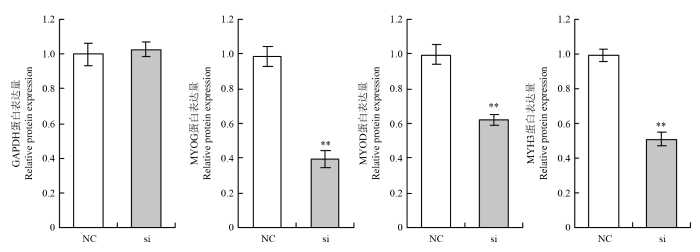 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}