IbMKP6, A Mitogen-Activated Protein Kinase, Confers Low Temperature Tolerance in Sweetpotato
JIN Rong,, LIU Ming, ZHAO Peng, ZHANG QiangQiang, ZHANG AiJun, TANG ZhongHou,Xuzhou Institute of Agricultural Sciences of Xuhuai District of Jiangsu Province/Xuzhou Sweetpotato Research Center of Jiangsu Province/Key Laboratory of Sweetpotato Biology and Genetic Breeding, Ministry of Agriculture, Xuzhou 221131, Jiangsu
Abstract 【Objective】Studying the function of mitogen activated protein kinase (MAPK) IbMPK6 in respect of low temperature stress tolerance in sweetpotato, that will help us to understand the mechanism of adaption to low temperature stress and play a fundamental role in molecular breeding of sweetpotato. 【Method】Agrobacterium tumefaciens strain EHA105 harbored the plasmid 35S::IbMPK6-GFP were transformed intosweetpotato cv. Xushu29 embryogenic callus. Molecular examination and qRT-PCR were used to screen and select transgenic lines. For low temperature stress assay, selected transgenic lines were performed to observe the phenotype and determine the physiological indexes such as Fv/Fm, the content of malondialdehyde (MDA) and hydrogen peroxide (H2O2) after low temperature treatment and recovery treatment. Diaminobenzidine (DAB) staining and nitro blue tetrazolium (NBT) staining analysis were performed to observe reactive oxygen species (ROS) accumulation. The expression level of the key transcription factor IbCBF3 and downstream gene IbCOR27 involved in low temperature signal transduction pathway were identified before and after low temperature treatment. 【Result】Twelve transgenic lines were generated and three transgenic lines (L3, L8 and L11) with a high expression level of IbMPK6 were selected for low temperature tolerance assay. Under low temperature stress, the level of Fv/Fm in transgenic lines L3, L8 and L11 was 0.79, 0.79 and 0.80, while that in WT was 0.05. After temperature recovery treatment, Fv/Fm in transgenic lines has recovered to former levels, whereas the level of Fv/Fm in WT was only 0.70, which was significantly lower than that in transgenic lines. MDA content of three transgenic (lines L3, L8 and L11) increased by 0.02, 0.04 and 0.02 μmol·g-1, and it of WT increased by 0.05 μmol·g-1 after low temperature stress treatment, respectively. After recovery treatment, MDA content in transgenic lines was 0.01 μmol·g-1 on average, whereas it of WT was 0.03 μmol·g-1. The results of DAB and NBT staining showed that the leaves of WT were stained deeper than those of transgenic lines, indicated that hydrogen peroxide and superoxide anion were accumulation less in transgenic lines than in WT. Furthermore, H2O2 level in WT was significantly higher than that in transgenic lines under low temperature stress condition and after recovery treatment. Low temperature regulated the expression level of IbCBF3 and IbCOR27 genes, but the expression level in transgenic lines was higher than that in WT. 【Conclusion】Overexpression of IbMPK6 in sweetpotato resulted in enhanced tolerance to low temperature stress, via alleviating the damage of membrane and photosynthetic system, and decreasing ROS accumulation. IbMPK6 involved in low temperature signaling transduction pathway by up-regulating the expression level of cold related genes IbCBF3 and IbCOR27. Keywords:sweetpotato;IbMPK6;transgenic lines;low temperature stress
PDF (1109KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 靳容, 刘明, 赵鹏, 张强强, 张爱君, 唐忠厚. 甘薯丝裂原活化蛋白激酶MPK6对低温胁迫的响应. 中国农业科学, 2021, 54(20): 4265-4273 doi:10.3864/j.issn.0578-1752.2021.20.002 JIN Rong, LIU Ming, ZHAO Peng, ZHANG QiangQiang, ZHANG AiJun, TANG ZhongHou. IbMKP6, A Mitogen-Activated Protein Kinase, Confers Low Temperature Tolerance in Sweetpotato. Scientia Acricultura Sinica, 2021, 54(20): 4265-4273 doi:10.3864/j.issn.0578-1752.2021.20.002
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】温度是植物生长发育过程中最重要的环境因素之一,不仅影响农作物的生长发育和产量,也制约农作物的耕作时间和地理分布[1]。甘薯(Ipomoea batatas (L.) Lam.)是世界上第七大粮食作物,然而,不论苗期生长还是收获后储藏都对低温都非常敏感,甘薯的不耐低温性严重制约了甘薯产业的发展[2,3]。克隆和低温应答相关的基因并阐释其功能,对开展基因靶向育种和改良甘薯耐低温胁迫的能力具有重要的理论意义。【前人研究进展】有丝分裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)是广泛存在于真核生物中的信号分子,由MAPKKK、MAPKK和MAPK组成,通过级联依次磷酸化反应方式参与胞外信号的放大和胞内信号的传递,参与生物或非生物的胁迫反应、激素反应、细胞分裂、生长发育与程序性凋亡等[4,5,6]。目前仅有少数的MAPK级联反应被详细报道,其中,拟南芥MEKKK1-MKK2-MPK4/MPK6级联反应和水稻OsMKK6-OsMPK3级联反应参与植物低温信号转导途径[7,8]。异源或同源过表达MAPK级联信号中的激酶可以提高植物耐低温性。突变MKK2的磷酸化位点,在拟南芥中过表达该突变基因MKK2DD,抑制其蛋白磷酸化活性,提高了拟南芥的耐低温性[9];在烟草中分离到一个拟南芥MAPKKK的同源基因NPK1,分别在烟草和玉米中过表达该基因,可以提高烟草和玉米的耐低温性[10,11];在拟南芥中异源表达玉米ZmMKK4,可以提高拟南芥耐低温和耐盐性[12];在烟草中过表达马铃薯SlMPK3或玉米ZmMPK17,可通过增加可溶性糖和脯氨酸含量,增加烟草耐低温性[13,14]。拟南芥中共有20个MAPK,目前,研究较为详尽的MAPK激酶有MPK3、MPK4和MPK6。根据序列磷酸化位点的保守性,MPK6和MPK3序列最为相近,功能也相似,同属于A类MAPK,均参与植物生长发育并响应多种生物和非生物胁迫[4,15]。在拟南芥中,MPK6响应茉莉酸和乙烯信号传导、参与病原菌应答、调控气孔发育、参与植物抗毒素的合成等。低温胁迫、渗透胁迫和盐胁迫等非生物胁迫均能够诱导拟南芥、水稻和玉米等多种植物中MPK6的表达[16,17,18]。在甘薯中,IbMPK6含有11个保守的亚结构域和磷酸化基序TEY。IbMPK6激酶活性可以被早期的NaCl、SA、H2O2和ABA诱导。在烟草中瞬时过表达IbMPK6,可以提高病原相关(protection related,PR)基因的表达水平,增强叶片对细菌病原菌的耐受性[19]。以上数据表明IbMPK6在调控甘薯逆境胁迫的反应过程中可能起着重要的作用。【本研究切入点】低温胁迫严重影响甘薯产业的发展,但有关甘薯抵御低温胁迫生理和分子机制方面的研究却很少。前人研究发现低温胁迫下IbMPK6的表达随着时间的变化先增后降,在处理30 min时,相对表达量达到顶峰,比处理前增加2.5倍,说明IbMPK6可能参与调控低温信号转导途径[19]。【拟解决的关键问题】本研究通过在甘薯中过表达IbMPK6获得甘薯转基因株系,开展甘薯耐低温性鉴定工作,明确IbMPK6响应低温胁迫的作用,为改良及培育甘薯耐低温品种提供思路。
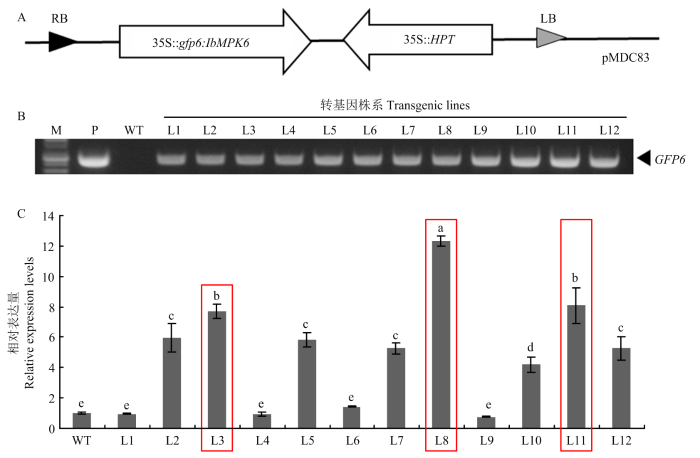
A:IbMPK6过表达载体结构示意图。B:IbMPK6过表达株系PCR鉴定。M:Marker;P:质粒;WT:非转基因对照植株,L1—L12:转基因株系。C:qRT-PCR检测筛选IbMPK6过表达株系。不同小写字母表示在P<0.01水平差异显著。红色边框内表示IbMPK6过表达最高的3个株系L3、L8和L11 Fig. 1Generation of IbMPK6-overexpression transgenic sweetpotato plants
A: Schematic diagram of IbMPK6-overexprssing constructs. B: Genomic DNA PCR analysis of IbMPK6-overexpressing transgenic sweetpotato lines. M: Marker; P: Plasmid; WT: Wide type; L1-L12: Transgenic plants. C: qRT-PCR analysis of IbMPK6-overexpressing transgenic sweetpotato lines. Lower-case letters indicated significantly different at P<0.01. The three lines (L3, L8 and L11) with the highest expression level of IbMPK6 were marked with red border
A:表型分析;B:PSⅡ最大光化学量子产量;C:丙二醛含量。*和**分别表示在P<0.05和P<0.01水平差异显著 Fig. 2Low temperature-resistance detection of IbMPK6-overexpressing sweetpotato
A: Phenotype analysis; B: Optimal/maximal quantum yield of PSⅡ; C: MDA content. * and ** indicated significantly different at P<0.05 and P<0.01, respectively
XIEZ Y. Identifies chilling responsive miRNAs and their targets in sweetpotato (Ipomoea batatas Lam.) during storage [D]. Xuzhou: Jiangsu Normal University, 2017. (in Chinese) [本文引用: 1]
MAD F, LIQ, CAOQ H, NIUF X, XIEY P, TANGJ, LIH M. Development and prospect of sweetpotato industry and its technologies in China Jiangsu Journal of Agricultural Sciences, 2012, 28(5):969-973. (in Chinese) [本文引用: 2]
MIZOGUCHIT, ICHIMURAK, SHINOZAKIK. Environmental stress response in plants: The role of mitogen-activated protein kinases Trends in Biotechnology, 1997, 15(1):15-19. DOI:10.1016/S0167-7799(96)10074-3URL [本文引用: 1]
DEZ A, COLCOMBETJ, HIRTH. The role of MAPK modules and ABA during abiotic stress signaling Trends in Plant Science, 2016, 21(8):677-685. DOI:10.1016/j.tplants.2016.04.004URL [本文引用: 1]
FURUYAT, MATSUOKAD, NANMORIT. Membrane rigidification functions upstream of the MEKK1-MKK2-MPK4 cascade during cold acclimation in Arabidopsis thaliana FEBS Letters, 2014, 588(11):2025-2030. DOI:10.1016/j.febslet.2014.04.032URL [本文引用: 1]
XIEG, KATOH, IMAIR. Biochemical identification of the OsMKK6-OsMPK3 signalling pathway for chilling stress tolerance in rice Biochemical Journal, 2012, 443(1):95-102. DOI:10.1042/BJ20111792URL [本文引用: 1]
TEIGEM, SCHEIKLE, EULGEMT, DOCZIF, ICHIMURAK, SHINOZAKIK, DANGLJ L, HIRTH. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis Molecular Cell, 2004, 15(1):141-152. DOI:10.1016/j.molcel.2004.06.023URL [本文引用: 1]
SHOUH, BORDALLOP, FANJ B, YEAKLEYJ M, BIBIKOVAM, SHEENJ, WANGK. Expression of an active tobacco mitogen- activated protein kinase kinase kinase enhances freezing tolerance in transgenic maize Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(9):3298-3303. [本文引用: 1]
KOVTUNY, CHIUW L, TENAG, SHEENJ. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(6):2940-2945. [本文引用: 1]
KONGX P, PANJ W, ZHANGM Y, XINGX, ZHOUY, LIUY, LID P, LID Q. ZmMKK4, a novel group C mitogen-activated protein kinase kinase in maize (Zea mays), confers salt and cold tolerance in transgenic Arabidopsis Plant, Cell & Environment, 2011, 34(8):1291-1303. [本文引用: 1]
YUL, YANJ, YANGY, ZHUW M. Overexpression of tomato mitogen-activated protein kinase SlMPK3 in tobacco increases tolerance to low temperature stress Plant Cell, Tissue and Organ Culture, 2015, 121(1):21-34. DOI:10.1007/s11240-014-0675-1URL [本文引用: 1]
PANJ, ZHANGM, KONGX, XINGX, LIUY K, ZHOUY, LIUY, SUNL P, LID Q. ZmMPK17, a novel maize group D MAP kinase gene, is involved in multiple stress responses Planta, 2012, 235(4):661-676. DOI:10.1007/s00425-011-1510-0URL [本文引用: 1]
DROILLARDM, BOUDSOCQM, BARBIER-BRYGOOH, LAURIERE,C. Different protein kinase families are activated by osmotic stresses in Arabidopsis cell suspensions: Involvement of the MAP kinase AtMPK3 and AtMPK6 FEBS Letters, 2002, 527:43-50. DOI:10.1016/S0014-5793(02)03162-9URL [本文引用: 1]
LIUY, ZHANGD, WANGL, LID Q. Genome-wide analysis of mitogen-activated protein kinase gene family in maize Plant Molecular Biology Reporter, 2013, 31(6):1446-1460. DOI:10.1007/s11105-013-0623-yURL [本文引用: 1]
XIEG, KATOH, SASAKIK, IMAIR. A cold-induced thioredoxin h of rice, OsTrx23, negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro FEBS Letters, 2009, 583(17):2734-2738. DOI:10.1016/j.febslet.2009.07.057URL [本文引用: 1]
KIMH S, PARKS C, JIC Y, PARKS, JEONGJ C, LEEH S, KWAKS S. Molecular characterization of biotic and abiotic stress-responsive MAP kinase genes, IbMPK3 and IbMPK6, in sweetpotato Plant Physiology & Biochemistry, 2016, 108:37-48. [本文引用: 2]
HODGESD M, DELONGJ M, PRANGEF. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds Planta, 1999, 207(4):604-611. DOI:10.1007/s004250050524URL [本文引用: 1]
ZHOUB, WANGJ, GUOZ, TANH Q, ZHUX C. A simple colorimetric method for determination of hydrogen peroxide in plant tissues Plant Growth Regulation, 2006, 49(2/3):113-118. DOI:10.1007/s10725-006-9000-2URL [本文引用: 1]
BAXTERA, MITTLERR, SUZUKIN. ROS as key players in plant stress signalling Journal of Experimental Botany, 2014, 65(5):1229-1240. DOI:10.1093/jxb/ert375URL [本文引用: 1]
Jiangsu Academy of Agricultural Sciences. Sweetpotato Cultivation in China. 12th ed. Shanghai: Shanghai Science and Technology Press, 1984. (in Chinese) [本文引用: 1]
KIMY H, KIMM D, PARKS C, YANGK S, JEONGJ C, LEEH S, KWAKS S. SCOF-1-expressing transgenic sweetpotato plants show enhanced tolerance to low-temperature stress Plant Physiology Biochemistry, 2011, 49(12):1436-1441. DOI:10.1016/j.plaphy.2011.09.002URL [本文引用: 1]
JIC Y, JINR, XUZ, KIMH S, LEEC J, KANGL, KIMS E, LEEH U, LEEJ S, KANGC H, CHIY H, LEES Y, XIEY P, LIH M, MAD F, KWAKS S. Overexpression of Arabidopsis P3B increases heat and low temperature stress tolerance in transgenic sweetpotato BMC Plant Biology, 2017, 17:139. DOI:10.1186/s12870-017-1087-2URL [本文引用: 1]
GONGZ Z, XIONGL M, SHIH Z, YANGS H, HERRERA- ESTRELLAL R, XUG H, CHAOD Y, LIJ R, WANGP Y, QINF, LIJ J, DINGY L, SHIY T, WANGY, TANGY Q, GUOY, ZHUJ K. Plant abiotic stress response and nutrient use efficiency Science China-Life sciences, 2020, 63(5):635-674. DOI:10.1007/s11427-020-1683-xURL [本文引用: 1]
HUW H, WUY, ZENGJ Z, HEL, ZENGQ M. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery Photosynthetica, 2010, 48(4):537-544. DOI:10.1007/s11099-010-0071-yURL [本文引用: 1]
THEOCHARISA, CLÉMENTC, BARKAE A. Physiological and molecular changes in plants grown at low temperatures Planta, 2012, 35(6):1091-1105. [本文引用: 1]
VOLKOVR A, PANCHUKI I, MULLINEAUXP M, SCHÖFFLF. Heat stress-induced H2O2 is required for effective expression of heat shock genes in Arabidopsis Plant Molecular Biology, 2006, 61(4/5):733-746. DOI:10.1007/s11103-006-0045-4URL [本文引用: 1]
SHIY T, DINGY L, YANGS H. Cold signal transduction and its interplay with phytohormones during cold acclimation Plant Cell Physiology, 2015, 56(1):7-15. DOI:10.1093/pcp/pcu115URL [本文引用: 1]
CHINNUSAMYV, OHTAM, KANRARS, LEEB H, HONGX, AGARWALM, ZHUJ K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis Genes Development, 2003, 17(8):1043-1054. DOI:10.1101/gad.1077503URL [本文引用: 1]
DONGM A, FARRÉE M, THOMASHOWM F. Circadian clock associated 1 and late elongated hypocotyl regulate expression of the C-repeat binding factor (CBF) pathway in Arabidopsis Proceedings of the National Academy Sciences of the United States of America, 2011, 108(17):7241-7246. [本文引用: 1]
JIANGB C, SHIY T, ZHANGX Y, XINX Y, QIL J, GUOH W, LIJ W, YANGS H. PIF3 is a negative regulator of the CBF pathway and freezing tolerance in Arabidopsis Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(32):E6695-E6702. [本文引用: 1]
KIMW Y, FUJIWARAS, SUHS S, KIMJ, KIMY, HANL, DAVIDK, PUTTERILLJ, NAMH G, SOMERSD E. ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light Nature, 2007, 449(7160):356-360. DOI:10.1038/nature06132URL [本文引用: 1]
NORÉNL, KINDGRENP, STACHULAP, RÜHLM, ERIKSSONM E, HURRYV, STRANDÅ. HSP90, ZTL, PRR5 and HY5 integrate circadian and plastid signaling pathways to regulate CBF and COR expression Plant Physiology, 2016, 171:1392-1406. [本文引用: 1]
JINR, KIMB H, JIC Y, KIMH S, MAD F, KWAKS S. Overexpressing IbCBF3 increases low temperature and drought stress tolerance in transgenic sweetpotato Plant Physiology and Biochemistry, 2017, 118:45-54. DOI:10.1016/j.plaphy.2017.06.002URL [本文引用: 1]
PITZSCHKEA, HIRTH. Mitogen-activated protein kinase and reactive oxygen species signaling in plants Plant Physiology, 2006, 141(2):351-356. DOI:10.1104/pp.106.079160URL [本文引用: 1]
NISHIHAMAR, BANNOH, KAWAHARAE, IRIEK, MACHIDAY. Possible involvement of differential splicing in regulation of the activity of Arabidopsis ANP1 that is related to mitogen-activated protein kinase kinase kinases (MAPKKKs) The Plant Journal, 1997, 12(1):39-48. DOI:10.1046/j.1365-313X.1997.12010039.xURL [本文引用: 1]
LIH, DINGY L, SHIY T, ZHANGX Y, ZHANGS Q, GONGZ Z, YANGS H. MPK3-and MPK6-mediated ICE1 phosphorylation negatively regulates ICE1 stability and freezing tolerance in Arabidopsis Developmental Cell, 2017, 43(5):630-642. DOI:10.1016/j.devcel.2017.09.025URL [本文引用: 1]
ZHANGZ Y, LIJ H, LIF, LIUH H, YANGW, CHONGK, XUY Y. OsMAPK3 phosphorylates OsbHLH002/OsICE1 and inhibits its ubiquitination to activate OsTPP1 and enhances rice chilling tolerance Developmental Cell, 2017, 43(6):731-743. DOI:10.1016/j.devcel.2017.11.016URL [本文引用: 1]
 ,, 刘明, 赵鹏, 张强强, 张爱君, 唐忠厚
,, 刘明, 赵鹏, 张强强, 张爱君, 唐忠厚

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT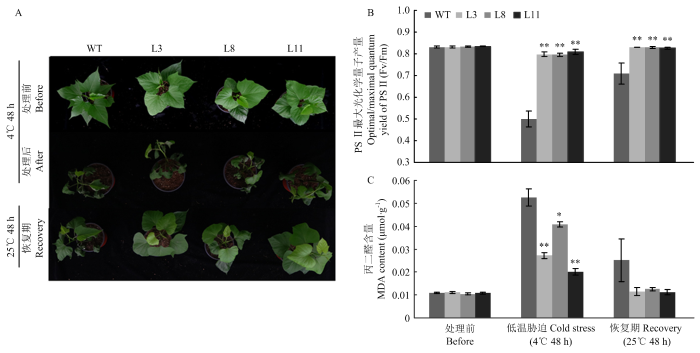 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT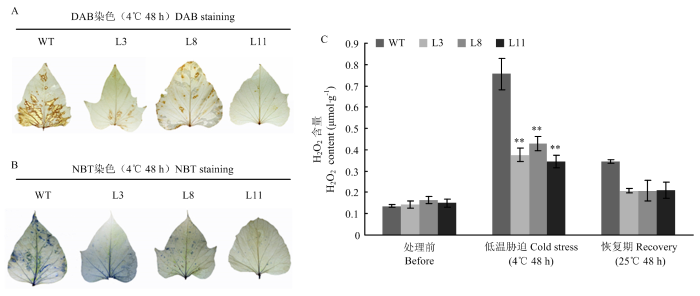 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT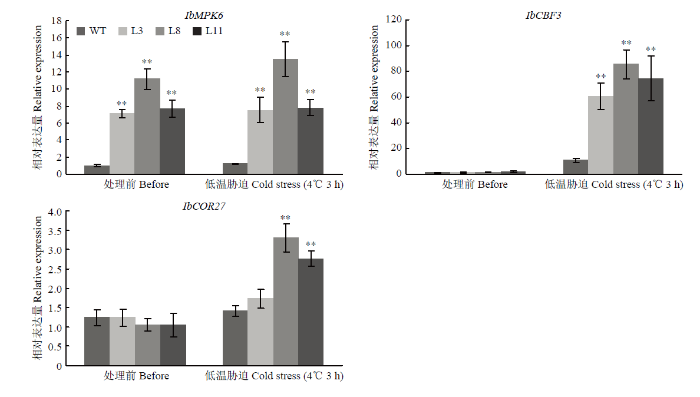 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}