Abstract 【Background】 Branching is one of the key yield components, which plays an important role in alfalfa (Medicago sativa L.) breeding. Exploring and functional characterization of key branching-related genes are of significance in accelerating breeding of alfalfa with high yield and quality. MAX2 is an important branching-related gene, which is involved in the regulation of branching in several plant species. 【Objective】 Our research on the functional characterization of MsMAX2 in alfalfa will lay a foundation for the molecular mechanism of MsMAX2 in regulating branch development in alfalfa. 【Method】 The gene sequence of MAX2 in alfalfa was isolated by using homologous cloning. Sequence characteristics and phylogenetic tree of MsMAX2 were analyzed by using bioinformatics tools including Expy Protparatam, DNAMAN, and MEGA-X. The real-time quantitative PCR (qPCR) was applied to detect the tissue-specific expression pattern of MsMAX2 in alfalfa. The subcellular localization of the MsMAX2 protein was determined by using transient expression system in tobacco. The biological function of MsMAX2 was clarified by transformation in the Arabidopsis mutant via Agrobacterium-mediated transformation. Proteins interacting with MsMAX2 were determined by using yeast two-hybrid assay. 【Result】The length of MsMAX2 CDS is 2 136 bp, encoding a protein of 711 amino acids, and it belongs to the F-box protein super-family. Phylogenetic analysis showed that the evolution of MAX2 homologs was highly similar to the differentiation of species, indicating that MsMAX2 was a functionally conserved gene. It was showed that MsMAX2 was expressed in the neck at the highest level, followed by in the leaves of seedling, the inflorescences on pollination day and the roots; the expression level of MsMAX2 was relatively low in other tissues, indicating it functions in multiple tissues. Subcellular localization assay showed that the MsMAX2 protein was localized in the nucleus. Complementation assay in Arabidopsis max2 mutant showed that the multi-branch phenotype was recovered by the ectopic expression of MsMAX2. Yeast two hybrid assay demonstrated that the interaction between MsMAX2 and hormone receptor D14 depended on the existence of strigolactones. 【Conclusion】The MsMAX2 was obtained from alfalfa and it was highly expressed in the neck and the encoding MsMAX2 protein was localized in nucleus. When the MsMAX2 was over-expressed in the Arabidopsis max2 mutant, its multi-branch phenotype was recovered, indicating that MsMAX2 regulates branch development in alfalfa plant, and its function was conserved. Keywords:alfalfa;MsMAX2;branching;complementation assay;yeast two hybrid assay
PDF (4431KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 马琳, 温红雨, 王学敏, 高洪文, 庞永珍. 紫花苜蓿MsMAX2的克隆及功能研究. 中国农业科学, 2021, 54(19): 4061-4069 doi:10.3864/j.issn.0578-1752.2021.19.003 MA Lin, WEN HongYu, WANG XueMin, GAO HongWen, PANG YongZhen. Cloning and Function Analysis of MsMAX2 Gene in Alfalfa (Medicago sativa L.). Scientia Acricultura Sinica, 2021, 54(19): 4061-4069 doi:10.3864/j.issn.0578-1752.2021.19.003
使用改良CTAB法提取紫花苜蓿的DNA,经Nanodrop检测质量后,稀释至50 ng·μL-1备用。收集紫花苜蓿苗期、分枝期、开花期、荚果期的13个不同组织,使用Eastep® Super Total RNA Extraction Kit试剂盒(上海普洛麦格生物科技有限公司)提取总RNA。使用TransScript Green One-Step qRT-PCR mix(北京全式金生物科技有限公司)将4 μg的总RNA反转录为cDNA,稀释8倍后备用。
AtMAX2:拟南芥,NP_565979.1;GmMAX2:大豆,XP_003540983.1;RMS4:豌豆,ABD67495.1;CaMAX2:鹰嘴豆,XP_004505491.1;TpMAX2:红三叶,PNX93440.1;MtMAX2:蒺藜苜蓿,XP_003607592.1;OsD3:水稻,NP_001174608.1;BdMAX2:短柄草,XP_003564315.1;TaMAX2:小麦,AZS54115.1;ZmMAX2:玉米,AQL02030.1;SbMAX2:高粱,XP_002436499.1;SiMAX2:谷子,XP_004964817.1。红色阴影为双子叶植物,蓝色阴影为单子叶植物 Fig. 2Phylogenetic analysis of MAX2 proteins from 13 plant species
AtMAX2: Arabidopsis thaliana, NP_565979.1; GmMAX2: Glycine max, XP_003540983.1; RMS4: Pisum sativum, ABD67495.1; CaMAX2: Cicer arietinum, XP_004505491.1; TpMAX2: Trifolium pratense, PNX93440.1; MtMAX2: Medicago truncatula, XP_003607592.1; OsD3: Orzya sativa, NP_001174608.1; BdMAX2: Brachypodium distachyon, XP_003564315.1; TaMAX2: Triticum aestivum, AZS54115.1; ZmMAX2: Zea may, AQL02030.1; SbMAX2: Sorghum bicolor, XP_002436499.1; SiMAX2: Setaria italica, XP_004964817.1. Proteins for dicotyledons are highlighted with red shadow, and monocotyledons with blue shadow
1:苗期叶片;2:苗期根;3:苗期颈部;4:苗期枝;5:苗期顶部;6:分枝期茎;7:分枝期叶片;8:分枝期顶部;9:开花期茎;10:开花期叶片;11:开花期花序;12:授粉当天的花序;13:授粉5 d的果荚 Fig. 3Expression levels of MsMAX2 in various tissues
1: Leaf at seedling stage; 2: Root at seedling stage; 3: Neck at seedling stage; 4: Branch at seedling stage; 5: Head at seedling stage; 6: Stem at branching stage; 7: Leaf at branching stage; 8: Head at branching stage; 9: Stem at flowering stage; 10: Leaf at flowering stage; 11: Inflorescence at flowering stage; 12: Capsule at 0 day after pollination; 13: Capsule at 5 day after pollination
野生型、max2及max2背景下过表达MsMAX2拟南芥植株的表型(A)、基因表达量(B)和分枝数(C) Fig. 5Complementation assays of MsMAX2 in the Arabidopsis max2 mutant
The phenotypes (A), MsMAX2 and AtMAX2 relatively expression levels (B) and brunch numbers (C) of WT, max2 and max2 transformed with 35S::MsMAX2 constructs
HONGB Z. Alfalfa Science. Beijing: China Agriculture Press, 2009. (in Chinese) [本文引用: 1]
RADOVIĆJ, SOKOLOVIĆD, MARKOVIĆJ J B A H. Alfalfa-most important perennial forage legume in animal husbandry Biotechnology in Animal Husbandry, 2009, 25:465-475. DOI:10.2298/BAH0906465RURL [本文引用: 1]
BAI ZH, MA WQ, MAL, VELTHOF GL, WEI ZB, HAVLÍKP, OENEMAO, LEE M RF, ZHANGF S. China’s livestock transition: Driving forces, impacts, consequences Science Advances, 2018, 4:eaar8534. DOI:10.1126/sciadv.aar8534URL [本文引用: 1]
ZHANG TJ, YANG QC, KANG JM, SUNY, GUO WS. Advances in genetic basic research of alfalfa breeding for yield Chinese Journal of Grassland, 2011, 33(1):102-106. (in Chinese) [本文引用: 1]
GOU JQ, DEBNATHS, SUNL, FLANAGANA, TANG YH, JIANG QZ, WEN JQ, WANG ZY. From model to crop: Functional characterization of SPL8 in M. truncatula led to genetic improvement of biomass yield and abiotic stress tolerance in alfalfa Plant Biotechnology Journal, 2018, 16(4):951-962. DOI:10.1111/pbi.2018.16.issue-4URL [本文引用: 1]
YAN YY, ZHAON, TANG HM, GONGB, SHI QH. Shoot branching regulation and signaling Plant Growth Regulation, 2020, 92:131-140. DOI:10.1007/s10725-020-00640-1URL [本文引用: 1]
DOMAGALSKA MA, LEYSERO. Signal integration in the control of shoot branching Nature Review, 2011, 12(4):211-221. DOI:10.1038/nrm3088URL [本文引用: 1]
MULLERD, LEYSERO. Auxin, cytokinin and the control of shoot branching Annals of Botany, 2011, 107:1203-1212. DOI:10.1093/aob/mcr069URL [本文引用: 1]
STIRNBERGP, FURNER IJ, LEYSERH M O. MAX2 participates in an SCF complex which acts locally at the node to suppress shoot branching The Plant Journal, 2007, 50(1):80-94. DOI:10.1111/j.1365-313X.2007.03032.xURL [本文引用: 2]
ISHIKAWAS, MAEKAWAM, ARITET, TAKAMUREI, KYOZUKAJ. Suppression of tiller bud activity in tillering dwarf mutants of rice Plant and Cell Physiology, 2005, 46:79-86. DOI:10.1093/pcp/pci022URL [本文引用: 3]
JOHNSONX, BRCICHT, DUN EA, GOUSSOTM, HAUROGNEK, BEVERIDGE CA, RAMEAUC. Branching genes are conserved across species. Genes controlling a novel signal in pea are coregulated by other long-distance signals Plant Physiology, 2006, 142:1014-1026. DOI:10.1104/pp.106.087676URL [本文引用: 1]
STIRNBERGP, VAN DESANDE K, LEYSERH M O. MAX1 and MAX2 control shoot lateral branching in Arabidopsis Development, 2002, 129(5):1131-1141. DOI:10.1242/dev.129.5.1131URL [本文引用: 1]
YAO RF, MING ZH, YAN LM, LI SH, WANGF, MAS, YU CT, YANGM, CHENL, CHEN LH, LI YW, YANC, MIAOD, SUN ZY, YAN JB, SUN YN, WANGL, CHU JF, FAN SL, HEW, DENG HT, NAN FJ, LI JY, RAO ZH, LOU ZY, XIE DX. DWARF14 is a non-canonical hormone receptor for strigolactone Nature, 2016, 536(7617):469-473. DOI:10.1038/nature19073URL [本文引用: 3]
WANGY, SUN SY, ZHU WJ, JIA KP, YANG HQ, WANG XL. Strigolactone/MAX2-induced degradation of brassinosteroid transcriptional effector BES1 regulates shoot branching Developmental Cell, 2013, 27(6):681-688. DOI:10.1016/j.devcel.2013.11.010URL [本文引用: 2]
WANGL, WANGB, JIANGL, LIUX, LI XL, LU ZF, MENG XB, WANG YH, SMITH SM, LI JY. Strigolactone signaling in Arabidopsis regulates shoot development by targeting D53-Like SMXL repressor proteins for ubiquitination and degradation The Plant Cell, 2015, 27(11):3128-3142. DOI:10.1105/tpc.15.00605URL [本文引用: 3]
ZHOUF, LIN QB, ZHU LH, REN YL, ZHOU KN, SHABEKN, WU FQ, MAO HB, DONGW, GANL, MA WW, GAOH, CHENJ, YANGC, WANGD, TAN JJ, ZHANGX, GUO XP, WANG JL, JIANGL, LIUX, CHEN WQ, CHU JF, YAN CY, UENOK, ITOS, ASAMIT, CHENG ZJ, WANGJ, LEI CL, ZHAI HQ, WU CY, WANG HY, ZHENGN, WAN JM. D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling Nature, 2013, 504(7480):406-410. DOI:10.1038/nature12878URL [本文引用: 3]
KAPULNIKY, KOLTAIH. Strigolactone involvement in root development, response to abiotic stress, and interactions with the biotic soil environment Plant Physiology, 2014, 166(2):560-569. DOI:10.1104/pp.114.244939URL [本文引用: 2]
WOO HR, CHUNG KM, PARK JH, OH SA, AHNT, HONG SH, JANG SK, NAM HG. ORE9, an F-box protein that regulates leaf senescence in Arabidopsis The Plant Cell, 2001, 13:1779-1790. DOI:10.1105/TPC.010061URL [本文引用: 2]
WATERS MT, SMITH SM, NELSON DC. Smoke signals and seed dormancy Plant Signaling & Behavior, 2011, 6:1418-1422. [本文引用: 1]
NELSON DC, SCAFFIDIA, DUN EA, WATERS MT, FLEMATTI GR, DIXON KW, BEVERIDGE CA, GHISALBERTI EL, SMITH SM. F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana Proceedings of the National Academy of Sciences of the USA, 2011, 108:8897-8902. [本文引用: 1]
BU QY, LV TX, SHENH, LUONGP, WANGJ, WANG ZY, HUANG ZG, XIAO LT, ENGINEERC, KIM TH, SCHROEDER JI, HUQE. Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis Plant Physiology, 2014, 164:424-439. DOI:10.1104/pp.113.226837URL [本文引用: 1]
KALLIOLAM, JAKOBSONL, DAVIDSSONP, PENNANENV, WASZCZAKC, YARMOLINSKYD, ZAMORAO, PALVA ET, KARIOLAT, KOLLISTH, BROSCHÉM. Differential role of MAX2 and strigolactones in pathogen, ozone, and stomatal responses Plant Direct, 2020, 4(2):1-14. [本文引用: 1]
SHENH, ZHUL, BU QY, HUQE. MAX2 affects multiple hormones to promote photomorphogenesis Molecular Plant, 2012, 5(3):750-762. DOI:10.1093/mp/sss029URL [本文引用: 1]
YANG QC, KANG JM, ZHANG TJ, LIU FQ, LONG RC, SUNY. Distribution, breeding and utilization of alfalfa germplasm resources Chinese Science Bulletin, 2016, 61(2):261-270. (in Chinese) [本文引用: 1]
SHI SL, NAN LL, SMITH KF, FORSTER JW. The current status, problems, and prospects of alfalfa (Medicago sativa L.) breeding in China Agronomy, 2017, 7(1):1. DOI:10.3390/agronomy7010001URL [本文引用: 1]
CAPSTAFF NM, MILLER AJ. Improving the yield and nutritional quality of forage crops Frontiers in Plant Science, 2018, 9:535. DOI:10.3389/fpls.2018.00535URL [本文引用: 1]
CHEN HT, ZENGY, YANG YZ, HUANG LL, TANG BL, ZHANGH, HAOF, LIUW, LI YH, LIU YB, ZHANG XS, ZHANGR, ZHANG YS, LI YX, WANGK, HEH, WANG ZK, FAN GY, YANGH, BAO AK, SHANG ZH, CHEN JH, WANGW, QIUQ. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa Nature Communications, 2020, 11:2494. DOI:10.1038/s41467-020-16338-xURL [本文引用: 1]
SHENC, DU HL, CHENZ, LU HW, ZHU FG, MENG HC, XZ, LIUQ W, LIUP, ZHENGL H, LIX X, DONGJ L, LIANGC Z, WANGT. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research Molecular Plant, 2020, 13(9):1250-1261. DOI:10.1016/j.molp.2020.07.003URL [本文引用: 1]
 ,, 温红雨, 王学敏, 高洪文, 庞永珍
,, 温红雨, 王学敏, 高洪文, 庞永珍

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT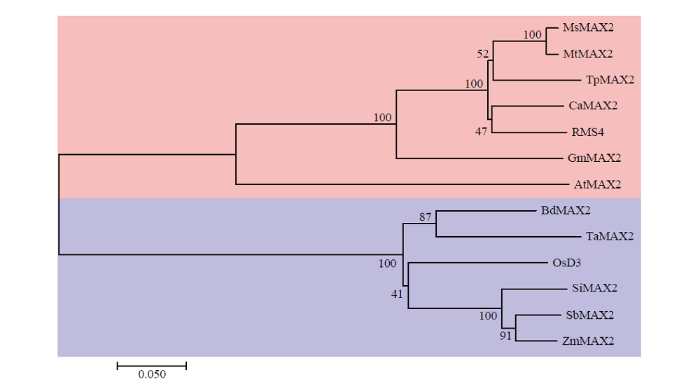 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT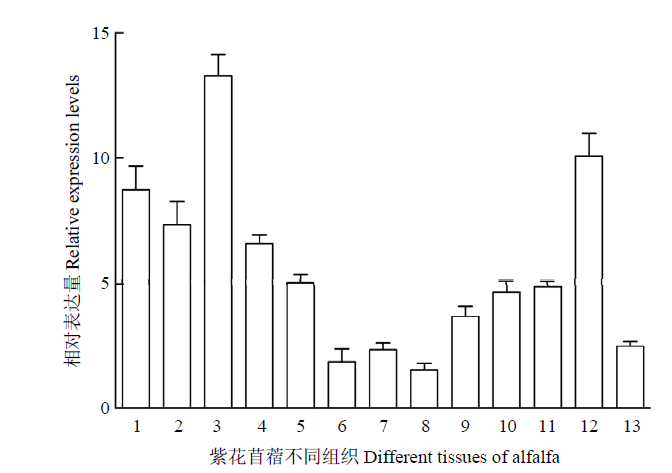 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT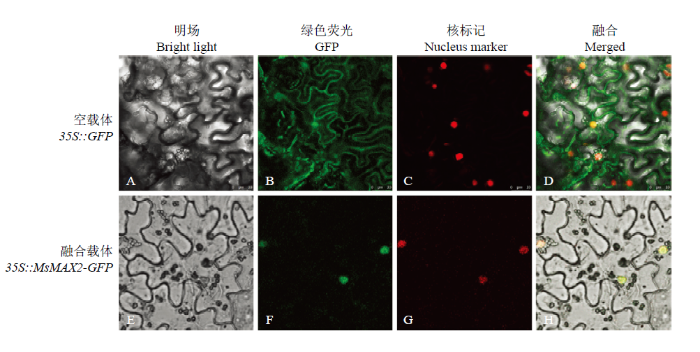 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT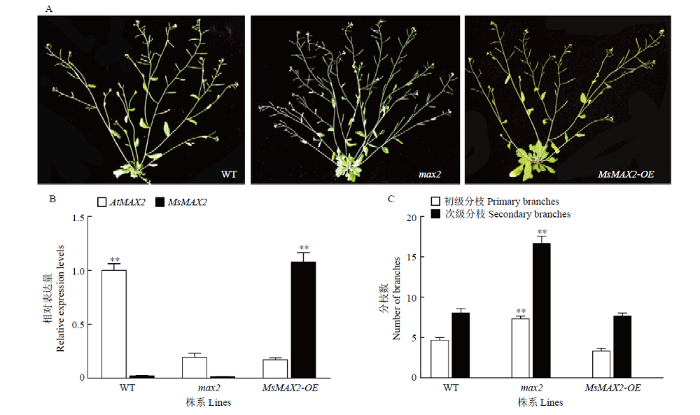 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}