Cytological Characters Analysis and Low-Resolution Mapping of Stripe-Leaf MutantA36-S in Foxtail Millet
ZHANG Shuo,1,2, ZHI Hui1, TANG ChanJuan1, LUO MingZhao1, TANG Sha1, JIA GuanQing1, JIA YanChao1, DIAO XianMin,11Institute of Crop Science, Chinese Academy of Agricultural Science, Beijing 100081 2Food Crop Research Institute, Hubei Academy of Agricultural Science, Wuhan 430064
Abstract 【Objective】 Foxtail millet is a C4 model plant, and its leaf color mutants are good materials for C4 photosynthetic pathway research. Through the cytological characters analysis and gene initial mapping of the stripe-leaf mutant A36-S in foxtail millet, it laid the foundation for cloning the mutant gene, analyzing the chloroplast biogenesis and development, and further understanding the C4 photosynthetic regulation mechanism in foxtail millet. 【Method】 The stripe-leaf mutant of foxtail millet A36-S was naturally mutated from intermediate material A36 created by breeding. Comparing the phenotypic characteristics of A36-S and its isogenic line A36-N, which showed normal phenotypes, and investigating the agronomic traits, such as plant height, leaf width, leaf length, panicle weight, thousand-grain weight, and seed setting rate. To analyze the photosynthetic characters of A36-S, the chlorophyll content, net photosynthetic rate, intercellular CO2 concentration, stomatal conductance, and transpiration rate of A36-S and A36-Nwere determined. By observing the leaf transverse section and ultrathin section of A36-S and the control variety Yugu1, the leaf anatomical structure characters were analyzed, by counting the numbers and areas of the chloroplasts in mesophyll cells and bundle sheath cells respectively, the chloroplast biogenesis and development were assessed. An F2 segregation population of A36-S×SSR41 were created, and genetic analysis was conducted by counting the number of normal phenotype single plant and stripe-leaf single plant in the population. The DNA mixed pools of normal single plants and stripe-leaf single plants of the F2 segregation population were constructed separately, and the method of Bulked Segregation Analysis (BSA) was used to locate the mutant gene. By screening the stripe-leaf plants in the F2 generation using SSR and In-Del markers, the mutant gene were furtherly located.【Result】The stripe-leaf mutant of foxtail millet A36-S showed the phenotype of irregular white stripe-leaf in the whole growth period. Agronomic traits analysis showed that compared with its isogenic line A36-N, A36-S decreased significantly in plant height, leaf width, panicle weight, thousand-grain weight, and setting percentage. Photosynthetic index measurement showed that the chlorophyll contents of A36-S were also reduced significantly, especially the chlorophyll b content declined more severely, additionally, the net photosynthetic rate was also decreased significantly. Observation of the leaf anatomical structure showed that the chloroplasts number and area were significantly lower than that of the contrast Yugu1, while the changes in Kranz structure were not obvious. Furtherly, the ultrastructure of chloroplast was observed and showed that the chloroplast development situation in different cells was quite different. And the leaf cells ofA36-S could be classified into three types: type Ⅰ cells had normal chloroplasts, type Ⅱ cells had chloroplasts with reduced grana and lamellar structures, while type Ⅲ cells had severely abnormal chloroplasts or even had no chloroplast. Genetic analysis suggested that the stripe-leaf trait of A36-S was controlled by a single recessive gene, and the mutant gene was located to the region from 7.66 Mb to 27.90 Mb of chromosome 4 by F2 population. 【Conclusion】 Stripe-leaf mutant of foxtail millet A36-S represented decreased agronomic traits and photosynthetic capacity, and the number, size, and ultrastructure of leaf cell chloroplast were significantly abnormal. The stripe-leaf trait of A36-S was controlled by a single recessive gene, which was mapped to a region from 7.66 Mb to 27.90 Mb of chromosome 4. Keywords:Setaria italica (L.) P. Beauv. ;stripe-leaf mutant;chloroplast;gene mapping
PDF (4565KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张硕, 智慧, 唐婵娟, 罗明昭, 汤沙, 贾冠清, 贾彦超, 刁现民. 谷子条纹叶突变体A36-S的细胞学特性分析及基因定位[J]. 中国农业科学, 2021, 54(14): 2952-2964 doi:10.3864/j.issn.0578-1752.2021.14.003 ZHANG Shuo, ZHI Hui, TANG ChanJuan, LUO MingZhao, TANG Sha, JIA GuanQing, JIA YanChao, DIAO XianMin. Cytological Characters Analysis and Low-Resolution Mapping of Stripe-Leaf MutantA36-S in Foxtail Millet [J]. Scientia Acricultura Sinica, 2021, 54(14): 2952-2964 doi:10.3864/j.issn.0578-1752.2021.14.003
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】谷子(Setaria italica (L.) P. Beauv.)属于禾本科(Granineae)黍亚科(Panicoideae)狗尾草属(Setaria),是中国北方传统粮食作物[1]。谷子为二倍体植物,基因组小(仅430 Mb左右)。近年来,随着谷子高质量基因组序列的获得[2,3,4],以及一些矮秆早熟品种高效遗传转化的突破[5],谷子作为功能基因组研究的模式作物而逐渐受到重视[6,7]。谷子同时也是典型的C4植物,随着环境问题及粮食安全问题的日趋严重,科学家提出将C4光合途径引入C3水稻及其他C3主要粮食和经济作物中,从而提高其产量和耐逆性的设想。谷子与传统的C4模式植物玉米和高粱相比,生育期短且基因组简单,加之其与水稻具有高度的基因组共线性,使其成为研究C4光合途径、构建水稻C4光合系统的新的模式植物。突变体是基因挖掘与功能研究的重要工具,目前,许多国内外机构都在广泛开展谷子及其野生近缘种狗尾草(Setaria viridis (L.) P. Beauv.)C4光合相关突变体的筛选及研究工作[8,9,10]。叶色突变体通常具有光合能力下降、光合器官结构异常等特点,是研究光合作用的重要材料。利用谷子叶色突变体开展光合生理学及细胞学观察,进而开展叶色调控基因的功能研究,对于进一步理解C4光合途径机理具有重要意义。【前人研究进展】植物叶色突变体具有多种类型,调控叶色的基因及机理也各不相同。目前,研究较为广泛且深入的叶色调控基因主要包括以下几类:(1)参与叶绿素合成及代谢的基因,比较典型的包括镁离子螯合酶D/I/H亚基的编码基因Chlorina-1、Chlorina-9和GUN4[11,12]、叶绿素a加氧酶基因OsCAO1[13]、叶绿素b还原酶基因OsNYC1[14]等,这些基因的变异往往导致叶片发生白化、黄化或持绿等均匀的叶色变化,有些还伴随叶绿体结构的异常;(2)参与叶绿体形成及发育的基因,这些基因中有些是直接编码某些叶绿体蛋白或结构组分的,例如叶绿体蛋白酶基因VYL1[15]、类囊体结合蛋白基因ZN[16]等,这些基因变异会导致叶绿体结构异常,有些则参与调控叶绿体基因的转录,从而影响叶绿体的形成及发育,这类基因的突变很多都会导致白化甚至致死等严重后果,例如叶绿体核糖体L13蛋白基因WLP1[17]、质体RNA聚合酶相关蛋白基因WLP2[18]、参与叶绿体RNA编辑及剪切的OsPPR6[19]等;(3)一些参与钾、铁等金属离子吸收及运输的基因,这些基因突变后,通常会影响叶绿体的正常发育,导致叶色变异,例如铁螯合物运输基因OsYSL15[20]、钾离子外排逆向转运蛋白基因AM1[21]等;(4)一些参与核酸合成及代谢的基因,在其功能丧失后,机体为保证核基因及线粒体基因的正常复制,会被迫牺牲叶绿体基因的复制与转录,从而导致叶绿体合成发育受到抑制,发生条纹或白化等叶色异常,例如尿嘧啶核苷酸激酶基因YGL8[22]、核糖核苷酸还原酶小亚基基因SDL/RNRS1[23]等。【本研究切入点】目前,已经在谷子上克隆了若干调控叶色的基因,包括镁离子螯合酶D亚基编码基因SiYGL1[24]、核糖核苷酸还原酶大亚基基因SiSTL1[25]、脱氧胞苷脱氨酶基因SiSTL2[26]、金属肽链内切酶SiYGL2[27]、类PsbP蛋白编码基因[28]等。然而,对于C4模式植物谷子而言,目前克隆的叶色调控基因尚少,C4植物叶绿体合成、发育及光合作用相关基因的研究基础仍然缺乏。【拟解决的关键问题】本研究中谷子A36-S突变体是来源于育种创制材料A36的一个自然突变体,叶片具有不规则白色条纹。通过对A36-S的光合参数、叶片细胞叶绿体合成发育情况进行观察分析,以探究叶色变异造成的光合生理和细胞学影响,并利用BSA图位克隆技术对条纹叶致变基因进行定位,以期为谷子光合作用相关基因的鉴定和功能分析提供一定的参考。
A:豫谷1号叶片横截切片(标尺50 μm);B: A36-S叶片横截切片(标尺50 μm);红色箭头指示正常叶肉细胞叶绿体;蓝色箭头指示正常维管束鞘细胞叶绿体;红色星号指示不含叶绿体的叶肉细胞;蓝色星号指示不含叶绿体的维管束鞘细胞 Fig. 3Leaf transverse sections of Yugu1 and A36-S
A: Leaf transverse section of Yugu1 (Bar = 50 μm); B: Leaf transverse section of A36-S (Bar=50 μm); Red arrows showed normal chloroplasts of mesophyll cells; Blue arrows showed normal chloroplasts of bundle sheath cells; Red stars showed the mesophyll cells containing no chloroplast; Blue stars show the bundle sheath cells containing no chloroplast
A:豫谷1号维管束鞘细胞叶绿体;B:豫谷1号叶肉细胞叶绿体;C:A36-S的Ⅰ类维管束鞘细胞;D:A36-S的Ⅰ类叶肉细胞;E:A36-S Ⅱ类维管束鞘细胞叶绿体;F: A36-S Ⅱ类叶肉细胞叶绿体;G: A36-S的Ⅲ类细胞;H:A36-SⅢ类细胞中的叶绿体,白色箭头指示泡状结构;SG:淀粉颗粒;SL:基质片层;GR:基粒;CLP:叶绿体 Fig. 5Ultrastructure of Yugu1 and A36-S chloroplast
A: Chloroplast of Yugu1 bundle sheath cell; B: Chloroplast of Yugu1 mesophyll cell; C: Type Ⅰ bundle sheath cells of A36-S; D: Type Ⅰ mesophyll cells of A36-S; E: Chloroplast in type Ⅱ bundle sheath cells of A36-S; F: Chloroplast in type Ⅱ mesophyll cells of A36-S; G: Type Ⅲ cell of A36-S; H: Chloroplast in type Ⅲ cell of A36-S, white arrows showed the foam-like structure; SG: Starch granules; SL: Stroma lamella; GR: Grana; CLP: Chloroplast
贾冠清, 刁现民. 谷子(Setaria italica (L.) P. Beauv.)作为功能基因组研究模式植物的发展现状及趋势 生命科学, 2017, 29(3):292-301. [本文引用: 1]
JIA GQ, DIAO XM. Current status and perspectives of researches on foxtail millet (Setaria italica (L.) P. Beauv.): A potential model of plant functional genomics studies Chinese Bulletin of Life Science, 2017, 29(3):292-301. (in Chinese) [本文引用: 1]
BENNETZEN JL, SCHMUTZJ, WANGH, PERCIFIELDR, HAWKINSJ, PONTAROLI AC, ESTEPM, FENGL, VAUGHN JN, GRIMWOODJ, JENKINSJ, BARRYK, LINDQUISTE, HELLSTENU, DESHPANDES, WANG XW, WU XM, MITROST, TRIPLETTJ, YANG XH, YE CY, MAURO-HERRERAM, WANGL, LI PH, SHARMAM, SHARMAR, RONALD PC, PANAUDO, KELLOGG EA, BRUTNELL TP, DOUST AN, TUSKAN GA, ROKHSARD, DEVOS KM. Reference genome sequence of the model plant Setaria Nature Biotechnology, 2012, 30:556-561. [本文引用: 1]
ZHANG GY, LIUX, QUAN ZW, CHENG SF, XUX, PAN SK, XIEM, ZENGP, YUEZ, WANG WL. Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential Nature Biotechnology, 2012, 30:549-554. DOI:10.1038/nbt.2195URL [本文引用: 1]
JIA GQ, HUANG XH, ZHIH, ZHAOY, ZHAOQ, LI WJ, CHAIY, YANG LF, LIU KY, LU HY, ZHU CR, LU YQ, ZHOU CC, FAN DL, WENG QJ, GUO YL, HUANGT, ZHANGL, LU TT, FENGQ, HAO HF, LIU HK, LUP, ZHANGN, LI YH, GUO EH, WANG SJ, WANG SY, LIU JR, ZHANG WF, CHEN GQ, ZHANG BJ, LIW, WANG YF, LI HQ, ZHAO BH, LI JY, DIAO XM, HANB. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica) Nature Genetics, 2013, 45:957-961. DOI:10.1038/ng.2673URL [本文引用: 1]
YANG ZY, ZHANG HS, LI XK, SHEN HM, GAO JH, HOU SY, ZHANGB, MAYESS, BENNETTM, MA JX, WU CY, SUIY, HAN YH, WANG XC. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 system Nature Plants, 2020, 6(9):1167-1178. DOI:10.1038/s41477-020-0747-7URL [本文引用: 1]
LI PH, BRUTNELL TP. Setaria viridis and Setaria italica, model genetic systems for the Panicoid grasses Journal of Experiment Botany, 2011, 62:3031-3037. DOI:10.1093/jxb/err096URL [本文引用: 1]
DIAO XM, SCHNABLEJ, BENNETZEN JL, LI JY. Initiation of Setaria as a model plant Frontiers of Agricultural Science and Engineering, 2014, 1:16-20. DOI:10.15302/J-FASE-2014011URL [本文引用: 1]
CAEMMERER SVON, QUICK WP, FURBANK RT. The development of C4 rice: Current progress and future challenges Science, 2012, 336:1671-1672. DOI:10.1126/science.1220177URL [本文引用: 1]
LUO MZ, ZHANGS, TANG CJ, JIA GQ, TANGS, ZHIH, DIAO XM. Screening of mutants related to the C4 photosynthetic Kranz structure in foxtail millet Frontiers in Plant Science, 2018, 9:1650. DOI:10.3389/fpls.2018.01650URL [本文引用: 1]
LUO MZ, TANG CJ, ZHANGS, ZHIH, TANGS, JIA GQ, DIAO XM. Screening of C4 photosynthesis-related mutants in foxtail millet (Setaria italica) by employmeny of low CO2 concentration incubator Journal of Plant Genetic Resources, 2018, 19(3):554-560. (in Chinese) [本文引用: 1]
ZHANG HT, LI JJ, YOO JH, YOO HC, CHO SH, KOH HJ, SEO HS, PAEK NC. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development Plant Molecular Biology, 2006, 62(3):325-337. DOI:10.1007/s11103-006-9024-zURL [本文引用: 1]
ZHOU SX, SAWICKIA, WILLOWS RD, LUO MZ. C-terminal residues of oryza sativa GUN4 are required for the activation of the ChlH subunit of magnesium chelatase in chlorophyll synthesis FEBS Letters, 2012, 586:205-210. DOI:10.1016/j.febslet.2011.12.026URL [本文引用: 1]
YANG YL, XU.J, HUANG LC, LENG YJ, DAI LP, RAO YC, CHENL, TU ZJ, HUJ, REN DY, ZHANG GH, ZHUL, GUO LB, QIANQ, ZENG DL. PGL, encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice Journal of Experimental Botany, 2016, 5:1297-1310. [本文引用: 1]
KUSABAM, ITOH, MORITAR, IIDAS, SATOY, FUJIMOTOM, KAWASAKIS, TANAKAR, HIROCHIKAH, NISHIMURAM, TANAKAA. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence The Plant Cell, 2017, 19:1362-1375. DOI:10.1105/tpc.106.042911URL [本文引用: 1]
DONGH, FEI GL, WU CY, WU FQ, SUN YY, CHEN MJ, REN YL, ZHOU KN, CHENG ZY, WANG JL, JIANGL, ZHANGX, GUO XP, LEI CL, SUN, WANG HY, WAN JM. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants Plant Physiology, 2013, 162:1867-1880. DOI:10.1104/pp.113.217604URL [本文引用: 1]
LI JJ, PANDEYAD, NATHK, ZULFUGAROV IS, YOO SC, ZHANG HT, YOO JH, CHO SH, KOH HJ, KIM DS, SEO HS, KANG BC, LEE ZH, PAEK NC. ZEBRA-NECROSIS, a thylakoid-bound protein, is critical for the photoprotection of developing chloroplasts during early leaf development The Plant Journal, 2010, 62:713-725. DOI:10.1111/tpj.2010.62.issue-4URL [本文引用: 1]
SONGJ, WEI XJ, SHAO GN, SHENG ZH, CHEN DB, LIU CL, JIAO GA, XIE LH, TANG SQ, HU PS. The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions Plant Molecular Biology, 2014, 84:301-314. DOI:10.1007/s11103-013-0134-0URL [本文引用: 1]
LV YS, SHAO GN, QIU JH, JIAO GA, SHENG ZH, XIE LH, WU YW, TANG SQ, WEI XJ, HU PS. White Leaf and Panicle 2, encoding a PEP-associated protein, is required for chloroplast biogenesis under heat stress in rice Journal of Experiment Botany, 2017, 68:5147-5160. DOI:10.1093/jxb/erx332URL [本文引用: 1]
TANG JP, ZHANG WW, WENK, CHEN GM, SUNJ, TIAN YL, TANG WJ, YUJ, AN HZ, WU TT, KONGF, TERZAGHIW, WANG CM, WAN JM. OsPPR6, a pentatricopeptide repeat protein involved in editing and splicing chloroplast RNA, is required for chloroplast biogenesis in rice Plant Molecular Biology, 2017, 95:345-357. DOI:10.1007/s11103-017-0654-0URL [本文引用: 1]
LEES, CHIECKO CJ, KIM SA, WALKER EL, LEEY, GUERINOT ML, ANN GG. Disruption of OsYSL15 leads to iron inefficiency in rice plants Plant Physiology, 2009, 150:786-800. DOI:10.1104/pp.109.135418URL [本文引用: 1]
SHENG PK, TAN JJ, JIN MN, WU FQ, ZHOU KN, MA WW, HENG YQ, WANG JL, GUO XP, ZHANGX, CHENG ZJ, LIU LL, WANG CM, LIU XM, WAN JM. Albino midrib 1, encoding a putative potassium efflux antiporter, affects chloroplast development and drought tolerance in rice Plant Cell Reports, 2014, 33:1581-1594. DOI:10.1007/s00299-014-1639-yURL [本文引用: 1]
ZHU XY, GUOS, WANG ZW, XING YD, ZHANG TQ, SHEN WQ, SANG XC, LING YH, HE GH. Map-based cloning and functional analysis of YGL8, which controls leaf colour in rice (Oryza sativa) BMC Plant Biology, 2016, 16:134-148. DOI:10.1186/s12870-016-0821-5URL [本文引用: 1]
QINR, ZENG DD, LIANGR, YANG CC, AKHTERD, ALAMINM, JIN XL, SHI CH. Rice gene SDL/RNRS1, encoding the small subunit of ribonucleotide reductase, is required for chlorophyll synthesis and plant growth development Gene, 2017, 627:351-362. DOI:10.1016/j.gene.2017.05.059URL [本文引用: 1]
LIW, TANGS, ZHANGS, SHAN JG, TANG CJ, CHEN QN, JIA GQ, HAN YH, ZHIH, DIAO XM. Gene mapping and functional analysis of the novel leaf color gene SiYGL1 in foxtail millet [Setaria italica (L.) P. Beauv.] Physiologia Plantarum, 2015, 157:24-37. DOI:10.1111/ppl.2016.157.issue-1URL [本文引用: 2]
TANG CJ, TANGS, ZHANGS, LUO MZ, JIA GQ, ZHIH, DIAO XM. SiSTL1, encoding a large subunit of ribonucleotide reductase, is crucial for plant growth, chloroplast biogenesis, and cell cycle progression in Setaria italica Journal of Experimental Botany, 2019, 70(4):1167-1182. DOI:10.1093/jxb/ery429URL [本文引用: 6]
ZHANGS, TANGS, TANG CJ, LUO MZ, JIA GQ, ZHIH, DIAO XM. SiSTL2 is required for cell cycle leaf organ development, chloroplast biogenesis and influences C4 photosynthesis in Setaria italica (L.) P. Beauv Frontiers in Plant Science, 2018, 9:1308. DOI:10.3389/fpls.2018.01308URL [本文引用: 3]
ZHANGS, ZHIH, LIW, SHAN JG, TANG CJ, JIA GQ, TANGS, DIAO XM. SiYGL2 is involved in the regulation of leaf senescence and photosystem II efficiency in Setaria italica (L.) P. Beauv Frontiers in Plant Science, 2018, 9:1103. DOI:10.3389/fpls.2018.01103URL [本文引用: 3]
WANG QL, WANG ZL, HANF, DU XF, LIAN SC, HAN KN, ZHOUX, LI HJ, ZHANG LY, WANGJ, GUO EH. Identified and candidate gene analysis of a white stripe leaf mutant wsl2 in foxtail millet Acta Agriculturae Boreali-Sinica, 2020, 35(1):214-221. (in Chinese) [本文引用: 1]
LICHTENTHALER HK. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes Methods in Enzymology, 1987, 148:350-382. [本文引用: 1]
XIANG JS, TANGS, ZHIH, JIA GQ, WANG HJ, DIAO XM. Loose Panicle1 encoding a novel WRKY transcription factor, regulates panicle development, stem elongation, and seed size in foxtail millet [Setaria italica (L.) P. Beauv.] PLoS ONE, 2017, 12(6):e0178730. DOI:10.1371/journal.pone.0178730URL [本文引用: 1]
ZHANGS, TANG CJ, ZHAOQ, LIJ, YANG LF, QIE LF, FAN XK, LIL, ZHANGN, ZHAO MC, LIU XT, CHAIY, ZHANGX, WANG HL, LI YT, LIW, ZHIH, JIA GQ, DIAO XM. Development of highly polymorphic simple sequence repeat markers using genome- wide microsatellite variant analysis in foxtail millet [Setaria italica (L.) P. Beauv.] BMC Genomics, 2014, 15:78. DOI:10.1186/1471-2164-15-78URL [本文引用: 4]
ZHOU HL, HUANG SM. Effects of sink source relationship on chlorophyll content and photosynthetic characteristics of rice Journal of Green Science and Technology, 2017, 24:147-149. (in Chinese) [本文引用: 1]
ZHAN JC, WANG LJ, HUANG WD. Effects of low light environment on the growth and photosynthetic characteristics of grape leaves Journal of China Agricultural University, 2002, 7(3):75-78. (in Chinese) [本文引用: 1]
ZHANG MS, TANF. Relationship between ratio of chlorophyll a and b under water stress and drought resistance of different sweet potato varieties Seed, 2001, 4:23-25. (in Chinese) [本文引用: 1]
WANG JH, XUT. Effects of simulated acid rain on content of chlorophyll a, b and their ratio in cotton cotyledon Journal of Hubei Agricultural College, 1991, 11(2):1-10. (in Chinese) [本文引用: 1]
GE CL, WANGL, YE WJ, WU LW, CUI YT, CHENP, PAN JJ, ZHANGD, HUJ, ZENG DL, DONG GJ, QIANQ, GUO LB, XUE DW. Single-point mutation of an histidine-aspartic domain containing gene involving in chloroplast ribosome biogenesis leads to white fine stripe leaf in rice Scientific Reports, 2017, 7:2398. DOI:10.1038/s41598-017-02724-xURL [本文引用: 1]
WANGY, REN YL, ZHOU KN, LIU LL, WANG JL, XUY, ZHANGH, ZHANGL, FENG ZM, WANG LW, MA WW, WANG YL, GUO XP, ZHANGX, LEI CL, WAN JM. WHITE STRIPE LEAF4 encodes a novel P-type PPR protein required for chloroplast biogenesis during early leaf development Frontiers in Plant Science, 2017, 8:1116. DOI:10.3389/fpls.2017.01116URL [本文引用: 1]
NIUM, WANG YH, WANG CM, LYUJ, WANG YL, DONGH, LONG WH, WANGD, KONG WY, WANG LW, GUO XP, SUN LT, HU TT, ZHAI HQ, WANG HY, WAN JM. ALR encoding dCMP deaminase is critical for DNA damage repair, cell cycle progression and plant development in rice Journal of Experimental Botany, 2017, 68:5773-5786. DOI:10.1093/jxb/erx380URL [本文引用: 1]
LIU LL, YOUJ, ZHUZ, CHEN KY, ·HU MM, GUH, LIU ZW, WANG ZY, WANG YH, LIU SJ, CHEN LM, LIUX, TIAN YL, ZHOU SR, JIANGL, WAN JM. WHITE STRIPE LEAF8, encoding a deoxyribonucleoside kinase, is involved in chloroplast development in rice Plant Cell Reports, 2020, 39:19-33. DOI:10.1007/s00299-019-02470-6URL [本文引用: 1]
YE WJ, HU SK, WU LW, GE CW, CUI YT, CHENP, WANGX, XUJ, REN DY, DONG GJ, QIANQ, GUO LB. White stripe leaf 12 (WSL12), encoding a nucleoside diphosphate kinase 2 (OsNDPK2), regulates chloroplast development and abiotic stress response in rice (Oryza sativa L.) Molecular Breeding, 2016, 36:57. DOI:10.1007/s11032-016-0479-6URL [本文引用: 1]
WANGP, KHOSHRAVESHR, KARKIS, TAPIAR, BALAHADIA CP, BANDYOPADHYAYA, QUICK WP, FURBANKR, SAGE TL, LANGDALE JA. Re-creation of a key step in the evolutionary switch from C3 to C4 leaf anatomy Current Biology, 2017, 27(21):3278-3287. DOI:10.1016/j.cub.2017.09.040URL [本文引用: 1]
SLEWINSKI TL, ANDERSON AA, ZHANG CK. Scarecrow plays a role in establishing Kranz anatomy in maize leaves Plant Cell Physiology, 2012, 53(12):2030-2037. DOI:10.1093/pcp/pcs147URL [本文引用: 1]
 ,1,2, 智慧1, 唐婵娟1, 罗明昭1, 汤沙1, 贾冠清1, 贾彦超1, 刁现民
,1,2, 智慧1, 唐婵娟1, 罗明昭1, 汤沙1, 贾冠清1, 贾彦超1, 刁现民

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT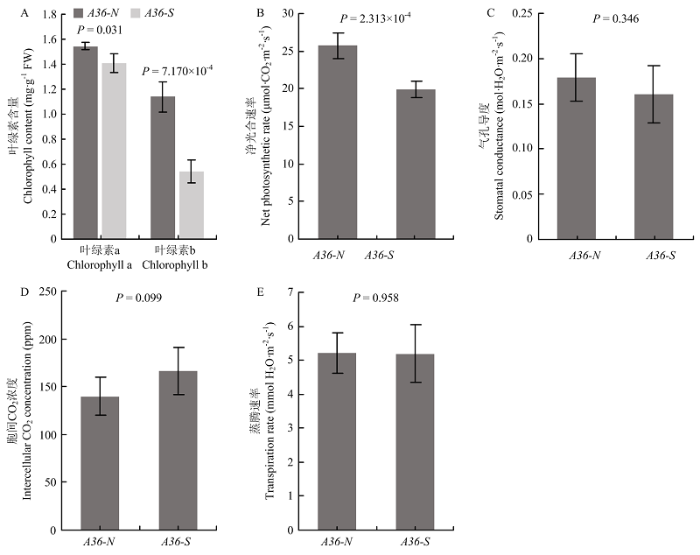 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT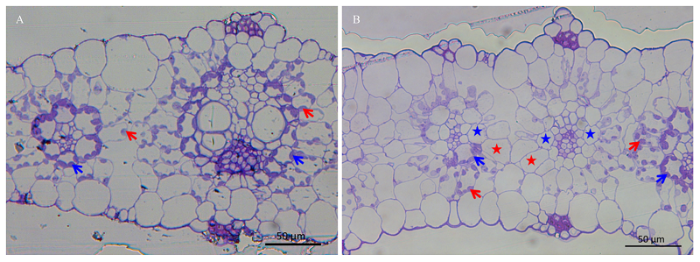 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT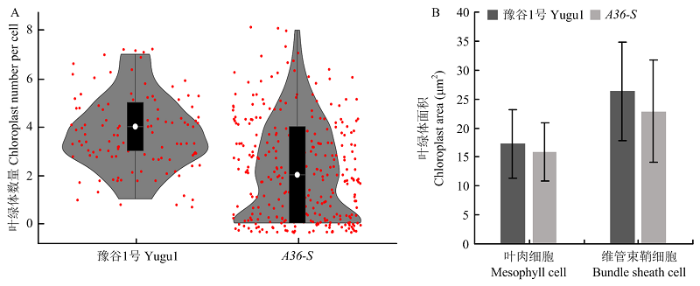 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT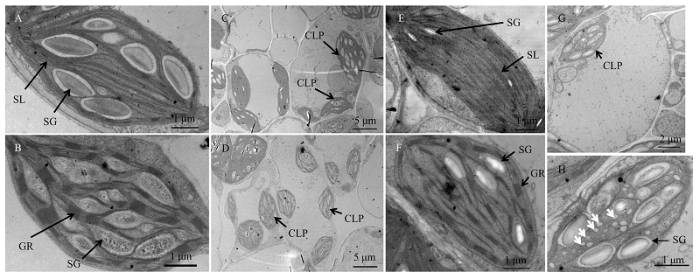 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}