Knockout of Single Allele of fl(2)d Significantly Decreases the Fecundity and Fertility inPlutella xylostella
LI FeiFei,, WANG BeiBei, LAI YingFang, YANG FeiYing, YOU MinSheng,, HE WeiYi,State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops/Institute of Applied Ecology, Fujian Agriculture and Forestry/International Joint Research Laboratory of Ecological Pest Control, Ministry of Education, Fujian Agriculture and Forestry University/Key Laboratory of Integrated Pest Management for Fujian-Taiwan Crops, Ministry of Agriculture and Rural Affairs, Fujian Agriculture and Forestry/Ministerial and Provincial Joint Innovation Centre for Safety Production of Cross-Strait Crops, Fuzhou 350002
Abstract 【Objective】 RNA methylation is the main form of epigenetic modification at post-transcriptional level, which is involved in many important cellular processes. The diamondback moth, Plutella xylostella, is an important oligophagous insect pest, causing serious loss on the production of cruciferous vegetables. However, the function of RNA methylation-related genes in P. xylostella is still unclear. The present study aims to identify and clone the homologous fl(2)d, one of the members of the RNA methylation protein complex (writers), to determine the expression pattern offl(2)d, and to knockout fl(2)d using CRISPR/Cas9 for the investigation of its biological functions in P. xylostella.【Method】The sequence of homologous fl(2)d was identified in the genome database ofP. xylostella, which was used for PCR amplification of the coding sequence (CDS). Quantitative real-time PCR (qRT-PCR) was used to study the relative expression levels of fl(2)d in different developmental stages and adult gonads of P. xylostella. The fl(2)d was edited using CRISPR/Cas9 combined with egg injection. Each of the adults that developed from the injected eggs was used to pair with a wild-type adult for reproduction. Offspring of the same population was forced to inbreed by single-pair mating to establish the mutant strains. The differences of genetic characters, biological parameters and phenotypes between mutants and wild-type individuals were recorded and compared to decipher the function offl(2)d.【Result】The CDS of fl(2)d with length of 912 bp was isolated, the expression of which was high in female pupa, adult and egg, moderate in male adult and pupa, the lowest in larva, and significantly higher in ovary than in testis of adult. The sgRNAs targeting fl(2)d and the Cas9 protein were mixed to inject eggs, and the offsprings carrying mutant alleles were screened for homozygous strains based on single-pair inbreeding for 10 generations. Three types of heterozygous mutant strains both predicted to cause frameshift of the CDS were obtained, with the deletion of 4 (Δfl(2)d213-4), 5 (Δfl(2)d213-5) and 7 (Δfl(2)d213-7) bases. During the screening process, six and two homozygous mutants from Δ fl(2)d213-4 and Δ fl(2)d213-5 strains were identified, respectively. The homozygous mutants of Δ fl(2)d213-4 successfully mated in two pairs, but no eggs were produced. Meanwhile, each two male adults of homozygous mutants of either Δ fl(2)d213-4 or Δ fl(2)d213-5 were mated with the same type of female heterozygous mutant, and also no eggs were produced. The results indicated that individuals with homozygousfl(2)d mutation may have extremely low survival rate and not be able to produce offspring. Through analyzing separation ratio of the genotypes of offspring from the inbreeding of heterozygous mutants and the hybridization between heterozygous mutants and wild-type, it was found that the ratio of heterozygous mutant individuals to wild-type was slightly less than 2 and 1, respectively, indicating that heterozygous mutation of fl(2)d would affect the normal growth and development of P. xylostella, and in some cases would lead to death. The offsprings of mutant individuals, which carry a mutant allele, showed a sex ratio close to 1﹕1 (P<0.05). It was speculated that thefl(2)d might not be involved in sex determination in P. xylostella. For the mating consists of mutant adults, the fecundity and hatchability were significantly lower (P<0.01) than the mating between wild-type adults. Most of the eggs produced from the mutant parents look abnormal, and could not hatch normally due to water loss and shrinkage. Based on the dissection of adult gonads, it was found that the number of attached eggs on the ovary of the mutant female adult and the wild-type female adult that has mated with mutant male adult was less than that of the wild-type virgin female adult, while no obvious abnormality was found for the testis of mutant male adult. Some of the hatched heterozygous mutants showed different degrees of distortion during the whole developmental process, resulting in the failure to complete life cycle. A small part of the heterozygous mutant individuals could develop normally, and thus transmit the mutant allele to their offspring. According to our findings, a model of genetic control ofP. xylostella based on fl(2)d was proposed.【Conclusion】The fl(2)d is involved in the reproductive process and embryonic development ofP. xylostella, mutation of which significantly affects the population size of the offspring, making it an ideal target for the genetic control of P. xylostella. Keywords:Plutella xylostella;fl(2)d;heterozygous mutant;fecundity;fertility
PDF (2751KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 李飞飞, 王贝贝, 赖颖芳, 杨菲颖, 尤民生, 何玮毅. fl(2)d单等位基因的敲除显著降低小菜蛾的生殖力和育性[J]. 中国农业科学, 2021, 54(14): 3029-3042 doi:10.3864/j.issn.0578-1752.2021.14.009 LI FeiFei, WANG BeiBei, LAI YingFang, YANG FeiYing, YOU MinSheng, HE WeiYi. Knockout of Single Allele of fl(2)d Significantly Decreases the Fecundity and Fertility inPlutella xylostella[J]. Scientia Acricultura Sinica, 2021, 54(14): 3029-3042 doi:10.3864/j.issn.0578-1752.2021.14.009
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】小菜蛾(Plutella xylostella)属鳞翅目(Lepidoptera)菜蛾科(Plutellidae),是危害十字花科蔬菜的重要害虫,全球每年用于其防治的费用及其造成的损失高达40—50亿美元[1]。在我国南方菜区,小菜蛾可终年发生危害,世代重叠严重、抗药性强,促使研究人员不断探索控制其种群增长的方法[2,3]。随着有害生物综合治理(integrated pest management,IPM)理念的提出,人们不再拘泥于传统的防治策略,而是致力于开发绿色、环保、有利于生态平衡的可持续治理方法[4,5]。当前,害虫灾变机理的研究已迈入组学时代,基于害虫遗传控制的原理和方法,研发小菜蛾绿色防控技术显示出了巨大的潜力和优势[6,7,8]。筛选合适的靶标基因,研究配套的遗传操作系统,将进一步提升害虫遗传控制的效率和稳定性[9,10]。RNA甲基化是基因转录后水平表观修饰的主要形式,rRNA、mRNA、tRNA甚至是非编码RNA都会出现超过100多种的修饰类型,其中以m6A(N6-腺苷酸甲基化)最为常见[11,12]。m6A甲基化存在于绝大多数生物体内,包括哺乳动物、昆虫、细菌和病毒。m6A影响生物体基因表达、mRNA稳定性、RNA可变剪接、蛋白翻译效率和X染色体失活等重要的细胞学过程[13,14]。参与m6A甲基化过程蛋白的失活会影响生物体正常的生命活动,探明小菜蛾中m6A甲基化蛋白成员female- lethal-2-d(fl(2)d)的生物学功能,对今后以该基因作为靶标建立小菜蛾遗传防控系统具有重要意义。【前人研究进展】近年来,人们将RNA甲基化修饰抗体富集技术和高通量测序技术相结合,实现了对m6A甲基化在转录组水平的分布和丰度检测[15,16]。在人类和小鼠的基因组中,m6A甲基化位点高度保守,大多集中在终止密码子附近和长的内部外显子,说明这种修饰在基因表达的表观遗传调控中发挥了重要作用[17]。参与m6A代谢和信号途径的蛋白有三大类,分别是RNA甲基化蛋白(writers)、去甲基化蛋白(erasers)和甲基化阅读蛋白(readers),它们均以复合物的形式发挥功能[18]。一旦哺乳动物体内的这些蛋白出现异常,就会引发一系列的疾病,包括肿瘤、神经性疾病和发育迟缓等[15,19-20]。Writers主要包括methyltransferase-like 3(METTL3)、methyltransferase-like 14(METTL14)、fl(2)d、vir-Like m6A methyltransferase associated(KIAA1429/VIRMA)和RNA-binding motif protein 15(RBM15)[18,21-22],在昆虫中的报道较少。家蚕(Bombyx mori)中发现,METTL3和METTL14在滞育品系中的表达量高于非滞育品系,说明m6A甲基化介导的转录后表观遗传调控可能参与了家蚕滞育的发生[23];在细胞水平利用RNA干扰(RNAi)技术沉默METTL3和METTL14,会导致家蚕细胞周期进程停止、染色体联会和分离失败[24]。METTL3、METTL14和fl(2)d的表达水平以及m6A含量在工蜂和蜂王幼虫阶段不同时期发生显著变化,说明m6A甲基化影响了蜜蜂的幼虫发育和级型分化[25]。fl(2)d的研究常见于果蝇[26,27,28],该基因主要有3个靶基因:sex-lethal(Sxl)、transformer和ultrabithorax,前两个基因是性别决定通路的上游基因,fl(2)d主要通过影响这两个基因前体mRNA的可变剪接影响果蝇的性别决定、剂量补偿、卵子形成与分化等[29,30,31,32]。fl(2)d也可以调控第3个靶基因的可变剪接,但对雌、雄两性的影响并无差异[33]。此外,fl(2)d在果蝇胚胎和卵巢中高表达,在卵子形成的早期,fl(2)d蛋白在不同的细胞核间移动,说明它是一种影响卵子发生的关键核蛋白[34]。【本研究切入点】昆虫RNA甲基化的研究目前仅见于家蚕、蜜蜂和果蝇,在小菜蛾上还未见报道。fl(2)d通过参与RNA甲基化过程影响了众多生物学过程,是开展害虫遗传控制潜在的靶标。由于鳞翅目昆虫中Sxl同源基因不具有决定昆虫性别的功能[35,36],利用基因编辑技术clustered regular interspaced short palindromic repeat sequences/CRISPR-associated protein 9(CRISPR/Cas9)探明小菜蛾fl(2)d同源基因是否参与性别决定、生殖及其他重要生长发育过程,可为今后利用该基因进行害虫的遗传防控打下理论基础。【拟解决的关键问题】小菜蛾fl(2)d的序列及其表达模式;建立小菜蛾fl(2)d突变品系;突变品系遗传特性、生物学参数和表型的变化;fl(2)d在小菜蛾生长发育中的功能。
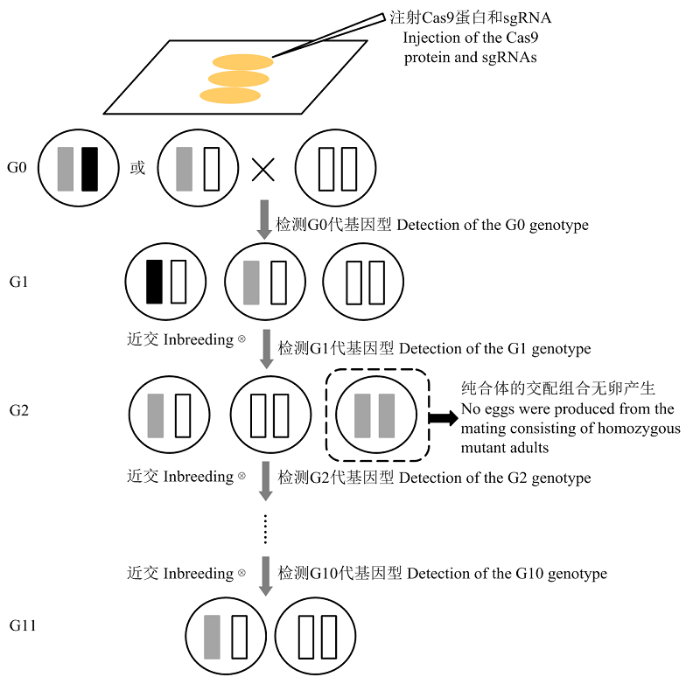
圆圈表示细胞核,圈中长方形表示编辑的目标染色体,空白填充表示野生型,不同颜色的填充表示不同类型的突变 Fig. 1Flow chart for screening mutant strains of fl(2)d in P. xylostella
The circle denotes nucleus. The rectangle inside circle denotes the chromosome targeted for gene editing. The rectangle filled with blank indicates the wild-type, and the rectangles filled with different colors indicate different types of mutations
A:fl(2)d CDS序列的克隆CDS cloning of fl(2)d;M:DNA分子标准DNA marker。B:预测的fl(2)d蛋白质序列Predicted protein sequence of fl(2)d;红色字母:保守功能域Red letters indicate the conserved domain。C:fl(2)d在不同发育阶段的表达Expression of fl(2)d in different developmental stages;E:卵Egg;FL:雌幼虫Female larva;ML:雄幼虫Male larva;FP:雌蛹Female pupa;MP:雄蛹Male pupa;FA:雌成虫Female adult;MA:雄成虫Male adult;采用单因素方差分析法进行差异显著性检验,使用Tukey法进行多重比较,不同字母表示差异显著(P<0.05)Significant difference analysis was performed using one-way ANOVA followed by a Tukey’s HSD post hoc test. Different letters indicate significant difference atP<0.05 level。D:fl(2)d在成虫生殖腺的表达Expression offl(2)d in adult gonads;O:卵巢Ovary;T:精巢Testis;采用t检验进行差异显著性分析,**表示检验性水平P<0.01 Significant difference analysis was performed usingt test. Double asterisks indicate significant difference of P<0.01 Fig. 2Cloning and expression profiling of fl(2)d in P. xylostella
A:fl(2)d基因结构及sgRNA靶点示意图Schematic diagram of the fl(2)d gene structure and sgRNA target site。B:G0代野生型(上)和突变体(下)成虫fl(2)d测序峰图;红色框是sgRNA,蓝色框是PAM结构Sequencing chromatograms of fl(2)d of wild-type (top) and mutant (bottom) adults; The edited site is indicated by a red rectangle and the protospacer adjacent motif (PAM) sequence is showed by a blue rectangle。C:突变序列,虚线表示缺失的序列Mutant sequences, the dashed lines represent the deleted bases caused by CRISPR/Cas9 Fig. 3Gene editing of fl(2)din P. xylostella
Table 2 表2 表2G2—G10代fl(2)d基因型分离情况 Table 2The segregation of fl(2)dgenotypes from G2 to G10 generation
G2代至G10代不同基因型个体数 Numbers of different genotypes from G2 to G10
AA×Aa(亲本基因型Parental genotype)
Aa×Aa(亲本基因型Parental genotype)
Δfl(2)d-4
Δfl(2)d-5
Δfl(2)d-7
P=0.05, df=1, χ2=3.841
Δfl(2)d-4
Δfl(2)d-5
Δfl(2)d-7
P=0.05, df=1, χ2=3.841
AA
Aa
AA
Aa
AA
Aa
Aa/AA
χ2
AA
Aa
aa
AA
Aa
aa
AA
Aa
aa
Aa/AA
χ2
G2
13
13
11
11
22
20
0.957
0.044
6
16
0
/
/
/
3
9
0
2.778
0.721
G3
19
19
/
/
40
30
0.831
0.926
15
24
3
7
5
2
10
16
0
1.406
2.506
G4
/
/
/
/
14
14
1.000
0
7
16
3
14
30
0
11
19
0
2.031
0.095
G5
20
14
25
37
1
3
1.174
0.640
/
/
/
11
17
0
13
23
0
1.667
0.500
G6
5
3
23
21
20
8
0.667
3.200
/
/
/
15
25
0
2
6
0
1.824
0.094
G7
17
7
18
12
19
15
0.630
4.545
/
/
/
2
10
0
/
/
/
5.000
2.063
G8
6
6
6
4
19
15
0.806
0.643
/
/
/
3
7
0
1
3
0
2.500
0.339
G9
/
/
7
9
11
9
1.000
0
/
/
/
/
/
/
/
/
/
/
/
G10
/
/
/
/
18
26
1.444
1.455
/
/
/
3
5
0
/
/
/
1.667
0.031
合计Total
80
62
90
94
164
140
0.886
0.292
28
56
6
55
99
2
40
76
0
1.878
0.488
斜杠表示未检测。亲本只统计基因型,不考虑性别The slash denotes the missing data. Only genotypes of the parents were recorded regardless of the sexuality。A:野生型基因型Wild-type genotype;a:突变型基因型Mutant genotype
A:突变品系的产卵量The fecundity of mutant strains。B:突变品系的孵化率The hatchability of mutant strains。WT:野生型成虫杂交Hybridization between wild-type adults;F-4:杂合雌性Δfl(2)d213-4成虫与野生型杂交Hybridization of heterozygous female adult of Δ fl(2)d213-4 with wild-type;M-4:杂合雄性Δfl(2)d213-4成虫与野生型杂交Hybridization of heterozygous male adult of Δ fl(2)d213-4 with wild-type;-4:杂合Δfl(2)d213-4成虫近交Inbreeding between heterozygous adults of Δ fl(2)d213-4;F-5:杂合雌性Δfl(2)d213-5成虫与野生型杂交Hybridization of heterozygous female adult of Δ fl(2)d213-5 with wild-type;M-5:杂合雄性Δfl(2)d213-5成虫与野生型杂交Hybridization of heterozygous male adult of Δ fl(2)d213-5 with wild-type;-5:杂合Δfl(2)d213-5成虫近交Inbreeding between heterozygous adults of Δ fl(2)d213-5;F-7:杂合雌性Δfl(2)d214-7成虫与野生型杂交Hybridization of heterozygous female adult of Δ fl(2)d214-7 with wild-type;M-7:杂合雄性Δfl(2)d214-7成虫与野生型杂交Hybridization of heterozygous male adult of Δ fl(2)d214-7 with wild-type;-7:杂合Δfl(2)d214-7成虫近交Inbreeding between heterozygous adults of Δ fl(2)d214-7。C:性别比例The sex ratios of mutant offspring;采用t检验进行差异显著性分析,**表示显著性水平P<0.01 Significant difference analysis was performed usingt test. Double asterisks indicate significant difference of P<0.01 Fig. 4Effects of fl(2)dmutation on the fecundity, hatchability and sex ratio of P. xylostella
A:野生型雌成虫产卵情况The fecundity of wild-type female adult。B:突变体雌成虫产卵情况The fecundity of mutant female adult。C、D:失水皱缩的卵The shrinking and dehydrated eggs。E:未能正常孵化的卵Eggs that do not hatch successfully。F:未能正常出卵的幼虫The larvae that fail to come out of eggs。G:未交配野生型雌成虫卵巢Ovary of the wild-type virgin female adult。H:与突变体雄成虫交配后野生型雌成虫卵巢Ovary of the wild-type female adult mated with the mutant male adult。除A图外,所观察的卵均为突变体雌成虫与野生型雄成虫单对交配后所产的后代,突变体类型为Δfl(2)d213-5 Except for figure A, the presented phenotypes of the eggs are all produced from the single-pair mating of mutant female adult and wild-type male adult, and the type of mutation is Δ fl(2)d213-5 Fig. 5Comparison of reproductive phenotypes of fl(2)dmutant and wild-type female adults of P. xylostella
FURLONG MJ, WRIGHT DJ, DOSDALL LM. Diamondback moth ecology and management: Problems, progress, and prospects Annual Reviews of Entomology, 2013, 58:517-541. DOI:10.1146/annurev-ento-120811-153605URL [本文引用: 1]
LI ZY, FENGX, LIU SS, YOU MS, FURLONG MJ. Biology, ecology, and management of the diamondback moth in China Annual Review of Entomology, 2016, 61:277-296. DOI:10.1146/annurev-ento-010715-023622URL [本文引用: 1]
GURR GM, REYNOLDS OL, JOHNSON AC, DESNEUXN, ZALUCKI MP, FURLONG MJ, LI ZY, AKUTSE KS, CHEN JH, GAO XW, YOU MS. Landscape ecology and expanding range of biocontrol agent taxa enhance prospects for diamondback moth management. A review Agronomy for Sustainable Development, 2018, 38:23. DOI:10.1007/s13593-018-0500-zURL [本文引用: 1]
XU XJ, HE WY, YANGJ, CHENW, YOU MS. Research and applications of genetics-based methods for pest control Scientia Sinica Vitae, 2019, 49(8):938-950. (in Chinese) DOI:10.1360/N052018-00256URL [本文引用: 1]
CHENW, YANG FY, XU XJ, KUMARU, HE WY, YOU MS. Genetic control of Plutella xylostella in omics era Archives of Insect Biochemistry and Physiology, 2019, 102(3):e21621. [本文引用: 1]
SHELTON AM, LONG SJ, WALKER AS, BOLTONM, COLLINS HL, REVUELTAL, JOHNSON LM, MORRISON NI. First field release of a genetically engineered, self-limiting agricultural pest insect: Evaluating its potential for future crop protection Frontiers in Bioengineering and Biotechnology, 2020, 7:482. DOI:10.3389/fbioe.2019.00482URL [本文引用: 1]
YOU MS, YUEZ, HE WY, YANG XH, YANGG, XIEM, ZHAN DL, BAXTER SW, VASSEURL, GURR GM, et al. A heterozygous moth genome provides insights into herbivory and detoxification Nature Genetics, 2013, 45(2):220-225. DOI:10.1038/ng.2524URL [本文引用: 2]
JINL, WALKER AS, FU GL, HARVEY-SAMUELT, DAFA’ALLAT, MILESA, MARUBBIT, GRANVILLED, HUMPHREY-JONESN, O’CONNELLS, MORRISON NI, ALPHEYL. Engineered female-specific lethality for control of pest Lepidoptera ACS Synthetic Biology, 2013, 2(3):160-166. DOI:10.1021/sb300123mURL [本文引用: 1]
HARVEY-SAMUELT, MORRISON NI, WALKER AS, MARUBBIT, YAOJ, COLLINS HL, GORMANK, DAVIES TG, ALPHEYN, WARNERS, SHELTON AM, ALPHEYL. Pest control and resistance management through release of insects carrying a male-selecting transgene BMC Biology, 2015, 13:49. DOI:10.1186/s12915-015-0161-1URL [本文引用: 1]
PANT. N6-methyl-adenosine modification in messenger and long non-coding RNA Trends in Biochemical Sciences, 2013, 38(4):204-209. DOI:10.1016/j.tibs.2012.12.006URL [本文引用: 1]
DOMINISSINID, MOSHITCH-MOSHKOVITZS, SCHWARTZS, SALMON-DIVONM, UNGARL, OSENBERGS, CESARKASK, JACOB-HIRSCHJ, AMARIGLION, KUPIECM, SOREKR, RECHAVIG. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq Nature, 2012, 485(7397):201-206. DOI:10.1038/nature11112URL [本文引用: 2]
MEYER KD, SALETOREY, ZUMBOP, ELEMENTOO, MASON CE, JAFFREY SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons Cell, 2012, 149(7):1635-1646. DOI:10.1016/j.cell.2012.05.003URL [本文引用: 1]
DOMINISSINID, MOSHITCH-MOSHKOVITZS, SALMON- DIVONM, AMARIGLION, RECHAVIG. Transcriptome-wide mapping of N6-methyladenosine by m6A-seq based on immunocapturing and massively parallel sequencing Nature Protocols, 2013, 8(1):176-189. DOI:10.1038/nprot.2012.148URL [本文引用: 1]
WU BX, LIL, HUANGY, MAJ, MIN JB, MIN JR. Readers, writers and erasers of N6-methylated adenosine modification Current Opinion in Structural Biology, 2017, 47:67-76. DOI:10.1016/j.sbi.2017.05.011URL [本文引用: 2]
ZHOU ZL, LICKLIDER LJ, GYGI SP, REEDR. Comprehensive proteomic analysis of the human spliceosome Nature, 2002, 419(6903):182-185. DOI:10.1038/nature01031URL [本文引用: 1]
HORIUCHIK, KAWAMURAT, IWANARIH, OHASHIR, NAITOM, KODAMAT, HAMAKUBOT. Identification of Wilms’ tumor 1-associating protein complex and its role in alternative splicing and the cell cycle Journal of Biological Chemistry, 2013, 288(46):33292-33302. DOI:10.1074/jbc.M113.500397URL [本文引用: 1]
LIU JZ, YUE YN, HAN DL, WANGX, FUY, ZHANGL, JIA GF, YUM, LU ZK, DENGX, DAIQ, CHEN WZ, HEC. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation Nature Chemical Biology, 2014, 10(2):93-95. DOI:10.1038/nchembio.1432URL [本文引用: 1]
BANSALH, YIHUAQ, IYER SP, GANAPATHYS, PROIAD, PENALVA LO, UREN PJ, SURESHU, CAREW JS, KARNAD AB, WEITMANS, TOMLINSON GE, RAO MK, KORNBLAU SM, BANSALS. WTAP is a novel oncogenic protein in acute myeloid leukemia Leukemia, 2014, 28(5):1171-1174. DOI:10.1038/leu.2014.16URL [本文引用: 1]
JIANGT, LI JS, QIANP, XUEP, XUJ, CHEN YR, ZHUJ, TANG SM, ZHAO QL, QIAN HY, SHEN XJ. The role of N6- methyladenosine modification on diapause in silkworm (Bombyx mori) strains that exhibit different voltinism Molecular Reproduction and Development, 2019, 86(12):1981-1992. DOI:10.1002/mrd.v86.12URL [本文引用: 2]
LI BQ, WANG XY, LI ZQ, LU CC, ZHANGQ, CHANGL, LI WS, CHENG TC, XIA QY, ZHAOP. Transcriptome-wide analysis of N6-methyladenosine uncovers its regulatory role in gene expression in the LepidopteranBombyx mori Insect Molecular Biology, 2019, 28(5):703-715. DOI:10.1111/imb.v28.5URL [本文引用: 1]
WANGM, XIAOY, LIY, WANG XY, QI SZ, WANGY, ZHAO LW, WANGK, PENG WJ, LUO GZ, XUE XF, JIA GF, WU LM. RNA m6A modification functions in larval development and caste differentiation in honeybee (Apis mellifera) Cell Reports, 2021, 34(1):108580. DOI:10.1016/j.celrep.2020.108580URL [本文引用: 2]
GUOJ, TANG HW, LIJ, PERRIMONN, YAND. Xio is a component of theDrosophila sex determination pathway and RNA N6-methyladenosine methyltransferase complex Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(14):3674-3679. [本文引用: 1]
LENCET, AKHTARJ, BAYERM, SCHMIDK, SPINDLERL, HO CH, KREIMN, ANDRADE-NAVARRO MA, POECKB, HELMM, ROIGNANT JY. m6A modulates neuronal functions and sex determination in Drosophila Nature, 2016, 540(7632):242-247. DOI:10.1038/nature20568URL [本文引用: 5]
GRANADINOB, SAN JUANA, SANTAMARIAP, SANCHEZL. Evidence of a dual function infl(2)d, a gene needed forsex-lethal expression in Drosophila melanogaster Genetics, 1992, 130(3):597-612. DOI:10.1093/genetics/130.3.597URL [本文引用: 1]
ORTEGAA, NIKSICM, BACHIA, WILMM, SANCHEZL, HASTIEN, VALCARCELJ. Biochemical function of female-lethal (2)D/Wilms’ tumor suppressor-1-associated proteins in alternative pre-mRNA splicing Journal of Biological Chemistry, 2003, 278(5):3040-3047. DOI:10.1074/jbc.M210737200URL [本文引用: 1]
GRANADINOB, CAMPUZANOS, SANCHEZL. The Drosophila melanogaster fl(2)dgene is needed for the female-specific splicing ofsex-lethal RNA The EMBO Journal, 1990, 9(8):2597-2602. DOI:10.1002/embj.1990.9.issue-8URL [本文引用: 2]
GRANADINOB, PENALVA LO, SANCHEZL. The gene fl(2)d is needed for the sex-specific splicing of transformer pre-mRNA but not fordouble-sex pre-mRNA in Drosophila melanogaster Molecular and General Genetics, 1996, 253(1/2):26-31. DOI:10.1007/s004380050292URL [本文引用: 1]
BURNETTE JM, HATTON AR, LOPEZ AJ. Trans-acting factors required for inclusion of regulated exons in the Ultrabithorax mRNAs of Drosophila melanogaster Genetics, 1999, 151(4):1517-1529. DOI:10.1093/genetics/151.4.1517URL [本文引用: 1]
ORTEGAA. Localization of the Drosophilaprotein fl(2)d in somatic cells and female gonads Cell and Tissue Research, 2005, 320(2):361-367. DOI:10.1007/s00441-004-1049-5URL [本文引用: 1]
TRAUTW, NIIMIT, IKEOK, SAHARAK. Phylogeny of the sex-determining gene sex-lethal in insects Genome, 2006, 49(3):254-262. DOI:10.1139/g05-107URL [本文引用: 1]
FUJIIT, SHIMADAT. Sex determination in the silkworm, Bombyx mori: A female determinant on the W chromosome and the sex-determining gene cascade Seminars in Cell and Developmental Biology, 2007, 18(3):379-388. DOI:10.1016/j.semcdb.2007.02.008URL [本文引用: 1]
TANG WQ, YU LY, HE WY, YANGG, KE FS, BAXTER SW, YOU SJ, DOUGLAS CJ, YOU MS. DBM-DB: The diamondback moth genome database Database, 2014, 2014: bat087. DOI:10.1093/database/bat087URL [本文引用: 1]
CHENW, DONG YH, SAQIB H SA, VASSEURL, ZHOU WW, ZHENGL, LAI YF, MA XL, LIN LY, XU XJ, BAI JL, HE WY, YOU MS. Functions of duplicated glucosinolate sulfatases in the development and host adaptation of Plutella xylostella Insect Biochemistry and Molecular Biology, 2020, 119:103316. DOI:10.1016/j.ibmb.2020.103316URL [本文引用: 2]
PING XL, SUN BF, WANGL, XIAOW, YANGX, WANG WJ, ADHIKARIS, SHIY, LVY, CHEN YS, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase Cell Research, 2014, 24(2):177-189. DOI:10.1038/cr.2014.3URL [本文引用: 1]
FUKUSUMIY, NARUSEC, ASANOM. WTAP is required for differentiation of endoderm and mesoderm in the mouse embryo Developmental Dynamics, 2008, 237(3):618-629. DOI:10.1002/dvdy.v237:3URL [本文引用: 1]
HORIUCHIK, UMETANIM, MINAMIT, OKAYAMAH, TAKADAS, YAMAMOTOM, ABURATANIH, REID PC, HOUSMAN DE, HAMAKUBOT, KODAMAT. Wilms’ tumor 1-associating protein regulates G2/M transition through stabilization of cyclin A2 mRNA Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(46):17278-17283. [本文引用: 1]
NARUSEC, FUKUSUMIY, KAKIUCHID, ASANOM. A novel gene trapping for identifying genes expressed under the control of specific transcription factors Biochemical and Biophysical Research Communications, 2007, 361(1):109-115. DOI:10.1016/j.bbrc.2007.06.161URL [本文引用: 1]
HONGAY CF, ORR-WEAVER TL. Drosophila inducer of MEiosis 4 (IME4) is required for Notch signaling during oogenesis Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(36):14855-14860. [本文引用: 1]
KAN LJ, GROZHIK AV, VEDANAYAGAMJ, PATIL DP, PANGN, LIM KS, HUANG YC, JOSEPHB, LIN CJ, DESPICV, et al. The m6A pathway facilitates sex determination in Drosophila Nature Communications, 2017, 8:15737. DOI:10.1038/ncomms15737URL [本文引用: 2]
SINGHA, BUEHNER NA, LINH, BARANOWSKI KJ, FINDLAY GD, WOLFNER MF. Long-term interaction betweenDrosophila sperm and sex peptide is mediated by other seminal proteins that bind only transiently to sperm Insect Biochemistry and Molecular Biology, 2018, 102:43-51. DOI:10.1016/j.ibmb.2018.09.004URL [本文引用: 1]
WIGBYS, SIROT LK, LINKLATER JR, BUEHNERN, CALBOLI F CF, BRETMANA, WOLFNER MF, CHAPMANT. Seminal fluid protein allocation and male reproductive success Current Biology, 2009, 19(9):751-757. DOI:10.1016/j.cub.2009.03.036URL [本文引用: 1]
SIMMONS LW, LOVEGROVEM. Socially cued seminal fluid gene expression mediates responses in ejaculate quality to sperm competition risk Proceedings of the Royal Society B: Biological Sciences, 2017, 284(1861):20171486. [本文引用: 1]
NEUBAUM DM, WOLFNER MF. Mated Drosophila melanogaster females require a seminal fluid protein, Acp36DE, to store sperm efficiently Genetics, 1999, 153(2):845-857. DOI:10.1093/genetics/153.2.845URL [本文引用: 1]
SIROT LK, BUEHNER NA, FIUMERA AC, WOLFNER MF. Seminal fluid protein depletion and replenishment in the fruit fly, Drosophila melanogaster: An ELISA-based method for tracking individual ejaculates Behavioral Ecology and Sociobiology, 2009, 63(10):1505-1513. DOI:10.1007/s00265-009-0806-6URL [本文引用: 1]
GRANADINOB, SAN JUÁN AB, SÁNCHEZL. The gene fl(2)d is required for various Sxl-controlled processes in Drosophila females Roux’s Archives of Developmental Biology, 1991, 200(3):172-176. DOI:10.1007/BF00190237URL [本文引用: 1]
BOURTZISK, LEES RS, HENDRICHSJ, VREYSEN M JB. More than one rabbit out of the hat: Radiation, transgenic and symbiont- based approaches for sustainable management of mosquito and tsetse fly populations Acta Tropica, 2016, 157:115-130. DOI:10.1016/j.actatropica.2016.01.009URL [本文引用: 1]
FLORES HA, O’NEILL SL. Controlling vector-borne diseases by releasing modified mosquitoes Nature Reviews. Microbiology, 2018, 16(8):508-518. DOI:10.1038/s41579-018-0025-0URL [本文引用: 1]
MEZA JS, UL HAQI, VREYSEN M JB, BOURTZISK, KYRITSIS GA, CÁCERESC. Comparison of classical and transgenic genetic sexing strains of Mediterranean fruit fly (Diptera: Tephritidae) for application of the sterile insect technique PLoS ONE, 2018, 13(12):e0208880. DOI:10.1371/journal.pone.0208880URL [本文引用: 1]
ZHANG ZJ, NIU BL, JI DF, LI MW, LIK, JAMES AA, TAN AJ, HUANG YP. Silkworm genetic sexing through W chromosome- linked, targeted gene integration Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(35):8752-8756. [本文引用: 2]
GANTZ VM, JASINSKIENEN, TATARENKOVAO, FAZEKASA, MACIAS VM, BIERE, JAMES AA. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(49):E6736-6743. [本文引用: 1]
HAMMONDA, GALIZIR, KYROUK, SIMONIA, SINISCALCHIC, KATSANOSD, GRIBBLEM, BAKERD, MAROISE, RUSSELLS, BURTA, WINDBICHLERN, CRISANTIA, NOLANT. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae Nature Biotechnology, 2016, 34(1):78-83. DOI:10.1038/nbt.3439URL [本文引用: 1]
 ,, 王贝贝, 赖颖芳, 杨菲颖, 尤民生
,, 王贝贝, 赖颖芳, 杨菲颖, 尤民生

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT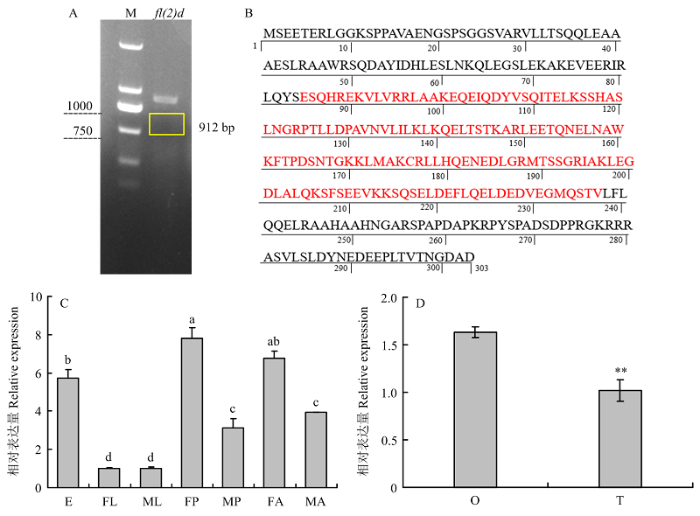 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT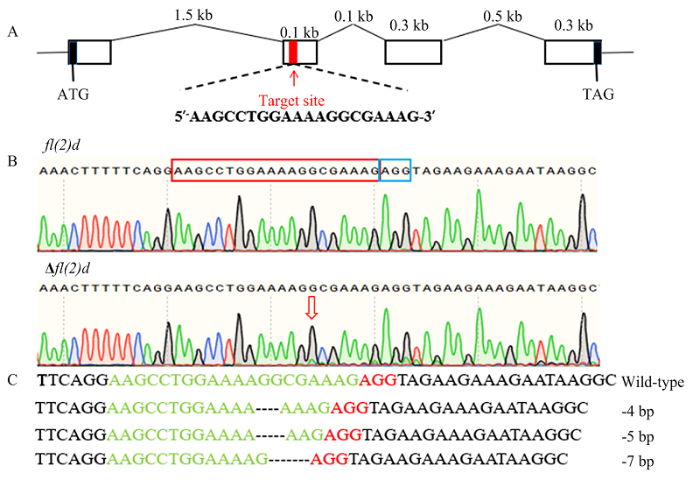 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT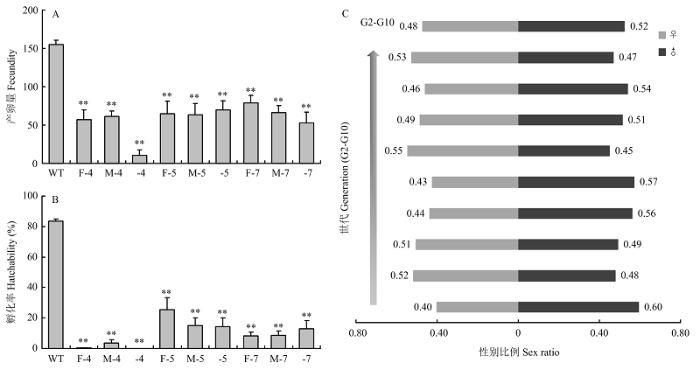 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT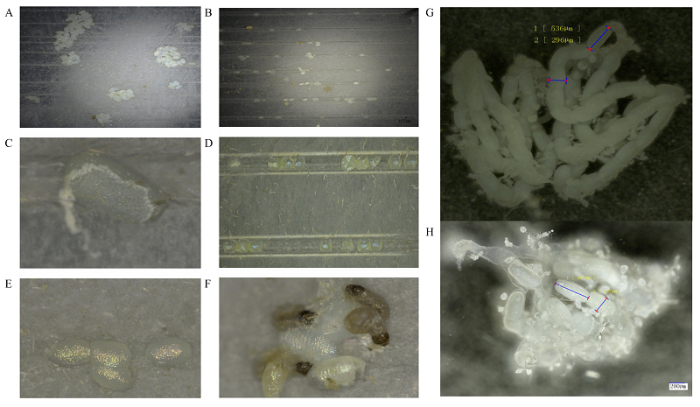 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT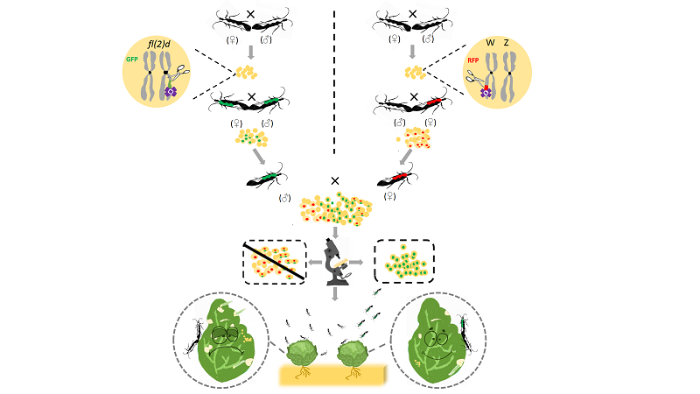 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}