Genetic Research Advances on Maize Stalk Lodging Resistance
WANG XiaQing,, SONG Wei,, ZHANG RuYang, CHEN YiNing, SUN Xuan, ZHAO JiuRan,Maize Research Center, Beijing Academy of Agriculture & Forestry Sciences/Beijing Key Laboratory of Maize DNA Fingerprinting and Molecular Breeding, Beijing 100097
Abstract Maize stalk lodging has a great adverse effect on yield, quality and mechanized harvesting, and is one of the main problems to be solved urgently in current maize production and breeding. Strengthening the research on the lodging resistance of maize stalk will have great significance for improving the lodging resistance of maize. In this paper, we summarize the main factors affecting maize stalk lodging resistance, and their genetic mechanisms. The stalk lodging resistance is closely related to the stalk strength. The greater the stalk strength, the stronger the lodging resistance. The stalk strength is affected by the developmental stage, the internal and external structures of the stalk, and the components of the stalk cell wall. The meristem zone has vigorously dividing cells and is easily broken. After entering the reproductive growth, the rind and sclerenchyma tissue of the stalk are thickened, the vascular bundles are mature, and thus the stalk strength is enhanced. The main components of the stalk cell wall, including cellulose, hemicellulose, lignin, soluble sugars, inorganic substances, can improve the strength of the stalk. To date, based on the high-throughput phenotyping platforms, various maize linkage and natural populations, and mapping methods, researchers have identified a series of QTLs and candidate genes that affect stalk morphology, strength, and cell wall components. The studies have shown that the haplotype-based mapping method is better than SNP-based mapping method. Meta-QTL analysis integrates the mapping results of different genetic populations and can improve the versatility of QTLs. The genetic basis of stalk strength is complex, which is determined by polygenes with minor effect and additive effect. Candidate genes in the QTLs involve cell wall metabolism, transcription factors, protein kinases, and so on. MAIZEWALL is an important database of genes related to maize cell wall. So far, the database contains 1 156 candidate genes related to maize cell wall biology, which provides a powerful resource for research in this field. A series of genes affecting cell wall components, stalk morphology and stalk strength in maize have been identified. Their functions of these genes are related to cellulose synthesis pathways, such as genes of cellulose synthase, Cobra, glycosyltransferase and ribose transport; phenylpropane pathway genes, such as genes regulating bm1-bm5; plant hormones genes, such as genes related to gibberellin, auxin and brassinosteroid; transcription factors such as NAC, MYB; miRNA (ZmmiR528) and F-box genes (stiff1). In the future research, it is needed to explore the mechanical mechanism of stalk lodging at different developmental stages. Develop diverse natural populations and breeding materials for genetic analysis. Employ a various of mapping strategies to improve the efficiency of identification of the QTL and genes related to lodging resistance. Design various molecular markers based on the favorable alleles to improve the molecular marker assisted selection for lodging resistance. These efforts will promote the research of the genetic mechanism of stalk lodging resistance, and provide a reference for the molecular breeding of new varieties with strong lodging resistance. Keywords:maize;lodging;stalk;cell wall;genetic mechanism
PDF (2585KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王夏青, 宋伟, 张如养, 陈怡凝, 孙轩, 赵久然. 玉米茎秆抗倒伏遗传的研究进展[J]. 中国农业科学, 2021, 54(11): 2261-2272 doi:10.3864/j.issn.0578-1752.2021.11.002 WANG XiaQing, SONG Wei, ZHANG RuYang, CHEN YiNing, SUN Xuan, ZHAO JiuRan. Genetic Research Advances on Maize Stalk Lodging Resistance[J]. Scientia Acricultura Sinica, 2021, 54(11): 2261-2272 doi:10.3864/j.issn.0578-1752.2021.11.002
A:拔节期玉米根倒;B:抽雄前期茎弯曲和茎折;C:抽雄前期茎折;D:灌浆期的根茎复合倒伏;E:籽粒成熟后根倒伏和茎弯曲;F:籽粒成熟后茎秆基部第3节茎折 Fig. 1The common types of lodging in maize production
A: Root lodging in the jointing stage; B: The stalk bending and fracture in the early stage of tasseling; C: The stalk fracture in the early stage of tasseling; D: The compound lodging of the root and stalk after the grain filling stage; E: The root lodging and stalk bending after maturity; F: The fracture position was at the third internode after maturity
目前,玉米茎秆强度的研究多集中于对茎秆硬度遗传位点的挖掘,而关于茎秆柔韧性的研究较少(表1)。茎秆强度的遗传力较低,需要在不同环境下检测,以最优线性无偏预测方法(best linear unbiased prediction,BLUP)来提高表型的准确性,基于BLUP数据得到的QTL结果优于单环境下的定位结果[8,9,10]。茎皮穿刺强度比茎秆弯折强度的遗传力高,且该性状与茎秆强度和倒伏的相关性较高[12]。茎秆强度的遗传基础复杂,受大量微效位点控制,各位点间存在加性效应,对茎秆强度的改良可以通过多个优良基因的聚合实现[11,13]。目前,已经鉴定到控制玉米茎秆强度的基因stiff1[30]。针对玉米茎秆柔韧性,WANG等[14]鉴定到一个与茎秆弯折角度相关的QTL,并提出候选基因可能与RING/U box泛素蛋白和MADS转录因子相关。
Table 1 表1 表1玉米茎秆抗倒伏相关性状遗传定位统计 Table 1The summary of genetic studies for stalk lodging resistance traits in maize
XUEJ, WANGK R, XIER Z, GOUL, ZHANGW F, MINGB, HOUP, LIS K. Research progress of maize lodging during late stage Scientia Agricultura Sinica, 2018,51(10):1845-1854. (in Chinese) [本文引用: 2]
SUNS X, GUW L, DAIJ Y. Effect of density on corn lodging and yield Journal of Shenyang Agricultural University, 1989,20:413-416. (in Chinese) [本文引用: 2]
WANGH L, WUR H, ZHUK, ZHANGY C, ZHANGY J, SUNJ W. Reviews of causes and control of maize lodging Journal of Henan Agricultural Sciences, 2011,40(10):1-5. (in Chinese) [本文引用: 2]
WANGL M, YAOG Q, MUC H, LIJ S, DAIJ R. Advances in genetic research of maize lodging resistance Journal of Maize Sciences, 2011,19(4):1-4. (in Chinese) [本文引用: 1]
JINY J, LIH P, ANP P, CHENGS X, ZHAOX Y, YUT Y, LIC H. Research progress on the lodging resistance of maize Journal of Maize Sciences, 2019,27(2):94-98. (in Chinese) [本文引用: 1]
GOUL, ZHAOM, HUANGJ J, ZHANGB, LIT, SUNR. Bending mechanical properties of stalk and lodging-resistance of maize (Zea mays L.) Acta Agronomica Sinica, 2008,34(4):653-661. (in Chinese) DOI:10.3724/SP.J.1006.2008.00653URL [本文引用: 3]
WANGX, ZHANGR, SHIZ, ZHANGY, SUNX, JIY L, ZHAOY, WANGJ, ZHANGY, XINGJ, WANGY, WANGR, SONGW, ZHAOJ. Multi-omics analysis of the development and fracture resistance for maize internode Scientific Reports, 2019,9(1):8183. DOI:10.1038/s41598-019-44690-6URL [本文引用: 5]
HUH X, LIUW X, FUZ Y, HOMANNL, TECHNOWF, WANGH W, SONGC L, LIS T, MELCHINGERA E, CHENS J. QTL mapping of stalk bending strength in a recombinant inbred line maize population Theoretical and Applied Genetics, 2013,126(9):2257-2266. DOI:10.1007/s00122-013-2132-7URL [本文引用: 3]
PEIFFERJ A, FLINT-GARCIAS A, DELEON N, MCMULLENM D, KAEPPLERS M, BUCKLERE S. The genetic architecture of maize stalk strength PLoS ONE, 2013,8(6):e67066. DOI:10.1371/journal.pone.0067066URL [本文引用: 3]
LIK, YANJ, LIJ, YANGX. Genetic architecture of rind penetrometer resistance in two maize recombinant inbred line populations BMC Plant Biology, 2014,14(1):152. DOI:10.1186/1471-2229-14-152URL [本文引用: 3]
ZHANGY, LIUP, ZHANGX, ZHENGQ, CHENM, GEF, LIZ, SUNW, GUANZ, LIANGT. Multi-locus genome-wide association study reveals the genetic architecture of stalk lodging resistance- related traits in maize Frontiers in Plant Science, 2018,9:611. DOI:10.3389/fpls.2018.00611URL [本文引用: 4]
WANGX, SHIZ, ZHANGR, SUNX, WANGJ, WANGS, ZHANGY, ZHAOY, SUA, LIC, WANGR, ZHANGY, WANGS, WANGY, SONGW, ZHAOJ. Stalk architecture, cell wall composition, and QTL underlying high stalk flexibility for improved lodging resistance in maize BMC Plant Biology, 2020,20(1):515. DOI:10.1186/s12870-020-02728-2URL [本文引用: 3]
KENDEH, VANDER KNAAP E, CHOH T. Deepwater rice: A model plant to study stem elongation Plant Physiology, 1998,118(4):1105-1110. DOI:10.1104/pp.118.4.1105URL [本文引用: 2]
SCOBBIEL, RUSSELLW, PROVANG J, CHESSONA. The newly extended maize internode: A model for the study of secondary cell wall formation and consequences for digestibility Journal of the Science of Food & Agriculture, 1993,61(2):217-225. [本文引用: 1]
ZHANGQ, CHEETAMUNR, DHUGGAK S, RAFALSKIJ A, TINGEYS V, SHIRLEYN J, TAYLORJ, HAYESK R, BEATTYM, BACICA. Spatial gradients in cell wall composition and transcriptional profiles along elongating maize internodes BMC Plant Biology, 2014,14(1):27. DOI:10.1186/1471-2229-14-27URL [本文引用: 1]
WANGT J, ZHANGL, HANQ, ZHENGF X, WANGT Q, FENGN N, WANGT X. Effects of stalk cell wall and tissue on the compressive strength of maize Plant Science Journal, 2015,33(1):109-115. (in Chinese) [本文引用: 1]
SINDHUA, LANGEWISCHT, OLEKA, MULTANID S, MCCANNM C, VERMERRISW, CARPITAN C, JOHALG. Maize brittle stalk2 encodes a COBRA-like protein expressed in early organ development but required for tissue flexibility at maturity Plant Physiology, 2007,145(4):1444-1459. DOI:10.1104/pp.107.102582URL [本文引用: 4]
JIAOS, HAZEBROEKJ P, CHAMBERLINM A, PERKINSM, SANDHUA S, GUPTAR, SIMCOXK D, YINGHONGL, PRALLA, HEETLANDL, MEELEYR B, MULTANID S. Chitinase-like1 plays a role in stalk tensile strength in maize Plant Physiology, 2019,181(3):1127-1147. DOI:10.1104/pp.19.00615URL [本文引用: 3]
CAOQ J, CUIJ H, WANGH Y, WENH J, GAOY N, LUOL H, HANH F. Effect of water treatments after jointing stage on plant characters and water use efficiency of maize Maize Science, 2011,19(3):105-109. (in Chinese) [本文引用: 1]
QIUX Y, HUS W. “Smart” materials based on cellulose: A review of the preparations, properties, and applications Materials, 2013,6(3):738-781. DOI:10.3390/ma6030738URL [本文引用: 1]
ZHONGR, RIPPERGERA, YEZ H. Ectopic deposition of lignin in the pith of stems of two Arabidopsis mutants Plant Physiology, 2000,123(1):59-70. DOI:10.1104/pp.123.1.59URL [本文引用: 1]
YANGS M, XIEL, ZHENGS L, LIJ, YUANJ C. Effects of nitrogen rate and transplanting density on physical and chemical characteristics and lodging resistance of culms in hybrid rice Acta Agronomica Sinica, 2009,35(1):93-103. (in Chinese) DOI:10.3724/SP.J.1006.2009.00093URL [本文引用: 1]
LIN, LIJ M, ZHAIZ X, LIZ H, DUANL S. Effects of chemical regulator on the lodging resistance traits, agricultural character and yield of maize Maize Science, 2010,18(6):38-42. (in Chinese) [本文引用: 1]
ZHANGZ, ZHANGX, LINZ, WANGJ, LIUH, ZHOUL, ZHONGS, LIY, ZHUC, LAIJ, LIX, YUJ, LINZ. A large transposon insertion in the stiff1 promoter increases stalk strength in maize The Plant Cell, 2020,32(1):152-165. DOI:10.1105/tpc.19.00486URL [本文引用: 5]
LIK, WANGH, HUX, LIUZ, WUY, HUANGC. Genome-wide association study reveals the genetic basis of stalk cell wall components in maize PLoS ONE, 2016,11(8):e0158906. DOI:10.1371/journal.pone.0158906URL [本文引用: 2]
WANGH, LIK, HUX, LIUZ, WUY, HUANGC. Genome-wide association analysis of forage quality in maize mature stalk BMC Plant Biology, 2016,16(1):227. DOI:10.1186/s12870-016-0919-9URL [本文引用: 2]
WANGQ, LIK, HUX, SHIH, LIUZ, WUY, WANGH, HUANGC. Genetic analysis and QTL mapping of stalk cell wall components and digestibility in maize recombinant inbred lines from B73×By804 The Crop Journal, 2020,8(1):132-139. DOI:10.1016/j.cj.2019.06.009URL [本文引用: 2]
PENNINGB W, SYKESR W, BABCOCKN C, DUGARDC K, HELDM A, KLIMEKJ F, SHREVEJ T, FOWLERM, ZIEBELLA, DAVISM F, DECKERS R, TURNERG B, MOSIERN S, SPRINGERN M, THIMMAPURAMJ, WEILC F, MCCANNM C, CARPITAN C. Genetic determinants for enzymatic digestion of lignocellulosic biomass are independent of those for lignin abundance in a maize recombinant inbred population Plant Physiology, 2014,165(4):1475-1487. DOI:10.1104/pp.114.242446URL [本文引用: 2]
BARRIèREY, THOMASJ, DENOUED. QTL mapping for lignin content, lignin monomeric composition, p-hydroxycinnamate content, and cell wall digestibility in the maize recombinant inbred line progeny F838×F286 Plant Science, 2008,175(4):585-595. DOI:10.1016/j.plantsci.2008.06.009URL [本文引用: 2]
TRUNTZLERM, BARRIèREY, SAWKINSMC, LESPINASSED, BETRANJ, CHARCOSSETA, MOREAUL. Meta-analysis of QTL involved in silage quality of maize and comparison with the position of candidate genes Theoretical and Applied Genetics, 2010,121(8):1465-1482. DOI:10.1007/s00122-010-1402-xURL [本文引用: 2]
BIANY L, SUND L, GUX, WANGY G, YINZ T, DENGD X, WANGY Q, WUF F, LIG S. Identification of QTL for stalk sugar-related traits in a population of recombinant inbred lines of maize Euphytica, 2014,198(1):79-89. DOI:10.1007/s10681-014-1085-5URL [本文引用: 2]
MALDONADOC, MORAF, SCAPIMC A, COANM. Genome-wide haplotype-based association analysis of key traits of plant lodging and architecture of maize identifies major determinants for leaf angle: hapLA4 PLoS ONE, 2019,14(3):e0212925. DOI:10.1371/journal.pone.0212925URL [本文引用: 2]
LIUF P, QUW L, FANGH Y, LIL L, JINF X, WUW L. Analysis of Meta- QTL and candidate genes related to stem diameter in maize Journal of Northeast Agricultural Sciences, 2019,44(5):30-33. (in Chinese) [本文引用: 2]
HUANGC, CHENQ, XUG, XUD, TIANJ, TIANF. Identification and fine mapping of quantitative trait loci for the number of vascular bundle in maize stem Journal of Integrative Plant Biology, 2016,58(1):81-90. DOI:10.1111/jipb.12358URL [本文引用: 2]
MAZAHERIM, HECKWOLFM, VAILLANCOURTB, GAGEJ L, BURDOB, HECKWOLFS, BARRYK, LIPZENA, RIBEIROC B, KONOT J Y. Genome-wide association analysis of stalk biomass and anatomical traits in maize BMC Plant Biology, 2019,19(1):45. DOI:10.1186/s12870-019-1653-xURL [本文引用: 3]
ZHANGY, WANGJ, DUJ, ZHAOY, LUX, WENW, GUS, FANJ, WANGC, WUS, WANGY, LIAOS, ZHAOC, GUOX. Dissecting the phenotypic components and genetic architecture of maize stem vascular bundles using high-throughput phenotypic analysis Plant Biotechnology Journal, 2021,19(1):35-50 . DOI:10.1111/pbi.v19.1URL [本文引用: 3]
GUILLAUMIES, SANCLEMENTEH, DESWARTEC, MARTINEZY, LAPIERREC, MURIGNEUXA, BARRIEREY, PICHONM, MAIZEWALLG D. database and developmental gene expression profiling of cell wall biosynthesis and assembly in maize Plant Physiology, 2006,143(1):339-363. DOI:10.1104/pp.106.086405URL [本文引用: 1]
MALEKIS S, MOHAMMADIK, JIK S. Characterization of cellulose synthesis in plant cells Scientific World Journal, 2016,2016:8641373. [本文引用: 2]
KOTAKET, AOHARAT, HIRANOK, SATOA, KANEKOY, TSUMURAYAY, TAKATSUJIH, KAWASAKIS. Rice Brittle culm 6 encodes a dominant-negative form of CesA protein that perturbs cellulose synthesis in secondary cell walls Journal of Experimental Botany, 2011,62(6):2053-2062. DOI:10.1093/jxb/erq395URL [本文引用: 1]
ZHOUY, LIS, QIANQ, ZENGD, ZHANGM, GUOL, LIUX, ZHANGB, DENGL, LIUX, LUOG, WANGX, LIJ. BC10, a DUF266-containing and Golgi-located type II membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.) The Plant Journal, 2009,57(3):446-462. DOI:10.1111/tpj.2009.57.issue-3URL [本文引用: 1]
ZHANGB, LIUX, QIANQ, LIUL, DONGG, XIONGG, ZENGD, ZHOUY. Golgi nucleotide sugar transporter modulates cell wall biosynthesis and plant growth in rice Proceedings of the National Academy of Sciences of the United States of America, 2011,108(12):5110-5115. [本文引用: 1]
YOONJ, CHOIH, ANG. Roles of lignin biosynthesis and regulatory genes in plant development Journal of Integrative Plant Biology, 2015,57:902-912. DOI:10.1111/jipb.v57.11URL [本文引用: 1]
HALPINC, HOLTK, CHOJECKIJ, OLIVERD, CHABBERTB, MONTIESB, EDWARDSK, BARAKATEA, FOXONG A. Brown-midrib maize (bm1)-A mutation affecting the cinnamyl alcohol dehydrogenase gene The Plant Journal, 1998,14(5):545-553. DOI:10.1046/j.1365-313X.1998.00153.xURL [本文引用: 2]
TANGH M, LIUS, HILLSKINNERS, WUW, REEDD, YEHC T, NETTLETOND, SCHNABLEP S. The maize brown midrib2 (bm2) gene encodes a methylenetetrahydrofolate reductase that contributes to lignin accumulation The Plant Journal, 2014,77(3):380-392. DOI:10.1111/tpj.2014.77.issue-3URL [本文引用: 2]
VIGNOLSF, RIGAUJ, TORRESM A, CAPELLADESM, PUIGDOMèNECHP. The brown midrib3 (bm3) mutation in maize occurs in the gene encoding caffeic acid O-methyltransferase The Plant Cell, 1995,7(4):407-416. [本文引用: 1]
LIL, HILL-SKINNERS, LIUS, BEUCHLED, TANGH M, YEHC T, NETTLETOND, SCHNABLEP S. The maize brown midrib4 (bm4) gene encodes a functional folylpolyglutamate synthase The Plant Journal, 2015,81(3):493-504. DOI:10.1111/tpj.2015.81.issue-3URL [本文引用: 1]
XIONGW, WUZ, LIUY, LIY, SUK, BAIZ, GUOS, HUZ, ZHANGZ, BAOY, SUNJ, YANGG, FUC. Mutation of 4-coumarate: coenzyme A ligase 1 gene affects lignin biosynthesis and increases the cell wall digestibility in maize brown midrib5 mutants Biotechnology for Biofuels, 2019,12:82. DOI:10.1186/s13068-019-1421-zURL [本文引用: 2]
BENSENR J, JOHALG S, CRANEV C, TOSSBERGJ T, SCHNABLEP S, MEELEYR B, BRIGGSS P. Cloning and characterization of the maize An1 gene The Plant Cell, 1995,7(1):75-84. [本文引用: 2]
WINKLERR G, HELENTJARIST. The maize Dwarf3 gene encodes a cytochrome P450-mediated early step in gibberellin biosynthesis The Plant Cell, 1995,7(8):1307-1317. [本文引用: 2]
CASSANIE, BERTOLINIE, BADONEF C, LANDONIM, GAVINAD, SIRIZZOTTIA, PILUR. Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of the dwarf8 gene Molecular Breeding, 2009,24(4):375-385. DOI:10.1007/s11032-009-9298-3URL [本文引用: 2]
HARTWIGT, CHUCKG, FUJIOKAS, KLEMPIENA, WEIZBAUERR, POTLURID P V, CHOES, JOHALG S, SCHULZB. Brassinosteroid control of sex determination in maize Proceedings of the National Academy of Sciences of the United States of America, 2011,108(49):19814-19819. [本文引用: 2]
MAKAREVITCHI, THOMPSONA, MUEHLBAUERG J, SPRINGERN M. Brd1 gene in maize encodes a brassinosteroid C-6 oxidase PLoS ONE, 2012,7(1):e30798. DOI:10.1371/journal.pone.0030798URL [本文引用: 2]
XINGA, GAOY, YEL, ZHANGW, CAIL, CHINGA, LLACAV, JOHNSONB E, LIUL, YANGX. A rare SNP mutation in Brachytic2 moderately reduces plant height and increases yield potential in maize Journal of Experimental Botany, 2015,66(13):3791-3802. DOI:10.1093/jxb/erv182URL [本文引用: 3]
AVILAL M, CERRUDOD, SWANTONC J, LUKENSL. Brevis plant1, a putative inositol polyphosphate 5-phosphatase, is required for internode elongation in maize Journal of Experimental Botany, 2016,67(5):1577-1588. DOI:10.1093/jxb/erv554URL [本文引用: 2]
MULTANID S, BRIGGSS P, CHAMBERLINM A, BLAKESLEEJ J, MURPHYA S, JOHALG S. Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants Science, 2003,302(5642):81-84. DOI:10.1126/science.1086072URL [本文引用: 1]
ZHONGR, LEEC, ZHOUJ, MCCARTHYR L, YEZ H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis The Plant Cell, 2008,20(10):2763-2782. DOI:10.1105/tpc.108.061325URL [本文引用: 2]
MITSUDAN, SEKIM, SHINOZAKIK, OHMETAKAGIM. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence The Plant Cell, 2005,17(11):2993-3006. DOI:10.1105/tpc.105.036004URL [本文引用: 1]
OLSENA N, ERNSTH A, LEGGIOL L, SKRIVERK. NAC transcription factors: Structurally distinct, functionally diverse Trends in Plant Science, 2005,10(2):79-87. DOI:10.1016/j.tplants.2004.12.010URL [本文引用: 1]
XIAOW, YANGY, YUJ. ZmNST3 and ZmNST4 are master switches for secondary wall deposition in maize (Zea mays L.) Plant Science, 2018; 266:83-94. DOI:10.1016/j.plantsci.2017.03.012URL [本文引用: 1]
SUNQ, LIUX, YANGJ, LIUW, DUQ, WANGH, FUC, LIW X. MicroRNA528 affects lodging resistance of maize by regulating lignin biosynthesis under nitrogen-luxury conditions Molecular Plant, 2018,11(6):806-814. DOI:10.1016/j.molp.2018.03.013URL [本文引用: 1]
ABD-HAMIDN A, AHMAD-FAUZIM I, ZAINALZ, ISMAILI. Diverse and dynamic roles of F-box proteins in plant biology Planta, 2020,251(3):68. DOI:10.1007/s00425-020-03356-8URL [本文引用: 1]
CAVANAGHC, MORELLM, MACKAYI, POWELLW. From mutations to MAGIC: resources for gene discovery, validation and delivery in crop plants Current Opinion in Plant Biology, 2008,11(2):215-221. DOI:10.1016/j.pbi.2008.01.002URL [本文引用: 1]
SUC F, ZHAOT J, GAIJ Y. Simulation Comparisons of effectiveness among QTL mapping procedures of different statistical genetic models Acta Agronomica Sinica, 2010,36(7):1100-1107. (in Chinese) DOI:10.3724/SP.J.1006.2010.01100URL [本文引用: 1]
WENJ Y, FENGY J, ZHANGJ, ZHANGY M. Advances on methodologies for genome-wide association studies in plants Acta Agronomica Sinica, 2016,42(7):945-956. (in Chinese) DOI:10.3724/SP.J.1006.2016.00945URL [本文引用: 1]
 ,, 宋伟
,, 宋伟

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT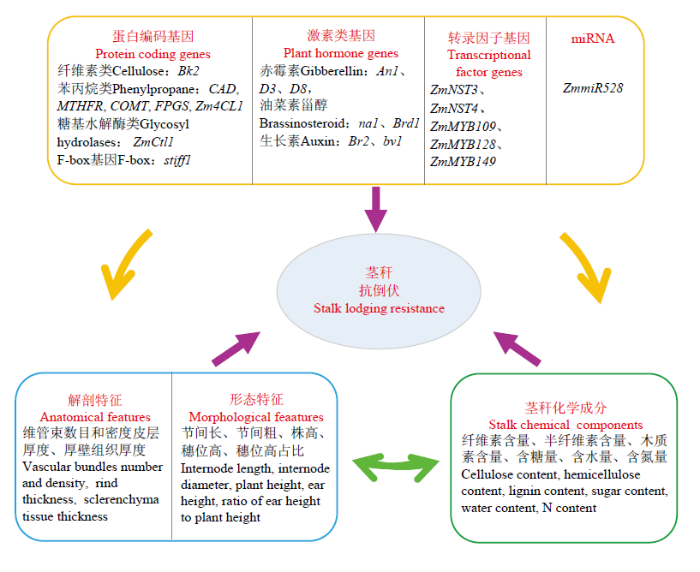 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}