 ,, 罗燚佳, 向雯, 张明磊, 张剑侠,西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业部西北地区园艺作物生物与种质创制重点实验室,陕西杨凌 712100
,, 罗燚佳, 向雯, 张明磊, 张剑侠,西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业部西北地区园艺作物生物与种质创制重点实验室,陕西杨凌 712100Rescue and Molecular Marker Assisted-Selection of the Cold-Resistant Seedless Grape Hybrid Embryo
ZHU PeiPei,, LUO YiJia, XIANG Wen, ZHANG MingLei, ZHANG JianXia,College of Horticulture, Northwest Agriculture and Forestry University/State Key Laboratory of Crop Stress Biology in Arid Areas/Key Laboratory of Horticultural Plant Germplasm Resource Utilization in Northwest China, Ministry of Agriculture, Yangling 712100, Shaanxi通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-06-30接受日期:2020-08-28网络出版日期:2021-03-16
| 基金资助: |
Received:2020-06-30Accepted:2020-08-28Online:2021-03-16
作者简介 About authors
朱佩佩,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1754KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
朱佩佩, 罗燚佳, 向雯, 张明磊, 张剑侠. 抗寒无核葡萄杂种胚挽救及分子标记辅助选择[J]. 中国农业科学, 2021, 54(6): 1218-1228 doi:10.3864/j.issn.0578-1752.2021.06.012
ZHU PeiPei, LUO YiJia, XIANG Wen, ZHANG MingLei, ZHANG JianXia.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】无核葡萄是育种的一个重要目标。目前世界上栽培的无核葡萄品种大多属于欧亚种葡萄(Vitis vinifera L.),品质优良,但抗寒性差,生产上缺乏抗寒无核葡萄品种,限制了其在寒冷地区的发展。原产中国的野生山葡萄(V. amurensis Rupr.)可耐 -40℃的低温,是葡萄属中极为重要的抗寒种质资源[1,2],此外,原产北美洲的美洲葡萄(V. labrusca L.)也是重要的抗寒种质资源[2]。利用抗寒的欧山杂种(V. vinifera×V. amurensis)或欧美杂种(V. vinifera×V. labrusca)作父本,与种子败育型欧亚种无核葡萄品种作母本杂交,借助胚挽救技术,在杂种胚败育前提供种胚正常发育所需的营养物质使其萌发成苗,是创制抗寒无核葡萄新种质的有效途径,将为进一步选育抗寒无核葡萄新品种奠定基础。【前人研究进展】1982年,美国葡萄育种家RAMMING和EMERSHAD[3]首次以无核葡萄品种的胚珠进行离体培养,获得了2株杂种,标志着无核葡萄胚挽救技术的创立。由于传统的无核葡萄育种只能以有核品种作母本与无核品种作父本杂交,杂交后代中无核杂种的几率极低,育种效率低,周期长;而胚挽救技术以无核葡萄作母本与无核或有核品种作父本杂交,扩大了母本选择的范围,提高了育种的效率,从而使该技术在许多国家得到广泛的研究和应用[4,5,6,7]。迄今,国内外已通过该技术选育出‘Melissa’‘Autumn Crisp’‘Early Sugar’‘秦红无核’和‘秦红1号’等47个无核葡萄新品种[8]。作为一项新的无核葡萄育种技术,影响其成功的因素很多,如父母本基因型、花前喷施激素、取样时期、基本培养基及其形态、添加有机物种类与浓度、培养条件等[9,10,11,12,13]。亲本基因型是影响胚挽救效果的一个关键因素,特别是母本基因型,前人研究认为‘Flame Seedless’[4]、‘Ruby Seedless’[4,9]、‘Perlette’[9]、‘秦红2号’[9]和‘Askari’[13]等适宜作胚挽救母本。徐海英等[6]研究认为胚过早发生败育的品种不适宜作胚挽救的母本,如‘Thompson Seedless’和‘Himrod’等,但POMMER等[12]认为早熟和中熟品种比晚熟品种作母本的胚挽救效果更好。此外,父本对胚挽救效果也有一定的影响[9,10]。培养基及其形态是决定胚挽救成败的另一关键因素。RAZI等[13]认为NN培养基培养效果好,也有使用ER培养基作为胚发育培养基[10],POMMER等[12]使用Cain培养基作为胚发育培养基,但笔者课题组研究获得的MM3培养基(改良ER培养基)被认为更适合以中国野生葡萄作为父本的胚挽救育种[9,14]。关于培养基的形态,SPIEGEL-ROY等[11]研究认为固相培养基最好,但RAMMING等[15]认为液体培养基效果好。TIAN等[16]认为固-液双相培养基最适合作为胚发育培养基。由此可见,胚发育培养基及其形态有必要做进一步的研究。对于通过胚挽救技术获得的杂种,一般是在树体结果后通过田间鉴定是否为无核性状,来确定取舍,育种速度较慢,而利用分子标记技术可以实现对目标性状在幼苗期的鉴定,极大缩短品种选育时间[17]。目前已获得的葡萄无核基因分子标记主要有SCAR标记SCC8-1018[18]、SCF27-2000[19],RAPD标记GSLP1-569[20],SSR标记p3-VvAGL11-216[21]、VMC7F2-198[22]和VvSD10[23]。其中以SCF27-2000对无核葡萄品种检测具有较广泛的适用性[14,24]。【本研究切入点】针对目前无核葡萄胚挽救中胚发育率、萌发率和成苗率仍较低,以及生产上缺乏抗寒无核葡萄品种的问题,本研究对胚挽救的主要影响因素(母本和胚发育培养基)进行研究,配置无核品种×抗寒欧山杂种、无核品种×抗寒欧美杂种杂交组合,以筛选出适宜作胚挽救的母本,比较2种胚发育培养基MM3和ER的胚挽救效果,并筛选出适宜畸形苗转化为正常苗的培养基,同时对胚挽救后代进行早期无核性状的分子标记辅助选择。【拟解决的关键问题】通过比较母本基因型、筛选胚发育培养基、筛选畸形苗转化正常苗培养基及分子标记辅助选择,为提高无核葡萄胚挽救效率提供参考依据,同时创制抗寒无核葡萄新种质,为进一步选育抗寒无核葡萄新品种奠定材料基础。1 材料与方法
2018年4月至2019年5月在西北农林科技大学葡萄种质资源圃、旱区作物逆境生物学国家重点实验室与园艺学院葡萄育种与分子生物学实验室进行。1.1 亲本材料
共设计5个杂交组合,分别是‘秦红2号’ב00-1-5’、‘秦秀’ב00-1-5’、‘Ruby Seedless’בJupiter’、‘秦红1号’בJupiter’和‘Crimson Seedless’בJupiter’。其中父本‘Jupiter’为抗寒欧美杂交无核品种,引自美国;另一父本‘00-1-5’(‘玫瑰香’ב山葡萄黑龙江实生’)为抗寒欧山杂种有核优系,由笔者课题组选育。父、母本均为二倍体品种(系)。1.2 方法
1.2.1 葡萄田间杂交 在父本植株上一个花序中有5%小花开放时取花穗,制备新鲜花粉,然后干燥、低温保存。母本在开花前3 d,选择健壮植株上发育一致的花穗进行人工去雄(图1-A)。2—3 d后,于上午7:00—9:00进行授粉,连续授粉3 d以保证授粉效率。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1抗寒无核葡萄胚挽救过程
A:去雄后的花穗;B:胚;C—E:幼胚萌发成苗;F:胚挽救苗移栽至田间
Fig. 1Procedure charts of breeding new cold-resistant seedless grapes by embryo rescue
A: Flowers after emasculation; B: Embryo; C-E: Young embryo germinates into a seedling; F: Transplanting hybrid seedlings to field
1.2.2 胚珠培养 5个母本品种‘秦秀’‘Crimson Seedless’‘秦红2号’‘秦红1号’和‘Ruby Seedless’的取样时期分别为花后42 d[9]、51 d[9]、51 d[24]、51 d[24]和61 d[9]。对采集的幼果先用自来水进行表面冲洗,然后在超净工作台中依次用浓度为75%乙醇和1%次氯酸钠进行表面消毒,再用无菌水冲洗3次,最后用手术刀剥取胚珠,接种于胚发育培养基中。
将5个组合‘秦秀’ב00-1-5’、‘秦红2号’ב00-1-5’、‘Ruby Seedless’בJupiter’、‘Crimson Seedless’בJupiter’和‘秦红1号’בJupiter’的胚珠接种在MM3固-液双相培养基中,研究不同母本对胚挽救效果的影响。
将材料较多的2个组合‘秦红2号’ב00-1-5’和‘Ruby Seedless’בJupiter’的胚珠,分别接种在固-液双相培养基ER和MM3中,比较不同培养基对胚挽救效果的影响。
两种培养基添加物均为0.5 g?L-1水解酪蛋白+1 mmol?L-1丝氨酸+0.5 mg?L-1 GA3+1.0 mg?L-1 IAA+ 0.5 mg?L-1 6-BA+60 g?L-1蔗糖+3 g?L-1活性炭+7 g?L-1琼脂(pH 6),其中液相部分不加琼脂。采用容量为200 mL的培养瓶,每瓶接种20个胚珠。培养瓶上覆盖黑布进行遮光培养,温度为(25±1)℃,培养9—10周。
1.2.3 胚萌发培养 胚珠培养9—10周后,在超净工作台中借助解剖镜剥取裸胚,接种到胚萌发培养基中(图1-B),置于(25±1)℃、每日光照16 h的培养室培养。40 d后,胚萌发成苗(图1-C—E)。萌发培养基为WPM固体培养基,培养基成分为:WPM+0.2 mg?L-1 6-BA+0.1 mg?L-1 IAA+20 g?L-1蔗糖+1.5 g?L-1活性炭+7 g?L-1琼脂(pH 6)。
幼苗在炼苗移栽前需要进行继代培养,培养基为1/2 MS+0.2 mg?L-1 IBA+30 g?L-1蔗糖+7 g?L-1琼脂(pH 6)。
1.2.4 畸形苗观察及转化培养基筛选 胚萌发培养5—6周后,观察不能正常萌发的畸形苗,包括子叶扭曲无根苗、白化苗、玻璃化苗、矮小苗等,比较不同母本组合及同一组合分别在MM3和ER固-液双相培养基上的畸形苗率。
将‘秦红2号’ב00-1-5’组合产生的畸形苗,分别接种至添加0、1.0和1.6 mg?L-1 ZnSO4的2MS+0.2 mg?L-1 6-BA+0.1 mg?L-1 IAA培养基中,培养30 d后统计畸形苗转化为正常苗的百分率。
1.2.5 无核基因分子标记辅助选择 采用CTAB法提取杂交后代株系基因组DNA,利用葡萄无核基因SCAR标记SCF27-2000的引物(F:5′-CAGGTGGGA GTAGTGGAATG-3′;R:5′-CAGGTGGGAGTAAGA TTTGT-3′)对杂交组合亲本及后代进行PCR检测,凡能扩增出2 000 bp大小的条带即初步确定为具有无核性状。
PCR反应程序为:94℃预变性4 min;95℃变性30 s,62℃复性90 s,72℃延伸1 min,共35个循环;72℃延伸1 min,12℃终止。
1.2.6 数据统计与分析 记录各组合胚的发育数、萌发数、成苗数和畸形苗数,统计胚的发育率、萌发率、成苗率、畸形苗率及畸形苗转化率,计算方法如下:
胚的发育率(%)=从胚珠中剥取胚的个数/胚珠数×100
胚的萌发率(%)=萌发胚的个数/胚珠数×100
胚的成苗率(%)=成苗数/胚珠数×100
畸形苗率(%)= 畸形苗数/胚珠数×100
畸形苗转化率(%)=转化成的正常苗数/畸形苗数×100
使用SPSS 22.0软件,对同一杂交组合3次生物
学重复中胚的发育率、萌发率、成苗率、畸形苗率和畸形苗转化率进行差异显著性分析(P<0.05)。
2 结果
2.1 不同母本对胚挽救效果的影响
5个杂交组合中不同母本的胚发育率、萌发率和成苗率的结果(表1)显示,以MM3为胚发育培养基,当以‘00-1-5’为父本,‘秦红2号’为母本的杂交组合的胚发育率和成苗率显著高于‘秦秀’ב00-1-5’,分别为17.04%和7.41%。以‘Jupiter’为父本,‘Ruby Seedless’为母本胚挽救效果最好,发育率和成苗率为13.71%和10.67%;其次是‘秦红1号’为母本的杂交组合,而‘Crimson Seedless’作母本胚的发育率和成苗率最低,与其他两个组合均具有显著性差异(P<0.05)。Table 1
表1
表1不同母本对胚挽救效果的影响
Table 1
| 杂交组合 Crossing combination | 胚珠数 No. of ovules | 剥胚数 No. of embryos developed | 萌发数 No. of embryos germinated | 成苗数 No. of normal seedings | 发育率 Proportion of embryos developed (%) | 萌发率 Proportion of embryos germinated (%) | 成苗率 Proportion of seedings (%) |
|---|---|---|---|---|---|---|---|
| 秦秀×00-1-5 Qinxiu×00-1-5 | 427 | 18 | 11 | 6 | 4.22e | 2.58d | 1.41e |
| 秦红2号×00-1-5 Qinhong No.2×00-1-5 | 405 | 69 | 38 | 30 | 17.04a | 9.38b | 7.41b |
| Crimson Seedless × Jupiter | 398 | 38 | 27 | 7 | 9.55d | 6.78c | 1.76d |
| Ruby Seedless × Jupiter | 525 | 72 | 56 | 56 | 13.71c | 10.67a | 10.67a |
| 秦红1号×Jupiter Qinhong No.1×Jupiter | 403 | 58 | 43 | 19 | 14.39b | 10.67a | 4.71c |
| 总计 Total | 2158 | 255 | 175 | 118 | 11.82 | 8.11 | 5.47 |
新窗口打开|下载CSV
2.2 不同发育培养基对胚挽救效果的影响
将‘Ruby Seedless’בJupiter’和‘秦红2号’ב00-1-5’的胚珠分别接种至MM3和ER基本培养基。对于‘Ruby Seedless’בJupiter’组合,接种至MM3培养基所获得的胚发育率和成苗率显著高于ER培养基(P<0.05)(表2);组合‘秦红2号’ב00-1-5’在MM3培养基中,胚的发育率和成苗率比ER培养基高,两种培养基中胚的发育率和成苗率之间差异显著(P<0.05)。Table 2
表2
表2胚发育培养基MM3与ER对胚挽救成苗的影响
Table 2
| 杂交组合 Crossing combination | 培养基类型 Types of medium | 胚珠数 No.of ovules cultured | 剥胚数 No. of embryos developed | 萌发数 No. of embryos germinated | 成苗数 No. of Normal seedings | 发育率 Proportion of Embryos developed (%) | 萌发率 Proportion of Embryos germinated (%) | 成苗率 Proportion of seedings (%) |
|---|---|---|---|---|---|---|---|---|
| Ruby Seedless × Jupiter | MM3 | 525 | 72 | 56 | 56 | 13.71a | 10.67a | 10.67a |
| Ruby Seedless × Jupiter | ER | 508 | 58 | 35 | 35 | 11.42b | 6.89b | 6.89b |
| 秦红2号×00-1-5 Qinhong No.2 × 00-1-5 | MM3 | 405 | 69 | 38 | 30 | 17.04a | 9.38b | 7.41a |
| 秦红2号×00-1-5 Qinhong No.2 × 00-1-5 | ER | 386 | 50 | 39 | 23 | 12.95b | 10.10a | 5.96b |
新窗口打开|下载CSV
2.3 不同母本及发育培养基对畸形苗率的影响
对5个杂交组合幼胚在MM3培养基中的畸形苗率的调查结果(表3)表明,畸形苗率最高的是‘秦红1号’בJupiter’,为5.96%,而组合‘Ruby Seedless’בJupiter’未出现畸形苗,其他3个组合介于1.17%—5.03%。Table 3
表3
表3母本对畸形苗形成的影响
Table 3
| 杂交组合 Crossing combination | 胚珠数 No. of ovules | 正常苗 Normal plantlet | 畸形苗 Abnormal plantlet | ||
|---|---|---|---|---|---|
| 数量 No. | 百分比Percentage (%) | 数量 No. | 百分比Percentage (%) | ||
| 秦秀×00-1-5 Qinxiu × 00-1-5 | 427 | 6 | 1.41c | 5 | 1.17bc |
| 秦红2号×00-1-5 Qinhong No.2 × 00-1-5 | 405 | 30 | 7.41b | 8 | 1.98cd |
| Crimson Seedless × Jupiter | 398 | 7 | 1.76d | 20 | 5.03a |
| Ruby Seedless × Jupiter | 525 | 56 | 10.67a | 0 | 0d |
| 秦红1号×Jupiter Qinhong No.1 × Jupiter | 403 | 19 | 4.71c | 24 | 5.96ab |
| 总计 Total | 2158 | 118 | 5.47 | 57 | 2.64 |
新窗口打开|下载CSV
不同发育培养基对不同杂交组合畸形苗率的影响结果见表4。对于‘秦红2号’ב00-1-5’组合,在ER培养基上获得的畸形苗率更高,显著高于MM3培养基,为4.15%。而‘Ruby Seedless’בJupiter’在MM3和ER培养基上均未出现畸形苗。
Table 4
表4
表4MM3和ER培养基对畸形苗率的影响
Table 4
| 杂交组合 Crossing combination | 培养基类型 Types of medium | 胚珠数 No. of ovules | 正常苗 Normal plantlets | 畸形苗 Abnormal plantlet | ||
|---|---|---|---|---|---|---|
| 数量 No. | 百分比Percentage (%) | 数量 No. | 百分比Percentage (%) | |||
| Ruby Seedless × Jupiter | MM3 | 525 | 56 | 10.67a | 0 | 0.00b |
| Ruby Seedless × Jupiter | ER | 508 | 35 | 6.89b | 0 | 0.00b |
| 秦红2号×00-1-5 Qinhong No.2 × 00-1-5 | MM3 | 405 | 30 | 7.41b | 8 | 1.98b |
| 秦红2号×00-1-5 Qinhong No.2 × 00-1-5 | ER | 386 | 23 | 5.96b | 16 | 4.15a |
新窗口打开|下载CSV
2.4 畸形苗转化培养基的筛选
‘秦红2号’ב00-1-5’组合的45株畸形苗转化结果(表5)表明,畸形苗转接至2MS+1.6 mg?L-1 ZnSO4+0.2 mg?L-1 6-BA+ 0.1 mg?L-1 IAA转化培养基上效果较好,转化率为40.00%,显著高于对照组。Table 5
表5
表5不同ZnSO4浓度对畸形苗转化率的影响
Table 5
| 培养基类型 Types of medium | 畸形苗数 No. of abnormal plantlet | 畸形苗 Abnormal plantlet | 转化的正常苗 Transformation normal plantlet | ||
|---|---|---|---|---|---|
| 数量 No. | 百分比Percentage (%) | 数量 No. | 百分比Percentage (%) | ||
| 2MS+0.2mg?L-1 6-BA+0.1mg?L-1 IAA | 15 | 15 | 100a | 0 | 0b |
| 2MS+1.0mg?L-1 ZnSO4+0.2mg?L-1 6-BA+0.1mg?L-1 IAA | 15 | 10 | 66.67b | 5 | 33.33a |
| 2MS+1.6mg?L-1 ZnSO4+0.2mg?L-1 6-BA+0.1mg?L-1 IAA | 15 | 9 | 60.00b | 6 | 40.00a |
新窗口打开|下载CSV
2.5 利用无核基因分子标记对杂交后代的无核性状鉴定
利用无核基因分子标记SCF27-2000对‘秦红2号’ב00-1-5’组合的53个杂种株系进行检测,表明51个株系扩增出特异条带(图2),该组合无核率为96.23%。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2无核基因分子标记SCF27-2000对‘秦红2号’ב00-1-5’组合亲本及后代的检测
M:Marker (2k Plus DNA marker) 1:秦红2号;2:00-1-5;3—55:‘秦红2号’ב00-1-5’组合杂交后代。“+”表示具有2000 bp特异带;“-”表示无2000 bp特异带。下同
Fig. 2Detection of seedless marker SCF27-2000 to the hybrids of combination ‘Qinhong No.2’ × ‘00-1-5’
M: Marker (2k Plus DNA marker); 1: Qinhong No.2; 2: 00-1-5; 3-55: hybrid seedlings from crosses between ‘Qinhong No.2’ × ‘00-1-5’. “+” indicates the 2000 bp specific band was present; “-” indicates the 2000 bp specific band was absent. The same as below
利用无核标记SCF27-2000对‘秦红1号’בJupiter’组合的19个杂种株系进行检测,所有株系均扩增出特异条带(图3),该组合无核率为100%。
图3
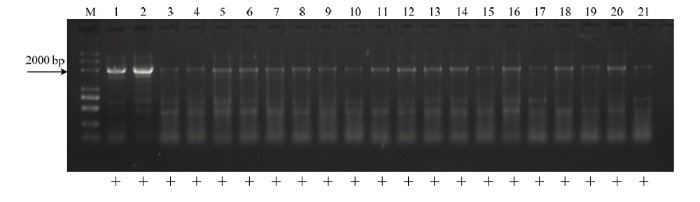 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3无核基因分子标记SCF27-2000对‘秦红1号’בJupiter’组合亲本及后代的检测
M:Marker (2k Plus DNA marker);1:秦红1号;2:Jupiter;3—21:‘秦红1号’בJupiter’组合杂交后代
Fig. 3Detection of seedless marker SCF27-2000 to the hybrids of combination ‘Qinhong No.1’ × ‘Jupiter’
M:Marker (2k Plus DNA marker); 1: Qinhong No.1; 2: Jupiter; 3-21: hybrid seedlings from crosses between ‘Qinhong No.1’ × ‘Jupiter’
利用无核基因分子标记SCF27-2000分别对‘秦秀’ב00-1-5’‘Ruby Seedless’בJupiter’和‘Crimson Seedless’בJupiter’组合的杂种株系进行检测,无核率分别为83.33%、90.59%和100%。
3 讨论
SAHIJRAM等[7]认为,利用传统育种方法对种子败育型无核葡萄作亲本进行杂交,很难获得无核杂种后代。而胚挽救技术能够解决这一问题,但影响胚挽救成功的因素很多,如亲本类型、激素处理、取样时期、培养基、培养条件等[9,24-26]。NOTSUKA等[5]研究认为,由于无核葡萄胚挽救中选用的母本不同,杂交亲和性不同,形成合子胚的比率不同,合子胚败育时间也不相同,用不同品种作母本进行杂交,胚的发育率不同。本试验结果表明,‘Ruby Seedless’‘秦红1号?’和‘秦红2号?’较适合作母本,这与LIU等[14]和VALDEZ[25]及笔者课题组[9]前期的结果相一致。而‘秦秀’和‘Crimson Seedless’作母本的胚挽救效果较差,但LIU等[14]和SIMIN等[26]认为‘Crimson Seedless’较适合作胚挽救母本,可能是由于每年物候期不同、取样时期不同和选用的培养基不同影响胚挽救的效果。不同母本由于基因型不同,胚珠内营养状况不同,受精卵发育程度不同,幼胚离体培养后进一步发育的程度也不同,造成部分幼胚不能萌发成正常植株。当‘Crimson Seedless’‘秦红1号’和‘秦秀’作母本时,畸形苗率较高,而‘Ruby Seedless’‘秦红2号’作母本时,畸形苗率较低。以上说明‘Ruby Seedless’‘秦红1号’和‘秦红2号’较适合作胚挽救母本。胚发育培养基为离体胚珠提供了幼胚体外发育所需的生长环境和营养物质,是影响胚挽救效果的关键因素之一。前人对胚发育培养基和萌发培养基进行了大量研究[5-7,11-13],目前应用较多的胚发育培养基主要有Bouquet and Davis(BD)培养基(1989)[27]、Nitsch and Nitsch(NN)培养基(1969)[28]、Cain培养基(1983)[12]、Emershad and Ramming(ER)培养基(1984)和MM3培养基等[29,30,31,32]。EBADI等[33]以‘Flame Seedless’自然授粉胚珠为研究对象,认为NN培养基比ER培养基效果更好。BURGER等[29]认为采用BD培养基并添加活性炭效果最好。PONCE等[34]认为MS培养基效果最好。KHOSHANDAM等[32]认为NN培养基适合作为胚发育培养基。赵雅楠等[35]认为MM3培养基效果比ER培养基效果好。本研究发现‘Ruby Seedless’בJupiter’接种在MM3培养基中,胚挽救效果最好,胚的成苗率最高,这与LIU等[14]、LI[24]等的结果相一致。MM3比ER培养基中多添加了半胱氨酸盐酸盐,半胱氨酸盐酸盐具有抗氧化和防止非酶褐变的作用,推测其可以减轻剥取胚珠时造成的损伤,促进胚的进一步发育,减少畸形苗的形成。
发育培养基选定后,在培养基中添加适量激素配比也会显著促进胚的发育。KUMAR等[30]认为植物激素在植物发育各个方面的调节中具有重要作用,因此被广泛研究。LI等[4]认为在WPM培养基中添加0.1 μmol?L-1 6-BA作胚发育培养基效果最好。RAZI等[13]认为IAA的浓度对胚萌发具有显著影响,‘Askari’בRuby Seedless’和‘Askari’בBidane Sefid’组合在NN培养基上添加1 mg?L-1 IAA效果最好。KHOSHANDAM等[32]认为在NN培养基中添加0.35 mg?L-1 GA3和1 mg?L-1 IAA效果最好。GRAY等[36]发现培养基中添加BA能显著提高胚萌发率。本研究表明,当以MM3为胚发育培养基时,除添加IAA浓度与RAZI等[13]和KHOSHANDAM等[32]相同外,还应添加0.5 mg?L-1 GA3和0.5 mg?L-1 6-BA,适宜作为抗寒无核葡萄胚挽救的胚发育培养基。KHOSHANDAM等[32]认为NN培养基也可以作为胚萌发培养基。POMMER等[12]用WPM液体培养基在滤纸桥上培养幼胚。EBADI等[33]也认为WPM培养基作为胚萌发培养基效果较好。
有研究表明,在胚挽救过程中,部分萌发的胚畸形生长,不能形成正常幼苗[25]。LESHEM等[37]认为培养基中添加激素后能促进和诱导畸形苗的产生,可能是由于生长素和细胞分裂素的比例不协调所造成。LIU等[38]认为胚挽救过程中畸形苗的出现可能与胚发育水平相关。JI等[39]认为2MS+0.4 mg?L-1 6-BA+0.1 mg?L-1 IBA+10 μmol?L-1 ZnSO4作转化正常植株的培养基,转化率最高,为27.9%,本研究与此结果类似。
胚挽救苗获得后,需要对其进行无核性状检测,保留具有目的性状的后代。田间鉴定方法准确有效,但等待结果的时间较长。分子标记辅助选择实现了对基因型的直接选择,是早期辅助选择的有效手段[17]。目前应用较多的葡萄无核基因分子标记有SCAR标记SCC8-1018、SCF27-2000和RAPD标记GSLP1- 569[9,14,35]。前期研究表明,SCF27-2000标记可以有效区分无核与有核亲本,被许多研究者使用[9,14,24],利用该分子标记鉴定的无核率与田间测定的实际无核率相关性达81%[19]。而SCC8-1018和GSLP1-569只能区分部分无核与有核品种,特别是GSLP1-569仅对‘无核白’及其亲缘关系较近的无核品种具有良好的适用性[9,14]。因此,本研究采用SCF27-2000对亲本及其杂交后代进行无核性状鉴定。由于分子标记与无核基因的连锁程度不同,连锁程度越高,检测无核性状的准确率越高,因此,筛选连锁紧密的无核基因分子标记仍然是本领域研究的重要内容。对于已获得的胚挽救后代,除了采用无核基因分子标记SCF27-2000进行早期鉴定外,仍需要与传统的田间鉴定相结合,以筛选出真正的无核杂种单株。
胚挽救杂种后代无核性状的遗传较为复杂,目前已有许多无核性状遗传的假说,包括单显性基因假说[40]、单隐性基因假说[41]、数量性状假说[42]、2隐性基因假说[43]、3互补显性基因假说[44]、1显性3隐性假说[45]与1主效基因3微效基因假说[46]等。由于葡萄无核机理尚未研究清楚,现有的假说都不够全面[8,47]。RAMMING等[15]认为无核品种×无核品种无核后代比率理论上可达100%,本研究中无核品种×无核品种杂交后代无核率在90.59%—100%,与RAMMING等[15]的假说基本一致。但是,无核品种×有核品种杂交后代无核率达到83.33%—96.23%,与笔者前期研究结果一致[9],但高于NOTSUKA等[5]、LIU等[14]和赵雅楠等[35]的结果,推测不同组合对无核性状的遗传力不同,还可能与细胞质遗传有关[48]。因此,选择无核性状传递力强的无核葡萄品种作为胚挽救的母本,有利于获得无核杂种。
4 结论
母本基因型对胚挽救效果有显著影响。以‘00-1-5’作父本时,‘秦红2号’作母本的胚挽救效果较好;以‘Jupiter’作为父本时,‘Ruby Seedless’作母本的胚挽救效果较好。以‘Ruby Seedless’和‘秦红2号’作母本时,杂种胚的成苗率较高且畸形苗率较低,说明‘Ruby Seedless’和‘秦红2号’适宜作胚挽救母本。‘秦红1号’作母本杂种胚的成苗率较高,但是畸形苗率也比较高;而‘Crimson Seedless’和‘秦秀’作母本时,杂种胚的成苗率低且畸形苗率较高,说明‘Crimson Seedless’和‘秦秀’不适合作母本。对于‘Ruby Seedless’בJupiter’和‘秦红2号’ב00-1-5’2个组合,在MM3固-液双相培养基上的成苗率均比ER固-液双相培养基高,畸形苗率比ER固-液双相培养基低,表明MM3比ER培养基更适合作为胚发育培养基。利用无核基因分子标记SCF27-2000对胚挽救后代进行无核性状早期鉴定,初步判定获得的159个胚挽救苗是潜在的无核株系。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1007/s13205-019-1978-zURLPMID:31815083 [本文引用: 4]

In this research, the binding of cellulolytic enzymes in Cellic((R)) CTec2 on six lignin isolates obtained from alkali (0.5, 1.0, and 1.5% NaOH at 121 degrees C for 30 min) and acid (1, 2, and 3% H2SO4 at 121 degrees C for 60 min) pretreated switchgrass was investigated. Briefly, the hydrolysis of cellobiose and Avicel with and without (control) lignin isolates was performed via CTec2 (5 and 10 FPU g(-1) carbohydrate) to determine whether the presence of lignin and binding of cellulolytic enzymes to the isolated lignin can affect the sugar production using three carbohydrate-lignin loadings, namely, 0.5:0.25, 0.5:0.5, and 0.5:1.0% (wv(-1)). Based on SDS-PAGE results, beta-glucosidase (BG) was significantly bound to all lignin isolates. Some enzymes in CTec2 presumed to be cellobiohydrolases, endo-1,4-beta-glucanases, and xylanase, were also observed to partially bind to the lignin isolates. Up to 0.97 g glucose g(-1) cellobiose was produced via hydrolysis (72 h and pH 4.8) with CTec2 (5 and 10 FPU g(-1) carbohydrate). Similarly, up to 0.23 and 0.46 g glucose g(-1) Avicel were produced via hydrolysis (72 h and pH 4.8) with 5 and 10 FPU g(-1) carbohydrate, respectively. Results indicated that the addition of lignin isolates during cellobiose and Avicel hydrolysis did not significantly (p > 0.05) reduce glucose production regardless of type and amount of lignin isolate. Hence, even though BG was significantly bound to lignin isolates, it could maintain its functionality as a biological catalyst in this study.
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 15]
[本文引用: 3]
[本文引用: 3]
[本文引用: 5]
[本文引用: 6]
[本文引用: 9]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17426772 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOI:10.1093/jxb/eru277URL [本文引用: 2]
[本文引用: 1]
[本文引用: 5]
[本文引用: 2]
URLPMID:12240561 [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
DOI:10.1093/oxfordjournals.aob.a087051URL [本文引用: 1]
DOI:10.1071/AR03053URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/oxfordjournals.jhered.a111419URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-002-0951-zURLPMID:12582493 [本文引用: 1]
Parental and consensus genetic maps of Vitis vinifera L. (2n = 38) were constructed using a F(1) progeny of 139 individuals from a cross between two partially seedless genotypes. The consensus map contained 301 markers [250 amplification fragment length polymorphisms (AFLPs), 44 simple sequence repeats (SSRs), three isozymes, two random amplified polymorphic DNAs (RAPDs), one sequence-characterized amplified region (SCAR), and one phenotypic marker, berry color] mapped onto 20 linkage groups, and covered 1,002 cM. The maternal map consisted of 157 markers covering 767 cM (22 groups). The paternal map consisted of 144 markers covering 816 cM (23 groups). Differences in recombination rates between these maps and another unpublished map are discussed. The major gene for berry color was mapped on both the paternal and consensus maps. Quantitative trait loci (QTLs) for several quantitative subtraits of seedlessness in 3 successive years were searched for, based on parental maps: berry weight, seed number, seed total fresh and dry weights, seed percent dry matter, and seed mean fresh and dry weights. QTLs with large effects (R(2) up to 51%) were detected for all traits and years at the same location on one linkage group, with some evidence for the existence of a second linked major QTL for some of them. For these major QTLs, differences in relative parental effects were observed between traits. Three QTLs with small effects (R(2) from 6% to 11%) were also found on three other linkage groups, for berry weight and seed number in a single year, and for seed dry matter in 2 different years.
[本文引用: 1]
URLPMID:7278311 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}