 ,, �쿭, ��־��, ʯ��˳, ����, ������,����ʡũҵ��ѧԺ�����о���,���� 110161
,, �쿭, ��־��, ʯ��˳, ����, ������,����ʡũҵ��ѧԺ�����о���,���� 110161Formation Regulating and Micro-Structure of Sorghum Starch with Different Types of Endosperm
KE FuLai,, ZHU Kai, LI ZhiHua, SHI YongShun, ZOU JianQiu, WANG YanQiu,Sorghum Research Institute, Liaoning Academy of Agricultural Sciences, Shenyang 110161ͨѶ����:
���α༭: ����
�ո�����:2019-07-31��������:2020-01-2�����������:2020-07-16
| ��������: |
Received:2019-07-31Accepted:2020-01-2Online:2020-07-16
����� About authors
�¸���,E-mail��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (5347KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
�¸���, �쿭, ��־��, ʯ��˳, ����, ������. ��ͬŴ�Ը�����������γ�����۹��̵�øѧ���ػ��Ƽ����ṹ�仯[J]. �й�ũҵ��ѧ, 2020, 53(14): 2774-2785 doi:10.3864/j.issn.0578-1752.2020.14.002
KE FuLai, ZHU Kai, LI ZhiHua, SHI YongShun, ZOU JianQiu, WANG YanQiu.
0 ����
���о����塿�������й���Ҫ�ĺ�������,��������Ҫ��������ƺͳ´�,���������ưƺͳ´����Ը���Ϊԭ����������ɡ����۵ĺ�������ֶ��ڰƺͳ´�Ʒ�ʺͲ���������Ҫ����[1],���,��ͬ�������͵��۽ṹ�������о���Ϊ��������Ʒ�ʺͲ��������ṩ���ۻ�������ǰ���о���չ������ɵ�[2]�Բ�ͬ���ۺ����ĸ���Ϊ�о�����,�������������γɹ�������ص��ۺϳ�ø�ĵ�������,��ΪADPG��GBSS��SBE��DBE���ԵIJ��켰��ͬ���öԸ������ۻ��۲��������Ҫ�����������á������[3]����ɨ��羵��ӫ��������������Ƭ�����Ը���ӱ����������������͵��������̬�����˹۲�,���ָ����������γɹ����д��ڲ�ͬ����������ķ������Ľṹ���������[4]���ù�ѧ�����͵����������ϵļ����۲��������ϸ������,�ص��о��˸�������ϸ���е�����ķ������,��Ϊ��������������ͨ����ʵ�����γɵġ������Ե�[5]��19����Ƹ���Ϊ����,�����Ƥ����Ƥ�����Ƥ�㣩��ȼ��������������ṹ�о�,��Ϊ��ͬ�������������������������HUBER��[6]ͨ���Ի�ѧ�ձ���ձ��õ������ʸߡ������Ậ���ߵĸ������۽���ɨ��羵�۲�,��������ؿ�����һЩ���ۿ��������пɹ������Ŀס�ʷ�����[7]��ʯ�����[8]�����µ�[9]�ڸ��������ס�ˮ���ϵ��о��������۽ṹ��Ӱ������Ʒ�ʵ���Ҫ���ء������о�����㡿��ˮ����С�������������,���ڸ��������е��ۺϳɻ��Ƶ��о�����,�Ե��۽ṹ������о�����,�����ǶԸ�����ͬ�������͵��۽ṹ�����ص��о��ʼ��������������Ĺؼ����⡿���о���Ŵ�͡��뾬��Ŵ�ͺ;���3�ֲ�ͬ���͵ĸ���Ʒ��Ϊ�Բ�,�о���ͬ���������ۻ��۹��������ø��ı仯,ͬʱ����ɨ��羵�����۲���������γɡ���С�Լ��ṹ�仯�ص�,��ʾ���ظ����������ۻ��ۼ���ͬ������ֱ��ʵ���������,Ϊ�ߵ��������Ǹ�֧�����۸���Ʒ�ֵ�ѡ���������ṩ���ۻ�����1 �����뷽��
1.1 ���Բ���
ѡ��3�ֲ�ͬŴ�ԵĴ����Ը���Ʒ�ָ�1������1��,����,Ŵ��Ʒ��Ϊ��ճ3�ţ���ճA-2/��ճR-2��,�뾬��Ŵ��Ʒ��Ϊ����19��7050A/0-01��,����Ʒ��Ϊ����10�ţ�7050A/9198����Table 1
��1
��1����Ʒ�������ĵ������
Table 1
| Ʒ�� Varieties | �ܵ��ۺ��� Total starch content (%) | ֧�����ۺ��� Amylopectin content (%) | ֱ�����ۺ��� Amylose content (%) | ֧������ռ�ܵ��۵ı���a Ratio of amylopectin to total starch (%) |
|---|---|---|---|---|
| ��ճ3�� Liaonian 3 | 72.3 | 70.1 | 2.2 | 97.0 |
| ����19 Liaoza19 | 70.3 | 62.2 | 8.1 | 88.5 |
| ����10 Liaoza10 | 74.3 | 58.6 | 15.7 | 78.9 |
�´��ڴ�|����CSV
1.2 ���鷽��
�������,ÿƷ��20��,�г�3m,�о�0.6m,3���ظ�,����������С�2016��5��8�ղ���,������ѡ��������һ�µ�ֲ��100����Ʊ��,����ȡ���ⶨ���ӿ�����ʼ,ÿ��ȡ��һ��,������7��42d,ÿ7��ȡ��һ��,ÿ��ȡ3��ֲ�����в����������,һ������Ʒ150��ɱ��30min,75����������;һ����������Һ��Ѹ���䶳,-80�汣��;һ���������ù̶�Һ�̶���,���þƾ���ˮ����,��ȫ����Ʒȡ���,ͳһ�ⶨ������,�����Կ��ʵ�[10]�����ⶨ���ն����������ǽ����ữø��uridine diphosphate glucose pyrophosphorylase,UDPG�������ն����������ǽ����ữø��adenosine diphosphate glucose pyrophosphorylase,ADPG���������Ե��ۺϳ�ø��soluble starch synthase,SSS���ͽ��̬���ۺϳ�ø��granule- bound starch synthase,GBSS����ø���ԡ�����NAKAMURA��[11]��NAKAMURA��[12]���Է�ï��[13]�����ⶨ���۷�֧ø��starch branching enzyme,SBE���͵���ȥ��֧ø��starch-debranching enzymes,DBE����ø���ԡ����Ϻ��������\������˾����ɨ��羵�ⶨ�������ṹ,����SPSS19.0�������ݷ�����2 ���
2.1 ��ͬ�������������ۻ��۹��������ø���Զ�̬�仯
������,������ʼ�ཬ���������ཬ������,UDPG��ADPGø�����ڷۺ�Ѹ������,�ڷۺ�21 d�ﵽ���ֵ,���UDPGø���������½�,ADPGø����Ѹ���½���ͼ1����3��Ʒ����Ƚ�,�ཬǰ�ڲ��첻��,�ཬ���ڲ�������,����Ϊ����10��>����19>��ճ3��,�ཬ����UDPGø���Բ����Դ�����ˮƽ,ADPGø���Բ��첻������ͼ1
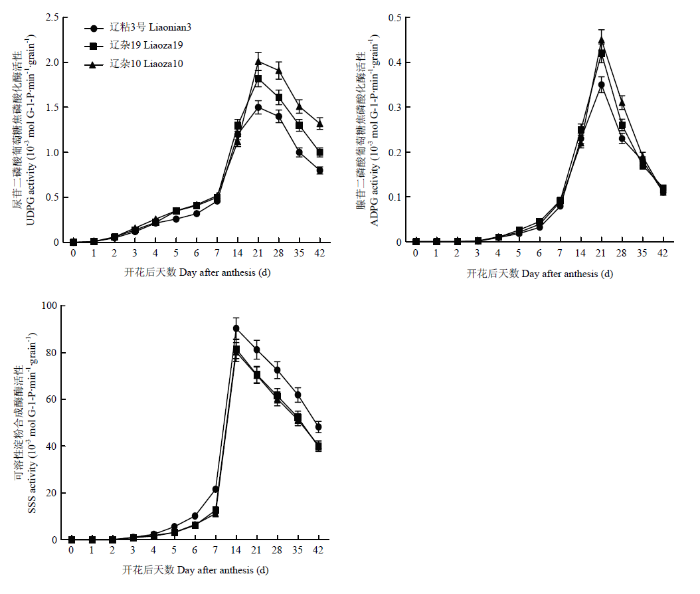 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1UDPG��ADPG��SSSø���Զ�̬�仯
Fig. 1Dynamics of activities of UDPG, ADPG and SSS in grains
�ڸ��������ཬ������,SSSø���Ա��ֺ�UDPG�������Ƶı仯����,������Ϊ�ڷۺ�Ѹ������,���������½�,��SSSø���Եķ�ֵ���ֽ���,�ڿ�����15 d������ߡ������ཬ������,��ճ3��SSSø����������������10�ź�����19,����10��������19����첻������
SBE��Ҫ������ˮ���-1,4���ռ�,�����µĶ���ͨ����-1,6���ռ����ӵ�������,�Ӷ��γɷ�֧����������ͬ������������SBE���Ծ��ʵ������߱仯,�����ڷۺ�21 d�ﵽ��ֵ�������ཬ�����ж�����Ϊ��ճ3��SBE����������������19������10��,����19������10��֮�����Ҳ������ˮƽ��ͼ2����
ͼ2
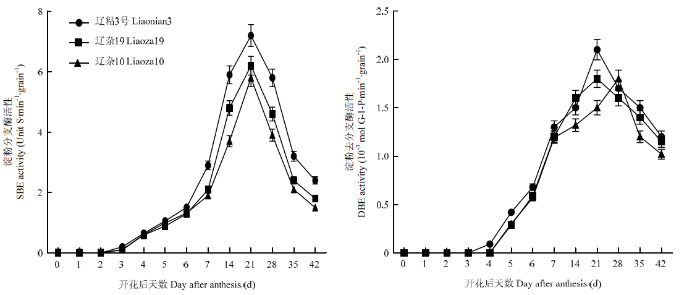 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2SBE��DBEø���Զ�̬�仯
Fig. 2Dynamics of activities of SBE, DBE in grains
DBE��Ҫ������ˮ��a-1,6-���ռ�,�Ե��۵����սṹ�������Ρ���ͼ2�ɼ�,��ͬ������������DBE���Եı仯Ҳ�ʵ�������,Ŵ����Ʒ����ճ3�ŷ�ֵ���,���ڷۺ�21 d�ﵽ��ֵ;�������Ͱ뾬��Ŵ����Ʒ�ֵķ�ֵ�ϵ�,���������ֵ��ʱ��Ҳ����,����10�����ڷۺ�28 d������DBE���Դﵽ��ֵ��ͼ2����
���������ཬ������,GBSSø���Բ����ϴ�,����10�š�����19���ֳ����Ե�˫�����߱仯,��ֵ�������ڷۺ�15��35 d,��ǰ�ڷ�ֵ�ϸ�;��ճ3�ű���Ϊ��������,�ڷۺ�Լ20 d�ﵽ��ֵ�������ཬ������,��ճ3��GBSSø����������������10�ź�����19;����10��������19�����Ҳ������ˮƽ,���ཬ������,������Ϊ����10��������������19��ͼ3����
ͼ3
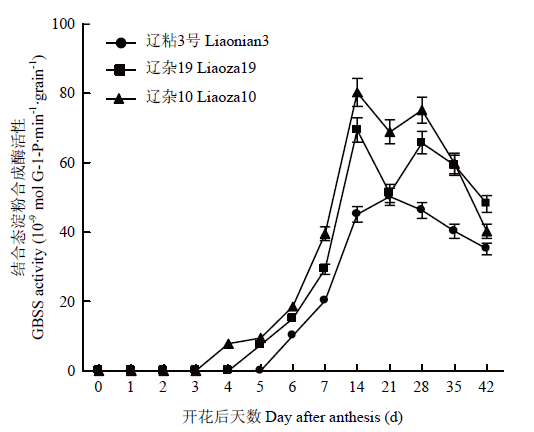 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3��ͬ����������GBSSø���Ա仯
Fig. 3Dynamics of activities of GBSS in grains
2.2 ���ۺϳ����ø��������ۻ������ʼ���ط���
Ϊ�˽�һ����ȷ���ۺϳ����ø��Ե��ۺϳɵĵ�������,�Բ�ͬ�ϳ�ø������ۻ������ʼ������Խ����˷������������2������,UDPG��ADPG��SSSø���ԺͲ�ͬ������ֱ�����ۡ�֧�����۵Ļ������ʾ�����������ء�GBSSø����������19������10��ֱ�����ۻ����������������,����ճ3��ֱ�����ۻ�����������Բ�����,�벻ͬƷ�ֵ�֧�����ۻ�����������Բ�������SBE��DBE�����벻ͬƷ�ֵ�֧�����ۻ����������������,��ֱ�����ۻ�����������Բ�������Table 2
��2
��2��ͬ�����������е��ۺϳ����ø��������ۻ������ʼ���ط���
Table 2
| ø Enzyme | ֱ�����ۻ�������Amylose accumulation rate | ֧�����ۻ�������Amylopectin accumulation rate | ||||
|---|---|---|---|---|---|---|
| ��ճ3 Liaonian3 | ����19 Liaoza19 | ����10 Liaoza10 | ��ճ3�� Liaonian3 | ����19 Liaoza19 | ����10 Liaoza10 | |
| ���ն����������ǽ����ữøUDPG | 0.85* | 0.82* | 0.89* | 0.79* | 0.84* | 0.86* |
| ���ն����������ǽ����ữøADPG | 0.87* | 0.89* | 0.84* | 0.92* | 0.85* | 0.94* |
| �����Ե��ۺϳ�øSSS | 0.79* | 0.76* | 0.74* | 0.91* | 0.94* | 0.92* |
| ���̬���ۺϳ�øGBSS | 0.64 | 0.72* | 0.75* | 0.59 | 0.49 | 0.36 |
| ���۷�֧øSBE | 0.47 | 0.58 | 0.49 | 0.98* | 0.95* | 0.86* |
| ����ȥ��֧øDBE | 0.42 | 0.51 | 0.38 | 0.78* | 0.75* | 0.73* |
�´��ڴ�|����CSV
2.3 ���۳��ṹ�����۲�
2.3.1 ����ϸ���е������ij�ʵ���� ����������10�ſ�����7 d,����ϸ�����Ѿ����ֵ���������,��ʱ,��������������,�����Բ�λ���Բ��,����ϸ�����Ѿ����ֵ������ϳ����ģ�ͼ4��,�ںϳ����ĵ������������С,�ɺϳ���������,�������ʷ����Է�ɢ,��Χ���������������������14 d,��������ϸ���е�������ʵ�϶�,�������Ļ������ʼӿ�,�����д�ϸ�����������۽��١�������21 d,���ۺϳ����ĵ�������������,���µ������Ļ���������������,������28 d,������ϸ���ж��ɹ۲쵽����������,����ϸ�����������Թ۲����������γ����ġ�������35 d,����ϸ���ж�����������,������֮����Թ۲���Եĵ����塣������42 d,��������ϸ���ж�����������,�������γ����ĵ�����Ҳ�����½���������,���ڵ������Ļ��༷ѹ,����ϸ���е������������ɽ�Բ�α�Ϊ������Ķ����,������֮������������塣ͼ4
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4���������е��������۹��̵����۲�
1������7 d;2������14 d;3������21 d;4������28 d;5������35 d;6������42 d;A��������;������������;B��������
Fig. 4Starch granule accumulation process
1: 7th after anthesis; 2: Day 14 after anthesis; 3: Day 21 after anthesis; 4: Day 28 after anthesis; 5: Day 35 after anthesis; 6: Day 42 after anthesis; A: Starch granule; ��: Starch granulation Center; B: Protein body
Ŵ������ճ3������ϸ���е������Ļ��۹���Ҳ�������Ƶı仯����,������7 d���۽���;������14��35 d�������Ļ��۽϶ࡣ�������������״Ҳ�ʽ�Բ��,�������ڵ������以�༷ѹ,Ҳ��Ϊ������Ķ���Ρ�������֮��Ҳ���������塣
2.3.2 ���������γ����� ����7 d������ϸ���пɹ۲�������ķֲ��Ϸ�ɢ,ֻ������������,��������ֱ�����ض���Ϊ����,�����ɴ�С�ı仯����,�����ĵ���ڵ�״�ʴصļ�����,����ǵ������γ����ĵij��Ρ���������ϸ���ij�ʵ����,�������γ�����������,��������35 d,���Թ۲���������ض���Ϊ����,�������Ե���״�ֲ�,��Ȧ��������ֱ����С,��Ȧ��������ֱ���ϴ���45 d,�γ����Ĵ��ĵ���������ѵ���ѹ�������Ե��α䣨ͼ5����
ͼ5
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5�������ϳ�����
1��35 d�۲�;2��7 d�۲�;3��42 d�۲⡣A��������;��������γ�����;C������
Fig. 5Starch granulation Center
1: Day 35 after anthesis; 2: day7 after anthesis; 3: Day 42 after anthesis. A: Starch granule; ��: Starch granulation center; C: Plastid
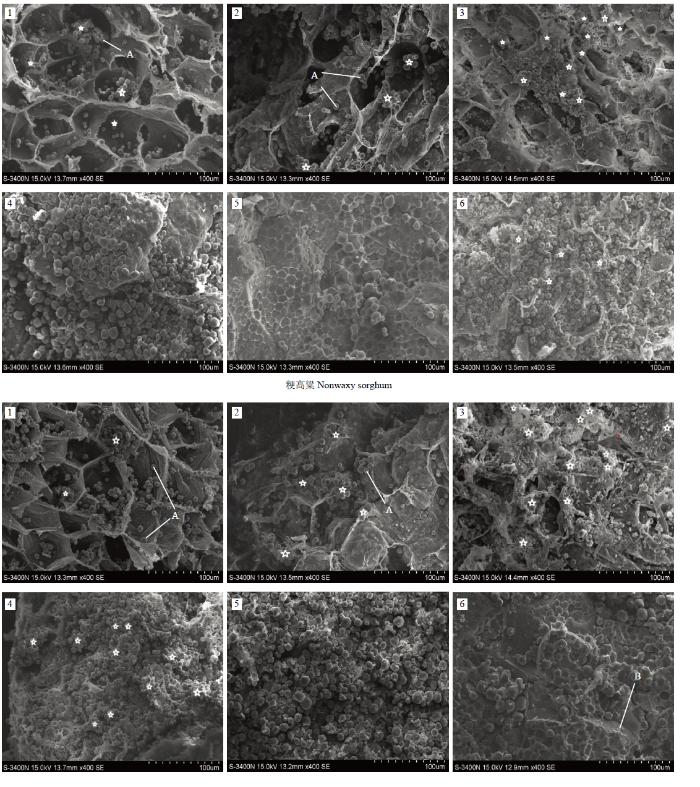
2.3.3 ����������̬���� ��ͬ����������������,����������̬��������ΪŴ�͵�������Խ�С,�־���10 ��m����,���氼�ӽ϶�;���͵�������Խϴ�,�־���10 ��m����,����������,����ȽϹ⻬;�뾬��Ŵ�͵��������ȷֲ��Ϲ�,��Ϊ������,���氼�ӽ��٣�ͼ6����
ͼ6
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6��ͬ��������������̬����
1��Ŵ����;2���뾬��Ŵ����;3��������
Fig. 6Morphological characteristics of starch granule in sorghum with different endosperm types
1: Glutinous sorghum; 2: Semi-glutinous sorghum; 3: Japonica sorghum
2.3.4 �������ڲ��ṹ���� ��ͬ���������������ڲ��ṹ����ϴ�,Ŵ���������������ڲ��ֲ��϶�Բ�ο�,ƽ��ֱ��Լ3.5 ��m;�������������ڲ��ȽϹ⻬,δ�����Կ�,�뾬��Ŵ�����������ڲ�������,�Ҿ��ʲ�������״,���������6��8 ��m��ͼ7����
ͼ7
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7�����������������ṹͼ
1��Ŵ����;2���뾬��Ŵ����;3��������
Fig. 7Submicroscopic diagram of transverse section of starch granule
1: Waxy sorghum; 2: Semi-waxy sorghum; 3: Nonwaxy sorghum
Ϊ����֤�����۲���,��������8����ͬ���͵�Ʒ��,���۲������ǵĵ������ڲ��ṹ���������������ڲ�ֻ�м�����ij���Բ�οն�,�ն���ֱ����С,���Ҳ����,�еĽ���С�İ��ӣ�ͼ8-c��ͼ8-e�����뾬��Ŵ�����������ڲ��ֲ��϶������Ρ�Ш�λ������εĿն���ͼ8-a��ͼ8-b����Ŵ�����������ڲ�Ҳ�ɹ۲�ն��Ĵ��ڣ�ͼ8-f��ͼ8-h��,�ն�����״��ΪШ�λ�Բ��,�ڸ���Ʒ�ֵĵ���������Ҳ�ɹ۲���ƿ��İߵ㣨ͼ8-h����
ͼ8
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ8�������ڲ���̬�ṹ���۲�
a��b���뾬��Ŵ���������������ṹͼ;c��d��e�������������������ṹͼ;f��g��h��Ŵ�͵���������ṹͼ
Fig. 8Microscopic observation on the internal structure of starch granule
a, b: Semi-glutinous sorghum; c, d, e: Japonica sorghum; f, g, h: Glutinous sorghum
3 ����
3.1 ��ͬ���������ۺϳɵĵ���
�������۵ĺϳ�����غϳ�ø��ĵ���,��Ҫ����UDPG��ADPG��SSS��GBSS��SBE��DBE�ȡ�UDPG��Ҫ�����γɵ��ۺϳɵ������ǹ���G-1-P��1-���������ǣ�,ADPG����G-1-P�ϳɵ��ۺϳɵ�ǰ��,���������ø����Ϊ�ǵ��ۺϳɵ�����ø[14,15,16,17,18]�����о�����,��ͬ������������UDPG��ADPG���Ա仯���ʵ�������,�ڿ�����21 dø�������,���ۺ����ϸߵ�����10��2��ø�Ļ����������ڵ��ۺ����ϵ͵�2��Ʒ��,��ط�������,��ͬ����Ʒ�ֵ�ֱ�����ۡ�֧�����۵ĺϳ���������������������,�����ڸ������ۺϳɹ�����,UDPG��ADPG��Ҫ���ص��ۺ���,��ֱ�����ۺ�֧�����۵ı�����û����������,����ǰ�˵��о��������һ��[14,15,16,17,18]�������о������ʾ,֧�����ۺ�����Ҫ��SSS��SBE��DBE 3��ø����,����SSS�����ڦ�-1,4���ռ�,������������ӳ�;SBE�����ڦ�-1,6���ռ�,����ϳ�֧��;DBE�����г���������֧��,����֧���������Ľṹ[19,20]����ӱ�ݵ�[21]��ʢ�[22]�����¸���[23]���Բ����[24]�о�����,SSS���Ժ�֧�����ۺϳ����������,GAO��[25]��ABEL��[26]����SSS����ͻ������о�Ҳ֤��,SSS���Ե��½�ʹ��֧�����۵ĺϳ��ܵ�����,��ֱ�����۵ĺ������ӡ����о���ͬ������SSS��SBE��DBE�����ֵ������߱仯����,SSS��ֵ���ֽ���,SBE��DBE��ֵ���ֽ���,����,������DBE��ֵ���ֵ�ʱ����������Ŵ��������ͬ������SSS��SBE��DBE���Բ�������,Ŵ����3��ø�Ļ���ʼ�ո��ھ�����,�������Ŵ����֧�����ۺ����ϸߵ���Ҫԭ����ط���Ҳ����,֧�����۵Ļ���������3��ø��������������ع�ϵ��
��ճ3�ſ�����5 d�ż�GBSS����,��������10��,���ֵ���ֵ�ʱ��Ҳ��������10��,���������ཬ������ø����ֵ��������������10��,���������ֱ�����ۺ����ϵ͵���Ҫ���ء�
ֱ�����ۺϳɹ�����,����ø�������Ҳ���ܺ��ӡ��Կ��ʵ�[10]��9����ֱͬ�����ۺ�����С����о�����,SSS�����ڹཬ������ֱ�����۵Ļ��۸���ء���ӱ�ݵ�[21]�о�С��ͬ���������������ۻ�����ø���Ե������,֤��SSS��SBE������ֱ�����۵ĺϳ�������������ء���������[27]Ҳ����,GBSS��SBE���Ժ�֧�����ۡ�ֱ�����ۻ������ʾ�����������ء����о�����,SSS�����벻ͬ����Ʒ�ֵ�ֱ�����ۺ�֧�����ۻ������ʾ������������,SBE��DBE��ֱ�����۵Ļ�������û�����������,��ǰ�˵�������������ͬ,Ʒ�����Բ�������������һ�������Ҫԭ��,��Ҳ��һ������˵��SSSø���ԶԵ��������ĵ������ÿ�����Ʒ�����Ե�Ӱ�졣����[28]��27�ݸ����������۵����о�Ҳ����,SSSø��������ۺϳɵĹ�ϵ,��Ʒ���Ŵ�����Ӱ�졣���о���,GBSS�����뾬�������뾬��Ŵ����ֱ�����ۻ����������������,��Ŵ����ֱ�����ۻ���������ز��������ݴ��Ʋ�GBSS��ֱ�����۵ĵ��������ܵ��ﺬ����Ӱ��,Ҳ������Ŵ���������нϸߵ�SBE��DBE����������GBSS������,����GBSS���Զ�ֱ�����۵ĵ������������ơ���骵�[29]���ֽϸߵ�SBE��������������ֱ�����۵��γɡ�
���ۺϳ����ø���������༼����ʩ��Ӱ��,�о�����,�����ס��ء���ˮ��Ӱ����ۺϳ�ø��Ļ���[30,31,32,33],���,�ڸ������������,�ɲ����ʵ����༼����ʩͨ��ø���Ա仯���ص��ۺϳɡ�
3.2 �����γɹ��̵����ṹ�仯
3.2.1 �������γ����� �����[3]��KS-304Ϊ�������,���õ羵�����۲������е����γɹ���,��Ϊ����35 d,�������Ѿ����ϸ�������о�����,����35 d,ϸ�����Կɹ۲�������γ�����,���е������Ļ��ۡ��������[4]ͬ����KS-304Ϊ�������,���õ羵�����۲������е��۵ķ�������,��Ϊ���������ڵ������ϳ������������γɵ�,���������γɻ�Ĥ�ṹ,Ȼ���γɵ�����,��������������γ���������ʷ���״���ӡ����о�Ҳ�۲�������γ����ĵ��γ�,�������Ĵ�С���γ������������ʽ����,����û�й۲�����γɵĻ�Ĥ�ṹ��3.2.2 �������γɹ�������ۻ��ۼ��ϵ ͨ��������,���Թ۲�������Ļ��۳̶�,������7 d�Ѿ��۲����������,������14 d����������������,������21��35 d���Թ۲������Ѹ������,��������ϸ��,������۵Ļ��۹���һ��,����ͨ�����������۹��̵����۲����ֱ�۵ķ�ӳ���۵Ļ������������ʢ�[34]���о����һ�¡�
3.3 �������ṹ�����۲�
3.3.1 �������ⲿ���������۲� ���Һ��[35]������ѩ��[36]��ʷ�����[7]����,��ͬ��Դ�������Ĵ�С����̬������ͬ��������[37]�Բ�ͬ��֧�������ۿ����������о�����,���������Ĵ�С�ܵ�����֧�ȵ�Ӱ�졣�ྲ��[38]�۲��˲�ͬŴ��С��������������̬,����Ŵ��С�������ƽ������С�ڷ�Ŵ�͡����о�����,��ͬ���������������������Ĵ�С��������,Ŵ��������������������С,�ձ���10 ��m����;�������������ϴ�,һ����15 ��m����;�뾬��Ŵ���������������ֲ��Ϲ㡣�����͵�������̬����,������Ϊ����������Ρ�3.3.2 �������ڲ����������۲� HUBER��[6]ͨ������ɨ�輤�������۲���������������ṹ,���ֵ��������������,��Щ�����������������,���ڵ��������ڲ��γɲ�ͬ��״�Ŀն������о�Ҳ���ֵ������ڲ����ڿն�,���ǿն���������״�������������,��������������û�з����пն��ķֲ�,Ŵ�����������Ŀն���Բ��,ֻ�а뾬��Ŵ�����ĵ������д������εĿն�����Ʒ����֤���Ҳ֤��,�������������ڲ��ն����ֵļ��ʺ�С,Ŵ�����������ڲ�����Բ�εĿն���,Ҳ�н϶�Ш�εĿն������о��Ʋ�ÿ����������ֱ����֧�����۵ı������ܲ�ͬ,�������ڲ��ṹ������ֱ��/֧�����۱�ֵ�������,ֱ������ռ�ݱ����ߵĵ������ڲ��γɿն��ļ��ʽ�С,֧��������ռ�����ϸߵĵ������ڲ��γɿն��ļ��ʽϸ�,���ڿն�����״����С�Ŀ���ԭ����Ҫ��һ�����о���ղ���ܵ�[39]�о�����,��ͬ���۽ṹ��������Ʋ�����������,Ŵ������ˮ���ʿ�,�����¶ȸ�,Ŵ�����������ڲ��϶�Ŀն������������һ�������Ҫԭ��
4 ����
UDPG��ADPG��SSS�ǵ��ظ����������ۻ������Ĺؼ�ø��SBE��DBE�ǵ���֧�����ۻ��۵Ĺؼ�ø��GBSS�ǵ���ֱ�����ۻ��۵Ĺؼ�ø��Ŵ������������С,�������������ϴ�Ŵ�����������ڲ����ڿն�,�������������ڲ����������ڿն�,�뾬��Ŵ�����������ڲ��ն���״�����������Ĵ�С���ڲ��ṹ��ֱ������/֧�����۵ı���������ء�
�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
//
[��������: 1]
//
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2014.17.002URL [��������: 2]

��Objective��The objective of this research was to clarify the relationship between endosperm and its maternal tissue during development process of the sorghum caryopsis. ��Method��Sorghum KS-304 was used as the experimental material with its caryopsis development days precisely recorded and its caryopsis development closely observed. Structural changes of endosperm cells and relation with their maternal tissue were observed through semi-thin sections under light microscopy by applying Spurr resin tissue embedding. The ultrastructure of cells from different tissues and the starch granules wherein in developing and full ripe caryopsis were observed under SEM. The relationships of testa and aleurone layer of full ripe caryopsis were studied through fluorescent microscopy using cryosectioning.��Result��Four stages could be found in caryopsis development, they were formation stage, milky stage, dough stage, and full maturity stage. Accordingly, endosperm development was divided into five stages, i.e. coenocyte stage, cellurization stage, differentiation stage, developmental stage and the final maturation stage, and the first three of which are equal to that of the formation stage of caryopsis; while the last maturation stage corresponds to the last two stages of caryopsis development. It took as long as 15 days for the nucellar epidermis to be fully degraded. The outer peripheral endosperm cells started to accumulate lipid bodies as early as 7 DPA (days after pollination) and, turned into aleurone cells in 11 DAP. Only 1 layer of aleurone cells formed in full ripe caryopsis. Besides commonly seen aleurone granules and globoids, some single amyloplasts were also found on ripe aleurone cells, circa. 3 μm in diameter. Growth and development of endosperm cells also varied according to different locations, amyloplasts in cells surrounding embryo tended to have a slow growth rate and were loosely packed than other areas, making them floury endosperm compared to others’ corneous endosperms when finally matured. Amyloplasts constitution in subaleurone cells were quite composite, in a manner that mingles both aleurone and starchy endosperm cells. Amyloplast formation in starchy endosperm was unique, which has an “occurrence center” during initiation. Starch “grow” inside a tube-like plastid, and when mature, falls apart, leaving the mature amyloplast with an uneven oval shape. Starch/amyloplasts in mesocarp may not be exhausted approaching mature stage, and on the contrary, there seems to be a secondary growth in both quantity and diameter.��Conclusion�� Development of starchy endosperm in sorghum KS-304 showed a similar pattern to the corn. Starch formation of amyloplasts in starchy endosperm followed a unique way and is independent of all other crop spiecies. Cells of mesocarp layer may act as an extra “sink” during later stages in caryopsis development.
DOI:10.3864/j.issn.0578-1752.2014.17.002URL [��������: 2]
��Objective��The objective of this research was to clarify the relationship between endosperm and its maternal tissue during development process of the sorghum caryopsis. ��Method��Sorghum KS-304 was used as the experimental material with its caryopsis development days precisely recorded and its caryopsis development closely observed. Structural changes of endosperm cells and relation with their maternal tissue were observed through semi-thin sections under light microscopy by applying Spurr resin tissue embedding. The ultrastructure of cells from different tissues and the starch granules wherein in developing and full ripe caryopsis were observed under SEM. The relationships of testa and aleurone layer of full ripe caryopsis were studied through fluorescent microscopy using cryosectioning.��Result��Four stages could be found in caryopsis development, they were formation stage, milky stage, dough stage, and full maturity stage. Accordingly, endosperm development was divided into five stages, i.e. coenocyte stage, cellurization stage, differentiation stage, developmental stage and the final maturation stage, and the first three of which are equal to that of the formation stage of caryopsis; while the last maturation stage corresponds to the last two stages of caryopsis development. It took as long as 15 days for the nucellar epidermis to be fully degraded. The outer peripheral endosperm cells started to accumulate lipid bodies as early as 7 DPA (days after pollination) and, turned into aleurone cells in 11 DAP. Only 1 layer of aleurone cells formed in full ripe caryopsis. Besides commonly seen aleurone granules and globoids, some single amyloplasts were also found on ripe aleurone cells, circa. 3 μm in diameter. Growth and development of endosperm cells also varied according to different locations, amyloplasts in cells surrounding embryo tended to have a slow growth rate and were loosely packed than other areas, making them floury endosperm compared to others’ corneous endosperms when finally matured. Amyloplasts constitution in subaleurone cells were quite composite, in a manner that mingles both aleurone and starchy endosperm cells. Amyloplast formation in starchy endosperm was unique, which has an “occurrence center” during initiation. Starch “grow” inside a tube-like plastid, and when mature, falls apart, leaving the mature amyloplast with an uneven oval shape. Starch/amyloplasts in mesocarp may not be exhausted approaching mature stage, and on the contrary, there seems to be a secondary growth in both quantity and diameter.��Conclusion�� Development of starchy endosperm in sorghum KS-304 showed a similar pattern to the corn. Starch formation of amyloplasts in starchy endosperm followed a unique way and is independent of all other crop spiecies. Cells of mesocarp layer may act as an extra “sink” during later stages in caryopsis development.
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1016/S0144-8617(99)00145-9URL [��������: 2]
DOI:10.3864/j.issn.0578-1752.2011.21.023URL [��������: 2]
��Objective�� An experiment was conducted to clarify the starch granule size distribution and the relationship between starch granule size and starch quality properties in storage roots of different types of sweetpotato cultivars. ��Method�� Six starchy sweetpotato cultivars, five edible sweetpotato cultivars, and one dual-purpose sweetpotato cultivar were planted at Taian experimental site in Shandong Province, with randomized block design in 2009-2010 growing season. The volume distribution and number distribution of starch granule, and their relationship to the content of total starch and its components in storage root were determined. ��Result��The starch granule in storage root was 0.39-55.14 µm in diameter, containing smaller type��diameter��3.36 µm��, mid-type��diameter 3.36-19.76 µm��and larger type��diameter��19.76 µm��granules. The distribution frequency showed typical two-peak curve in starch granule volume and number, and the grain diameters were all equal to 3.36 µm. The volume of smaller type granule accounted for 9.8%-18.5% of the total starch granule, and the number of smaller type granule accounted for 97.7%-99.1% of the total starch granule. The volume percentage of ��3.36 µm starch granule and the number percentage of ��1.00µm starch granule were higher in edible sweetpotato than in starchy sweetpotato. The contents of total starch , amylose and amylopectin were negatively correlated with the volume percentage of smaller type��r=-0.772**, -0.730**, -0.774**��and middling type��r=-0.723**, -0.715**, -0.712**�� starch granules, respectively, but positively correlated to larger type��r=0.746**, 0.729**, 0.738**�� starch granule. The ratio of amylopectin to amylose was positively correlated with the volume percentage of smaller type��r=0.716**��and middling type��r=0.740**�� starch granules, respectively, but negatively correlated to larger type��r=-0.745**�� starch granule. ��Conclusion��The rates of small starch grains in storage root of edible sweetpotato were higher, but the rates of big starch grains in storage root of starchy sweetpotato were higher.
DOI:10.3864/j.issn.0578-1752.2011.21.023URL [��������: 2]
��Objective�� An experiment was conducted to clarify the starch granule size distribution and the relationship between starch granule size and starch quality properties in storage roots of different types of sweetpotato cultivars. ��Method�� Six starchy sweetpotato cultivars, five edible sweetpotato cultivars, and one dual-purpose sweetpotato cultivar were planted at Taian experimental site in Shandong Province, with randomized block design in 2009-2010 growing season. The volume distribution and number distribution of starch granule, and their relationship to the content of total starch and its components in storage root were determined. ��Result��The starch granule in storage root was 0.39-55.14 µm in diameter, containing smaller type��diameter��3.36 µm��, mid-type��diameter 3.36-19.76 µm��and larger type��diameter��19.76 µm��granules. The distribution frequency showed typical two-peak curve in starch granule volume and number, and the grain diameters were all equal to 3.36 µm. The volume of smaller type granule accounted for 9.8%-18.5% of the total starch granule, and the number of smaller type granule accounted for 97.7%-99.1% of the total starch granule. The volume percentage of ��3.36 µm starch granule and the number percentage of ��1.00µm starch granule were higher in edible sweetpotato than in starchy sweetpotato. The contents of total starch , amylose and amylopectin were negatively correlated with the volume percentage of smaller type��r=-0.772**, -0.730**, -0.774**��and middling type��r=-0.723**, -0.715**, -0.712**�� starch granules, respectively, but positively correlated to larger type��r=0.746**, 0.729**, 0.738**�� starch granule. The ratio of amylopectin to amylose was positively correlated with the volume percentage of smaller type��r=0.716**��and middling type��r=0.740**�� starch granules, respectively, but negatively correlated to larger type��r=-0.745**�� starch granule. ��Conclusion��The rates of small starch grains in storage root of edible sweetpotato were higher, but the rates of big starch grains in storage root of starchy sweetpotato were higher.
DOI:10.3864/j.issn.0578-1752.2014.04.003URL [��������: 1]
��Objective��Starch granule size distribution is an important factor to evaluate starch quality and is greatly influenced by environmental factor. Irrigation and nitrogen are the major agronomic measures for high yield and good quality of maize. Therefore, it is very important for quality control and directional utilization of starch to clarify the effects of supplemental irrigation and nitrogen application on starch granule size distribution, and analyze the relationship between starch granule size distribution, grain weight, starch pasting properties and starch components of maize grain.��Method��In this study, Zhengdan 958 was used. Two water treatments were carried out, rain-fed (W1) and supplemental irrigation (W2). Supplemental irrigation treatment and irrigation amount were determined by soil water content. If soil water content was lower than 60% of field capacity, irrigation was applied. If soil water content after irrigation arrived 80% of field capacity, irrigation was stopped. Irrigation amount was calculated according to the formula of m=10ρbH(?i -?j), where m is the amount of supplementary irrigation (mm), H is the supplementary irrigation depth (cm) of the period, ρb is soil bulk density(g•cm-3) of the supplementary irrigation depth, βi is the objective water content (field capacity×objective and relative water content), βj is the natural water content. The highest nitrogen application rate was determined according to 3 kg nitrogen requirement for 100 kg kernels and the yield standard of 10 500 kg•hm-2 in 67 500 plants•hm-2 density. So, three nitrogen treatments were carried out, 0 (N1), 160 (N2) and 320 (N3) kg•hm-2. The volume, number and surface area distribution of starch granule in mature maize endosperm in irrigation and nitrogen application treatments were determined using laser particle size analyzer. Starch pasting properties were measured by RVA analyzer. Starch components were determined according to the double wavelengh method. The correlations between volume distribution of starch granule and starch pasting properties, kernel weight, yield and starch components were analyzed. ��Result�� Starch granule of maize was in the range of 0.38-39.78 μm, and the upper limit was 30.07-39.78 μm. Starch granule volume and surface area showed a three-peak curve. According to the concave point of the double peak curve, taking 3.5 μm and 7.4 μm as limit, starch granules were divided into three types: small (<3.5 μm), middle (3.5-7.4 μm) and large (>7.4 μm). Starch granule number showed a single peak curve. Starch granule of maize grain was mainly composed of small starch granule, which accounted for over 98% of total starch granule. Nitrogen fertilization and supplemental irrigation reduced the percentages of volume, surface area and number of small starch granule, and increased the volume percentage of large starch granule. Nitrogen fertilization and supplementary irrigation increased starch peak viscosity, trough viscosity, breakdown, final viscosity, setback, grain yield and weight, total starch and amylopectin content, reduced peak time, pasting temperature, amylose content and the ratio of amylose to amylopectin. Correlation analysis indicated that amylose content, the ratio of amylose to amylopectin, peak time and pasting temperature were positively correlated with volume percentage of small starch granule, and negatively correlated with volume percentage of large starch granule. Grain yield and weight, amylopectin and total starch content, peak viscosity, trough viscosity, breakdown, final viscosity and setback were negatively correlated with volume percentage of small starch granule, and positively correlated with volume percentage of large starch granule. ��Conclusion��Water and nitrogen fertilization significantly affected starch granule size distribution and pasting properties. Supplemental irrigation and 320 kg•hm-2 nitrogen fertilization treatment had the lowest volume percentage of small starch granule, the highest volume percentage of large starch granule and the best starch pasting properties.
DOI:10.3864/j.issn.0578-1752.2014.04.003URL [��������: 1]
��Objective��Starch granule size distribution is an important factor to evaluate starch quality and is greatly influenced by environmental factor. Irrigation and nitrogen are the major agronomic measures for high yield and good quality of maize. Therefore, it is very important for quality control and directional utilization of starch to clarify the effects of supplemental irrigation and nitrogen application on starch granule size distribution, and analyze the relationship between starch granule size distribution, grain weight, starch pasting properties and starch components of maize grain.��Method��In this study, Zhengdan 958 was used. Two water treatments were carried out, rain-fed (W1) and supplemental irrigation (W2). Supplemental irrigation treatment and irrigation amount were determined by soil water content. If soil water content was lower than 60% of field capacity, irrigation was applied. If soil water content after irrigation arrived 80% of field capacity, irrigation was stopped. Irrigation amount was calculated according to the formula of m=10ρbH(?i -?j), where m is the amount of supplementary irrigation (mm), H is the supplementary irrigation depth (cm) of the period, ρb is soil bulk density(g•cm-3) of the supplementary irrigation depth, βi is the objective water content (field capacity×objective and relative water content), βj is the natural water content. The highest nitrogen application rate was determined according to 3 kg nitrogen requirement for 100 kg kernels and the yield standard of 10 500 kg•hm-2 in 67 500 plants•hm-2 density. So, three nitrogen treatments were carried out, 0 (N1), 160 (N2) and 320 (N3) kg•hm-2. The volume, number and surface area distribution of starch granule in mature maize endosperm in irrigation and nitrogen application treatments were determined using laser particle size analyzer. Starch pasting properties were measured by RVA analyzer. Starch components were determined according to the double wavelengh method. The correlations between volume distribution of starch granule and starch pasting properties, kernel weight, yield and starch components were analyzed. ��Result�� Starch granule of maize was in the range of 0.38-39.78 μm, and the upper limit was 30.07-39.78 μm. Starch granule volume and surface area showed a three-peak curve. According to the concave point of the double peak curve, taking 3.5 μm and 7.4 μm as limit, starch granules were divided into three types: small (<3.5 μm), middle (3.5-7.4 μm) and large (>7.4 μm). Starch granule number showed a single peak curve. Starch granule of maize grain was mainly composed of small starch granule, which accounted for over 98% of total starch granule. Nitrogen fertilization and supplemental irrigation reduced the percentages of volume, surface area and number of small starch granule, and increased the volume percentage of large starch granule. Nitrogen fertilization and supplementary irrigation increased starch peak viscosity, trough viscosity, breakdown, final viscosity, setback, grain yield and weight, total starch and amylopectin content, reduced peak time, pasting temperature, amylose content and the ratio of amylose to amylopectin. Correlation analysis indicated that amylose content, the ratio of amylose to amylopectin, peak time and pasting temperature were positively correlated with volume percentage of small starch granule, and negatively correlated with volume percentage of large starch granule. Grain yield and weight, amylopectin and total starch content, peak viscosity, trough viscosity, breakdown, final viscosity and setback were negatively correlated with volume percentage of small starch granule, and positively correlated with volume percentage of large starch granule. ��Conclusion��Water and nitrogen fertilization significantly affected starch granule size distribution and pasting properties. Supplemental irrigation and 320 kg•hm-2 nitrogen fertilization treatment had the lowest volume percentage of small starch granule, the highest volume percentage of large starch granule and the best starch pasting properties.
DOI:10.3724/SP.J.1006.2019.82064URL [��������: 1]
Nowadays, the Wx b allele has been widely used to improve grain quality of indica rice. However, some indica varieties carrying Wx b allele usually has a much softer texture, which is not favored by consumers in South China. So the grain quality of these varieties needs to be further improved. To understand the effect of Wx in allele on rice eating quality and physicochemical properties in indica rice, we developed two Near-Isogenic Lines (NILs) carrying Wx in and Wx b alleles by crossing an indica variety 3611 (receptor, carrying Wx a) with IR64 (carrying Wx in) and 9311 (carrying Wx b), and seven times of backcrossing based on molecular marker assistant selection (MAS). The Wx effects in controlling the synthesis of amylose, grain quality, and physicochemical properties were investigated. There were non-significant differences in the agronomic traits among the NILs. However, for grain quality characters, we found that the NIL(Wx in) rice showed significantly lower apparent amylose content (AAC) and higher gel consistency (GC), compared with the wild type 3611. Besides, the NIL(Wx b) rice showed the lowest AAC and highest GC among three lines. The NIL(Wx in) rice had a significantly higher taste value than the wild type 3611, while the NIL(Wx b) rice exhibited the highest taste value among the three samples. The granule-bound starch synthase I (GBSSI) level was the highest in 3611, moderate in NIL(Wx in) and lowest in NIL(Wx b), which showed a positive correlation with the AAC level. Also, the starch viscosity, thermal gelatinization property and crystal structure of different rice flours had a high correlation with the AAC level. To sum up, our results proved that both Wx in and Wx b allele can improve the grain quality in 3611 background, and what is more, the Wx in allele might be more useful for the improvement of grain quality in indica rice.
DOI:10.3724/SP.J.1006.2019.82064URL [��������: 1]
Nowadays, the Wx b allele has been widely used to improve grain quality of indica rice. However, some indica varieties carrying Wx b allele usually has a much softer texture, which is not favored by consumers in South China. So the grain quality of these varieties needs to be further improved. To understand the effect of Wx in allele on rice eating quality and physicochemical properties in indica rice, we developed two Near-Isogenic Lines (NILs) carrying Wx in and Wx b alleles by crossing an indica variety 3611 (receptor, carrying Wx a) with IR64 (carrying Wx in) and 9311 (carrying Wx b), and seven times of backcrossing based on molecular marker assistant selection (MAS). The Wx effects in controlling the synthesis of amylose, grain quality, and physicochemical properties were investigated. There were non-significant differences in the agronomic traits among the NILs. However, for grain quality characters, we found that the NIL(Wx in) rice showed significantly lower apparent amylose content (AAC) and higher gel consistency (GC), compared with the wild type 3611. Besides, the NIL(Wx b) rice showed the lowest AAC and highest GC among three lines. The NIL(Wx in) rice had a significantly higher taste value than the wild type 3611, while the NIL(Wx b) rice exhibited the highest taste value among the three samples. The granule-bound starch synthase I (GBSSI) level was the highest in 3611, moderate in NIL(Wx in) and lowest in NIL(Wx b), which showed a positive correlation with the AAC level. Also, the starch viscosity, thermal gelatinization property and crystal structure of different rice flours had a high correlation with the AAC level. To sum up, our results proved that both Wx in and Wx b allele can improve the grain quality in 3611 background, and what is more, the Wx in allele might be more useful for the improvement of grain quality in indica rice.
URL [��������: 2]
Nine wheat cultivars with similar total starch content and different amylose and amylopectin content in mature stage were classified into three groups: low amylose, medium amylose and high amylose. The dynamic changes in ratio of amylose to amylopectin, and the activity changes of four enzymes in re
URL [��������: 2]
Nine wheat cultivars with similar total starch content and different amylose and amylopectin content in mature stage were classified into three groups: low amylose, medium amylose and high amylose. The dynamic changes in ratio of amylose to amylopectin, and the activity changes of four enzymes in re
[��������: 1]
DOI:10.1007/BF00196561URLPMID:8680309 [��������: 1]
Starch debranching enzyme (R-enzyme or pullulanase) was purified to homogeneity from developing endosperm of rice (Oryza sativa L. cv. Fujihikari) using a variety of high-performance liquid chromatography columns, and characterized. A cDNA clone encoding the full length of the rice endosperm debranching enzyme was isolated and its nucleotide sequence was determined. The cDNA contains an open reading frame of 2958 bp. The mature debranching enzyme of rice appears to be composed of 912 amino acids with a predicted relative molecular mass (Mr) of 102,069 Da, similar in size to its Mr of about 100,000 Da estimated by polyacrylamide gel electrophoresis in sodium dodecyl sulfate. The amino acid sequence of rice debranching enzyme is substantially similar to that of bacterial pullulanase, while it bears little similarity to that of bacterial isoamylase or to glycogen debranching enzymes from human muscle and rabbit muscle. Southern blot analyses strongly suggest that the debranching enzyme gene is present as a single copy in the rice genome. Analysis by restriction fragment length polymorphism with a probe including the 3'-untranslated region of cDNA for rice debranching enzyme confirmed that the debranching enzyme gene is located on chromosome 4.
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[D].
[��������: 1]
[D].
[��������: 1]
URL [��������: 1]
URL [��������: 1]
[��������: 1]
[��������: 1]
//
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.16819/j.1001-7216.2016.5099URL [��������: 1]
A field experiment was conducted to reveal the influence of cold��water stress(17��,lasting 3,6,9,12,15d) on grain starch synthesis and accumulation, yield components of japonica rice (Dongnong 428, Songjing 10 and Longdao 7) in cold��region during grain��filling stage, as well as the relationships between the changes in key enzymes activities related to starch synthesis and starch composition under cold��water stress. The results showed that compared with the control, soluble starch synthase (SSS) activity and starch branching enzyme (SBE) activity decreased, as well as the total starch contents and amylopectin content, while the amylose content increased under 17�� cold��water stress during grain��filling stage. According to the correlation analysis,the activities of soluble starch synthase (SSS) and starch branching enzyme (SBE) were significantly correlated with grain starch accumulation, playing an equally important role in the changes of starch and its component contents. Meanwhile,the grain number per panicle, thousand seed weight and seed setting rate were significantly reduced during grain��filling stage. The amplitude of variation for every indicator increased gradually with lengthening cold��water treatment time. The indicators related to the yield components of japonica rice in cold��region varied with cold��water stress duration and can be used for the identification of cold resistance. According to the indexes under the cold��water stress for different japonica rice varieties in cold��region, Songjing 10 was the most susceptible, followed by Longdao 7 and Dongnong 428.
DOI:10.16819/j.1001-7216.2016.5099URL [��������: 1]
A field experiment was conducted to reveal the influence of cold��water stress(17��,lasting 3,6,9,12,15d) on grain starch synthesis and accumulation, yield components of japonica rice (Dongnong 428, Songjing 10 and Longdao 7) in cold��region during grain��filling stage, as well as the relationships between the changes in key enzymes activities related to starch synthesis and starch composition under cold��water stress. The results showed that compared with the control, soluble starch synthase (SSS) activity and starch branching enzyme (SBE) activity decreased, as well as the total starch contents and amylopectin content, while the amylose content increased under 17�� cold��water stress during grain��filling stage. According to the correlation analysis,the activities of soluble starch synthase (SSS) and starch branching enzyme (SBE) were significantly correlated with grain starch accumulation, playing an equally important role in the changes of starch and its component contents. Meanwhile,the grain number per panicle, thousand seed weight and seed setting rate were significantly reduced during grain��filling stage. The amplitude of variation for every indicator increased gradually with lengthening cold��water treatment time. The indicators related to the yield components of japonica rice in cold��region varied with cold��water stress duration and can be used for the identification of cold resistance. According to the indexes under the cold��water stress for different japonica rice varieties in cold��region, Songjing 10 was the most susceptible, followed by Longdao 7 and Dongnong 428.
DOI:10.3864/j.issn.0578-1752.2019.22.020URL [��������: 1]
��Objective��Responses of sorghum grain yield , quality and NPK utilization to inherent soil fertility, and their interaction were explored to provide theoretical basis for best nutrient management according to soil fertility. ��Method��Pot experiment was conducted in greenhouse. Soil was collected from three treatments of long-term fertilizer application experiment for 6 years, which were without fertilizer, NPK application and NPK application along with animal manure and straw returning to field, respectively. They were termed as low soil fertility (LSF), medium soil fertility (MSF) and high soil fertility (HSF), correspondingly. For each soil fertility, there were two treatments for pot experiment: without fertilizer (NF) and fertilizer application (CF). Plant and grain of sorghum were harvested after maturity. NPK accumulation in plant above ground and grain were calculated according to NPK concentration and biomass of each part of plant. The contents of starch, tannic and protein of grain were determined. ��Result��For LSF, MSF and HSF, a similar biomass and yield were gained if fertilizer was applied. They were significantly affected by inherent soil fertility if fertilizer was withdrawn. Soil fertility did not affect starch content of grain, which was 67.99%-69.33%, if fertilizer was not applied. However, HSF combined with fertilizer application resulted in 60.75% starch content in grain. For all treatments, the highest of tannin content was observed in grain of LSF without fertilizer. Fertilizer application significantly decreased tannin content of grain when sorghum was cultivated with LSF and HSF, only 70% of that was produced by LSF combined with NF. Protein content was promoted with the increase of inherent soil fertility, and was doubled by fertilizer application whatever soil fertility was. When fertilizer was applied, the contributions of soil fertility to yield were 90.25%, 51.75% and 8.5% for LSF, MSF and HSF, respectively. NPK accumulation of both grain and plant aboveground was regulated by inherent soil fertility. Fertilizer enhanced NPK absorption for all soil fertility treatments. For example, N accumulation in grain and plant, induced by fertilizer application, were 1.8 and 1.9 times of those when fertilizer was withdrawn under conditions of HSL. NPK harvest indices were enhanced by fertilizer application if soil fertility was either low or medium; whereas diminished NPK harvest indices were noticed with high soil fertility.��Conclusion��The potential sorghum grain yield can be gained for low soil fertility by means of fertilizer application. Without fertilizer, inherent soil fertility has a significant effect on grain yield. However its impacts on content of starch, tannin and protein can be neglected. Low soil fertility, combined withdrawn fertilizer, promotes starch and tannin accumulation in grain. Fertilizer application has relative more influence on grain protein content than soil fertility does. Fertilizer application diminishes nutrient harvest index and nutrient use efficiency if inherent soil fertility is high.
DOI:10.3864/j.issn.0578-1752.2019.22.020URL [��������: 1]
��Objective��Responses of sorghum grain yield , quality and NPK utilization to inherent soil fertility, and their interaction were explored to provide theoretical basis for best nutrient management according to soil fertility. ��Method��Pot experiment was conducted in greenhouse. Soil was collected from three treatments of long-term fertilizer application experiment for 6 years, which were without fertilizer, NPK application and NPK application along with animal manure and straw returning to field, respectively. They were termed as low soil fertility (LSF), medium soil fertility (MSF) and high soil fertility (HSF), correspondingly. For each soil fertility, there were two treatments for pot experiment: without fertilizer (NF) and fertilizer application (CF). Plant and grain of sorghum were harvested after maturity. NPK accumulation in plant above ground and grain were calculated according to NPK concentration and biomass of each part of plant. The contents of starch, tannic and protein of grain were determined. ��Result��For LSF, MSF and HSF, a similar biomass and yield were gained if fertilizer was applied. They were significantly affected by inherent soil fertility if fertilizer was withdrawn. Soil fertility did not affect starch content of grain, which was 67.99%-69.33%, if fertilizer was not applied. However, HSF combined with fertilizer application resulted in 60.75% starch content in grain. For all treatments, the highest of tannin content was observed in grain of LSF without fertilizer. Fertilizer application significantly decreased tannin content of grain when sorghum was cultivated with LSF and HSF, only 70% of that was produced by LSF combined with NF. Protein content was promoted with the increase of inherent soil fertility, and was doubled by fertilizer application whatever soil fertility was. When fertilizer was applied, the contributions of soil fertility to yield were 90.25%, 51.75% and 8.5% for LSF, MSF and HSF, respectively. NPK accumulation of both grain and plant aboveground was regulated by inherent soil fertility. Fertilizer enhanced NPK absorption for all soil fertility treatments. For example, N accumulation in grain and plant, induced by fertilizer application, were 1.8 and 1.9 times of those when fertilizer was withdrawn under conditions of HSL. NPK harvest indices were enhanced by fertilizer application if soil fertility was either low or medium; whereas diminished NPK harvest indices were noticed with high soil fertility.��Conclusion��The potential sorghum grain yield can be gained for low soil fertility by means of fertilizer application. Without fertilizer, inherent soil fertility has a significant effect on grain yield. However its impacts on content of starch, tannin and protein can be neglected. Low soil fertility, combined withdrawn fertilizer, promotes starch and tannin accumulation in grain. Fertilizer application has relative more influence on grain protein content than soil fertility does. Fertilizer application diminishes nutrient harvest index and nutrient use efficiency if inherent soil fertility is high.
URL [��������: 1]
��Ŀ�ġ�����������ʩ�ʶ�С����ۺϳɹؼ�ø���Ե�Ӱ�졣���������ڴ������������£�������Ʒ�����Ͳ�ͬ�Ķ�С��Ʒ�֡�����9�š�������͡�����10�š����нΪ���ϣ��о�������ʩ�ʶ�С�������е��ۺϳ����ø���Ժ͵��ۺ�����Ӱ�켰���뿪������Ҷ������Ӫ���Ĺ�ϵ����������벻ʩ�����طʵĶ��մ�����ȣ�������ʩ����������˻��������е����Ǻ��������Ǻϳ�ø��SS���������Ե��ۺϳ�ø��SSS��������̬���ۺϳ�ø��GBSS���Ļ��ԣ�����˵��۲��������������ܵ��ۺ�֧�����ۺ�������С��Ʒ�ֵı��ֻ���һ�£����е��ʵ����ô��ڼطʣ�ʩ��������ֱ�����ۺ�����ʩ�������ֱ�����ۺ���������9�Ÿ�������10�š������ۡ���ط������������ط��������߿�������Ҷ������Ӫ����ά�����˵ĵ�/�رȣ��ǵ�����ʩ���������SSS��GBSS���ԣ������ٽ�С���������ۺϳɡ���������Ʒ�ʵ���Ҫ�������ء�ʩ�������С��Ʒ��GBSS������ֱ�����ۺ�����ߵ�øѧ�������ϸߵ�SS��SSS��GBSS����������Ʒ������9�Žϸߵ��ۺ�����øѧ���ϡ�
URL [��������: 1]
��Ŀ�ġ�����������ʩ�ʶ�С����ۺϳɹؼ�ø���Ե�Ӱ�졣���������ڴ������������£�������Ʒ�����Ͳ�ͬ�Ķ�С��Ʒ�֡�����9�š�������͡�����10�š����нΪ���ϣ��о�������ʩ�ʶ�С�������е��ۺϳ����ø���Ժ͵��ۺ�����Ӱ�켰���뿪������Ҷ������Ӫ���Ĺ�ϵ����������벻ʩ�����طʵĶ��մ�����ȣ�������ʩ����������˻��������е����Ǻ��������Ǻϳ�ø��SS���������Ե��ۺϳ�ø��SSS��������̬���ۺϳ�ø��GBSS���Ļ��ԣ�����˵��۲��������������ܵ��ۺ�֧�����ۺ�������С��Ʒ�ֵı��ֻ���һ�£����е��ʵ����ô��ڼطʣ�ʩ��������ֱ�����ۺ�����ʩ�������ֱ�����ۺ���������9�Ÿ�������10�š������ۡ���ط������������ط��������߿�������Ҷ������Ӫ����ά�����˵ĵ�/�رȣ��ǵ�����ʩ���������SSS��GBSS���ԣ������ٽ�С���������ۺϳɡ���������Ʒ�ʵ���Ҫ�������ء�ʩ�������С��Ʒ��GBSS������ֱ�����ۺ�����ߵ�øѧ�������ϸߵ�SS��SSS��GBSS����������Ʒ������9�Žϸߵ��ۺ�����øѧ���ϡ�
[��������: 1]
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2014.01.005URL [��������: 1]
��Objective��A field experiment was conducted to clarify the regulatory effects of potassium application time on starch accumulation and its physiological mechanism, starch granule distribution of storage roots of sweetpotato variety Jixu 23. ��Method��Jixu 23 (cultivar for starch use) was planted at Tai’an experimental site in Shandong Province, with randomized block design in 2008-2009 growing season. There were four treatments in all, including the control (non-potassium fertilizer, T1), potassium applied as basal fertilizer (T2), as top cover fertilizer (T3) and as top-growth peak fertilizer (T4). Samples were obtained by using periodic sampling during storage roots development. The content of starch, amylose and amylopectin of storage root were determined at different growth stages. Meanwhile the medium diameter of starch grain was determined and the volume fraction of different diameters starch granules (��19.76 μm, 3.359-19.76 μm and ��3.359 μm) was calculated. The activities of enzymes (SS, ADPGPPase, UDPGPPase, GBSS and SSS) related to starch synthesis were also studied. The effects of potassium application time on starch and its components, starch granule distribution and activities of enzymes related to starch synthesis in storage roots at different growth stages were analyzed.��Result��Compared with the control (T1), dry matter accumulation and starch yield of storage roots in treatments with potassium applied increased, and the increase of starch yield of T2, T3 and T4 treatments was 24.90%, 16.57%, and 9.24%, respectively. And the increase decreased when the application time delayed. The amylose content of storage root in treatments of topdressing (T3, T4) increased while amylopectin content was decreased significantly. The volume fraction of large starch granules in treatment with potassium applied as basal fertilizer (T2) was larger, and the volume fraction of medium and small starch granules in treatment with potassium applied as top-growth peak fertilizer (T4) was larger than that in the other treatments. Potassium application increased the activities of SS, ADPGPPase and UDPGPPase, and enhanced the supply capability of precursors for the synthesis of starch. And the longest action time of potassium on SS and ADPGPPase activities was 130 days, and potassium applied as basal fertilizer (T2) worked effectively mainly at early and middle stages of storage roots development, while treatments of topdressing (T3, T4) worked effectively mainly at middle and late stages of storage roots development. For UDPGPPase activity, the earlier the potassium applied, the longer action time was got. Potassium applied as topdressing (T3, T4) increased SSS activity but reduced GBSS activity of storage roots during the later expanding period. There was a significant positive correlation between total starch content and SS and ADPGPPase activities, and the significant positive correlation was also found between amylose content and GBSS and SS activities. And amylopectin content had a significant positive correlation with ADPGPPase and SSS activities.��Conclusion��Potassium applied as basal fertilizer (T2) and top cover fertilizer (T3) increased dry matter accumulation and starch yield of storage root significantly. Potassium applied as topdressing increased amylopectin content but decreased amylase content significantly. Treatments with potassium applied as basal fertilizer got a higher volume fraction of large starch granules, while treatments with potassium applied as top-growth peak fertilizer got a higher volume fraction of medium and small starch granules. The significant effects of potassium application time on amylase content, amylopectin content and starch yield were closely related with the changes of SS, ADPGPPase, UDPGPPase, GBSS and SSS activities.
DOI:10.3864/j.issn.0578-1752.2014.01.005URL [��������: 1]
��Objective��A field experiment was conducted to clarify the regulatory effects of potassium application time on starch accumulation and its physiological mechanism, starch granule distribution of storage roots of sweetpotato variety Jixu 23. ��Method��Jixu 23 (cultivar for starch use) was planted at Tai’an experimental site in Shandong Province, with randomized block design in 2008-2009 growing season. There were four treatments in all, including the control (non-potassium fertilizer, T1), potassium applied as basal fertilizer (T2), as top cover fertilizer (T3) and as top-growth peak fertilizer (T4). Samples were obtained by using periodic sampling during storage roots development. The content of starch, amylose and amylopectin of storage root were determined at different growth stages. Meanwhile the medium diameter of starch grain was determined and the volume fraction of different diameters starch granules (��19.76 μm, 3.359-19.76 μm and ��3.359 μm) was calculated. The activities of enzymes (SS, ADPGPPase, UDPGPPase, GBSS and SSS) related to starch synthesis were also studied. The effects of potassium application time on starch and its components, starch granule distribution and activities of enzymes related to starch synthesis in storage roots at different growth stages were analyzed.��Result��Compared with the control (T1), dry matter accumulation and starch yield of storage roots in treatments with potassium applied increased, and the increase of starch yield of T2, T3 and T4 treatments was 24.90%, 16.57%, and 9.24%, respectively. And the increase decreased when the application time delayed. The amylose content of storage root in treatments of topdressing (T3, T4) increased while amylopectin content was decreased significantly. The volume fraction of large starch granules in treatment with potassium applied as basal fertilizer (T2) was larger, and the volume fraction of medium and small starch granules in treatment with potassium applied as top-growth peak fertilizer (T4) was larger than that in the other treatments. Potassium application increased the activities of SS, ADPGPPase and UDPGPPase, and enhanced the supply capability of precursors for the synthesis of starch. And the longest action time of potassium on SS and ADPGPPase activities was 130 days, and potassium applied as basal fertilizer (T2) worked effectively mainly at early and middle stages of storage roots development, while treatments of topdressing (T3, T4) worked effectively mainly at middle and late stages of storage roots development. For UDPGPPase activity, the earlier the potassium applied, the longer action time was got. Potassium applied as topdressing (T3, T4) increased SSS activity but reduced GBSS activity of storage roots during the later expanding period. There was a significant positive correlation between total starch content and SS and ADPGPPase activities, and the significant positive correlation was also found between amylose content and GBSS and SS activities. And amylopectin content had a significant positive correlation with ADPGPPase and SSS activities.��Conclusion��Potassium applied as basal fertilizer (T2) and top cover fertilizer (T3) increased dry matter accumulation and starch yield of storage root significantly. Potassium applied as topdressing increased amylopectin content but decreased amylase content significantly. Treatments with potassium applied as basal fertilizer got a higher volume fraction of large starch granules, while treatments with potassium applied as top-growth peak fertilizer got a higher volume fraction of medium and small starch granules. The significant effects of potassium application time on amylase content, amylopectin content and starch yield were closely related with the changes of SS, ADPGPPase, UDPGPPase, GBSS and SSS activities.
URL [��������: 1]
URL [��������: 1]
DOI:10.3864/j.issn.0578-1752.2017.03.010URL [��������: 1]
��Objective��Sweetpotato (Ipomoea batatas (L.) Lam) is a typical K-preferred starchy root crop. Potassium (K) deficiency (namely low potassium), an important abiotic stress in plants,has become a major limiting element for obtaining crop high yield and good quality. However, in China, K deficiency in soils exists commonly in sweetpotato-growing areas. Sweetpotato starch utilization is closely related with its physico-chemical properties. The influence of low potassium on starch physic-chemical properties was investigated in an experiment conducted by using three sweetpotato cultivars with different potassium utilization efficiencies (KUE) in response to low potassium(K) field condition, aiming at providing reference for improving sweetpotato root properties and potassium utilization efficiency. ��Method��Three sweetpotato cultivars with different potassium utilization efficiencies (KUE) were chosen and cultivated under different potassium field condition, including two treatments (low potassium treatment, LK, and normal potassium treatment, CK).��Result��The results showed that under low K condition, the main components (amylose, amylopectin, nitrogen, total phosphorus and total lipid) in sweetpotato storage root starch changed to a certain extent. There were some differences among the three cultivars in the main components. Three distribution peaks in ≤2.50 μm, 2.50-5.00 μm and 5.00-30.00 μm were observed in starch granule size obviously. The slight decrease was observed in the whole average granule size under low K, which of Xushu32 had the least decrease in volume distribution. However, there was a significant difference in decrease of average granule size of Ningzishu1. The percentage, average granule size and peak value of different granule sizes in three distributions had some differences among three cultivars. Starch T0 (gelatinization temperature), Tp (peak temperature) and ΔH (heat enthalpy) were basicially lower than that in CK treatment, and Ningzishu1 had significant differences in the three characteristic values. Peak viscosity and breakdown viscosity in three genotypes had an obvious decrease. Hot paste viscosity, cool paste viscosity in Xushu18 and Ningzishu1 significantly increased, but Xushu32 was on the contrary. Moisture absorption degree of starches in three genotypes was reduced differently. There was a significant difference in the decline of moisture absorption in Xushu18 and Ningzishu1. In addition, swelling power of starch increased differently in three storage root starchs. There was a significant difference in a rise of swelling power of starch in Ningzishu1. Xushu32 had high starch swelling power, but low K had a little effect on it. Correlation analysis showed that the main physico-chemical indicators of sweetpotato starch are closely related. The low K changes starch compositions and particle sizes in sweetpotato root, which led to different degrees of influence on starch gelatinization properties and thermal properties.��Conclusion��The results of the study findings indicated that potassium is a key plant element influencing sweetpotato starch physico-chemical properties. Some obvious effects on physico-chemical properties and main components exist in storage root starch under low K stress. The responses of sweetpotato genotypes differing in potassium utilization efficiency to low K stress are different.
DOI:10.3864/j.issn.0578-1752.2017.03.010URL [��������: 1]
��Objective��Sweetpotato (Ipomoea batatas (L.) Lam) is a typical K-preferred starchy root crop. Potassium (K) deficiency (namely low potassium), an important abiotic stress in plants,has become a major limiting element for obtaining crop high yield and good quality. However, in China, K deficiency in soils exists commonly in sweetpotato-growing areas. Sweetpotato starch utilization is closely related with its physico-chemical properties. The influence of low potassium on starch physic-chemical properties was investigated in an experiment conducted by using three sweetpotato cultivars with different potassium utilization efficiencies (KUE) in response to low potassium(K) field condition, aiming at providing reference for improving sweetpotato root properties and potassium utilization efficiency. ��Method��Three sweetpotato cultivars with different potassium utilization efficiencies (KUE) were chosen and cultivated under different potassium field condition, including two treatments (low potassium treatment, LK, and normal potassium treatment, CK).��Result��The results showed that under low K condition, the main components (amylose, amylopectin, nitrogen, total phosphorus and total lipid) in sweetpotato storage root starch changed to a certain extent. There were some differences among the three cultivars in the main components. Three distribution peaks in ≤2.50 μm, 2.50-5.00 μm and 5.00-30.00 μm were observed in starch granule size obviously. The slight decrease was observed in the whole average granule size under low K, which of Xushu32 had the least decrease in volume distribution. However, there was a significant difference in decrease of average granule size of Ningzishu1. The percentage, average granule size and peak value of different granule sizes in three distributions had some differences among three cultivars. Starch T0 (gelatinization temperature), Tp (peak temperature) and ΔH (heat enthalpy) were basicially lower than that in CK treatment, and Ningzishu1 had significant differences in the three characteristic values. Peak viscosity and breakdown viscosity in three genotypes had an obvious decrease. Hot paste viscosity, cool paste viscosity in Xushu18 and Ningzishu1 significantly increased, but Xushu32 was on the contrary. Moisture absorption degree of starches in three genotypes was reduced differently. There was a significant difference in the decline of moisture absorption in Xushu18 and Ningzishu1. In addition, swelling power of starch increased differently in three storage root starchs. There was a significant difference in a rise of swelling power of starch in Ningzishu1. Xushu32 had high starch swelling power, but low K had a little effect on it. Correlation analysis showed that the main physico-chemical indicators of sweetpotato starch are closely related. The low K changes starch compositions and particle sizes in sweetpotato root, which led to different degrees of influence on starch gelatinization properties and thermal properties.��Conclusion��The results of the study findings indicated that potassium is a key plant element influencing sweetpotato starch physico-chemical properties. Some obvious effects on physico-chemical properties and main components exist in storage root starch under low K stress. The responses of sweetpotato genotypes differing in potassium utilization efficiency to low K stress are different.
[��������: 1]
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2014.22.006URL [��������: 1]
��Objective�� The aim of this research is to study the endosperm development and morphological features of starch in waxy wheat Shannuo 1 and non-waxy wheat Xinong 1330, which will provide a theoretical basis for quality improvement of wheat breeding. ��Method�� Two wheat cultivars, Shannuo 1 (waxy wheat) and Xinong 1330 (non-waxy wheat) were used as materials in this paper. The development of endosperm cells were observed by optical microscope. In order to preserve the endosperm structure, the developing grains (5, 8, 12, 15, 18, 21, 25 and 28 d after fertilization) were immersed in glutaraldehyde and osmic acid after transected by freezing in liquid nitrogen. Before embedded with Epon812, samples were washed with phosphate buffer and dehydrated individually in a graded acetone solution. The semi-thin slices were obtained by Leica ULTRACUT slicer. Images were taken by optical microscope after stained by 1% toluidine. The cross sections of Shannuo 1 and Xinong 1330 grains and starch samples which isolated from the two wheat varieties were placed on an aluminum specimen holder, and sputter-coated with a thin ?lm of gold under vacuum condition. Samples were observed with a JEOL scanning electron microscope (JSM-6360LV, JEOL, Japan). Particle size distribution of the starch samples was measured by MASTERSIZER-2000 laser particle analyzer (Malvern UK company). All measurements were performed in triplicates. Analysis of variance (ANOVA) was performed using LSD’s test to compare treatment variations at a signi?cance level of P<0.05 with DPS software. The data figures were performed by Sigmaplot 12.0 software. ��Result�� The results indicated that the endosperm cells of waxy wheat Shannuo 1 were smaller and developed slower than those of non-waxy wheat Xinong 1330 during grain development. In the early period of the endosperm development, the starch granules of Shannuo 1 and Xinong 1330 could be stained well by toluidine. However, in the late period of the grain development, the starch granules of Shannuo 1 could also be stained well, but Xinong 1330 can not. Compared with Xinong 1330, the endosperm of Shannuo 1 contained less protein matrix and the combination between protein and starch granules was much looser. The shape of B type starch granules of Shannuo 1 was irregular polygon, while that of Xinong 1330 was spherical. There was no significant difference in the shape of A type starch granules between Shannuo 1 and Xinong 1330. The size distribution of starch granules varied between waxy and non-waxy wheat. Volume distribution of Shannuo 1 starch granules showed a typical four-peak distribution, while that of Xinong 1330 showed a bimodal distribution. Granule surface area distribution of both cultivars indicated a typical three-peak distribution. The number distribution of granules in waxy and non-waxy starch was a typical unimodal distribution��The percentage of starch granules of Shannuo 1 and Xinong 1330 was nearly identical. In contrast, the percentage of the volume and the surface area of Shannuo 1 differed significantly from those of Xinong 1330. For A (>10 μm) type starch granules, the percentage of the volume and the surface area of Shannuo 1 were both lower than those of Xinong 1330. While for B (<10 μm) type starch granules, those of Shannuo 1 were higher than those of Xinong 1330. The volume, surface area and number of SB (<1 μm) type starch granules of Shannuo 1 were lower than those of Xinong 1330 by 1.11%, 11.60% and 9.28%, respectively, whereas the LB (1-10 μm) type starch granules of Shannuo 1 were higher than those of Xinong 1330 by 8.27%, 15.88% and 9.27%, respectively. A minority LA (>53 μm) type starch granules was found in Shannuo 1, but not in Xinong 1330. ��Conclusion�� In conclusion, the development of endosperm and the morphological features of starch granules of waxy wheat Shannuo 1 were significantly different from those of non-waxy wheat Xinong 1330 during grain development. The starch granules of LB type had considerable effects on the B type starch granules size distribution of Shannuo 1 and Xinong 1330.
DOI:10.3864/j.issn.0578-1752.2014.22.006URL [��������: 1]
��Objective�� The aim of this research is to study the endosperm development and morphological features of starch in waxy wheat Shannuo 1 and non-waxy wheat Xinong 1330, which will provide a theoretical basis for quality improvement of wheat breeding. ��Method�� Two wheat cultivars, Shannuo 1 (waxy wheat) and Xinong 1330 (non-waxy wheat) were used as materials in this paper. The development of endosperm cells were observed by optical microscope. In order to preserve the endosperm structure, the developing grains (5, 8, 12, 15, 18, 21, 25 and 28 d after fertilization) were immersed in glutaraldehyde and osmic acid after transected by freezing in liquid nitrogen. Before embedded with Epon812, samples were washed with phosphate buffer and dehydrated individually in a graded acetone solution. The semi-thin slices were obtained by Leica ULTRACUT slicer. Images were taken by optical microscope after stained by 1% toluidine. The cross sections of Shannuo 1 and Xinong 1330 grains and starch samples which isolated from the two wheat varieties were placed on an aluminum specimen holder, and sputter-coated with a thin ?lm of gold under vacuum condition. Samples were observed with a JEOL scanning electron microscope (JSM-6360LV, JEOL, Japan). Particle size distribution of the starch samples was measured by MASTERSIZER-2000 laser particle analyzer (Malvern UK company). All measurements were performed in triplicates. Analysis of variance (ANOVA) was performed using LSD’s test to compare treatment variations at a signi?cance level of P<0.05 with DPS software. The data figures were performed by Sigmaplot 12.0 software. ��Result�� The results indicated that the endosperm cells of waxy wheat Shannuo 1 were smaller and developed slower than those of non-waxy wheat Xinong 1330 during grain development. In the early period of the endosperm development, the starch granules of Shannuo 1 and Xinong 1330 could be stained well by toluidine. However, in the late period of the grain development, the starch granules of Shannuo 1 could also be stained well, but Xinong 1330 can not. Compared with Xinong 1330, the endosperm of Shannuo 1 contained less protein matrix and the combination between protein and starch granules was much looser. The shape of B type starch granules of Shannuo 1 was irregular polygon, while that of Xinong 1330 was spherical. There was no significant difference in the shape of A type starch granules between Shannuo 1 and Xinong 1330. The size distribution of starch granules varied between waxy and non-waxy wheat. Volume distribution of Shannuo 1 starch granules showed a typical four-peak distribution, while that of Xinong 1330 showed a bimodal distribution. Granule surface area distribution of both cultivars indicated a typical three-peak distribution. The number distribution of granules in waxy and non-waxy starch was a typical unimodal distribution��The percentage of starch granules of Shannuo 1 and Xinong 1330 was nearly identical. In contrast, the percentage of the volume and the surface area of Shannuo 1 differed significantly from those of Xinong 1330. For A (>10 μm) type starch granules, the percentage of the volume and the surface area of Shannuo 1 were both lower than those of Xinong 1330. While for B (<10 μm) type starch granules, those of Shannuo 1 were higher than those of Xinong 1330. The volume, surface area and number of SB (<1 μm) type starch granules of Shannuo 1 were lower than those of Xinong 1330 by 1.11%, 11.60% and 9.28%, respectively, whereas the LB (1-10 μm) type starch granules of Shannuo 1 were higher than those of Xinong 1330 by 8.27%, 15.88% and 9.27%, respectively. A minority LA (>53 μm) type starch granules was found in Shannuo 1, but not in Xinong 1330. ��Conclusion�� In conclusion, the development of endosperm and the morphological features of starch granules of waxy wheat Shannuo 1 were significantly different from those of non-waxy wheat Xinong 1330 during grain development. The starch granules of LB type had considerable effects on the B type starch granules size distribution of Shannuo 1 and Xinong 1330.
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}