 ,1
,1RNA-Binding Motif Protein 3(RBM3) Expression is Regulated by Insulin-Like Growth Factor (IGF-1) for Protecting Yak (Bos grunniens) Cumulus Cells from Apoptosis During Hypothermia Stress
PAN YangYang1, WANG Meng1, RUI Xian2, WANG LiBin1, HE HongHong1, WANG JingLei1, MA Rui1, XU GengQuan1, CUI Yan1, FAN JiangFeng1, YU SiJiu,1通讯作者:
责任编辑: 林鉴非
收稿日期:2019-05-5接受日期:2019-10-28网络出版日期:2020-06-01
| 基金资助: |
Received:2019-05-5Accepted:2019-10-28Online:2020-06-01
作者简介 About authors
潘阳阳,E-mail: panyangyang_2007@126.com。

摘要
关键词:
Abstract
Keywords:
PDF (2797KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
潘阳阳, 王萌, 芮弦, 王立斌, 何翃闳, 王靖雷, 马睿, 徐庚全, 崔燕, 樊江峰, 余四九. IGF-1调控RBM3表达抑制低温应激诱导牦牛卵丘细胞凋亡[J]. 中国农业科学, 2020, 53(11): 2285-2296 doi:10.3864/j.issn.0578-1752.2020.11.014
PAN YangYang, WANG Meng, RUI Xian, WANG LiBin, HE HongHong, WANG JingLei, MA Rui, XU GengQuan, CUI Yan, FAN JiangFeng, YU SiJiu.
0 引言
【研究意义】RNA结合基序蛋白3(RNA-binding motif protein 3,RBM3)作为冷诱导结合蛋白(cold- inducible RNA-binding protein,CIRP)家族成员之一[1],动物机体和细胞受到低氧、低温应激均可诱导其表达[1,2],通过促进体内其他蛋白的合成,维护应激条件下细胞的正常生理功能。研究哺乳动物细胞,尤其是高寒环境哺乳动物细胞经受低温应激时RBM3的表达变化,有助于揭示其参与动物机体低温适应性的调控机制。通过生长因子的作用,探索其对细胞和机体RBM3表达的影响,降低低温应激对细胞的损伤,对为优化动物体细胞和生殖细胞的冷冻技术具有重要意义。【前人研究进展】动物机体内RBM3可维持AU含量较高mRNAs的稳定性[3],通过诱导糖原合成激酶GSK-3β/ Wnt/β、Mark、丝氨酸/苏氨酸激酶等信号通路维持其在细胞中的多能性,如细胞周期参与从G2到M期过渡的调节,胞内相关microRNAs的调节[3,4]。哺乳动物的研究证实,RBM3可在多种细胞内表达,但表达水平存在差异,表达水平增加时可参与细胞内mRNAs翻译和翻译后的修饰的调节,进而提升细胞对不利环境刺激的耐受能力[5,6],如细胞对低温的耐受能力[7,8]。相关研究通过RBM3抑制剂咖啡酸苯乙酯(caffeic acid phenethyl ester,CAPE)的作用证实其参与NF-κB p65信号调节,保护HeLa细胞免受中低温诱导的细胞凋亡[9]。胰岛素样生长因子(insulin-like growth factor,IGF-1)作为细胞生长调控的关键因子,体外可通过多种途径调节细胞生长与增殖,调控哺乳动物卵母细胞的成熟和早期胚胎发育[10,11]。研究发现IGF-1通过调控B-细胞淋巴瘤/白血病-2原癌基因(B-cell lymphoma/leukemia-2,Bcl-2)和B细胞淋巴瘤/白血病基因伴随蛋白x(B-cell lymphoma/leukemia associated x protein,Bax)的表达水平抑制多种类型细胞凋亡[12,13]。【本研究切入点】笔者前期研究显示IGF-1通过调控牦牛卵丘细胞HSP70、Bax和 Bcl-2表达,降低细胞凋亡率[14]。在牦牛卵母细胞体外成熟过程中加入不同浓度IGF-1,可增加成熟卵母细胞CIRP的表达,提升其冷冻-解冻后的发育能力[15],但IGF-1提高卵母细胞冷冻后发育能力与细胞凋亡是否存在相关性未见报道。牦牛(Bos grunniens)是我国青藏高原区域的典型家畜,长期生活在年均气温为0℃的坏境,低温相关蛋白对其生理调控具有重要作用[16]。【拟解决的关键问题】因此本研究以牦牛卵丘细胞为模型,分析IGF-1和温度应激对RBM3表达的影响,评估RBM3的变化对细胞凋亡的调控,探索牦牛适应低温调控的分子机制,为进一步阐明RBM3参与动物机体或细胞免受低温损伤的分子机制提供了关键信息。1 材料与方法
1.1 主要试剂
DMEM/F12培养粉、细胞培养级别犊牛血清、青霉素、链霉素购自于美国Gibco公司;透明质酸酶、胰蛋白酶、IGF-1、RBM3抑制剂CAPE购自美国 Sigma公司。Bax(ab32503)和Bcl-2(ab117115)抗体购自美国Abcam公司,RBM3(bs-5902R)抗体、荧光二抗bs-0295G-FITC)和普通二抗(bs-0294D、bs-0295G)购自北京博奥森生物公司。细胞总RNA提取试剂盒、反转录试剂盒、SYBR GreenⅡ荧光定量PCR试剂盒均购自美国OMEGA生物公司,DAPI及蛋白免疫印迹(Western blot,WB)所用试剂均购买于北京索莱宝生物公司,其他试剂均为国产分析纯。1.2 牦牛卵丘细胞分离培养
2018年10月至2019年9月,在青海西宁及甘肃临夏家畜屠宰场,牦牛屠宰后立即采集卵巢,置于30—35℃预热的含有双抗的生理盐水中,4 h内带回甘肃农业大学牛羊胚胎工程中心。取出卵巢,用37℃生理盐水清洗2—3次,用带有12号针头的注射器从直径约为8 mm的卵泡中抽取卵泡液。将其转移到培养皿中,置于体式显微镜下用捡卵针挑取胞质均一,含有3层以上卵丘细胞的卵丘卵母细胞复合体(cumulus- oocyte complex,COC),无血清的PBS清洗3次后,用1%的透明质酸酶37℃消化5 min,体式显微镜下挑出卵母细胞,将含有卵丘细胞得悬液1 000 r/min离心5 min,用含有10% 血清的培养液通过离心再洗涤2次,悬浮细胞,计数后在37℃、5% CO2的条件下进行培养,每48 h换液一次。培养液为:DMEM/F12培养基+10%犊牛血清+100 U·mL-1链霉素+100 U·mL-1青霉素。1.3 IGF-1和低温应激分别处理卵丘细胞
卵丘细胞培养至第2代对数生长期时,加入含有0.05%的胰蛋白酶进行消化,在培养液中调整细胞数至4×105个/mL,传代到6孔板,每孔含有2 mL细胞悬液,培养24h后根据试验设计做以下处理(1)IGF-1及CAPE处理组:对其进行换液,分别加入0、50、100、200 ng·mL-1 IGF-1、25 μg·mL-1 CAPE和100 ng·mL-1 IGF-1+25 μg·mL-1 CAPE 到细胞培养液,每个处理组24个重复,培养30 h后,收集部分细胞用于RBM3 mRNA和蛋白的检测;(2)依据(1)的结果,取0和100 ng·mL-1处理组各6个重复分别置于30℃和25℃条件下应激8 h,收集细胞,并以37℃为对照组,进行后续RBM3 mRNA和蛋白检测实验,并分析IGF-1作用对卵丘细胞低温应激RBM3表达的影响;(3)检测(1)中,0、100 ng·mL-1 IGF-1和100 ng·mL-1 IGF-1+25 μg·mL-1 CAPE处理组卵丘细胞,25℃条件下应激8 h的后Bax和Bcl-2的表达水平。IGF-1处理卵丘细胞时间及低温应激时间根据预实验确定。1.4 不同处理组卵丘细胞凋亡率的检测
将细胞培养板中0、100 ng·mL-1 IGF-1和100 ng·mL-1 IGF-1+25 μg·mL-1 CAPE处理组细胞,用无血清PBS清洗3遍后,37℃,用0.05%胰蛋白酶消化3—5 min,加入含血清培养液终止消化,温柔吹打使细胞悬浮,移入1.5 mL 离心管中。经 1 000 r/min 离心 10 min,用PBS重悬细胞,并使其浓度达到1×107个/mL。用 1.5 mL 细胞固定液 4℃固定卵丘细胞 30 min,离心清洗固定液,参照 Annexin VFITC 细胞凋亡检测试剂盒说明标记细胞。用流式细胞仪以每 10 000 个细胞为单位,分析细胞凋亡率。1.5 RBM3、Bax、Bcl-2 mRNA表达检测
1.5.1 细胞总RNA提取与cDNA合成 每个试验组3个重复,将作用后卵丘细胞用PBS清洗3遍,参照贴壁细胞总RNA提取试剂盒说明书提取细胞总RNA,并参照反转录试剂盒合成第一链cDNA,置于-20℃保存、备用。1.5.2 Real-time PCR检测RBM3、Bax、Bcl-2的表达 根据GenBank公布的牦牛的RBM3 mRNA序列设计RBM3检测引物,Bax和Bcl-2 mRNA检测引物参照笔者前期研究所用的引物序列[13, 17],具体信息及反应条件见表1。反应体系主要包含1 μL cDNA模板,终浓度为0.25 μmol·mL-1的上下游引物各0.8 μL,2×SYBR GreenⅡ PCR mix 10 μL,加ddH2O使终体积达到20 μL。反应条件:94℃预变性15 s,94℃预变性10 s,退火10 s(具体温度见表1)、72℃延伸10 s,共35个循环,所有荧光定量PCR仪为Roche 480。内参基因为β-actin,重复至少3次。采用相对定量分析法评估基因表达水平[18]。
Table 1
表1
表1Real-time PCR引物信息
Table 1
| 基因 Gene | 引物序列 Primer sequence | 温度 Tm (℃) | 产物大小 Product size (bp) | GenBank 登录号 GenBank accession No. |
|---|---|---|---|---|
| RBM3 | F:GCTTCATCACCTTCACCAATC R:GGTATGACAATCGACCTGGAGC | 60 | 220 | MF142258.1 |
| Bax | F:TTTGCTTCAGGGTTTCATC R:CAGCTGCGATCATCCTCT | 59 | 174 | NM173894.1 |
| Bcl-2 | F:CTGCACCTGACGCCCTTCAC R:GCGTCCCAGCCTCCGTTGT | 62 | 236 | NM001166486.1 |
| β-actin | F:CTTCAACACCCCTGCCAT R:CTCGGCTGTGGTGGTGAAG | 60 | 238 | JF830811 |
新窗口打开|下载CSV
1.6 RBM3、Bax、Bcl-2蛋白表达Western blot检测
不同处理组卵丘细胞经预冷PBS清洗3次,置于冰盒中,用蛋白裂解液裂解细胞4 h,低温下10 000×g 离心15 min,收集上清,BCA试剂盒检测蛋白浓度并进行平衡,分装置于-80℃备用。检测蛋白在离心管中与5×SDS上样缓冲液混匀,沸水中变性10 min,以 20 μg蛋白/泳道上样,经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)电泳后,电转膜至聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF),室温振荡封闭2 h,分别加入检测目标一抗,37℃摇床孵育2 h,DPBS振荡清洗3次,每次5 min,加辣根过氧化物酶(horse radish peroxidase,HRP)标记的二抗,37℃摇床孵育1 h,DPBS洗涤3次,电化学发光(electrochemiluminescence,ECL)试剂盒进行曝光约 2—5 min,待蛋白条带显色清晰时,立刻拍摄照片,记录试验结果,根据光密度分析蛋白表达水平。1.7 RBM3蛋白定位检测
用PBS清洗不同处理组卵丘细胞3次,加入含有0.1% TritonX-100的固定液常温固定细胞1 h,免疫染色洗涤液清洗3遍后进行2 h室温封闭,加入RBM3抗体(1﹕500)37℃摇床孵育2 h,清洗3遍,用FITC标记的二抗孵育1 h,清洗3遍后用5 ng·mL-1 DAPI室温作用3—5 min,清洗3遍后荧光显微镜观察并成像。1.8 数据分析
采用SPSS19.0统计软件对数据进行单因素方差分析,每组至少重复3次。P<0.05表示差异显著,所有结果以“X±SE”表示。2 结果
2.1 IGF-1及CAPE对牦牛卵丘细胞RBM3表达的调节
用不同浓度IGF-1作用培养至第三代的牦牛卵丘细胞,作用12 h检测发现,IGF-1可显著改变牦牛卵丘细胞RBM3的表达水平,其中IGF-1浓度为100 ng·mL-1处理组RBM3 mRNA和蛋白的表达水平均显著高于其他处理组(图1);50 和200 ng·mL-1 IGF-1处理组中RBM3 mRNA和蛋白的表达水平动态不一致,其中RBM3 mRNA水平在50 ng·mL-1处理组高于其在200 ng·mL-1处理组的表达,但蛋白水平在50 ng·mL-1处理组低于200 ng·mL-1处理组,但两组中RBM3 mRNA和蛋白的表达水平均显著低于100 ng·mL-1 IGF-1处理组,却又显著高于对照组(0 ng·mL-1 IGF-1,图1)。25 μg·mL-1 CAPE处理组中RBM3 mRNA和蛋白水平均显著低于对照组(图1)。图1
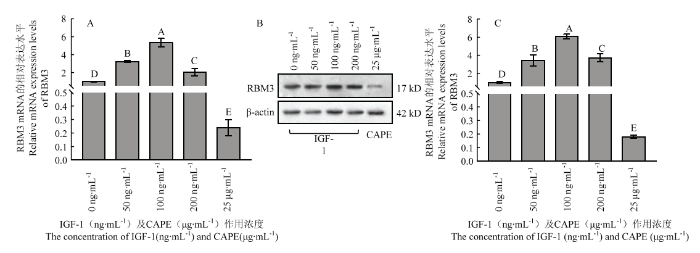 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同浓度IGF-1和CAPE对牦牛卵丘细胞RBM3表达的影响
A:不同浓度IGF-1和CAPE对牦牛卵丘细胞RBM mRNA表达的影响;B:不同浓度IGF-1和CAPE处理组牦牛卵丘细胞RBM3蛋白检测;C:不同浓度IGF-1和CAPE对牦牛卵丘细胞RBM 蛋白表达的影响;不同字母表示差异显著(P<0.05)。下同
Fig. 1Effect of different concentration of IGF-1 and CAPE on the levels of RBM3 in yak cumulus
A: Effect of different concentration of IGF-1 and CAPE on mRNA levels of RBM3 in yak cumulus; B: The detection of RBM3 protein in yak cumulus cells in different groups; C: Effect of different concentration of IGF-1 and CAPE on protein levels of RBM3 in yak cumulus; Different superscripts mean significantly different (P<0.05). The same below
2.2 低温应激对不同处理组牦牛卵丘细胞RBM3表达的调节
将培养至第三代的牦牛卵丘细胞分别置于37、30和25℃的添加下继续培养8 h后,检测RBM3的表达水平。如图2所示,在30和25℃低温应激下,牦牛卵丘细胞RBM3的表达水平显著增高,其mRNA和蛋白的水平均显著高于对照组(37℃),但在30和25℃两组之间RBM3的蛋白和mRNA表达水平差异不显著;将100 ng·mL-1 IGF-处理组的卵丘细胞不同温度低温应激培养8 h后,检测RBM3的表达水平如图3所示,其在30和25℃低温应激处理两组中的水平均显著高于对照组(37℃),但低温应激处理的两组中RBM3的表达水平差异并不显著;将6组卵丘细胞RBM3的表达水平进行对比,如图4所示,以未经IGF-1处理组37℃培养的细胞为对照组,发现经100 ng·mL-1 IGF-处理组的卵丘细胞低温应激后,细胞中RBM3 mRNA和蛋白表达水平均显著高于其中未经IGF-处理而经受低温应激的卵丘细胞中的表达水平。图2
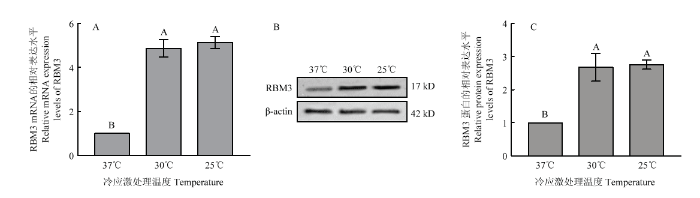 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同温度低温应激对牦牛卵丘细胞RBM3表达的影响
A:不同温度低温应激处理对牦牛卵丘细胞RBM mRNA表达的影响;B:不同温度低温应激处理组牦牛卵丘细胞RBM3蛋白检测;C:不同温度低温应激处理对牦牛卵丘细胞RBM 蛋白表达的影响;不同字母表示差异显著(P<0.05)
Fig. 2Effect of different of temperature on the levels of RBM3 in yak cumulus
A: Effect of different of temperature on mRNA levels of RBM3 in yak cumulus; B: The detection of RBM3 protein in yak cumulus cells in different groups; C: Effect of different of temperature on protein levels of RBM3 in yak cumulus
图3
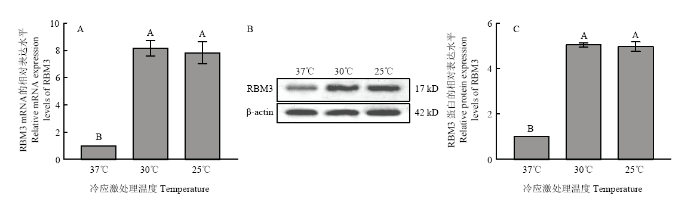 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同温度低温应激对100 ng·mL-1 IGF-1处理组牦牛卵丘细胞RBM3表达的影响
A:不同温度低温应激处理对100 ng·mL-1 IGF-1处理组牦牛卵丘细胞RBM mRNA表达的影响;B:不同温度低温应激处理组牦牛卵丘细胞RBM3蛋白检测;C:不同温度低温应激处理对100 ng·mL-1 IGF-1处理组牦牛卵丘细胞RBM 蛋白表达的影响
Fig. 3Effect of different of temperature on the levels of RBM3 in yak cumulus treated with 100 ng·mL-1 IGF-1
A: Effect of different of temperature on mRNA levels of RBM3 in yak cumulus treated with 100 ng·mL-1 IGF-1; B: The detection of RBM3 protein in yak cumulus cells in different groups; C: Effect of different of temperature on protein levels of RBM3 in yak cumulus treated with 100 ng·mL-1 IGF-1
图4
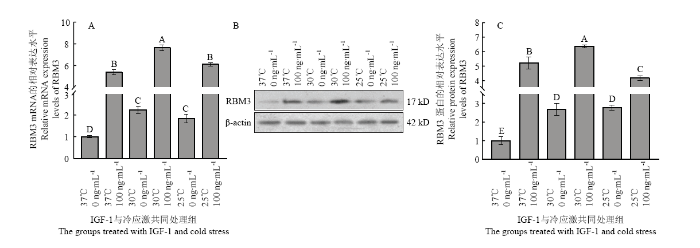 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同处理组卵丘细胞低温应激后RBM3表达比较
A:不同处理组卵丘细胞低温应激后RBM mRNA表达比较;B:不同处理组卵丘细胞低温应激后RBM蛋白表达的检测;C:不同处理组卵丘细胞低温应激后RBM蛋白表达比较
Fig. 4Compared RBM3 levels in yak cumulus stressed at different of temperature and treated with different concentration IGF-1
A: Compared RBM3 mRNA levels in yak cumulus stressed at different of temperature and treated with different concentration IGF-1; B: Detection of RBM3 protein in yak cumulus stressed at different of temperature and treated with different concentration IGF-1; C: Compared RBM3 protein levels in yak cumulus stressed at different of temperature and treated with different concentration IGF-1
检测6个处理组卵丘细胞RBM3的表达水平如图4所示,以未经IGF-1处理组37℃培养的细胞为对照组,发现经100 ng·mL-1IGF-处理组的卵丘细胞低温应激后,细胞中RBM3 mRNA和蛋白表达水平均显著高于其在未经IGF-处理而经受低温应激卵丘细胞中的表达水平。
2.3 RBM3蛋白在不同处理组卵丘细胞分布的检测
对IGF-1不同处理组卵丘细胞用RBM3蛋白进行免疫标记,如图5所示,荧光检测显示未经IGF-1处理组,RBM3主要在牦牛卵丘细胞的细胞质表达(图5-A),50 ng·mL-1 IGF-1处理组RBM 3蛋白也主要表达于细胞质(图5-B),但其荧光强度高于对照组(0 IGF-1)。100和200 ng·mL-1 IGF-1处理组中,卵丘细胞的胞质核细胞核均可强表达RBM蛋白(图5-C,D)。图5
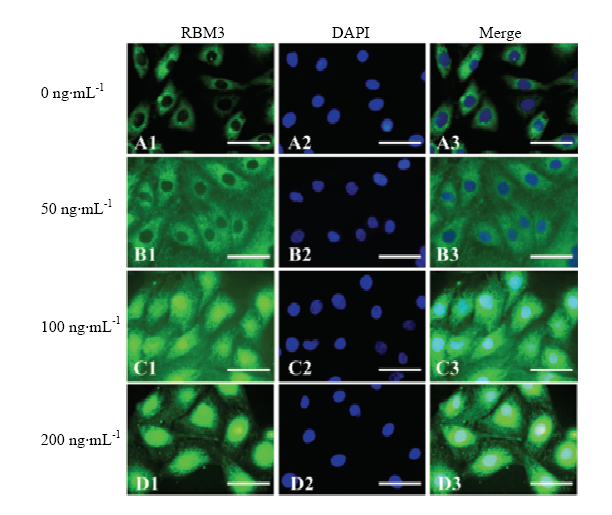 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同浓度IGF-1处理组卵丘细胞RBM3蛋白表达免疫荧光检测绿色荧光为RBM3蛋白;蓝色荧光为标记细胞核的DAPI
绿色荧光为RBM3蛋白;蓝色荧光为标记细胞核的DAPI
Fig. 5Immunofluorescence detection of RBM3 protein in yak cumulus treated with different concentration IGF-1
The HSP70 protein was stained with green fluorescence; Blue fluorescence is the DAPI for nucleus labeling
对照组和100 ng·mL-1 IGF-1处理组卵丘细胞经25℃低温应激8 h后检测后,用免疫荧光技术检测RBM3蛋白,结果如图6所示,低温应激后对照组卵丘细胞胞质和胞核均可检测到RBM3蛋白(图6-A),荧光强度高于未经低温应激处理的对照组(图6-B)。100 ng·mL-1 IGF-1处理组卵丘细胞25℃低温应激前后胞质和胞核均可检测到RBM3蛋白(图6-C),低温应激前经受100 ng·mL-1 IGF-1处理组的卵丘细胞中RBM3蛋白荧光强度高于未经IGF-1处理的低温应激卵丘细胞。
图6
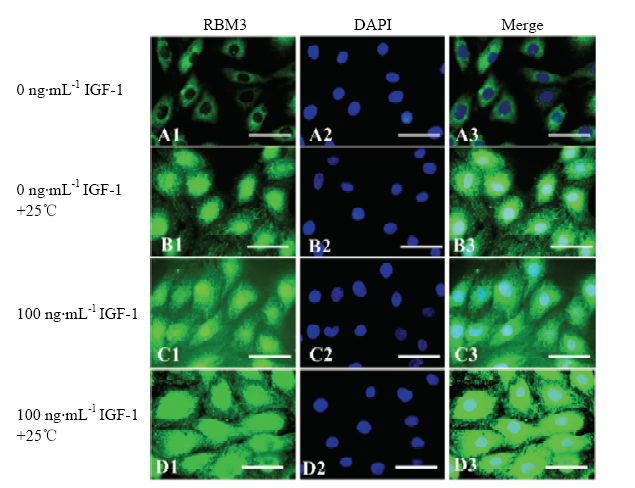 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同浓度IGF-1处理组卵丘细胞低温应激后RBM3蛋白表达免疫荧光检测
绿色荧光为RBM3蛋白;蓝色荧光为标记细胞核的DAPI
Fig. 6Immunofluorescence detection of RBM3 protein in yak cumulus treated with different concentration IGF-1 and stressed at 25℃ for 8 h
The RBM3 protein was stained with green fluorescence; Blue fluorescence is the DAPI for nucleus labeling
2.4 不同处理组后卵丘细胞凋亡率的检测
利用流行细胞仪对不同处理组的细胞进行凋亡检测,如图 7、表 2所示,30和25℃低温应激卵丘细胞8 h均可增加其凋亡率,分别为(16.23±2.81)%和(25.86±1.09)%,100 ng·mL-1 IGF-1处理后的卵丘细胞,其在25℃低温应激8 h凋亡率呈显著降低趋势,凋亡率为(15.94±2.03)%,100 ng·mL-1 IGF-1+CAPE处理组中卵丘细胞25℃低温应激8 h的凋亡率为(20.14±2.65)%,均显著高于对照组中的细胞凋亡率(7.36±0.65)%。图7
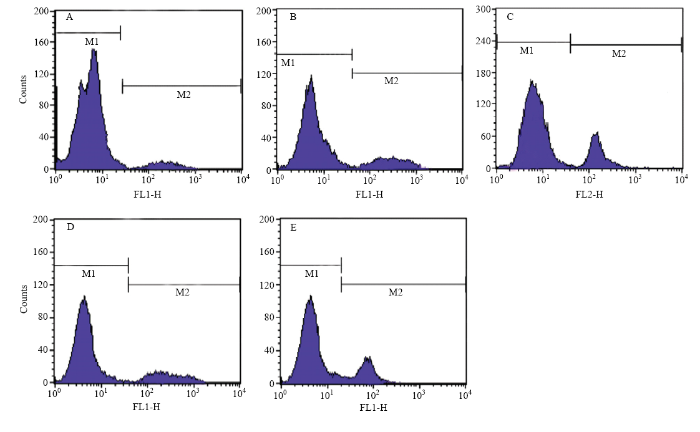 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同处理组牦牛卵丘细胞凋亡率的检测
A:37℃ 卵丘细胞;B:30℃ 应激8h卵丘细胞;C:25℃ 应激8 h卵丘细胞;D:100 ng·mL-1 IGF-1处理组牦牛卵丘细胞25℃ 应激8 h;E:100 ng·mL-1 IGF-1+CAPE处理组牦牛卵丘细胞25℃ 应激8 h
Fig. 7Detection the apoptosis ratio of yak cumulus cells in different groups
A: Yak cumulus at 37℃; B: Yak cumulus stressed at 30℃ for 8 h; C: Yak cumulus stressed at 25℃ for 8 h; D: Yak cumulus treated with 100 ng·mL-1 IGF-1 and stressed at 25℃ for 8 h; E: Yak cumulus treated with 100 ng·mL-1 IGF-1+CAPE and stressed at 25℃ for 8 h
Table 2
表2
表2不同处理组牦牛卵丘细胞凋亡率
Table 2
| 组别 Groups | 细胞凋亡率 Rate of apoptosis (%) |
|---|---|
| 37℃ | 7.36±0.65d |
| 30℃ | 16.23±2.81c |
| 25℃ | 25.86±1.09a |
| 100 ng·mL-1 IGF-1 + 25℃ | 15.94±2.03c |
| 100 ng·mL-1 IGF-1+CAPE+ 25℃ | 20.14±2.65b |
新窗口打开|下载CSV
2.5 不同处理组卵丘细胞低温应激后Bax和Bcl-2表达的变化
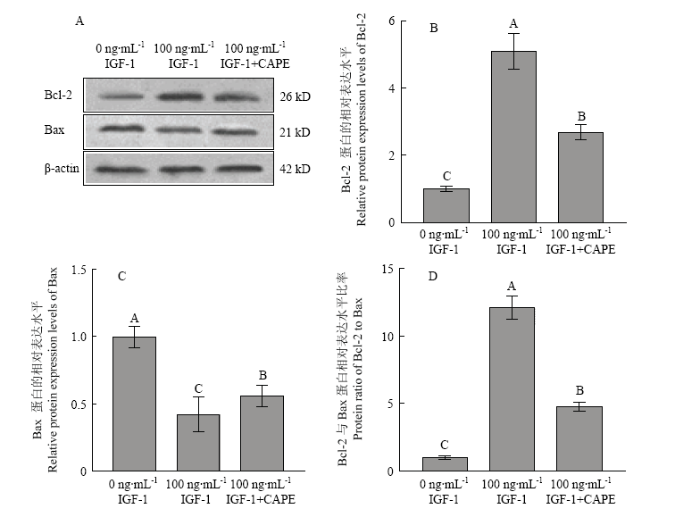
比较对照组、100 ng·mL-1 IGF-1和100 ng·mL-1 IGF-1+CAPE处理组卵丘细胞经25℃低温应激8 h后细胞凋亡相关基因Bax和Bcl-2的表达水平显示,100 ng·mL-1 IGF-1处理组卵丘细胞Bcl-2 mRNA和蛋白的表达水平最高,100 ng·mL-1 IGF-1+CAPE处理组显表达水平次之,对照组(0 IGF-1)表达水平最低(图8-A、9-A和B);而Bax mRNA和蛋白的表达水平在100 ng·mL-1 IGF-1处理组中最低,对照组中最高,100 ng·mL-1 IGF-1+CAPE处理组中Bax mRNA和蛋白的表达处于中间水平(图8-B、9-A和8-C);经100 ng·mL-1 IGF-1处理后,低温应激卵丘细胞中Bcl-2/Bax的比值显著高于其在对照组和100 ng·mL-1 IGF-1+CAPE处理组中的比值,且IGF-1+ CAPE处理组中的比值高于对照组。图8
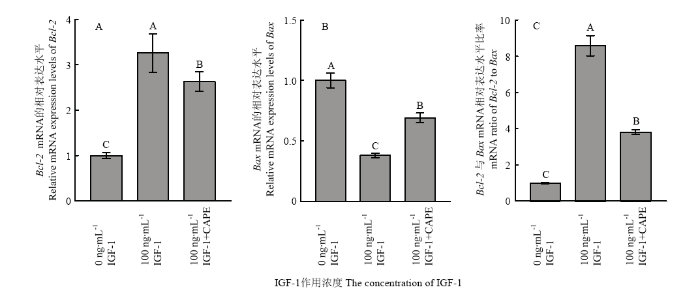 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同处理组卵丘细胞低温应激后Bcl-2, Bax mRNA的表达水平
A:Bcl-2 mRNA的表达水平;B:Bax mRNA的表达水平;C:Bcl-2和Bax mRNA表达水平的比率
Fig. 8Bcl-2 and Bax mRNA levels in yak cumulus from different groups and stressed at 25℃ for 8 h
A: Bcl-2 mRNA levels in yak cumulus; B: Bax mRNA levels; C: Ratio of Bcl-2 to Bax mRNA
图9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9不同处理组卵丘细胞低温应激后Bcl-2, Bax 蛋白的表达水平
A:Bcl-2和Bax蛋白表达Western blot检测;B:Bcl-2 蛋白的表达水平;C:Bax 蛋白的表达水平;D:Bcl-2和Bax蛋白表达水平的比率
Fig. 9Bcl-2 and Bax protein levels in yak cumulus from different groups and stressed at 25℃ for 8 h
A: Detection of Bcl-2 and Bax protein levels in yak cumulus treated with different concentration IGF-1, CAPE and stressed at 25℃ for 8 h; B: Bcl-2 protein levels; C: Bax protein levels; D: Ratio of Bcl-2 to Bax
3 讨论
RBM3和CIRP结合在一起,作为应激调节基因,在正常生理温度下保护体内细胞免受内在刺激造成的损伤外[19],机体遭受低温应激刺激也可诱导其表达[20]。相关****研究发现,由于哺乳动物睾丸组织位于体表,温度往往低于体内核心器官,该器官组织可高水平表达RBM3和CIRP蛋白,其中RBM的表达主要位于睾丸支持细胞,有助于动物精子生成[21]。CIRP和RBM3在体外的生理学功能较为相似,主要为参与细胞或其他器官组织冷冻,尤其是卵母细胞和胚胎的冷冻[22]。因此研究细胞低温应激过程中CIRP和RBM3表达的变化,对细胞冷冻技术的改良尤为重要。卵丘细胞与卵母细胞紧密结合在一起,两者之间可进行信息传导和物质交互,在体外和体内均可促进卵母细胞的成熟。本研究以卵丘细胞为模型,探索其遭受低温应激过程中RBM3表达水平的变化,发现在30℃和 25℃中低温温度应激时,RBM3水平增加,虽在25℃中呈现下降趋势,但差异并不显著,此结果更进一步证实25—34℃温度范围是诱导RBM3表达的最佳低温诱导温度[23,24]。卵丘细胞经受低温诱导高水平表达RBM3也为卵母细胞冷冻技术的优化提供了新的突破点。IGF-1作用动物卵母细胞成熟关键因子,可调控细胞凋亡、增殖与分化等多种信号通路[25]。本研究在卵母细胞体外培养过程中加入不同浓度IGF-1,检测RBM3的水平发现,100 ng·mL-1 IGF-1可显著提高卵丘细胞RBM3的表达,此作用浓度与课题组前提研究结果一致[14],进一步确定IGF-1对牦牛生殖相关细胞体外培养的最佳作用浓度,也与100 ng·mL-1 IGF-1可显著提高卵母细胞CIRP表达的结果相似[15]。为了进一步探索IGF-1对RBM3的提高与温度应激是否存在关联,本研究比较了0和100 ng·mL-1 IGF-1作用的卵丘细胞经受应激后RBM3的表达水平,结果发现IGF-1更进一步提高了卵丘细胞低温应激时RBM3的表达水平,但在IGF-1作用时抑制RBM3的表达,卵丘细胞凋亡水平增高,揭示IGF-1可通过上调牦牛卵丘RBM3的表达提升其对低温应激的适应性,但其在其他细胞中是否具有相似的生物学作用仍需证实。研究同时发现,在IGF-1+CAPE处理组中,卵丘细胞的凋亡水平显著低于对照组,表明除上调RBM3外,IGF-1也可以通过其他信号通路降低低温应激诱导的细胞凋亡。
免疫荧光检测显示,正常条件下RBM3蛋白的表达主要位于牦牛卵丘细胞的细胞质,低浓度IGF-1并不影响RBM3蛋白表达部位的改变。高浓度IGF-1和低温应激条件下卵丘细胞的胞质和胞核均可表达IGF-1,表明RBM3蛋白表达部位发生改变,且IGF-1最佳浓度作用组荧光强度更强,这种变化可能与IGF-1联合RBM3发挥其他生理学功能有关,如两者均可调控细胞生长、分化、胞内其他mRNA的转录和翻译等[5-6, 26]。人的骨髓瘤细胞中,IGF-1可联合AKT激酶细胞通路抑制细胞凋亡[27],本研究中高浓度IGF-1作用后,RBM3蛋白表达部位发生改变可能是由于IGF-1激活AKT激酶,从而改变RBM3磷酸化修饰,影响蛋白质的定位,但其具体机制仍需研究证实。除此之外,也可能与RBM3启动卵丘细胞适应低温应激分子信号有关。
RBM3和CIRP可通过多种信号通路抑制细胞凋亡的发生,如诱导Bcl-2的表达,抑制caspase的活性[28]。低温应激过程中RBM3和CIRP的高水平表达可增加细胞的生长能力,减少低温诱导对其的损害,该生物学功能与细胞凋亡水平减低存在一定关联[29,30]。研究发现,在IGF-1处理组卵丘细胞经低温诱导RBM3表达增加,细胞凋亡率也显著降低,且细胞中抗凋亡基因Bcl-2的表达水平显著高于对照组,Bax水平降低,该结果与IGF-1正常温度条件下可调控牦牛卵丘细胞Bax和Bcl-2表达降低细胞凋亡的结论相似[14]。表明IGF-1降低低温诱导卵丘细胞凋亡的水平与其调控RBM3的表达相关,也为探索IGF-1调控低温诱导相关因子RBM3和CIRP提高冷冻-解冻后细胞的发育能力提供了重要理论依据。
4 结论
牦牛卵丘细胞体外培养过程中100 ng·mL-1胰岛素样生长因子1和中低温诱导均可诱导RNA结合基序蛋白3的表达,且胰岛素样生长因子1可通过调控RNA结合基序蛋白3抑制低温诱导卵丘细胞中B细胞淋巴瘤/白血病基因伴随蛋白x和B细胞淋巴瘤/白血病-2原癌基因的表达,降低细胞凋亡水平。研究结果为揭示胰岛素样生长因子1和RNA结合基序蛋白3参与动物机体或细胞免受低温损伤的分子机制提供了关键信息。有助于通过相关生长因子的改善哺乳动物细胞和胚胎冷冻技术、提高冷冻-解冻后细胞的发育能力。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.neuroscience.2015.08.012URL [本文引用: 2]
DOI:10.1016/j.bbrc.2018.02.209URL [本文引用: 1]
DOI:10.1093/nar/gkw041URL [本文引用: 2]
DOI:10.1002/mc.v55.11URL [本文引用: 1]
[本文引用: 2]
DOI:10.1006/bbrc.1997.7059URL [本文引用: 2]
DOI:10.1038/srep41738URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1210/edrv-6-3-400URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
DOI:10.5603/FM.a2016.0015URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/nar/30.9.e36URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0037043URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00018-016-2253-7URL [本文引用: 1]
DOI:10.1002/(ISSN)1097-4547URL [本文引用: 1]
DOI:10.1038/sj.onc.1205664URL [本文引用: 1]
DOI:10.1016/j.ghir.2019.02.003URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10974-018-9496-xURL [本文引用: 1]
DOI:10.1038/s41598-017-02473-xURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}