Diversity of Endophytic Fungi in Transgenic Rice Seeds from Different Planting Sites Based on PTN System
ZHAO Yan, WANG TianQi, ZHU JunLiCollege of Food Science and Biotechnology, Zhejiang Gongshang University, Hangzhou 310018责任编辑: 李莉
收稿日期:2019-10-9接受日期:2020-02-18网络出版日期:2020-06-01
| 基金资助: |
Received:2019-10-9Accepted:2020-02-18Online:2020-06-01
作者简介 About authors
赵艳,Tel:0571-28008970;E-mail: yanzhao9918@163.com

摘要
关键词:
Abstract
Keywords:
PDF (1830KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
赵艳, 王天圻, 朱军莉. 基于PTN系统分析不同种植地转基因水稻种子 可培养内生真菌菌群的多样性[J]. 中国农业科学, 2020, 53(11): 2305-2320 doi:10.3864/j.issn.0578-1752.2020.11.016
ZHAO Yan, WANG TianQi, ZHU JunLi.
0 引言
【研究意义】水稻是最重要的主粮作物之一,世界上一半以上的人口以稻米为食。基因工程已被用于改善包括水稻、玉米和小麦在内的多种作物的农艺性状和营养价值。评估转基因植物的安全性对于其商业化至关重要,实质等同性是基本原则[1],公众对转基因(genetically modified,GM)作物非预期效应影响人类健康和生态环境的担忧是当前GM作物商品化应用的障碍[2]。研究GM水稻近等基因系的种子内生真菌多样性为进一步评估有关非预期效应奠定科学基础,对农业生物技术的健康发展意义重大。【前人研究进展】GM作物的非预期效应一直是国内外研究热点。JIAO等[3]研究表明GM水稻种子某些营养成分的含量存在非预期变异。赵艳等[2]利用PTN样本系统研究发现GM水稻种子蛋白质非预期变异主要来源于组织培养变异,基因插入的影响较小。PARK等[1]研究证实水稻种子的营养品质不受基因插入的影响,并且表明环境因素对稻谷营养成分的影响比遗传背景的影响更大。GM植物一些代谢物的非预期变异可能影响植物体内环境进而影响植物内生微生物的定植。寄居在植物组织和器官内部,并且不会对宿主造成损害的真菌和细菌等微生物被称为内生菌或内生微生物[4]。前人研究表明转Bt植物可直接或间接地影响农作物周围环境微生物[5]。PAN等[6]报道内生真菌的多样性与玉米植物的基因型有关。刘月廉等[7]发现与亲本相比,GM水稻根部内生真菌分离率、种类多样性以及真菌侵染率和侵染指数无显著差异,转基因对水稻根部内生真菌分布影响不显著。【本研究切入点】将种植地域环境和转基因因素结合探讨GM水稻种子内生菌类群的研究仍鲜见报道。植物内生菌可在植物系统间进行内部传播,水稻种子既是内生菌的携带者和传播者,又是主食稻米的直接原料,内生真菌菌群变异可能涉及病害菌和产毒菌,从而影响水稻的种植生产和食用安全。【拟解决的关键问题】本研究以粳稻品种日本晴(P1)和PJ574(P2)为材料,收集亲本品种(parent variety,P)、转基因株系(transgenic line,T)、及非转基因组培再生株系(non-transgenic regeneration line from tissue culture,NR)等样本,分别建立水稻PTN近等基因系。分析不同种植地、不同品种来源的PTN近等基因系种子内生真菌群落结构特征和生物多样性,比较样本间内生真菌种群结构的差异,探究种植地域环境、品种遗传背景对水稻种子内生真菌多样性的影响贡献大小,为从种子内生菌分布角度客观评估GM水稻的非预期效应奠定基础。1 材料与方法
1.1 供试材料
水稻转基因株系(T)以及非转基因对照样本(P、NR)的来源:水稻亲本(P)→愈伤组织的诱导培养和感受态细胞制备→外源基因2mG2-epsps转化愈伤组织→草甘膦抗性愈伤组织的筛选→再生植株→转基因株系(T)和非转基因组培再生株系(NR)。具体包括粳稻日本晴品种(P1)及其相应的GM株系T16和组培对照株系NR25组成的P1近等基因系(P1 near-isogenic line,P1L);粳稻品种PJ574(P2)及其相应的GM株系T23和组培对照株系NR18组成的P2近等基因系(P2 near-isogenic line,P2L)。供试水稻样本于2017年12月种植于海南省(Hainan province,H),2018年5月收获。2018年6月种植于浙江省富阳市(Fuyang,Zhejiang province,F),11月收获。收获种子自然晾干后,4℃保存,种子收获后2个月内完成微生物分离试验,尽量减少储存时间的影响。
1.2 水稻种子可培养内生真菌的分离纯化
水稻种子表面消毒:水稻种子脱壳后,参照CUI等[8]方法对水稻种子表面消毒,用无菌水冲洗种子5次,洗去表面灰尘杂质,再依次使用70%乙醇浸泡3 min、5%次氯酸钠浸泡5 min、70%乙醇清洗30 s、最后无菌水冲洗6次。取最后一次清洗液100 μL涂布在PDA培养基,28℃培养7 d,作为检验种子表面彻底消毒的对照。内生真菌的分离:采用组织块分离法[8],将表面消毒的种子放到垫有滤纸的无菌平板上,超净台上稍晾干,切成两段,切面朝下半埋接种于PDA培养基。在28℃恒温培养箱中培养6—10 d,待组织块周围长出菌丝体后,挑取菌丝接到PDA培养基上纯化培养。
1.3 水稻种子可培养内生真菌的鉴定
无菌条件下,将纯化后的真菌续接到PDA固体培养基,28℃培养7 d后观察菌落形态并拍照记录。刮取足量的菌丝体到灭菌洁净研钵中,用液氮充分研磨。采用CTAB法提取菌体DNA[9],以DNA为模板,以ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)为引物进行PCR扩增[8],PCR反应体系(50 μL):5 μL 10×PCR buffer(含Mg2+)、1 μL上游引物、1 μL下游引物、3 μL DNA模板、4 μL dNTP、1 μL Taq酶(最后加),dH2O补足50 μL。PCR扩增程序为94℃ 5 min;94℃ 45 s,54℃ 30 s,72℃ 90 s,32个循环;72℃ 10 min。PCR产物经1.0%琼脂糖凝胶电泳检测。PCR产物送生工生物工程(上海)股份有限公司测序。用Blast软件将测序结果与GenBank中已知菌株的ITS rDNA序列进行同源性比对鉴定。
1.4 数据处理与统计分析
对稻米内生真菌的分离率(isolation rate,IR)、分离频率(isolation frequency,IF)进行统计,用Margalef指数(D)分析真菌类群的丰富度,Shannon-Weiner多样性指数(H')、Evenness均匀度指数(E)及Sorenson相似性系数(Cs)分析内生真菌的多样性及分布特征[10]。分离率IR(%)=(稻米样本长菌组织块数/稻米全部样本组织块数)×100。分离率是衡量植物组织中内生真菌的定植丰度和每个组织块受侵染的发生频率。
分离频率IF(%)=(Ni/N)×100。其中:Ni为某种稻米内生真菌的菌株数;N为全部内生真菌菌株数。
IF>10%的内生真菌为优势菌,1%<IF≤10%的内生真菌为常见菌,IF≤1%为稀有菌[11]。当N取值为所有样本内生真菌合集时,某种真菌相应占比用IF表示;当N取值为部分特定样本组合时某种真菌相应占比称为相对分离频率(relative frequency,RF),便于样本间的直观比较。
Margalef指数D=(S-1)/ln N。其中:S为类群总数;N为内生真菌总数目。
Shannon-Wiener多样性指数H'=-$\sum _{i=\text{1}}^{k}$(Ni/N)×ln(Ni/N)。其中,k为所有真菌种类的数目;Ni为某种真菌的数目;N为内生真菌总数目。
均匀度指数E=H'/lnS。其中:H'为Shannon-Wiener指数,S为群落中内生真菌的类群总数。
Sorenson相似性指数Cs=2j/(a+b)。式中:j为两种植地(或品系或不同基因型)共有的种数或属数;a为一个种植地(或品系或不同基因型)内生真菌的种数或属数;b为另一种植地(或品系或不同基因型)内生真菌的种数或属数。当Cs为0.00—0.25时,极不相似;当Cs为0.25—0.50时,中等不相似;当Cs为0.50—0.75时,中等相似;当Cs为0.75—1.00时,极相似。
利用软件spss17.0进行Fisher精确检验(Fisher's exact test)分析比较样本间真菌菌群组成的差异显著性。
2 结果
2.1 H和F种植地水稻种子可培养内生真菌的组成
接种后在28℃培养7 d,洗涤液对照无菌落长出(图1-a),说明试验稻米材料表面已彻底消毒。待组织块在PDA中长出较明显菌落时(图1-b—图1-e)续接纯化培养并进行菌落形态观测。从不同种植地(H、F)PTN系统样本组成的2个近等基因系2 400个稻米组织块中共分离内生真菌121株,总分离率(IR)为5.04%(表1),根据形态结构合并为43种真菌(图2)。以提取的真菌DNA为模版,ITS1和ITS4为引物的PCR扩增条带约在550 bp,测序后进行BLAST分析,将相似度>97%的鉴定为同属,>99%的鉴定为同种[12],因此,将43个菌株归为3纲、4目、7科、15属(表1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1部分水稻种子内生真菌分离图
a:对照;b、c、d、e:菌落分离图
Fig. 1Partial separation mapping of endophytic fungi from rice seeds
a: Control; b, c, d, e: Colony separation diagram
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2水稻种子内生真菌在PDA平板上的菌落形态
上:培养皿正面;下:培养皿反面
Fig. 2Colony morphology of endophytic fungi from rice seeds on PDA plates
Upper: Petri dish front; Bottom: Petri dish reverse side
Table 1
表1
表1水稻种子可培养内生真菌的鉴定结果
Table 1
| 编号 Number | 纲 Class | 目 Order | 科 Family | 属 Genera | 比对相似种 Compare similar species | 覆盖率 Coverage (%) | 相似度 Similarity (%) | 比对结果 Comparison result | IR (%) |
|---|---|---|---|---|---|---|---|---|---|
| WF6 | Ascomycetes | Pleosporales | Pleosporaceae | Curvularia | Curvularia lunata strain S3-2 | 97 | 99 | Curvularia lunata strain WF6 | 0.54 |
| WF44 | Curvularia lunata isolate E16 | 97 | 99 | Curvularia lunata isolate WF44 | 0.08 | ||||
| 合计Total | 0.63 | ||||||||
| WF17 | Pleosporales | Pleosporales sp. strain APBSDSF36 | 97 | 100 | Pleosporales sp. strain WF17 | 0.04 | |||
| 合计Total | 0.04 | ||||||||
| WF22 | Phaeosphaeria | Phaeosphaeriopsis musae isolate DFFSCS029 | 99 | 99 | Phaeosphaeriopsis musae strain WF22 | 0.04 | |||
| WF55 | Phaeosphaeria papayae isolate LQ122404 | 98 | 99 | Phaeosphaeria papayae isolate WF55 | 0.04 | ||||
| 合计Total | 0.08 | ||||||||
| WF40 | Setophoma | Setophoma vernoniae strain CPC 23123 | 92 | 93 | Setophoma vernoniae strain .WF40 | 0.13 | |||
| 合计Total | 0.13 | ||||||||
| WF39 | Ascomycetes | Pleosporales | Pleosporaceae | Ophiosphaerella | Ophiosphaerella agrostidis strain MFLUCC 11-0152 | 95 | 99 | Ophiosphaerella agrostidis strain.WF39 | 0.13 |
| WF58 | Ophiosphaerella agrostidis isolate MFLUCC 16-0895 | 94 | 99 | Ophiosphaerella agrostidis isolate WF58 | 0.04 | ||||
| 合计Total | 0.17 | ||||||||
| WF41 | Dendryphiella | Dendryphiella sp. crri.34 | 98 | 99 | Dendryphiella sp. WF41 | 0.83 | |||
| WF42 | Dendryphiella sp. crri.34 | 97 | 99 | Dendryphiella sp. WF42 | 0.75 | ||||
| WF45 | Dendryphiella sp. crri.34 | 98 | 99 | Dendryphiella sp.WF45 | 0.08 | ||||
| WF46 | Dendryphiella sp. crri.34 | 98 | 99 | Dendryphiella sp.WF46 | 0.08 | ||||
| WF59 | Dendryphiella sp. crri.34 | 98 | 99 | Dendryphiella sp. WF59 | 0.13 | ||||
| 合计Total | 1.88 | ||||||||
| WF48 | Bipolaris | Bipolaris oryzae isolate BO1 | 97 | 99 | Bipolaris oryzae isolate WF48 | 0.04 | |||
| WF62 | Bipolaris sorokiniana strain A0606 | 95 | 97 | Bipolaris sorokiniana strain WF62 | 0.04 | ||||
| 合计Total | 0.08 | ||||||||
| WF49 | Pithomyces | Leptosphaerulina chartarum strain DH08111quan1 | 98 | 99 | Leptosphaerulina chartarum strain WF49 | 0.04 | |||
| 合计Total | 0.04 | ||||||||
| WF43 | Didymellaceae | Epicoccum | Epicoccum sorghinum strain BJ-F1 | 97 | 99 | Epicoccum sorghinum strain.WF43 | 0.08 | ||
| WF47 | Epicoccum sorghinum isolate M3 | 98 | 99 | Epicoccum sorghinum isolate WF47 | 0.13 | ||||
| WF52 | Epicoccum sorghinum isolate M3 | 97 | 99 | Epicoccum sorghinum isolate WF52 | 0.38 | ||||
| WF53 | Epicoccum sorghinum isolate M3 | 96 | 99 | Epicoccum sorghinum isolate WF53 | 0.04 | ||||
| 合计Total | 0.63 | ||||||||
| WF51 | Leptosphaeriaceae | Leptosphaeria | Leptosphaeria sp. SL14 | 99 | 99 | Leptosphaeria sp. WF51 | 0.04 | ||
| WF61 | Leptosphaeria sp. isolate LQ122416 | 98 | 99 | Leptosphaeria sp. isolate WF61 | 0.04 | ||||
| WF64 | Leptosphaeria sp. isolate LQ122417 | 98 | 99 | Leptosphaeria sp. isolate WF64 | 0.04 | ||||
| 合计Total | 0.13 | ||||||||
| WF20 | Moniliales | Dematiaceae | Nigrospora | Nigrospora sp. JL4-CGL24 | 99 | 99 | Nigrospora sp. WF20 | 0.04 | |
| WF57 | Nigrospora sp. HCH267 | 98 | 99 | Nigrospora sp. WF57 | 0.04 | ||||
| WF60 | Nigrospora oryzae strain APBSMLF76 | 98 | 99 | Nigrospora oryzae strain WF60 | 0.04 | ||||
| 合计Total | 0.13 | ||||||||
| WF3 | Cladosporium | Cladosporium oxysporum isolate DFFSCS018 | 100 | 99 | Cladosporium sp. WF3 | 0.04 | |||
| WF19 | Cladosporium oxysporum strain SCAU101 | 99 | 99 | Cladosporium oxysporum strain WF19 | 0.04 | ||||
| WF63 | Cladosporium xanthochromaticum | 98 | 99 | Cladosporium xanthochromaticum WF63 | 0.08 | ||||
| WF65 | Cladosporium tenuissimum strain APBSDSF6 | 99 | 99 | Cladosporium tenuissimum strain WF65 | 0.04 | ||||
| 合计Total | 0.21 | ||||||||
| WF4 | Alternaria | Alternaria sp. H6_E08_1104035148Q | 100 | 99 | Alternaria sp.WF4 | 0.04 | |||
| WF18 | Alternaria alternata strain EN24 | 98 | 100 | Alternaria alternata strain WF18 | 0.04 | ||||
| WF31 | Alternaria longissima strain CLB44 | 98 | 99 | Alternaria longissima strain WF31 | 0.04 | ||||
| 合计Total | 0.13 | ||||||||
| WF9 | Tuberculariaceae | Fusarium | Fusarium chlamydosporum NBAIM:236 | 98 | 100 | Fusarium chlamydosporum strain WF9 | 0.04 | ||
| WF14 | Fusarium chlamydosporum isolate LrSF22 | 99 | 99 | Fusarium chlamydosporum strain WF14 | 0.04 | ||||
| WF15 | Fusarium sp. AL-14 IRH-2012a | 98 | 99 | Fusarium sp. WF15 | 0.04 | ||||
| WF16 | Fusarium equiseti strain CP16 | 99 | 99 | Fusarium equiseti strain WF16 | 0.04 | ||||
| WF38 | Fusarium sp. Z13-09 | 96 | 99 | Fusarium sp.WF38 | 0.08 | ||||
| WF50 | Fusarium sp. AL-15 IRH-2012b | 98 | 99 | Fusarium sp.WF50 | 0.38 | ||||
| WF54 | Fusarium proliferatum isolate MC-23-F | 99 | 99 | Fusarium proliferatum isolate WF54 | 0.04 | ||||
| WF56 | Fusarium proliferatum isolate 10R-7 | 98 | 99 | Fusarium proliferatum isolate WF56 | 0.04 | ||||
| WF66 | Fusarium proliferatum isolate LrBF18 | 99 | 99 | Fusarium proliferatum isolate WF66 | 0.04 | ||||
| 合计Total | 0.75 | ||||||||
| WF23 | Fungi Imperfecti | Hypocreales | Hypocreaceae | Letendraea helminthicola | Letendraea helminthicola strain B1A0062SNA2CC1081 | 88 | 100 | Letendraea helminthicola strain WF23 | 0.04 |
| 合计Total | 0.04 | ||||||||
| 总分离率Total IR(%) | 5.04 | ||||||||
新窗口打开|下载CSV
2.2 不同种植地水稻种子内生真菌的多样性比较
2.2.1 海南省和浙江省富阳市两地水稻样本间的综合比较 海南省种植(H)的水稻样本共分离内生真菌20株,归为9个属,其中8个属为子囊菌纲,1个属为半知菌纲。浙江省富阳市(F)种植地分离内生真菌101株,归为12个属,全部属于子囊菌纲。可见水稻种子可培养内生真菌大多为子囊菌纲。H地水稻种子内生真菌的总IR(0.83%)显著低于F地样本(4.61%)。以两种植地所有水稻样本内生真菌菌株总数作为N值,计算不同样本中各内生真菌属的IF(表2),分析可知H和F两地内生真菌菌群的组成既有共性,又存在一定的差异。两地的共有真菌包括Curvularia、Phaeosphaeria、Setophoma、Nigrospora、Cladosporium和Fusarium 6个属,共46株,分别占H和F地总菌株数的75.00%和30.70%,占两地总菌株38.02%。H地特有Alternaria、Letendraea helminthicola和Pleosporales 3个菌属共5株,占两地总菌数4.13%。F地特有真菌包括6个属共70株,分别为Ophiosphaerella、Dendryphiella、Epicoccum、Bipolaris、Pithomyces和Leptosphaeria,占比高达F地总菌株的69.3%和两地总菌数的57.85%。表明水稻种子内生真菌种群具有明显的地理分布特性。
Table 2
表2
表2不同种植地水稻种子内生真菌分离频率比较(IF)(%)
Table 2
| 属 Genera | 海南省Hainan (H) | 浙江省富阳市Fuyang, Zhejiang (F) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HP1L | HP2L | FP1L | FP2L | |||||||||
| HP1 | HT16 | HNR25 | HP2 | HT23 | HNR18 | FP1 | FT16 | FNR25 | FP2 | FT23 | FNR18 | |
| Curvularia | — | 1.65 | — | 1.65 | 0.83 | — | 5.79 | — | — | — | 2.48 | — |
| Pleosporales | — | 0.83 | — | — | — | — | — | — | — | — | — | — |
| Phaeosphaeria | — | — | — | 0.83 | — | — | — | 0.83 | — | — | — | — |
| Setophoma | — | — | 1.65 | — | — | — | 0.83 | — | — | — | — | — |
| Ophiosphaerella | — | — | — | — | — | — | 2.48 | 0.83 | — | — | — | — |
| Dendryphiella | — | — | — | — | — | — | 12.40 | 5.79 | 8.26 | 3.31 | 0.83 | 6.61 |
| Bipolaris | — | — | — | — | — | — | 0.83 | — | — | 0.83 | — | — |
| Pithomyces | — | — | — | — | — | — | 0.83 | — | — | — | — | — |
| Epicoccum | — | — | — | — | — | — | 3.31 | 1.65 | 0.83 | 0.83 | 4.96 | 0.83 |
| Leptosphaeria | — | — | — | — | — | — | 0.83 | — | — | 0.83 | 0.83 | — |
| Nigrospora | — | — | — | 0.83 | — | — | — | 0.83 | — | 0.83 | — | — |
| Cladosporium | — | 0.83 | — | — | — | 0.83 | — | — | — | — | 2.48 | — |
| Alternaria | — | 0.83 | — | 0.83 | — | 0.83 | — | — | — | — | — | — |
| Fusarium | — | 1.65 | — | 0.83 | 0.83 | — | 3.31 | 4.13 | — | 1.65 | 1.65 | 0.83 |
| Letendraea helminthicola | — | — | — | — | 0.83 | — | — | — | — | — | — | — |
| 合计分离频率 Sum of IF (%) | 0.00 | 5.79 | 1.65 | 4.96 | 2.48 | 1.65 | 30.58 | 14.05 | 9.09 | 8.26 | 13.22 | 8.26 |
| 7.44 | 9.09 | 53.72 | 29.75 | |||||||||
| 16.53 | 83.47 | |||||||||||
新窗口打开|下载CSV
H地6个样本内生真菌的IF和所含真菌类群的Margalef(D)、Shannon-Wiener(H')以及均匀度指数(E)均比对应F地的低(表2和表3),表明F地内生真菌要比H地更加丰富和多样。两地水稻种子样本内生真菌类群属水平上Cs为0.57,为中等相似(表4),并且Fisher精确检验结果显示二者分析两地真菌菌群组成差异显著(P<0.05)(表5),表明水稻种子内生真菌群落的组成在一定程度上受地理环境影响。
Table 3
表3
表3海南省(H)和浙江省富阳市(F)种植的PTN近等基因系水稻种子内生真菌菌群多样性指数
Table 3
| 样本 Samples | 海南省Hainan (H) | 浙江省富阳市Fuyang, Zhejiang (F) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 菌株数 No. of strains | 属数 No. of genus | 丰富度指数 Margalef index (D) | 多样性指数 Shannon- Wiener index(H') | 均匀度指数 Evenness index (E) | 菌株数 No. of strains | 属数 No. of genus | 丰富度指数 Margalef index (D) | 多样性指数 Shannon- Wiener Index (H') | 均匀度指数 Evenness index (E) | |
| P1 | 0 | 0 | — | — | — | 37 | 9 | 1.92 | 1.32 | 0.60 |
| T16 | 7 | 5 | 1.82 | 1.40 | 0.87 | 17 | 6 | 1.20 | 0.74 | 0.41 |
| NR25 | 2 | 1 | 0.00 | 0.33 | 11 | 2 | 0.24 | 0.35 | 0.51 | |
| P1L | 9 | 6 | 1.67 | 1.14 | 0.64 | 65 | 11 | 2.17 | 1.35 | 0.56 |
| P2 | 6 | 5 | 1.67 | 1.18 | 0.73 | 10 | 6 | 1.40 | 0.80 | 0.45 |
| T23 | 3 | 3 | 0.83 | 0.65 | 0.60 | 16 | 6 | 1.40 | 1.07 | 0.60 |
| NR18 | 2 | 2 | 0.42 | 0.44 | 0.63 | 10 | 3 | 0.56 | 0.53 | 0.49 |
| P2L | 11 | 7 | 0.30 | 1.34 | 0.69 | 36 | 8 | 1.52 | 0.99 | 0.48 |
| 合计Tatol | 20 | 9 | 1.67 | 0.63 | 0.29 | 101 | 12 | 2.29 | 1.63 | 0.66 |
新窗口打开|下载CSV
Table 4
表4
表4不同种植地水稻PTN近等基因系样本种子内生真菌菌群的相似性系数(Cs)
Table 4
| 样本 Samples | 海南省Hainan (H) | 浙江省富阳市Fuyang, Zhejiang (F) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | T16 | NR25 | P2 | T23 | NR18 | P1 | T16 | NR25 | P2 | T23 | NR18 | |||
| P1 | 1.00 | 1.00 | ||||||||||||
| T16 | 0.00 | 1.00 | 0.53 | 1.00 | ||||||||||
| NR25 | 0.00 | 0.00 | 1.00 | 0.36 | 0.50 | 1.00 | ||||||||
| P2 | 0.00 | 1.00 | 0.67 | 1.00 | ||||||||||
| T23 | 0.50 | 1.00 | 0.67 | 1.00 | ||||||||||
| NR18 | 0.29 | 0.00 | 1.00 | 0.67 | 0.67 | 1.00 | ||||||||
| HP2L | FP2L | |||||||||||||
| P1L | 0.62 | 0.74 | ||||||||||||
| F | ||||||||||||||
| H | 0.57 | |||||||||||||
新窗口打开|下载CSV
Table 5
表5
表5不同种植地水稻PTN近等基因系样本种子内生真菌菌群组成的差异性分析(P)
Table 5
| 样本 Samples | 海南省Hainan (H) | 浙江省富阳市Fuyang, Zhejiang (F) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | T16 | NR25 | P2 | T23 | NR18 | P1 | T16 | NR25 | P2 | T23 | NR18 | |||
| P1 | 1.00 | 1.00 | ||||||||||||
| T16 | — | 1.00 | 0.23 | 1.00 | ||||||||||
| NR25 | — | 0.09 | 1.00 | 0.38 | 0.07 | 1.00 | ||||||||
| P2 | — | 1.00 | 0.33 | 1.00 | ||||||||||
| T23 | 1.00 | 1.00 | 0.05 | 1.00 | ||||||||||
| NR18 | 0.86 | 1.00 | 1.00 | 0.61 | 0.00 | 1.00 | ||||||||
| HP2L | FP2L | |||||||||||||
| P1L | 0.86 | 0.18 | ||||||||||||
| F | ||||||||||||||
| H | 0.00 | |||||||||||||
新窗口打开|下载CSV
2.2.2 亲本品种及其PTN近等基因系种子内生真菌分布的地域差异比较 按地域分组计算RF值比较两地样本的内生优势菌属(RF>10%,图3),Fusarium是两地水稻种子共有优势菌属。H地样本有3个优势菌属,其中Curvularia菌属在F地样本中丰度降为常见菌属,Alternaria在F地未检出。F地独有优势菌属Dendryphiella和Epicoccum广泛存在于6个样本中(表2),可见不同种植地域水稻种子内生真菌的优势菌群组成存在差异。
图3
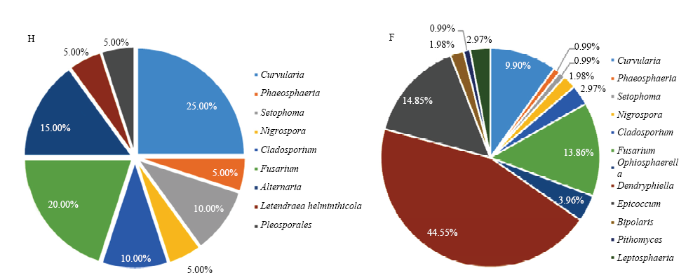 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3海南省(H)和浙江省富阳市(F)种植水稻种子内生真菌的相对分离频率(RF)
Fig. 3Relative isolation frequency ( RF ) of endophytic fungi from rice seeds grown in Hainan (H) and Fuyang, Zhejiang (F)
将两地种植的2个PTN近等基因系共6个供试水稻样本种子可培养内生真菌菌群组成和多样性指数汇总于表3。观察相同品种水稻种植于不同地点时的种子内生真菌类群丰富度和多样性。对P1品种而言,HP1和FP1样本表现为“无”和“有”的极端差异,可见天然亲本品种P1种子内生真菌类群分布受种植地域影响很大。将观测样本群体扩大至P1近等基因系P1L,除均匀度指数(E)外,浙江省富阳市种植的FP1L近等基因系分离的真菌株数、菌属数、丰富度指数(D)和多样性指数(H')都高于相应海南省种植样本(表3),这与上述H和F两种植地所有水稻样本的内生真菌多样性变化趋势一致。对P2品种而言,虽然FP2分离的内生真菌株数和属数比HP2多,但多样性指数均低于相应的HP2样本;将观测样本群体扩大至P2近等基因系P2L,FP2L分离的内生真菌株数和菌属数仍比HP2L多,Margalef(D)指数1.52也大幅高于海南省种植HP2L群体的0.30。
2.3 不同品种水稻种子内生真菌的多样性比较
综合表2和表3结果可分析不同样本间种子内生真菌的组成和分布差异,探测水稻种子非预期变异的特征和变异来源。以亲本品种P1和P2为比对组,分析天然水稻品种之间种子内生真菌菌群结构和多样性变异程度。海南省种植地样本中,HP1未检出真菌,HP2分离到内生真菌6株,归为Curvularia、Phaeosphaeria、Nigrospora、Alternaria和Fusarium 5个属(表2)。可见在海南省种植时P2品种内生真菌类群比P1丰富得多。浙江省富阳市种植地样本中,FP1分离37株内生真菌归为9个属,FP2获得10株内生真菌归为6个属,FP2的IF(8.26%)显著低于FP1(30.58%)。Curvularia、Setophoma、Ophiosphaerella和Pithomyces 4个为FP1特有菌属,Nigrospora为FP2特有菌属;FP1和FP2共有菌属5个,包括Dendryphiella、Bipolaris、Epicoccum、Leptosphaeria和Fusarium。FP2内生真菌的多样性指数Margalef(D)、Shannon-Wiener(H')以及均匀度指数(E)均低于FP1,下降幅度分别达27.08%、39.39%和25.00%,说明浙江省富阳市种植的P1品种内生真菌类群比P2更丰富多样。
综合两地,在属水平,P1和P2之间种子内生真菌菌群的相似性系数(Cs)分别为0(H)和0.67(F)(表4),表现极不相似和中等相似;近等基因系P1L和P2L之间内生真菌菌群的Cs分别为0.62(H)和0.74(H),为中等相似,Fisher' exact test检验显示二者差异均不显著(P>0.05)(表5)。
在内生真菌类群组成上,HP2和FP2样本都分离出了Nigrospora和Fusarium,说明这两个菌属是P2品种的固有菌属,只是Fusarium菌属丰度因地域而表现不同,FP2中Fusarium菌属的IF(1.65%)是HP2的2倍(表2)。综合表2和表3,可知HP2中的3个菌属Curvularia、Phaeosphaeria、Alternaria和FP2中的Dendryphiella等4个菌属在水稻种子内定植主要受地域影响。对P1品种而言,只在F地分离到9个属真菌,种植地域影响更为突出。
2.4 GM水稻种子内生真菌的非预期变异特征
海南省种植的GM水稻株系T16(HT16)种子共分离7株内生真菌,总IF为5.79%,归为Curvularia、Fusarium、Pleosporales、Cladosporium和Alternaria 5个属(表2),相应亲本HP1未检出真菌,二者相似性系数为0,完全不等同。浙江省富阳市种植时,GM水稻株系FT16种子内生真菌总IF(14.05%)极显著低于相应亲本FP1(30.58%),丰富度指数(D)、多样性指数(H')和均匀度指数(E)也都低于FP1样本。菌群结构上,FT16与FP1样本共有Ophiosphaerella、Dendryphiella、Epicoccum和Fusarium 4个内生真菌属,未检出FP1携带的Bipolaris、Leptosphaeria、Curvularia、Setophoma和Pithomyces菌属,却额外增加了2个特有菌属Phaeosphaeria和Nigrospora。FT16与FP1内生真菌类群的相似性系数0.53,为中等相似(表4),且二者内生真菌的组成不具有显著差异(P=0.23>0.05,表5)。海南省种植的GM水稻株系T23(HT23)种子内生真菌总IF(2.48%)显著低于亲本对照HP2(4.96%)。菌群组成上,与亲本HP2相比,HT23携带的3个菌属中Letendraea helminthicola为新增特有菌属,Curvularia和Fusarium与HP2所共有,此外未检出HP2特有菌属Phaeosphaeria、Nigrospora和Alternaria,二者的相似性系数为0.50,中等相似,且不具有显著差异(表5)。浙江省富阳市种植的FT23样本内生真菌总分离率13.22%显著高于FP2。FT23分离获得6个属共16株真菌,其中Dendryphiella、Epicoccum、Leptosphaeria和Fusarium 4个菌属与亲本FP2共有,Curvularia和Cladosporium为新增特有菌属,比FP2减少了2个菌属Bipolaris和Nigrospora。FT23种子内生真菌类群与FP2的相似性系数0.67,中等相似,二者内生真菌的组成差异不显著(P=0.05)。
综合两地样本,GM水稻品系T16种子固有的内生真菌只有Fusarium属,在H和F两地的IF分别为1.65%和4.13%;T23种子固有的内生真菌包括Curvularia和Fusarium。Nigrospora和Fusarium是亲本品种P2种子固有内生真菌(表2)。
2.5 GM水稻种子内生真菌类群的非预期变异技术来源解析
两地种植的P1L和P2L近等基因系中,与相应亲本相比,2个GM株系种子内生真菌总分离频率(IF)在海南省和浙江省富阳市的变异方向相反,但在相同种植地时又分别与相应组培对照NR株系变异方向相同(表2)。以T16为例,HT16种子内生真菌总IF(5.79%)显著高于HNR25(1.65%),变异方向一致只是大小不同,其中GM水稻品HT16种子内生真菌分离频率增加的变异效应中转基因插入突变效应>组织培养无性系变异效应。但浙江省富阳市种植时,FT16种子内生真菌总IF(14.05%)比亲本FP1降低的幅度显著低于FNR25(9.09%),因此,推测HT16种子内生真菌分离频率降低的变异效应中转基因插入突变效应<组织培养无性系变异效应。对GM水稻品系T23而言,海南省种植的HT23和组培对照HNR18的种子内生真菌总IF均显著低于亲本对照HP2(4.96%),二者变异方向一致而且HNR18样本变异幅度更大,似乎这种内生真菌总分离率降低的变异效应来源于组织培养无性系变异。但浙江省富阳市种植的FT23样本内生真菌总IF13.22%极显著高于FP2(8.26%),组培对照FNR18与FP2亲本对照相同(表2),又可将该变异效应归因于转基因插入突变。综合2个种植地变异方向相反且技术来源不甚明确的分析结果,认为GM水稻品系种子内生真菌总IF的波动变异受种植地域环境影响较大,非预期变异效应较小。菌群组成和结构上,HT16种子携带内生真菌Curvularia等5个属,HNR25却只有Setophoma 1个属(表2);FT16额外增加的2个菌属Phaeosphaeria和Nigrospora不存在于其相应组培对照样本FNR25中,说明T16种子定植真菌类群增加的变异主要为转基因插入突变效应。与FP1相比,FT16种子内生真菌减少了Bipolaris等5个菌属,组培对照样本FNR25却减少了7个菌属,而且其中5个菌属与FT16的一致,由此可知GM水稻株系T16种子内生真菌类群的消减变异效应为组织培养无性系变异。同理,HT23与亲本HP2相比增加了1个菌属Letendraea helminthicola并且该菌属只出现在该样本中,浙江省富阳市种植的FT23比亲本FP2增加了Curvularia和Cladosporium 2个菌属,这种内生真菌菌属增多的变异并未在相应组培参照样本中出现,可判断变异来源于转基因插入突变,需要重点关注。海南省种植的HT23比HP2减少3个菌属Phaeosphaeria、Nigrospora和Alternaria,HNR18比HP2减少4个菌属Curvularia、Phaeosphaeria、Nigrospora和Fusarium,浙江省富阳市种植的FT23与组培参照样本FNR18一样也减少了Bipolaris和Nigrospora 2个菌属,可知GM水稻品系T23种子内生真菌减少的变异主要受组织培养无性系变异效应的影响。
2.6 不同样本水稻种子内生真菌分布的聚类分析及影响因素主次评价
在属水平上,根据种子可培养内生真菌的种类和丰度进行聚类分析(图4),可见两地种植的12个水稻样本中,聚类距离最近的样本组包括FNR25和FNR18、FT16和HP2、HP1和HNR25。分析样本具体来源可知,HP1和HNR25为海南省种植的P1品种及其组培品系,遗传关系相近外;FNR25和FNR18对应P1和P2品种来源的组培品系之间;FT16和HP2分别属于不同近等基因系、不同种植地域、GM水稻品系和天然亲本P2,实际遗传关系较远,可见种子内生真菌类群分布受品种、种植地、转基因技术和组织培养技术改良的综合影响。在根据样本聚类的3个大组中,除FP1内生真菌类群和丰度最大单独归为一个大组外,FP2/FT23/FNR25/FNR18/FT16/HP2归为一组,HT16/HT23/HNR18/HP1/HNR25归为另一组,后2个大组近似于种植地域的样本集合,说明种植地域的影响较大。图4
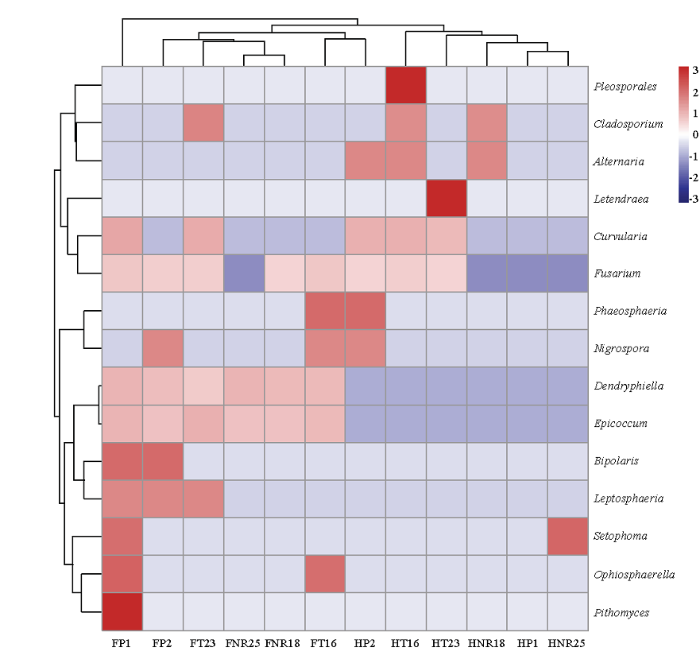 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4海南省(H)和浙江省富阳市(F)种植的6个水稻样本种子内生真菌组成的聚类分析
Fig. 4The cluster analysis of endophytic fungi composition of rice seeds from the 6 samples planted in Hainan (H) and Fuyang, Zhejiang (F)
综合分析两地种植的2个PTN近等基因系共12个样本的变异情况,将变异幅度峰值采取同向取高,反向绝对值相加的计算方法,得出最大变异幅度并根据大小进行排序,真菌总分离频率(IF)排序结果:P1和P2品种两种植地之间变异幅度3.30%—30.58%(30.58%)>P1和P2品种之间-22.32%—4.96%(27.28%)>NR株系变异幅度-21.49%—1.65%(23.14%)>GM株系变异幅度-16.53%—5.79%(22.32%),可见种植地域差异对水稻种子可培养内生真菌分离频率的影响最大,品种影响其次,组培技术的变异效应较小,转基因技术的变异效应最小。从携带内生真菌的总属数变异幅度排序结果:P1和P2品种两种植地之间变异幅度1—9(9)>P1和P2品种之间-3—5(8)=NR株系变异幅度-7—1(8)=GM株系变异幅度-3—5(8),可见在属水平上,种植地域差异对水稻种子可培养内生真菌类群组成的影响最大,其他3个因素的影响较小且相当。
3 讨论
“实质等同性”原则一直是转基因作物安全性评价的重要指标,尚未见基于种子内生菌评估GM作物非预期变异的研究报道。植物内生菌与植物生长发育密切相关,进而可能影响农作物的产量以及食用安全。有研究表明Curvularia lunata和Nigrospora oryzae能抑制胚芽生长,并且减少洋葱根分生组织的有丝分裂指数[13]。本研究发现水稻种子可培养内生真菌绝大多数为子囊菌纲,并且Fusarium为海南省和浙江省富阳市两种植地的优势菌属(图3)。TIAN等[14]也发现Fusarium为稻米中常见菌株。Fusarium是水稻恶苗病的病原体[15],也可能产生威胁食品安全的真菌毒素[16]。本研究发现地域对内生真菌的影响较为明显,H和F两地内生真菌菌群的组成既有共性,又存在一定的差异。Alternaria作为H地特有优势菌株未在F地中分离得到,Dendryphiella和Epicoccum作为在F地特有的优势菌株存在于6个稻米样本中(表2),表明内生真菌的分布具有地域的专化性,优势菌群也随着地理环境的变化而改变。这与李盼盼的研究结果一致[11]。并且F地6个样本中的IF、丰富度指数(D)、多样性指数(H')以及均匀度指数(E)均高于H地,表明F地稻米内生真菌更加丰富,薛庆婉等[17]研究也发现地理环境是影响植物内生真菌组成、丰富度和优势菌群的重要因素。本研究发现H和F地水稻种子内生真菌的组成存在显著性差异,差异的原因可能涉及土壤化学成分组成[14],环境条件和气候的影响[13]。
理论上,GM水稻种子的非预期变异效应包括公认安全的组织培养无性系变异和风险性较大的转基因插入突变,技术来源分别为GM株系培育过程中的组织培养和转基因插入整合。同等条件下,本研究PTN样本系统中NR的性状表现可作为解析GM水稻品系非预期变异技术来源的有效参照[2]。在H和F种植地中,某些菌属表现明显的品种特异性,如Nigrospora为P2特有菌株(表2),原因可能与水稻品种的遗传背景相关,也可能受感染种子从母代到子代的垂直遗传[18]。GM水稻T16和T23种子内生真菌分离频率的变异效应即包括组织培养无性系变异,又包括转基因插入突变,但变异幅度显著小于P1品种的种植地域差异,也小于浙江省富阳市种植地的FP1和FP2的品种间变异(表2)。可推定GM水稻品系种植内生真菌IF的变异幅度在安全范围内。与相应亲本相比,FT16额外增加2个菌属Phaeosphaeria和Nigrospora,HT23与亲本HP2相比增加了1个独有菌属Letendraea helminthicola,FT23比亲本FP2增加了Curvularia和Cladosporium 2个菌属,基于PTN系统分析可知这些内生真菌菌属种类增多的变异均来源于转基因插入突变效应,需要重点关注其安全性。
与前人关于转基因技术对包括水稻在内的植物根际土壤真菌群落、根际细菌群落和根内生真菌群落结构的影响不明显[19,20,21,22]的报道不同,本研究发现转基因技术和组织培养技术均能显著影响水稻种子内生真菌类群的结构。两地种植的PTN系统样本的最大变异幅度排序结果可知种植地域对水稻种子可培养内生真菌丰度和类群组成的影响最大,品种影响次之,组织培养无性系变异效应影响较小,转基因插入突变效应的影响最小。这与前人对GM马铃薯和GM水稻根际微生物群落分布的研究结果类似,与种植地点、土壤类型、植物生长阶段、植物品种相比,转基因插入突变对马铃薯根际细菌群落的影响较小,其中种植地域影响要大的多[22,23,24,25]。结合聚类分析(图4)和P1品种在海南省和浙江省富阳市种植时种子内生真菌多样性和丰富度的极端差异表现(表2),可明确种植地域对水稻种子内生真菌类群的影响最为显著。
4 结论
水稻种子内生真菌丰富多样,可培养内生真菌绝大多数为子囊菌纲。不同种植地域水稻种子内生真菌菌群组成存在差异,Fusarium为海南省和浙江省富阳市两种植地的水稻种子的共有优势内生菌属。转基因技术和组织培养技术均显著影响水稻种子内生真菌类群的结构,其中GM水稻种子内生真菌菌属种类增多的变异主要来源于转基因插入突变效应,需评估其安全性。对水稻种子可培养内生真菌丰度和类群组成的影响大小排序为种植地域>品种差异>组织培养无性系变异效应>转基因插入突变效应。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1469-8137.2007.02350.xURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
DOI:10.5846/stxb201501290233URL [本文引用: 1]

对我国古老特有植物青檀叶片进行内生和附生真菌的研究,以了解青檀叶片内生和附生真菌的组成特点和探讨内生和附生真菌菌群之间的可能联系,为研究真菌资源多样性、植物附生和内生真菌的相互演化关系及真菌与宿主植物协同进化等提供有益参考资料。研究结果表明,从健康的青檀叶片获得可培养内生真菌839株,附生真菌1857株,共计2696株,鉴定其分属于4目,5科,43属。在目的分类水平上,内生和附生真菌均以丛梗孢目Moniliales为优势菌群,分别占90.23%和 92.51%;在科的水平上,内生真菌以暗梗孢科 Dematiaceae和丛梗孢科Moniliaceae为优势菌群,分别占47.56%和42.67%,附生真菌以丛梗孢科Moniliaceae 和暗梗孢科 Dematiaceae为优势菌群,分别占67.04%和25.47%;在属的水平上,内生真菌以黑团孢属Periconia和青霉属Penicillium为优势菌群,分别占31.47%和10.73%,附生真菌以小球霉属Glomerularia、膝葡孢属Gonatobotrys和青霉属Penicillium为优势菌群,分别占20.03%、13.95%和12.22%。青檀叶片内生真菌和附生真菌均存在的菌群数量达到23个属,占53.49%。内生真菌特有的属有6个,共分离19株,占0.70%,附生真菌特有的属有14个,共分离120株,占4.45%。 内生真菌的Shannon-Wiener index (H') 多样性指数(2.44)和Margalef index (R) 丰富度指数(2.88)分别小于附生真菌Shannon-Wiener index (H') 多样性指数(2.57)和Margalef index (R) 丰富度指数(3.32),但两者的Evenness index (E) 均匀度指数几乎相等。青檀叶片内生和附生真菌菌群组成具有较高的相似性,相似性系数达0.70。通过Fisher's exact test分析表明青檀叶片内生和附生真菌菌群组成无明显差异(P=0.072)。
DOI:10.5846/stxb201501290233URL [本文引用: 1]
对我国古老特有植物青檀叶片进行内生和附生真菌的研究,以了解青檀叶片内生和附生真菌的组成特点和探讨内生和附生真菌菌群之间的可能联系,为研究真菌资源多样性、植物附生和内生真菌的相互演化关系及真菌与宿主植物协同进化等提供有益参考资料。研究结果表明,从健康的青檀叶片获得可培养内生真菌839株,附生真菌1857株,共计2696株,鉴定其分属于4目,5科,43属。在目的分类水平上,内生和附生真菌均以丛梗孢目Moniliales为优势菌群,分别占90.23%和 92.51%;在科的水平上,内生真菌以暗梗孢科 Dematiaceae和丛梗孢科Moniliaceae为优势菌群,分别占47.56%和42.67%,附生真菌以丛梗孢科Moniliaceae 和暗梗孢科 Dematiaceae为优势菌群,分别占67.04%和25.47%;在属的水平上,内生真菌以黑团孢属Periconia和青霉属Penicillium为优势菌群,分别占31.47%和10.73%,附生真菌以小球霉属Glomerularia、膝葡孢属Gonatobotrys和青霉属Penicillium为优势菌群,分别占20.03%、13.95%和12.22%。青檀叶片内生真菌和附生真菌均存在的菌群数量达到23个属,占53.49%。内生真菌特有的属有6个,共分离19株,占0.70%,附生真菌特有的属有14个,共分离120株,占4.45%。 内生真菌的Shannon-Wiener index (H') 多样性指数(2.44)和Margalef index (R) 丰富度指数(2.88)分别小于附生真菌Shannon-Wiener index (H') 多样性指数(2.57)和Margalef index (R) 丰富度指数(3.32),但两者的Evenness index (E) 均匀度指数几乎相等。青檀叶片内生和附生真菌菌群组成具有较高的相似性,相似性系数达0.70。通过Fisher's exact test分析表明青檀叶片内生和附生真菌菌群组成无明显差异(P=0.072)。
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOI:10.1023/B:WIBI.0000023843.83692.3fURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2014.18.012URL [本文引用: 1]
脱氧雪腐镰刀菌烯醇(DON),又称呕吐毒素,是由小麦赤霉病菌禾谷镰刀菌复合群(Fusariumgraminearum species complex)产生的单端孢霉烯族毒素,毒素在小麦籽粒中累积。作为B型单端孢霉烯族毒素,DON可以引起呕吐、拒食、腹泻、出血甚至死亡, 对猪的危害尤其严重。近年来,小麦赤霉病在世界各地高频率流行,尤其在中国长江中下游小麦生产区以及黄淮部分小麦产区、美国中西部小麦主产区,导致小麦中DON毒素严重超标,并引发了严重的食品安全问题。本文对国内外小麦中DON毒素的污染现状、产毒镰刀菌种类及其化学型的分布及其趋势、毒素产生的调控机制以及小麦中DON毒素的防控途径进行了论述,希望有助于镰刀菌毒素污染小麦质量安全的风险评估、监管以及科学处置。
DOI:10.3864/j.issn.0578-1752.2014.18.012URL [本文引用: 1]
脱氧雪腐镰刀菌烯醇(DON),又称呕吐毒素,是由小麦赤霉病菌禾谷镰刀菌复合群(Fusariumgraminearum species complex)产生的单端孢霉烯族毒素,毒素在小麦籽粒中累积。作为B型单端孢霉烯族毒素,DON可以引起呕吐、拒食、腹泻、出血甚至死亡, 对猪的危害尤其严重。近年来,小麦赤霉病在世界各地高频率流行,尤其在中国长江中下游小麦生产区以及黄淮部分小麦产区、美国中西部小麦主产区,导致小麦中DON毒素严重超标,并引发了严重的食品安全问题。本文对国内外小麦中DON毒素的污染现状、产毒镰刀菌种类及其化学型的分布及其趋势、毒素产生的调控机制以及小麦中DON毒素的防控途径进行了论述,希望有助于镰刀菌毒素污染小麦质量安全的风险评估、监管以及科学处置。
DOI:DOI: 10.13346/j.mycosystema.140030URL [本文引用: 1]
为认识和掌握常见瓜类植物内生真菌多样性及其变化规律,对从4个地点采集的9种70株健康瓜类植物进行了内生真菌的分离,共获得534个菌株,总体分离率为20.9%。结合形态学和ITS序列分析,鉴定了分属于55个属的455个菌株,有8个属的真菌属于优势类群,相对多度最高的3个属依次为Colletotrichum、Fusarium和Rhizopycnis。Colletotrichum主要存在于植株地上部,Fusarium主要存在于根部和下部茎,Rhizopycnis主要存在于根部,表现出明显的部位差异。河南原阳不同瓜类植物样品中主要内生真菌类群的相对多度和内生真菌的多样性存在明显差异,蛇瓜中Colletotrichum的相对多度达44.7%,显著高于丝瓜中Colletotrichum的相对多度8.0%;黄瓜中Fusarium的相对多度达32.4%,远高于在其他瓜类植物中的相对多度,而在南瓜中Fusarium的相对多度最低,仅为3.0%。江苏无锡和河南原阳的丝瓜植株样品中内生真菌的类群组成有很大不同,表明植株生长的地理环境可能对内生真菌的组成和多样性具有显著影响。
DOI:DOI: 10.13346/j.mycosystema.140030URL [本文引用: 1]
为认识和掌握常见瓜类植物内生真菌多样性及其变化规律,对从4个地点采集的9种70株健康瓜类植物进行了内生真菌的分离,共获得534个菌株,总体分离率为20.9%。结合形态学和ITS序列分析,鉴定了分属于55个属的455个菌株,有8个属的真菌属于优势类群,相对多度最高的3个属依次为Colletotrichum、Fusarium和Rhizopycnis。Colletotrichum主要存在于植株地上部,Fusarium主要存在于根部和下部茎,Rhizopycnis主要存在于根部,表现出明显的部位差异。河南原阳不同瓜类植物样品中主要内生真菌类群的相对多度和内生真菌的多样性存在明显差异,蛇瓜中Colletotrichum的相对多度达44.7%,显著高于丝瓜中Colletotrichum的相对多度8.0%;黄瓜中Fusarium的相对多度达32.4%,远高于在其他瓜类植物中的相对多度,而在南瓜中Fusarium的相对多度最低,仅为3.0%。江苏无锡和河南原阳的丝瓜植株样品中内生真菌的类群组成有很大不同,表明植株生长的地理环境可能对内生真菌的组成和多样性具有显著影响。
DOI:10.17521/cjpe.2015.0008URL [本文引用: 1]
通过田间试验, 研究了垂直传播的Neotyphodium属内生真菌和水平传播的Epichloë属内生真菌对羽茅(Achnatherum sibiricum)生理生态的影响。结果表明: Neotyphodium属内生真菌感染对羽茅的株高和叶长有显著的正效应, 而Epichloë属内生真菌感染对羽茅株高和叶长的增益作用较少。感染Neotyphodium属内生真菌的羽茅最大净光合速率显著高于感染Epichloë属内生真菌的羽茅, Neotyphodium属内生真菌显著提升了羽茅的气孔限制值和水分利用效率, 而实验测定的第一年结果显示: Epichloë属内生真菌对羽茅的各个光合指标具有明显的负效应(胞间CO2浓度除外)。Neotyphodium属内生真菌感染使得宿主植物积累的可溶性糖含量显著高于感染Epichloë属内生真菌的羽茅和不染菌的羽茅植株。感染Epichloë属内生真菌的羽茅中氮含量显著高于感染Neotyphodium属内生真菌的羽茅。总之, 内生真菌的传播方式是影响羽茅生理生态学特性的一个因素, 并且垂直传播的内生真菌对羽茅生长和生理特性的益处高于水平传播的内生真菌。
DOI:10.17521/cjpe.2015.0008URL [本文引用: 1]
通过田间试验, 研究了垂直传播的Neotyphodium属内生真菌和水平传播的Epichloë属内生真菌对羽茅(Achnatherum sibiricum)生理生态的影响。结果表明: Neotyphodium属内生真菌感染对羽茅的株高和叶长有显著的正效应, 而Epichloë属内生真菌感染对羽茅株高和叶长的增益作用较少。感染Neotyphodium属内生真菌的羽茅最大净光合速率显著高于感染Epichloë属内生真菌的羽茅, Neotyphodium属内生真菌显著提升了羽茅的气孔限制值和水分利用效率, 而实验测定的第一年结果显示: Epichloë属内生真菌对羽茅的各个光合指标具有明显的负效应(胞间CO2浓度除外)。Neotyphodium属内生真菌感染使得宿主植物积累的可溶性糖含量显著高于感染Epichloë属内生真菌的羽茅和不染菌的羽茅植株。感染Epichloë属内生真菌的羽茅中氮含量显著高于感染Neotyphodium属内生真菌的羽茅。总之, 内生真菌的传播方式是影响羽茅生理生态学特性的一个因素, 并且垂直传播的内生真菌对羽茅生长和生理特性的益处高于水平传播的内生真菌。
DOI:10.1002/ece3.305URL [本文引用: 1]
DOI:10.1128/AEM.68.3.1325-1335.2002URL [本文引用: 1]
DOI:10.1371/journal.pone.0158974URL [本文引用: 1]
DOI:10.1007/s00248-010-9776-5URL [本文引用: 2]
We assessed the temporal dynamics of bacterial and fungal communities in a soil ecosystem supporting genetically modified (GM) rice (Oryza sativa L., ABC-TPSP; fusion of trehalose-6-phosphate synthase and phosphatase). Using terminal restriction fragment length polymorphism analysis and real-time quantitative PCR, we compared bacterial and fungal communities in the soils underlying GM rice (ABC-TPSP), and its host cultivar (Nakdong) during growing seasons and non-growing seasons. Overall, the soils supporting GM and non-GM rice did not differ significantly in diversity indices, including ribotype numbers, for either bacteria or fungi. The diversity index (H) in both the bacterial and fungal communities was correlated with water content, dissolved organic carbon (DOC), and ammonium nitrogen, and the correlation was stronger in fungi than in bacteria. Multivariate analysis showed no differences in microbial community structures between the two crop genotypes, but such differences did appear in time, with significant changes observed after harvest. Gene copy number was estimated as 10(8)similar to 10(11) and 10(5)similar to 10(7) per gram of soil for bacteria and fungi, respectively. As observed for community structure, the rice genotypes did not differ significantly in either bacterial- or fungal-specific gene copy numbers, although we observed a seasonal change in number. We summarize the results of this study as follows. (1) GM rice did not influence soil bacterial and fungal community structures as compared to non-GM rice in our system, (2) both bacterial and fungal communities changed with the growth stage of either rice genotype, (3) fungal communities were less variable than bacterial communities, and (4) although several environmental factors, including ammonium nitrogen and DOC correlated with shifts in microbial community structure, no single factor stood out.
DOI:10.1111/fem.2006.56.issue-2URL [本文引用: 1]
DOI:10.1371/journal.pone.0033819URL [本文引用: 1]
DOI:10.1016/j.soilbio.2007.09.017URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}