 ,1,3
,1,3Correlation Analysis Between Amino Acids and Fruity Esters During Spine Grape Fermentation
ZHU ZiJian1, CHEN SiYu1, SU Jun2, TAO YongSheng,1,3通讯作者:
责任编辑: 赵伶俐
收稿日期:2019-09-30接受日期:2019-12-16网络出版日期:2020-06-01
| 基金资助: |
Received:2019-09-30Accepted:2019-12-16Online:2020-06-01
作者简介 About authors
朱子健,E-mail: zhuzijian@nwafu.edu.cn。

摘要
关键词:
Abstract
Keywords:
PDF (532KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
朱子健, 陈思雨, 粟俊, 陶永胜. 刺葡萄酒酒精发酵过程中氨基酸组成与果香酯类物质 生成的关联分析[J]. 中国农业科学, 2020, 53(11): 2272-2284 doi:10.3864/j.issn.0578-1752.2020.11.013
ZHU ZiJian, CHEN SiYu, SU Jun, TAO YongSheng.
0 引言
【研究意义】刺葡萄是中国特色野生葡萄,酿造而成的刺葡萄酒呈现深紫色或红宝石色,花青素和总酚含量高,具有很好的抗氧化特性[1]。研究表明,刺葡萄酒香气较弱,品种香气成分及其前体物质含量偏低,发酵形成的果香特征是刺葡萄酒的主要香气特征[2]。因此,刺葡萄酒的增香酿造应以改善发酵香气为主。酯类物质是红葡萄酒中重要的发酵香气成分[3],以乙酸酯和短-中链脂肪酸乙酯为主的果香酯类物质与葡萄酒的果香特征密切相关,通常作为葡萄酒增香酿造的目标产物[4]。葡萄酒发酵过程中果香酯类的形成与氨基酸密切相关,支链氨基酸在酵母的作用下通过Ehrlich途径脱羧、还原,生成相应的高级醇,并进一步转化生成酯和酸[5],从而影响葡萄酒发酵酯类物质的含量。通过调整发酵基质中氨基酸的组成,将有望针对性提高刺葡萄酒果香酯类物质的含量,提高发酵香气质量。但目前为止,针对刺葡萄酒发酵环境中氨基酸及果香酯类物质的研究较少。分析刺葡萄酒精发酵过程中的氨基酸组成与果香酯类物质生成的关联性,将有利于促进基于氨基酸组成调控的刺葡萄酒增香酿造研究的发展,并促进其他果酒增香酿造技术的开拓创新。【前人研究进展】此前,有关刺葡萄酒的研究主要集中在保健价值和功能性成分上,如花色苷和白藜芦醇[6,7],对刺葡萄酒香气匮乏的技术问题仍缺少针对性研究。刺葡萄酒增香酿造的研究起步较晚,本研究团队尝试通过混菌发酵优化刺葡萄酒的发酵香气,并在一定程度上提高了刺葡萄酒中果香酯类物质的含量[8],然而上述研究仍无法确定刺葡萄酒发酵香气淡弱、果香酯类含量低的内在原因。影响葡萄酒发酵香气成分的因素较多。其中,发酵葡萄汁中α-氨基酸的组成对葡萄酒发酵过程中高级醇和酯类物质的生成具有重要影响[9],异亮氨酸、亮氨酸、缬氨酸、苯丙氨酸是高级醇及其相关酯类物质的重要前体[10]。HERNáNDEZ-ORTE等[11]根据11种葡萄品种的氨基酸谱配制模拟葡萄汁进行发酵,发现大量挥发性香气化合物与葡萄汁氨基酸组成相关。PROCOPIO等[12]配置了34种不同氨基酸谱的模拟酒精发酵环境,通过PLS分析建立了理想发酵环境下氨基酸轮廓对主要香气成分影响的指纹图谱,发现缬氨酸、谷氨酸、脯氨酸、亮氨酸、异亮氨酸是影响香气活性成分含量的最重要解释变量。也有研究者通过相关性分析与线性拟合发现在模拟葡萄汁酒精发酵条件下氨基酸含量与挥发性香气成分含量之间存在线性关系[13]。然而,上述研究主要在模拟葡萄汁发酵环境中进行,对真实葡萄酒发酵环境中氨基酸组成及含量与果香酯类物质的关联性研究很少。大多数研究专注于在发酵前补充氨基酸,忽视了对葡萄醪及葡萄发酵过程中氨基酸组成的研究。测定葡萄醪初始状态及发酵过程中的氨基酸组成,明确氨基酸与果香酯类物质的关联,将有利于规避不必要的氨基酸补充,避免可同化氮含量过高导致的负面影响。【本研究切入点】国内对刺葡萄酒增香酿造的研究尚处于起步阶段,有必要从更多角度开发刺葡萄酒香气优化方案。【拟解决的关键问题】以刺葡萄品种‘湘珍珠’为试验对象,研究刺葡萄醪、刺葡萄酒发酵过程中氨基酸及果香酯类物质的组成和含量,并以‘赤霞珠’葡萄为发酵对照组,分析发酵过程中氨基酸组成与果香酯类物质的相关性,对比刺葡萄发酵体系与赤霞珠发酵体系之间上述相关性的差异,初步确定限制刺葡萄酒果香质量的氨基酸营养因素,为刺葡萄酒增香酿造提供理论指导。1 材料与方法
试验于2019年在西北农林科技大学葡萄酒学院酿造与风味实验室进行。1.1 葡萄原料与试验菌种
赤霞珠:于2018年8月采自陕西合阳葡萄基地,含糖量207.5 g?L-1,含酸量5 g?L-1(以酒石酸计)。刺葡萄(湘珍珠):于2018年8月采自湖南怀化,含糖量172.5 g?L-1,初始汁含酸量2.3 g?L-1(以酒石酸计)。
试验菌株:酿酒酵母(S. cerevisiae)ACTIFLORE F33,简称F33,购于法国Laffort公司。
1.2 仪器与试剂
HW.SY21-KP8电热恒温水浴锅(北京市长风仪器仪表公司);TGL-16M台式高速冷冻离心机(长沙高新技术开发区湘仪离心机仪器公司);ME203E电子天平(梅特勒-托利多仪器(上海)有限公司);10RTEX-5旋涡震荡仪(海门市其林贝尔仪器制造有限公司);FE28 pH计(梅特勒-托利多仪器(上海)有限公司);DW-YL270低温冰箱(中科美菱低温科技有限公司);GCMS-QP2020气相色谱-质谱联用仪(岛津公司);Waters 2695高效液相色谱仪、Waters 2996二极管阵列检测器(沃特世科技(上海)有限公司)。分析纯化学试剂:葡萄糖、亚硫酸溶液购于天津市科密欧化学试剂有限公司;五水合硫酸铜、氢氧化钠、氯化钠、盐酸(36%—38%)购于四川西陇科学有限公司;四水合酒石酸钾钠、邻苯二甲酸氢钾、磷酸二氢钾、碳酸氢钠、无水乙酸钠、2, 4-二硝基氟苯(DNFB)。
色谱纯化学试剂及标准品:超纯水,色谱纯乙腈。L-天冬氨酸、L-色氨酸、L-异亮氨酸、L-丙氨酸、L-精氨酸、γ-氨基丁酸、L-蛋氨酸、L-苯丙氨酸、L-酪氨酸、L-缬氨酸购于上海源叶生物科技有限公司;L-谷氨酸、L-脯氨酸、L-亮氨酸购于北京索莱宝科技有限公司。乙酸乙酯、乙酸丁酯、乙酸异丁酯、乙酸异戊酯、乙酸己酯、乙酸辛酯、2-乙酸苯乙酯、丁酸乙酯、异戊酸乙酯、己酸乙酯、庚酸乙酯、辛酸乙酯、癸酸乙酯、月桂酸乙酯、己酸甲酯、己酸异戊酯、辛酸异戊酯购于北京Sigma-Aldrich公司。
定容缓冲液:取KH2PO4 3.4 g于500 mL容量瓶中,加入0.1 mol?L-1 NaOH溶液145.5 mL,加超纯水定容。
1.3 试验方法
1.3.1 葡萄酒酿造 将‘赤霞珠’‘湘珍珠’葡萄进行除梗破碎,分别装入准备好的20 L透明玻璃发酵罐中,每个品种的酿造处理重复2次,每罐取样40 mL作为初始样品。加入60 mg?L-1 SO2并在4℃下浸渍24 h,随后向发酵罐中加入酿酒酵母,转移至20℃恒温箱中启动酒精发酵,每隔24 h取样,每次取样40 mL。残糖小于2 g?L-1后结束发酵[14],并取样40 mL作为最终酒样。样品置于-20℃冰箱中保存。1.3.2 氨基酸分析 氨基酸衍生方法:使用移液枪吸取经0.22 μm滤膜过滤后的样品或标准品溶液0.1 mL,加入1 mL离心管中,加0.05 mol?L-1 NaHCO3溶液0.1 mL后摇匀,再加入5% DNFB-乙腈溶液40 μL并摇匀。将离心管置于60℃恒温水浴锅内,水浴加热60 min后取出,置于避光环境下冷却至室温。随后加入定容缓冲液0.76 mL并摇匀,于黑暗处放置15 min后进行色谱分离[15]。
氨基酸色谱分析:色谱柱为C18分离柱(Agilent SB-C18色谱柱;Analytical 4.6 mm×250 mm;5-Micron);柱温33℃,进样量20 μL,采用四元梯度分析;流动相总流量为1 mL?min-1;二极管阵列扫描波长:320—400 nm。流动相A为色谱纯乙腈,流动相B为超纯水,流动相C为pH 6.4的乙酸钠溶液。色谱柱洗脱程序如下:初始8% A,8% B,84% C;0—2 min:8%—15% A,8%—15% B,84%—70% C;2—4 min:15%—17% A,15%—17% B,70%—66% C;4—8 min:17%—20% A,17%—20% B,66%—60% C;8—14 min:20%—21% A,20%—22% B,60%—57% C;14—24 min:21%—28% A,22%—27% B,57%—45% C;24—27 min:28%—28% A,27%—27% B,45%—45% C;27—36 min:28%—49% A,27%—49% B,45%—2% C;36—40 min:49%—8% A,49%—8% B,2%—84% C。
定性定量方法:通过氨基酸单标溶液与混标溶液的色谱分析结果,根据标准品保留时间进行定性。将混标溶液梯度稀释后,通过外标法定量,标准曲线采用5点法绘制。
1.3.3 香气物质分析 SPME样品处理:本试验香气物质的提取采用顶空固相微萃取(HS-SPME)的方法,使用DVB/CAR/PDMS萃取纤维(50/30 μm涂层厚度,2 cm可伸缩长度)及SPME 57330-U联用手柄(Supelco, Bellefonte PA, USA)。20 mL顶空瓶中加入8 mL酒样,2.0 g氯化钠,2-辛醇(内标,400 μg?L-1)和搅拌子在40℃水浴中平衡15 min后,再用萃取纤维在40℃下搅拌吸附30 min后取出,立即手动进样至气相色谱进样口(230℃)解析5 min。每个酒样的萃取操作重复2次。
GC-MS分析:使用气相色谱-质谱联用仪(岛津GCMS-QP2020),柱型号为DB-WAX(60 m×0.25 mm,涂层厚度0.25 μm,Agilent JandW,USA)。不分流进样,载气为高纯氦气(99.999%),载气流速为1.5 mL?min-1。程序升温:以3℃?min-1从40℃至130℃,再以4℃?min-1从130℃至250℃,保持8 min。进样口温度230℃,连接杆温度220℃,离子源温度200℃,电子电源电压70 eV,质谱为电离轰击(EI)模式,全扫描质谱范围25—350 amu,扫描频率0.2 s/次。
定性定量分析:配制酯类物质标准品的模拟酒溶液,进行上述HS/SPME-GC-MS分析,采用标准品保留时间对比、保留指数对比和NIST 17质谱谱库查询进行目标酯类物质的定性。采用内标-标准曲线法对目标酯类物质进行定量,2-辛醇为内标物,具体定量方法参考HU等[16]的方法。
1.4 数据处理
数据分析采用Pearson相关性分析(Pearson Correlation Analysis)对发酵过程中氨基酸与果香酯类物质进行关联性分析,试验数据处理主要使用SPSS 20.0(SPSS Inc., Chicago, IL, USA)软件。使用OriginPro 9.0绘制发酵过程中氨基酸含量变化趋势图。2 结果
2.1 葡萄酒精发酵过程中氨基酸的变化
本研究通过HPLC-PAD对刺葡萄和赤霞珠葡萄醪样品和发酵汁样品中的13种氨基酸成分进行定性定量分析(表1)。刺葡萄醪总氨基酸含量显著低于‘赤霞珠’葡萄,仅为‘赤霞珠’葡萄醪中总氨基酸含量的33%,但刺葡萄醪除脯氨酸外的氨基酸总量则显著高于‘赤霞珠’,约为‘赤霞珠’葡萄醪的3倍。除脯氨酸外,刺葡萄中的丙氨酸、苯丙氨酸、异亮氨酸含量显著低于‘赤霞珠’,仅为‘赤霞珠’葡萄醪中上述氨基酸含量的10%、35%和20%。刺葡萄中天冬氨酸、缬氨酸、谷氨酸、γ-氨基丁酸、精氨酸的含量则显著高于‘赤霞珠’,分别达到‘赤霞珠’葡萄醪中上述氨基酸含量的2.4倍、2.2倍、4.2倍、7.7倍、3倍。‘赤霞珠’葡萄与刺葡萄的氨基酸组成及含量具有明显差异。亮氨酸、色氨酸和蛋氨酸在两种葡萄醪样品中均未检出。Table 1
表1
表1葡萄醪中的氨基酸含量
Table 1
| 氨基酸 Amino acid | 赤霞珠葡萄 Cabernet Sauvignon grape | 刺葡萄 Spine grape |
|---|---|---|
| 天冬氨酸 Asp | 11.7±2.6b | 28.1±4a |
| 谷氨酸 Glu | 14.9±0.2b | 63.4±18a |
| 精氨酸 Arg | 273.2±1.5b | 814.7±33.8a |
| 脯氨酸 Pro | 10868.6±108a | 1316.4±79.4b |
| γ-氨基丁酸 GABA | 59.9±0.5b | 463±3.2a |
| 丙氨酸 Ala | 48.1±4.4a | 5±0.1b |
| 缬氨酸 Val | 14.3±0b | 31.6±0.6a |
| 蛋氨酸 Met | ND | ND |
| 异亮氨酸 Ile | 37.6±2.5a | 7.56±0.5b |
| 亮氨酸 Leu | ND | ND |
| 色氨酸 Trp | ND | ND |
| 苯丙氨酸 Phe | 15.4±0.9a | 5.4±1b |
| 酪氨酸 Tyr | 21.1±0.4a | 16.9±1.2ab |
| 总氨基酸 Total amino acids | 11350.4±100a | 3727.8±59.8b |
| 总氨基酸(除脯氨酸) Total amino acids (except Pro) | 481.8±11b | 2411.4±52.9a |
新窗口打开|下载CSV
发酵过程中氨基酸含量变化如图1所示。刺葡萄发酵体系中的天冬氨酸、谷氨酸、苯丙氨酸含量在发酵第1天有所升高。随着酒精发酵的进一步进行,除酪氨酸外,发酵酒样的氨基酸含量逐渐减少。酪氨酸的变化具有品种差异性,其含量在‘赤霞珠’发酵过程中略有下降,而在刺葡萄发酵中则有所增加。刺葡萄最终酒样中的酪氨酸含量高于‘赤霞珠’酒样。脯氨酸是两种葡萄发酵过程中含量最高的氨基酸。亮氨酸、色氨酸及蛋氨酸在两种葡萄发酵全程均未检出。
图1
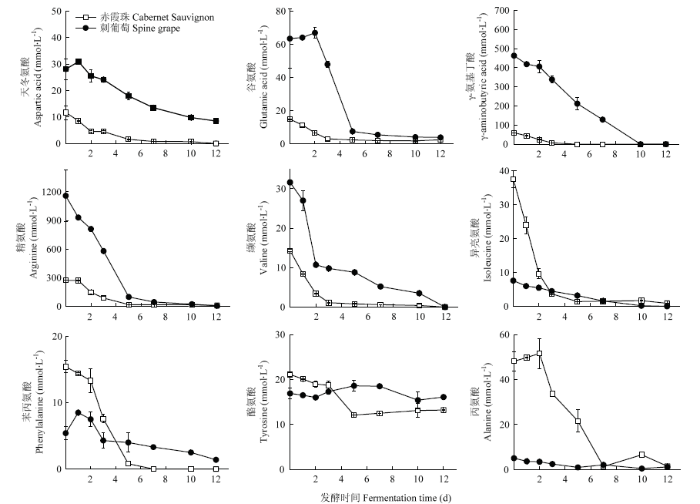 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1酒精发酵过程中氨基酸含量的变化
Fig. 1Evolution of amino acids during alcohol fermentation
2.2 葡萄酒精发酵过程中果香酯类物质的变化
经HS/SPME-GC-MS对发酵葡萄酒样品中香气成分的定性定量分析,本研究在刺葡萄酒和赤霞珠葡萄酒发酵体系中共检出17种酯类物质,包括7种乙酸酯、2种短链脂肪酸乙酯(SCFAEE,C2—C5)、5种中链脂肪酸乙酯(MCFAEE,C6—C13)和3种其他酯类物质,其中6种酯类物质的OAV>1(OAV:气味活性值,OAV=香气化合物浓度/该香气化合物气味阈值),包括乙酸乙酯、乙酸异戊酯、丁酸乙酯、异戊酸乙酯、己酸乙酯、辛酸乙酯。刺葡萄和‘赤霞珠’发酵体系中果香酯类含量变化分别见表2、表3。在刺葡萄发酵体系中,乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯含量始终低于‘赤霞珠’,在最终酒样内的含量分别为16 233、132.4和711.9 μg?L-1,仅为‘赤霞珠’最终酒样中乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯含量的55%、11%和69%。其他酯类含量在刺葡萄发酵过程中呈先增加后减少的变化趋势,在发酵第5天、第7天和第10天时含量高于‘赤霞珠’,而在发酵结束时,刺葡萄酒中其他酯类含量略低于‘赤霞珠’葡萄酒。从总体上看,刺葡萄酒中的果香酯类(乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯)含量显著低于‘赤霞珠’葡萄酒,尤其是对葡萄酒果香特征具有突出贡献的中链脂肪酸乙酯含量远远低于‘赤霞珠’葡萄酒。Table 2
表2
表2刺葡萄酒精发酵过程中酯类物质的含量
Table 2
| 化合物 Compound | 葡萄醪 Must | 1 d | 2 d | 3 d | 5 d | 7 d | 10 d | 12 d | 嗅觉阈值 Threshold (μg?L-1) | 气味描述 Odor description | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 乙酸酯 Acetate | 2352.3±0.1 | 1861.9±17.6 | 2914.9±105.4 | 3608.9±7.7 | 8826.9±567.5 | 11450.5±1056.8 | 13251.9±499.8 | 16233.0±129.2 | |||
| 1 | 乙酸乙酯 Ethyl acetate | 2352.3±0.1 | 1861.9±17.6 | 2913.8±105.5 | 3608.1±7.7 | 8644.5±526.1 | 11383.7±1051.5 | 13204.6±499.2 | 16168.6±117.5 | 7500[4] | 菠萝,果香,甜香 Pineapple, fruity, sweet |
| 2 | 乙酸异丁酯 Isobutyl acetate | ND | ND | ND | ND | 18.7±0.3 | 7.4±0.5 | ND | 0.4±0.1 | 1600[4] | 果香,梨,香蕉 Fruity, pear, banana |
| 3 | 乙酸异戊酯Isoamyl acetate | ND | ND | ND | ND | 161.4±41.5 | 57.6±5.5 | 46.0±0.5 | 63.1±11.7 | 30[17] | 果香,香蕉Fruity, banana |
| 4 | 乙酸丁酯 Butyl acetate | ND | ND | 1.0±0.1 | 0.8±0.1 | 1.6±0.2 | 1.3±0.2 | 1.1±0.1 | 1.1±0.2 | 1600[4] | 果香,梨,香蕉 Fruity, pear, banana |
| 5 | 乙酸己酯Hexyl acetate | ND | ND | ND | ND | ND | ND | ND | ND | 1500[17] | 梨,甜果Pear, sweet fruit |
| 6 | 乙酸辛酯Octyl acetate | ND | ND | ND | ND | 0.7±0.1 | 0.4±0.2 | 0.3±0.1 | ND | NF | |
| 7 | 2-乙酸苯乙酯 2-phenylethyl acetate | ND | ND | ND | 8.1±0.1 | 12.3±0.4 | 17.4±2.0 | 20.4±0.9 | 20.9±0.1 | 250[18] | 花香Floral |
| 短链脂肪酸乙酯 SCFAEEs | ND | ND | ND | 2.6±0.2 | 66.2±8.0 | 90.3±4.8 | 106.1±6.5 | 132.4±3.6 | |||
| 1 | 丁酸乙酯 Ethyl butyrate | ND | ND | ND | ND | 59.2±7.8 | 85.0±4.5 | 101.8±6.4 | 127.5±3.3 | 20[4] | 草莓,苹果,香蕉 Strawberry, apple, banana |
| 2 | 异戊酸乙酯 Ethyl isovalerate | ND | ND | ND | 2.6±0.2 | 7.0±0.2 | 5.3±0.3 | 4.3±0.1 | 4.9±0.3 | 3[19] | 香蕉,甜果 Banana, sweet fruity |
| 中链脂肪酸乙酯 MCFAEEs | ND | ND | 35.2±1.0 | 231.7±23.8 | 1263.0±99.7 | 933.1±33.4 | 711.4±22.7 | 711.9±13.5 | |||
| 1 | 己酸乙酯 Ethyl hexanoate | ND | ND | ND | 73.6±12.5 | 707.3±73.7 | 531.9±11.8 | 414.9±21.5 | 429.2±9.3 | 5[20] | 花香,果香,草莓 Floral, fruity, strawberry |
| 2 | 庚酸乙酯Ethyl heptanoate | ND | ND | ND | ND | 0.9±0.1 | 0.7±0.3 | 0.6±0.1 | 0.5±0.1 | 220[21] | 白兰地,果香Brandy, fruity |
| 3 | 辛酸乙酯 Ethyl octanoate | ND | ND | 35.0±1.0 | 132.9±10.0 | 485.3±22.1 | 373.2±18.6 | 295.6±1.2 | 282.0±4.2 | 5[4] | 苹果,梨,花香 Apple, pear, floral |
| 4 | 癸酸乙酯 Ethyl caprate | ND | ND | 0.2±0.1 | 0.4±0.1 | 0.8±0.1 | 0.5±0.1 | 0.3±0.1 | 0.2±0.1 | 200[4] | 蜡味,果香,玫瑰 Waxy, fruity, rose |
| 5 | 月桂酸乙酯 Ethyl dodecanoate | ND | ND | ND | 24.8±1.3 | 68.7±3.9 | 26.8±2.6 | ND | ND | 1500[19] | 甜香,花香,果香,奶油 Sweet, floral, fruity, cream |
| 其他酯类other esters | ND | ND | 6.3±0.3 | 8.0±0.2 | 65.1±2.2 | 33.5±3.7 | 11.7±0.3 | 6.9±0.1 | |||
| 1 | 己酸甲酯Methyl caproate | ND | ND | 6.3±0.3 | 7.9±0.1 | 44.1±0.7 | 17.9±1.4 | ND | ND | NF | |
| 2 | 己酸异戊酯Isoamyl hexanoate | ND | ND | ND | 0.1±0.1 | 6.5±0.5 | 4.0±0.2 | 2.4±0.1 | 1.5±0.1 | NF | 苹果,菠萝Apple, pineapple |
| 3 | 辛酸异戊酯Isoamyl octanoate | ND | ND | ND | ND | 14.5±1.0 | 11.6±2.1 | 9.3±0.2 | 5.4±0.1 | 125[22] | 甜香Sweat |
新窗口打开|下载CSV
Table 3
表3
表3赤霞珠酒精发酵过程中酯类物质的含量
Table 3
| 化合物 Compound | 发酵日程 Fermentation time | 嗅觉阈值 Threshold (μg?L-1) | 气味描述 Odor description | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 葡萄醪Must | 1 d | 2 d | 3 d | 5 d | 7 d | 10 d | 12 d | ||||
| 乙酸酯Acetate | 1846.1±2.5 | 1951.8±35.4 | 3447.1±240.9 | 9036.5±1418.0 | 26065.4±1197.7 | 26151.0±105.7 | 28865.8±594.1 | 29631.1±1381.0 | |||
| 1 | 乙酸乙酯 Ethyl acetate | 1846.0±2.5 | 1951.8±35.4 | 3437.4±240.8 | 8608.2±1403.7 | 24509.4±827.1 | 25681.0±69.4 | 26713.2±508.6 | 27764.9±1252.0 | 7500[4] | 菠萝,果香,甜香 Pineapple, fruity, sweet |
| 2 | 乙酸异丁酯 Isobutyl acetate | ND | ND | ND | 59.5±12.5 | 100.4±11.4 | 62.0±2.6 | 37.1±9.0 | 0.8±0.1 | 1600[4] | 果香,梨,香蕉 Fruity, pear, banana |
| 3 | 乙酸异戊酯 Isoamyl acetate | ND | ND | ND | 345.2±0.1 | 1383.3±355.5 | 306.7±19.0 | 1994.3±74.9 | 1753.5±127.5 | 30[17] | 果香,香蕉 Fruity, banana |
| 4 | 乙酸丁酯 Butyl acetate | 0.1±0.1 | ND | 2.1±0.1 | 5.0±0.6 | 8.3±0.5 | 8.3±0.1 | 8.3±0.2 | 6.4±0.3 | 1600[4] | 果香,梨,香蕉 Fruity, pear, banana |
| 5 | 乙酸己酯 Hexyl acetate | ND | ND | ND | ND | ND | 0.4±0.1 | 0.7±0.1 | 0.6±0.1 | 1500[17] | 梨,甜果 Pear, sweet fruit |
| 6 | 乙酸辛酯 Octyl acetate | ND | ND | ND | 0.3±0.1 | 0.8±0.1 | 0.6±0.1 | 0.4±0.1 | 0.3±0.1 | NF | |
| 7 | 2-乙酸苯乙酯 2-phenylethyl acetate | ND | ND | 7.6±0.1 | 18.3±1.1 | 63.2±3.1 | 92.0±14.6 | 111.8±1.4 | 104.6±1.2 | 250[18] | 花香 Floral |
| 短链脂肪酸乙酯 SCFAEEs | ND | ND | 12.7±0.1 | 752.2±146.9 | 2293.4±167.3 | 1683.0±79.6 | 1279.5±30.0 | 1253.4±83.5 | |||
| 1 | 丁酸乙酯 Ethyl butyrate | ND | ND | ND | 28.9±6.7 | 111.7±5.3 | 127.3±4.2 | 138±5.0 | 143.6±10.0 | 20[4] | 草莓,苹果,香蕉 Strawberry, apple, banana |
| 2 | 异戊酸乙酯 Ethyl isovalerate | ND | ND | ND | 440.2±92.8 | 1146.5±77.0 | 841.7±14.9 | 638.8±19.3 | 530.7±40.5 | 3[19] | 香蕉,甜果 Banana, sweet fruity |
| 中链脂肪酸乙酯 MCFAEEs | ND | ND | 12.1±0.5 | 715.3±139.4 | 2149.7±159.3 | 1492.9±95.3 | 1058.5±27.2 | 1038.0±69.7 | |||
| 1 | 己酸乙酯 Ethyl hexanoate | ND | ND | ND | 440.2±92.8 | 1146.5±77.0 | 841.7±14.9 | 638.8±19.3 | 530.7±40.5 | 5[20] | 花香,果香,草莓 Floral, fruity, strawberry |
| 2 | 庚酸乙酯 Ethyl heptanoate | ND | ND | 0.2±0.1 | 0.8±0.1 | 3.2±0.2 | 2.4±0.3 | 1.9±0.1 | 2.1±0.2 | 220[21] | 白兰地,果香 Brandy, fruity |
| 3 | 辛酸乙酯 Ethyl octanoate | ND | ND | 11.9±0.5 | 238.8±33.6 | 846.9±74.2 | 587.5±68.1 | 417.6±7.9 | 505.0±29.0 | 5[4] | 苹果,梨,花香 Apple, pear, floral |
| 4 | 癸酸乙酯 Ethyl caprate | ND | ND | ND | 0.6±0.1 | 0.8±0.1 | 0.5±0.1 | 0.2±0.1 | 0.2±0.1 | 200[4] | 蜡味,果香,玫瑰 Waxy, fruity, rose |
| 5 | 月桂酸乙酯 Ethyl dodecanoate | ND | ND | ND | 34.9±12.8 | 152.3±7.8 | 60.8±11.9 | ND | ND | 1500[19] | 甜香,花香,果香,奶油 Sweet, floral, fruity, cream |
| 其他酯类 other esters | ND | ND | 14.4±0.5 | 31.6±4.4 | 31.5±3.5 | 13.8±5.4 | 6.9±0.2 | 9.0±0.2 | |||
| 1 | 己酸甲酯Methyl caproate | ND | ND | 14.2±0.5 | 25.8±3.4 | 6.8±1.5 | 2.0±1.2 | ND | ND | NF | |
| 2 | 己酸异戊酯 Isoamyl hexanoate | ND | ND | 0.2±0.1 | 1.4±0.3 | 7.7±0.7 | 4.0±0.7 | 1.5±0.2 | 1.7±0.3 | NF | 苹果,菠萝 Apple, pineapple |
| 3 | 辛酸异戊酯 Isoamyl octanoate | ND | ND | ND | 4.4±0.7 | 17.0±1.3 | 7.8±3.5 | 5.4±0.2 | 7.3±0.1 | 125[22] | 甜香 Sweat |
新窗口打开|下载CSV
2.3 葡萄发酵过程中氨基酸与果香酯类的相关性分析
由于酪氨酸在发酵过程中的变化具有品种特异性,且在刺葡萄发酵中具有增加趋势,因此无法判断其消耗情况,不将其列入相关性分析对象。为确定发酵过程中的氨基酸含量变化是否与果香酯类物质含量变化具有明显关联性,以及两种葡萄发酵过程中上述关联性是否存在差异,本试验对葡萄发酵过程中氨基酸含量与果香酯类含量进行了Pearson相关性分析,如表4所示。发酵过程中大多数氨基酸与果香酯类变化趋势相反,因此结果以负相关为主。总氨基酸含量(除脯氨酸)与果香酯类物质的相关性分析表明,葡萄酒发酵过程中总氨基酸与乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯的相关性系数较高,但与其他酯类物质的相关性系数较低。对比刺葡萄与‘赤霞珠’对照组中总氨基酸含量与果香酯类含量的Pearson相关性系数可见,‘赤霞珠’中的总氨基酸含量(除脯氨酸)与乙酸酯、短链和中链脂肪酸乙酯含量具有极强相关性,分别达到0.919、0.875和0.825;刺葡萄发酵组的总氨基酸与乙酸酯(0.873)、短链脂肪酸乙酯(0.864)、中链脂肪酸乙酯(0.750)的相关性系数低于‘赤霞珠’葡萄发酵组的上述相关性系数。Table 4
表4
表4酒精发酵过程中氨基酸与发酵酯类物质的相关性系数
Table 4
| 品种 Variety | 化合物 Compound | 总氨基酸 Total amino acid | 天冬氨酸 Asp | 谷氨酸 Glu | 精氨酸 Arg | γ-氨基丁酸 GABA | 缬氨酸 Val | 丙氨酸 Ala | 异亮氨酸 Ile | 苯丙氨酸 Phe |
|---|---|---|---|---|---|---|---|---|---|---|
| 赤霞珠 Cabernet Sauvignon | 乙酸酯 Acetates | -0.919 | -0.865 | -0.810 | -0.910 | -0.848 | -0.750 | -0.946 | -0.764 | -0.978 |
| 短链脂肪酸乙酯 SCFAEEs | -0.875 | -0.831 | -0.754 | -0.858 | -0.791 | -0.695 | -0.961 | -0.705 | -0.945 | |
| 中链脂肪酸乙酯 MCFAEEs | -0.825 | -0.736 | -0.762 | -0.833 | -0.796 | -0.689 | -0.733 | -0.716 | -0.876 | |
| 其他酯类 Other esters | -0.505 | -0.419 | -0.612 | -0.541 | -0.593 | -0.591 | -0.139 | -0.607 | -0.378 | |
| 刺葡萄 Spine grape | 乙酸酯 Acetates | -0.873 | -0.927 | -0.889 | -0.888 | -0.976 | -0.821 | -0.837 | -0.738 | -0.694 |
| 短链脂肪酸乙酯 SCFAEEs | -0.864 | -0.929 | -0.913 | -0.886 | -0.970 | -0.759 | -0.805 | -0.791 | -0.661 | |
| 中链脂肪酸乙酯 MCFAEEs | -0.750 | -0.721 | -0.892 | -0.869 | -0.745 | -0.669 | -0.794 | -0.552 | -0.569 | |
| 其他酯类 Other esters | -0.399 | -0.331 | -0.571 | -0.558 | -0.298 | -0.500 | -0.370 | -0.178 | -0.216 |
新窗口打开|下载CSV
通过8种氨基酸含量与果香酯类物质含量的相关性分析可见,在‘赤霞珠’发酵体系中,天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸与乙酸酯、短链和中链脂肪酸乙酯具有强相关性或极强相关性,相关性系数范围为0.689—0.910;丙氨酸、苯丙氨酸、异亮氨酸与上述果香酯类的相关性系范围为0.705—0.978,与果香酯类物质具有强相关性或极强相关性,其中苯丙氨酸与乙酸酯相关性系数最高,丙氨酸与短链脂肪酸乙酯相关性系数最高。在刺葡萄发酵体系的相关性分析中,天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸与乙酸酯、短链和中链脂肪酸乙酯具有强相关性或极强相关性,相关性系数在0.669—0.976;丙氨酸、苯丙氨酸、异亮氨酸与上述果香酯类物质的相关性系数在0.552—0.837。
将刺葡萄发酵过程中氨基酸与果香酯类物质含量的相关性系数与‘赤霞珠’对比可见,在发酵过程中,刺葡萄的精氨酸、天冬氨酸、谷氨酸、γ-氨基丁酸、缬氨酸与果香酯类的相关性系数较‘赤霞珠’偏高。与上述情况相反,刺葡萄发酵过程中的苯丙氨酸与乙酸酯、各类脂肪酸乙酯的相关性系数低于‘赤霞珠’葡萄,异亮氨酸与乙酸酯、中链脂肪酸乙酯的相关性低于‘赤霞珠’葡萄,丙氨酸与乙酸酯和短链脂肪酸乙酯的相关性低于‘赤霞珠’葡萄,上述相关性系数较‘赤霞珠’葡萄发酵过程中对应的相关性系数偏低3%—54%。为直观展现‘赤霞珠’与刺葡萄发酵过程中丙氨酸、苯丙氨酸、异亮氨酸与果香酯类物质的相关性差异,对上述氨基酸含量变化与各类果香酯类物质的含量变化进行线性拟和,拟合结果如图2所示。通过线性拟和分析可以发现,‘赤霞珠’发酵体系中丙氨酸、苯丙氨酸、异亮氨酸与果香酯类的线性拟合效果较刺葡萄发酵体系存在明显差异,刺葡萄发酵体系中的上述氨基酸与果香酯类物质的线性拟和效果较差。在果香酯类物质含量分析中,‘赤霞珠’酒精发酵产生的乙酸酯和短-中链脂肪酸乙酯含量均高于刺葡萄酒,表明不同葡萄品种的发酵酒中果香酯类物质含量的差异可能与氨基酸-果香酯类的相关性差异具有一定关联。结合氨基酸含量研究可见,氨基酸的含量差异与相关性差异具有一定规律性:刺葡萄中精氨酸、天冬氨酸、谷氨酸、γ-氨基丁酸、缬氨酸含量高于‘赤霞珠’,丙氨酸、苯丙氨酸、异亮氨酸含量低于‘赤霞珠’。上述结果表明,刺葡萄中丙氨酸、苯丙氨酸、异亮氨酸含量偏低与刺葡萄酒果香酯类物质含量偏低具有一定关联,刺葡萄发酵过程中的氨基酸组成与果香酯类物质的生成之间具有“木桶效应”,刺葡萄中含量偏低的丙氨酸、苯丙氨酸、异亮氨酸可能限制了果香酯类物质在发酵过程中的合成,进而导致最终酒样中果香酯类含量的差异。
图2
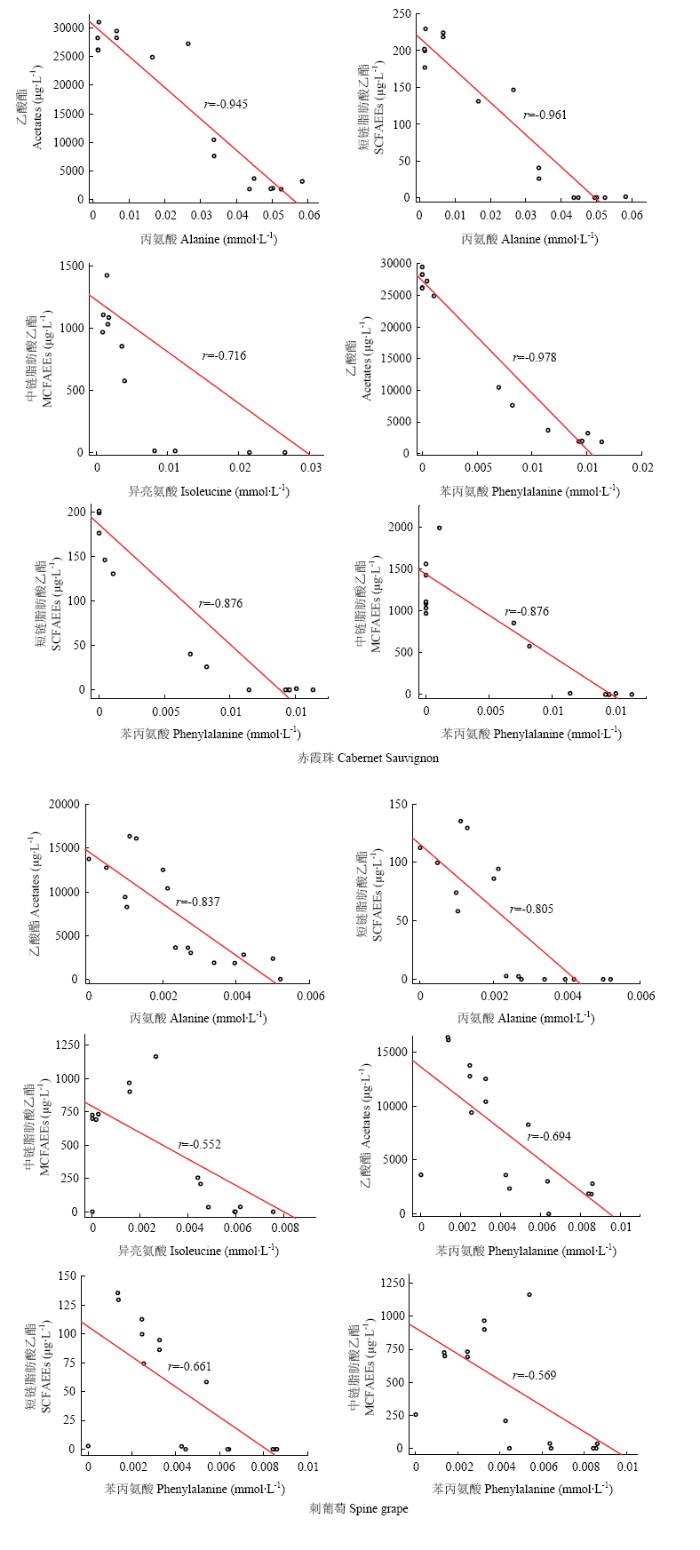 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2酒精发酵过程中部分氨基酸与酯类物质的线性拟合模型
Fig. 2Linear regression of part of amino acids and esters during alcohol fermentation
此外,刺葡萄中除脯氨酸外的总氨基酸含量高于‘赤霞珠’葡萄,但其发酵酒中的果香酯类含量仍然低于‘赤霞珠’葡萄酒,这表明不能将果香酯类合成的有机氮营养因素单一地归结于酵母可利用的总氨基酸含量或某种氨基酸含量的多少,发酵过程中发酵基质的氨基酸组成对果香酯类含量具有更加重要的影响,良好的氨基酸组成可能更有利于果香酯类物质的生成。
3 讨论
3.1 刺葡萄酒精发酵过程中氨基酸、果香酯类物质的含量变化规律
氨基酸存在于葡萄果实,经过葡萄浸渍后进入发酵体系中从而被酵母利用,参与葡萄酒中的发酵香气的合成。葡萄果实中氨基酸组成与葡萄品种关系密切[23],而其测定结果也根据测定方法不同而存在较大差异[24],本研究未在葡萄醪及发酵汁样品中检出亮氨酸,这与此前部分研究结果不同,推测可能受到特定年份葡萄种植条件及环境的影响。GUTIéRREZ- GAMBOA等[25]对智利莫莱谷产区的‘佳丽酿’葡萄醪氨基酸含量进行测定,其中色氨酸在多个样品中含量偏低,且蛋氨酸含量最低仅为0.14 mg?L-1;高年发等[26]对宁夏4个产区的葡萄醪样品进行氨基酸测定,7种被测样品中仅有3种检出色氨酸,且均未检出蛋氨酸。上述研究表明葡萄醪中色氨酸、蛋氨酸含量极低,而本研究未在刺葡萄与‘赤霞珠’葡萄发酵体系中检出色氨酸与蛋氨酸。目前为止尚无对刺葡萄醪及刺葡萄发酵过程中氨基酸含量的研究。本研究发现,在发酵第1天时天冬氨酸、谷氨酸、苯丙氨酸含量略有升高,这与高年发等[26]对浸渍前期氨基酸含量变化的研究结果相似,推测是在浸渍过程中,存在于果皮、果肉和果籽中的部分天冬氨酸、谷氨酸和苯丙氨酸浸出至发酵葡萄汁所致,但尚需更多研究予以证明。在刺葡萄和‘赤霞珠’对照组的酒精发酵过程中,天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸、异亮氨酸、苯丙氨酸含量在发酵过程中呈下降趋势,部分氨基酸最终被完全消耗。酪氨酸含量在刺葡萄发酵过程中逐渐下降,而在‘赤霞珠’中呈现波动变化,具有明显的品种差异性,这与高年发等[26]对不同产区、多个葡萄品种的研究结果基本一致,但与GARDE- CERDáN等[27]的研究结果不同。刺葡萄中天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸含量高于‘赤霞珠’,而苯丙氨酸、丙氨酸、异亮氨酸含量低于‘赤霞珠’。由于脯氨酸通常不被视为酵母可同化氮[28],因此本文不再讨论脯氨酸的含量变化及其与果香酯类物质的相关性情况。葡萄酒中的酯类物质对香气的贡献曾被长期低估,而如今有更多的研究证明其对香气质量的贡献不可忽视[29]。目前为止,对刺葡萄醪及刺葡萄发酵过程中香气物质的研究较少,鲍瑞峰[30]对刺葡萄果实及刺葡萄酒中部分果香酯类成分进行了研究,但其未对香气物质含量进行定量分析;本研究团队在前期研究中对刺葡萄酒中香气物质含量进行了定量测定,但未对发酵过程中的香气物质含量变化进行全程监控,且未将刺葡萄酒的香气物质组成与其他葡萄酒进行对比分析[8]。本研究对刺葡萄及‘赤霞珠’葡萄发酵过程中果香酯类物质含量进行测定,发现刺葡萄酒发酵过程中果香酯类物质含量始终低于‘赤霞珠’,其中乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯含量显著低于‘赤霞珠’,这可能是导致刺葡萄酒香气贫乏的直接原因。
3.2 刺葡萄酒发酵体系中氨基酸与果香酯类物质生成的关联
氨基酸组成与葡萄品种相关,通过葡萄发酵,这些氨基酸转化产生特异性的香气物质,形成酒的特异性香气,从而形成不同风格的葡萄酒[31]。氨基酸经过微生物分解代谢产生的香气物质具有一定对应性,支链氨基酸(Leu、Ile、Val)被转化为具有麦芽味、果香和汗味的香气化合物;芳香族氨基酸(Phe、Tyr、Trp)的分解代谢产生具有花香、化学溶剂味和粪便气味的化合物;天冬氨酸(Asp)被分解成具有黄油味的香气物质,含硫氨基酸(Met、Cys)被转化为具有煮白菜、肉味和大蒜味的化合物[32]。在酒精发酵过程中,氨基酸主要通过Ehrlich途径进行代谢,通过转氨、脱羧等过程生成对应的高级醇及脂肪酸[33],并进一步反应生成酯类物质。然而,更多研究表明在实际发酵环境中,氨基酸对香气物质生成的影响并不是严格一一对应的,酵母可能将氨基酸代谢过程中产生的α-酮酸重新分配至其他类似的生物化学反应途径[13]。因此,在具有多种可同化氮来源的复杂发酵基质中,某种氨基酸代谢的中间产物可能同时参与多种香气物质的合成,从而对化学结构相似的多种香气物质的含量产生影响。对于氨基酸与香气的研究不是一蹴而就的,研究者们将氨基酸归于氮源,又将氮源分为无机氮和氨基酸进行添加试验。HERNáNDEZ-ORTE等[34]根据11种葡萄品种的氨基酸谱配制模拟葡萄汁,进行模拟发酵,发现一些挥发性化合物(乙酸乙酯、高级醇、异丁酸、己酸、辛酸)存在显著差异,进行多元回归分析后发现,这些挥发性化合物与葡萄汁氨基酸组成相关。GARDE-CERDáN等[35]在氮缺乏葡萄醪中进行氨基酸补充试验发现,总酯、乙酸异戊酯、乙酸苯乙酯与添加的氨基酸量成正比;PROCOPIO等[12]通过对不同氨基酸谱的模拟汁发酵及统计学分析,发现谷氨酸、异亮氨酸、缬氨酸、苯丙氨酸含量会影响最终酒样中酯类物质的含量;CHEN等[36]发现荔枝酒中的苯丙氨酸含量影响到乙酸苯乙酯、异丁酸苯乙酯和己酸苯乙酯的含量;HERNáNDEZ-ORTE等[37]发现外加苯丙氨酸、丙氨酸、天冬氨酸会促进乙酸苯乙酯、己酸乙酯、辛酸乙酯等乙酸酯和中链脂肪酸乙酯含量的增加。上述研究印证了本研究中天冬氨酸、谷氨酸、缬氨酸、丙氨酸、苯丙氨酸、异亮氨酸与乙酸酯、中链脂肪酸乙酯具有较高相关性的结果。
本研究综合分析了刺葡萄发酵组与‘赤霞珠’葡萄间的氨基酸及果香酯类物质组成、氨基酸-果香酯类相关性数据,在前人对氨基酸与香气物质关联性研究的基础上,进一步明确真实发酵环境下刺葡萄的氨基酸组成与果香酯类物质的生成之间具有显著相关性,同时发现葡萄中氨基酸组成对果香酯类物质生成可能具有“木桶效应”:刺葡萄中含量显著低于‘赤霞珠’葡萄的丙氨酸、苯丙氨酸、异亮氨酸构成了刺葡萄氨基酸组成的“短板”,限制了刺葡萄酒发酵体系中果香酯类物质积累,导致刺葡萄酒中果香酯类物质含量低于‘赤霞珠’。尽管刺葡萄中天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸含量高于‘赤霞珠’葡萄,但上述含量较高的氨基酸无法弥补刺葡萄发酵环境中丙氨酸、苯丙氨酸、异亮氨酸含量偏低导致的果香酯类物质含量低下,在刺葡萄酒发酵过程中成为果香酯类物质合成的限制因素。这可能表明发酵葡萄汁中均衡且充足的氨基酸组成对果香酯类物质的生成具有重要作用。在刺葡萄发酵过程中补充上述“短板”氨基酸将可能提高刺葡萄酒中果香酯类物质的含量。尽管有研究显示在发酵过程中对任意单一氨基酸的补充都将在一定程度上影响特定果香酯类的含量[13],但对于优化刺葡萄酒果香酯类含量、提高香气质量而言,基于特定产区内葡萄的氨基酸组成,针对性地补充发酵体系中的“短板”氨基酸具有更高的经济价值和应用前景。此外,发酵基质中无机氮源的含量及补充时间[38]、有机氮源与无机氮源的组成及比例[39]等对葡萄酒果香酯类的含量也具有明显影响,而本研究未涉及无机氮源的考察。因此,有必要进行进一步研究,明确葡萄果实及葡萄发酵体系中以氨基酸为主的有机氮源的组成对葡萄酒果香酯类的影响,进一步阐释氨基酸对果香酯类生成的作用机理,为基于发酵氮营养调控的果酒增香酿造技术的开发提供更坚实的理论基础。
4 结论
刺葡萄中除脯氨酸外的总氨基酸含量是‘赤霞珠’葡萄的3倍,刺葡萄中天冬氨酸、谷氨酸、精氨酸、γ-氨基丁酸、缬氨酸含量高于‘赤霞珠’,但丙氨酸、苯丙氨酸、异亮氨酸含量显著低于‘赤霞珠’。刺葡萄发酵酒中的乙酸酯、短链脂肪酸乙酯和中链脂肪酸乙酯含量显著低于‘赤霞珠’。氨基酸与果香酯类物质的相关性分析显示,刺葡萄中丙氨酸、苯丙氨酸、异亮氨酸与果香酯类物质的相关性低于‘赤霞珠’中上述氨基酸与果香酯类物质的相关性,这可能是导致刺葡萄酒中果香酯类物质含量显著偏低的原因。致谢:
感谢西北农林科技大学测试中心李爱华老师为本研究提供的仪器分析技术支持。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.foodres.2013.09.039URL [本文引用: 1]

Spine grape (Vitis davidii Foex) is a wild grape species in South China. However, it has not yet been effectively developed and utilized. To provide sufficient experimental evidence for the characteristic flavor of the spine grapes (V. davidii Foex), nine clones of the spine grape from Zhongfang County, Hunan Province, China, were evaluated for their aromatic components. In this study, several new compounds, including geranylacetone, mequinol, and eugenol were isolated from the spine grape, that have not been reported for other Vitis species. These compounds likely contribute to the characteristic flavor of the spine grape. (E,E)-2,4-hexadienal, which is not produced by Vitis vinifera, was found in the spine grape in large concentration. The results showed that C-6 compounds are predominant aromatic components in various spine grape clones, accounting for 71% to 94% of the sum of aromatic compounds identified. (E)-2-hexenal was the predominant C-6 compound, ranging from 814 +/- 60 (Baiputao) to 3788 74 (Xiangzhenzhu #2) mu g/kg. Regarding terpenes and C-13-norisoprenoids, the highest content of nerol, linalool, menthol, and beta-ionone was present in Seputao grapes, while the highest content of beta-damascenone was detected in Miputao #2. Benzyl alcohol was the most abundant shikimic add derivative, and varied between 106 +/- 6.9 and 522 +/- 50 mu g/kg among the different spine grape cultivars. For most aromatic compounds identified in this study, the content in Seputao was significantly higher than the other eight spine grape clones. This presents Seputao to a candidate for development of higher utilization value. (C) 2013 Elsevier Ltd.
DOI:10.1016/j.lwt.2019.03.101URL [本文引用: 1]
DOI:10.1021/jf9000555URL [本文引用: 1]
DOI:10.1016/j.foodchem.2017.01.113URL [本文引用: 13]
DOI:10.1021/jf9803381URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ijfoodmicro.2015.04.003URL [本文引用: 1]
DOI:10.1021/jf011395oURL [本文引用: 1]
DOI:10.1016/j.lwt.2012.11.022URL [本文引用: 2]
The biochemical formation of aroma-active metabolites determines different characteristics of aroma and taste in fermented beverages. Amino acid utilization by yeast during brewer's wort fermentation is seen as linked to flavor profile. For major understanding of the relationship between amino acid composition in wort and their impact on the synthesis of aroma-active metabolites amino acids were varied in synthetic medium and fermented by Saccharomyces cerevisiae strain S81 and Saccharomyces pastorianus var. carlsbergensis strain S23, respectively. After fermentation of different amino acid combinations higher alcohols and esters were detected by gas chromatography to determine specific aroma compound spectrum. Partial least square (PLS) regression and variable importance in the projection (VIP) were used to establish a relation between amino acids and the resulting concentrations of aroma compounds by means of pattern recognition, indicating most representative amino acids in aroma compound synthesis during fermentation. Thus, a fingerprint of amino acid importance on the detected aroma compound spectrum was created. The most important explanatory variables affecting the synthesis of aroma-active substances of S. pastorianus var. carlsbergensis are leucine, isoleucine, valine, glutamine, cysteine and surprisingly proline. In case of S. cerevisiae the significant amino acids are leucine, isoleucine, valine, histidine, glutamine and again proline. (C) 2012 Elsevier Ltd.
DOI:10.3389/fmicb.2017.02554URL [本文引用: 3]
DOI:10.3864/j.issn.0578-1752.2018.11.015URL [本文引用: 1]
【目的】研究优选发酵毕赤酵母(Pichia fermentans)与酿酒酵母(Saccharomyces cerevisiae)混合发酵对‘爱格丽’(Ecolly)干白葡萄酒的增香作用,优化干白葡萄酒的酿造工艺。【方法】将发酵毕赤酵母菌株H5Y-28与酿酒酵母菌株F5以10﹕1、4﹕1、1﹕1、1﹕4、1﹕10比例接种,进行模拟葡萄汁的混合发酵,并以酿酒酵母纯发酵为对照,发酵过程中监测酵母的活菌数、总菌数,建立菌体生长动力学模型。按相同接种比例进行爱格丽干白葡萄酒的发酵应用,以酿酒酵母纯发酵和添加发酵毕赤酵母胞外酶处理作为对照,酿造次年4月,对所酿葡萄酒酒样进行香气的感官分析和GC-MS分析。【结果】菌体生长动力学表明,随着发酵毕赤酵母接种比例的升高,其存活数量和存活时间明显增加,但优选菌株H5Y-28的高比例(10﹕1、4﹕1)接种处理会降低酿酒酵母的最大生长数量。感官分析结果表明,混合发酵有利于增强‘爱格丽’葡萄酒的果香和花香,尤其是热带水果香气,但高发酵毕赤酵母接种比例(10﹕1)会给葡萄酒带来较强的生青味,而酒样1﹕1、4﹕1仅引入较弱的生青味。挥发性香气成分分析结果表明,品种香气成分的含量随着发酵毕赤酵母接种比例的升高而增加,尤其是萜烯类和C13-去甲类异戊二烯化合物。此外,添加胞外酶能显著提高葡萄酒品种香气的含量,尤其是C13-去甲类异戊二烯化合物的含量(高于纯酿酒酵母发酵酒样90%)。混合发酵显著增加了酒样中发酵香气中的乙酸酯、C6—C12脂肪酸及其酯类、异戊醇与苯乙醇的含量。与单一酿酒酵母发酵酒样相比,1﹕1接种比例有助于提高酒样中26%品种香气成分和39%发酵香气成分的含量,尤其是显著地提高了中链脂肪酸乙酯的含量(40%)。【结论】毕赤酵母与酿酒酵母1﹕1接种处理不影响酿酒酵母生长,且具有较强的增香酿造潜力。
DOI:10.3864/j.issn.0578-1752.2018.11.015URL [本文引用: 1]
【目的】研究优选发酵毕赤酵母(Pichia fermentans)与酿酒酵母(Saccharomyces cerevisiae)混合发酵对‘爱格丽’(Ecolly)干白葡萄酒的增香作用,优化干白葡萄酒的酿造工艺。【方法】将发酵毕赤酵母菌株H5Y-28与酿酒酵母菌株F5以10﹕1、4﹕1、1﹕1、1﹕4、1﹕10比例接种,进行模拟葡萄汁的混合发酵,并以酿酒酵母纯发酵为对照,发酵过程中监测酵母的活菌数、总菌数,建立菌体生长动力学模型。按相同接种比例进行爱格丽干白葡萄酒的发酵应用,以酿酒酵母纯发酵和添加发酵毕赤酵母胞外酶处理作为对照,酿造次年4月,对所酿葡萄酒酒样进行香气的感官分析和GC-MS分析。【结果】菌体生长动力学表明,随着发酵毕赤酵母接种比例的升高,其存活数量和存活时间明显增加,但优选菌株H5Y-28的高比例(10﹕1、4﹕1)接种处理会降低酿酒酵母的最大生长数量。感官分析结果表明,混合发酵有利于增强‘爱格丽’葡萄酒的果香和花香,尤其是热带水果香气,但高发酵毕赤酵母接种比例(10﹕1)会给葡萄酒带来较强的生青味,而酒样1﹕1、4﹕1仅引入较弱的生青味。挥发性香气成分分析结果表明,品种香气成分的含量随着发酵毕赤酵母接种比例的升高而增加,尤其是萜烯类和C13-去甲类异戊二烯化合物。此外,添加胞外酶能显著提高葡萄酒品种香气的含量,尤其是C13-去甲类异戊二烯化合物的含量(高于纯酿酒酵母发酵酒样90%)。混合发酵显著增加了酒样中发酵香气中的乙酸酯、C6—C12脂肪酸及其酯类、异戊醇与苯乙醇的含量。与单一酿酒酵母发酵酒样相比,1﹕1接种比例有助于提高酒样中26%品种香气成分和39%发酵香气成分的含量,尤其是显著地提高了中链脂肪酸乙酯的含量(40%)。【结论】毕赤酵母与酿酒酵母1﹕1接种处理不影响酿酒酵母生长,且具有较强的增香酿造潜力。
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1016/j.foodres.2019.05.030URL [本文引用: 1]
DOI:10.1016/j.lwt.2015.05.049URL [本文引用: 4]
DOI:10.1016/j.lwt.2015.05.044URL [本文引用: 2]
DOI:10.1002/(ISSN)1097-0010URL [本文引用: 4]
DOI:10.1021/jf3043874URL [本文引用: 2]
The influence of a prefermentative freezing process on changes of aromatic characteristics and volatile compounds in Meili wines was studied to optimize freezing parameters and reveal the mathematical relationship between aromatic characteristics and volatile compounds. The wines obtained were characterized by sensory evaluation and stir bar sorptive extraction (SBSE) followed by a thermal desorption-gas chromatography-mass spectrometry analysis. A total of 28 aromatic descriptors from 6 categories of wine aroma terminology were identified by judging with high "modified frequency (MF%)". In addition, 19 varietal aroma compounds and 36 fermentation aroma compounds were quantitated, followed by the determination of odor activity values (OAVs). On the basis of the data obtained, principal component analysis (PCA) was used to find the relationship between characteristic aroma terms and different freezing conditions, and then partial least-squares regression (PLSR) was proposed to establish the mathematical relationship between the resulting terms and impact odorants. Natural thawing treatment on frozen must resulted in higher aroma quality with higher extraction of varietal aroma compounds. Lower frozen maceration temperature contributed to higher esters and organic acids. Impact aroma compounds were related to models for floral, sweet fruit, temperate fruit, and vegetal, whereas the model of rose and strawberry contained only varietal volatile compounds, and temperate fruit could be regressed by impact fermentation aroma compounds.
DOI:10.1111/1750-3841.13253URL [本文引用: 2]
DOI:10.4236/health.2009.13029URL [本文引用: 2]
[本文引用: 1]
DOI:10.1016/j.foodchem.2009.06.045URL [本文引用: 1]
DOI:10.1016/j.foodres.2017.11.021URL [本文引用: 1]
DOI:10.3969/j.issn.0254-5071.2011.01.009URL [本文引用: 3]
应用HPLC对我国4个产区多个品种葡萄酒酿造过程样品中氨基酸含量变化趋势进行了研究,结果表明,L-脯氨酸是所有葡萄汁中氨基酸的主体,也是各样品含量差异最大的氨基酸;红葡萄酒中含脯氨酸、苏氨酸、赖氨酸等20种氨基酸,其中以脯氨酸含量最高,发现不同葡萄品种之间游离氨基酸种类和含量有差别,这与葡萄品种、气候、发酵工艺有关.
.
DOI:10.3969/j.issn.0254-5071.2011.01.009URL [本文引用: 3]
应用HPLC对我国4个产区多个品种葡萄酒酿造过程样品中氨基酸含量变化趋势进行了研究,结果表明,L-脯氨酸是所有葡萄汁中氨基酸的主体,也是各样品含量差异最大的氨基酸;红葡萄酒中含脯氨酸、苏氨酸、赖氨酸等20种氨基酸,其中以脯氨酸含量最高,发现不同葡萄品种之间游离氨基酸种类和含量有差别,这与葡萄品种、气候、发酵工艺有关.
DOI:10.1016/j.foodchem.2010.05.112URL [本文引用: 1]
The aim of this work was to study the influence of grape variety, cultivation system and stage of grape maturation on nitrogen compounds evolution during alcoholic fermentation. To do this, four grape varieties, Monastrell, Merlot, Syrah and Petit Verdot, traditionally cultivated and Monastrell cultivated using organic agriculture, which were collected in two different stages of maturation, were used. The results showed that, regardless of grape variety, cultivation system and stage of grape maturation, the consumption of nitrogen compounds was directly proportional (R(2) > 0.7) to their concentration in natural musts. This is very important as it is the first time that these results were obtained with natural musts without external addition of nitrogen compounds. (C) 2010 Elsevier Ltd.
DOI:10.1021/jf0487001URL [本文引用: 1]
DOI:10.1016/j.foodchem.2009.12.004URL [本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1007/s00217-013-2059-xURL [本文引用: 1]
DOI:10.1016/j.biotechadv.2005.11.005URL [本文引用: 1]
Abstract
Microbial catabolism of amino acids produces flavour compounds of importance for foods such as cheese, wine and fermented sausages. Lactic acid bacteria are equipped with enzyme systems for using the amino acids in their metabolism and are useful for flavour formation of foods. Branched-chain amino acids (Leu, Ile, Val) are converted into compounds contributing to malty, fruity and sweaty flavours; catabolism of aromatic amino acids (Phe, Tyr, Trp) produce floral, chemical and faecal flavours; aspartic acid (Asp) is catabolised into buttery flavours and sulphuric amino acids (Met, Cys) are transferred into compounds contributing to boiled cabbage, meaty and garlic flavours.[本文引用: 1]
DOI:10.1016/j.foodchem.2004.02.021URL [本文引用: 1]
DOI:10.1016/j.lwt.2007.03.018URL [本文引用: 1]
Abstract
In this work the effect of the addition of different quantities of amino acids to a nitrogen-deficient must on the formation of volatile compounds during the wine alcoholic fermentation was studied. To do so, fermentations of Mazuelo must were carried out, to which were added ammonium and 45, 120, and 250 mg/l of amino acids. The results showed that the formation of total esters, isoamyl acetate, and 2-phenylethyl acetate was directly proportional to the quantity of amino acids added to the must while the synthesis of diethyl succinate, and ethyl 3-hydroxybutyrate was inversely proportional to this addition. The alcohols, with the exception of tyrosol and 2-phenylethanol, did not show any direct correlation between their formation and the addition of amino acids. The quantity of amino acids added to the must favoured the formation of total acids but it had scant influence on the individual synthesis of the acids. So, it may be stated that, in general, the addition of amino acids in the concentrations under study to a nitrogen-deficient must, favoured the formation of volatile compounds in the wine.DOI:10.1016/j.ijfoodmicro.2013.10.025URL [本文引用: 1]
The impact of individual aromatic amino add addition (L-phenylalanine, L-tryptophan and L-tyrosine) on nonvolatile and volatile constituents in lychee wine fermented with Saccharomyces cerevisiae var. cerevisiae MERIT.ferm was studied. None of the added amino acids had any significant effect on the yeast cell count, pH, soluble solid contents, sugars and ethanol. The addition of L-phenylalanine significantly reduced the production of pyruvic and succinic acids. The addition of each amino acid dramatically reduced the consumption of proline and decreased the production of glycerol. Supplementation of the lychee juice with L-phenylalanine resulted in the formation of significantly higher amounts of 2-phenylethyl alcohol, 2-phenylethyl acetate, 2-phenylethyl isobutyrate and 2-phenylethyl hexanoate. In contrast, supplementation with L-tryptophan and L.-tyrosine had negligible effects on the volatile profile of lychee wines. These findings suggest that selectively adding amino acids may be used as a tool to modulate the volatile profile of lychee wines so as to diversify and/or intensify wine flavour and style. (C) 2013 Elsevier B.V.
DOI:10.1016/j.foodchem.2005.05.073URL [本文引用: 1]
DOI:10.1016/j.fm.2018.04.005URL [本文引用: 1]
DOI:10.1016/j.foodchem.2011.01.092URL [本文引用: 1]
Inorganic nitrogen salts, and to a growing extent organic nitrogen preparations, are widely used to ameliorate a nitrogen deficiency in wine fermentation, but the impact of nitrogen supplementation on perceived wine sensory profile is essentially unknown. Supplementation of a low nitrogen Chardonnay grape juice with either ammonium nitrogen or combined amino acid and ammonium nitrogen showed that the type of nitrogen and concentration in the range 160-480 mg N/l had a substantial impact on the formation of yeast volatile compounds and perceived wine aroma. Addition of amino acid and ammonium nitrogen increased both acetate and medium chain fatty acid esters to a greater extent and decreased higher alcohols to a lesser extent than ammonium nitrogen alone whereas ammonium nitrogen substantially increased ethyl acetate and acetic acid. Low nitrogen wines were rated relatively low in floral/fruity aroma descriptors, while moderate nitrogen wines showed a good balance between desirable and less desirable attributes, whereas high nitrogen produced either an acetic/solvent character or highest ratings for floral/fruity attributes, depending on nitrogen type. These results show that amount and type of nitrogen supplement can substantially modulate Chardonnay wine volatiles composition and perceived aroma. (C) 2011 Elsevier Ltd.

{kind=link}
{kind=link}
{kind=link}
{kind=link}