 ,2, 邢凯,1
,2, 邢凯,1Differential Expression of mRNA and lncRNA in Longissimus Dorsi Muscle of Songliao Black Pig and Landrace Pig Based on High-Throughput Sequencing Technique
WU YaoQun1, CHEN ShaoKang3, SHENG XiHui1, QI XiaoLong1, WANG XiangGuo1, NI HeMin1, GUO Yong1, WANG ChuDuan,2, XING Kai,1通讯作者:
责任编辑: 林鉴非
收稿日期:2018-12-10接受日期:2019-12-5网络出版日期:2020-02-16
| 基金资助: |
Received:2018-12-10Accepted:2019-12-5Online:2020-02-16
作者简介 About authors
吴垚群,E-mail:1530321769@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (1304KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
吴垚群, 陈少康, 盛熙晖, 齐晓龙, 王相国, 倪和民, 郭勇, 王楚端, 邢凯. 用高通量测序技术研究松辽黑猪与长白猪背最长肌 mRNA和lncRNA的差异表达[J]. 中国农业科学, 2020, 53(4): 836-847 doi:10.3864/j.issn.0578-1752.2020.04.015
WU YaoQun, CHEN ShaoKang, SHENG XiHui, QI XiaoLong, WANG XiangGuo, NI HeMin, GUO Yong, WANG ChuDuan, XING Kai.
0 引言
【研究意义】 长链非编码RNA(long non-coding RNA, lncRNA)是一类转录长度大于200nt的非编码RNAs。目前许多研究已证实,lncRNA 参与了广泛的基因表达的调控过程,在细胞增殖、分化、凋亡以及肿瘤的发生和发展中发挥重要作用[1,2]。随着高通量测序技术的广泛应用,已经有众多的lncRNA在肌肉发育和脂肪沉积中的作用被发现[3,4,5]。由此可见lncRNA是基因表达调控的重要因素,本研究的意义在于通过对不同品种猪背最长肌组织中lncRNA的研究,进一步解析猪骨骼肌生长发育和脂肪沉积的分子机理。【前人研究进展】骨骼肌占哺乳动物总体重的50%左右[6]。多个研究发现lncRNAs在骨骼肌发育过程中起着关键作用。在小鼠成肌细胞中,一个lncRNA位于发育多能性相关2(developmental pluripotency associated 2, Dppa2)基因上游,其转录产物可以招募多种DNA甲基化转移酶到Dppa2基因的启动子区域,使Dppa2 基因沉默,进而促进成肌细胞的分化和肌肉再生[7]。lncRNA H19由H19/胰岛素样生长因子2(insulin like growth factor 2, IGF2)基因座母体等位基因转录,在胚胎发育和成年机体的肌肉组织中表达较高,H19 通过结合某些复合物,起到抑制IGF2转录的作用[8]。LU等人发现lncRNA YY1相关肌发生RNA 1(YY1-associated myogenesis RNA 1, Yam-1),受转录因子YY1(YY1 transcription factor, YY1)的正向调控,Yam-1可以激活miR-715、抑制Wnt家庭成员7B(Wnt family member 7B, Wnt7b)的表达,可以作为肌细胞抑制因子发挥作用[9]。在牛骨骼肌组织中,lncRNA-133a可以促进牛骨骼肌卫星细胞的增殖,对牛骨骼肌卫星细胞的增殖有着正向调控作用[10]。lncRNA 作为基因表达的重要调控分子,可以通过多种作用机制,从转录水平、转录后水平以及染色质水平调节靶基因的表达,进而调控生命活动[11]。【本研究切入点】猪骨骼肌的生长发育与其生长速度和瘦肉率息息相关,直接关系着生猪养殖的生产效益。猪肌肉生长发育的差异是导致不同品种猪肉品质差异的重要原因。松辽黑猪是我国培育的第一个瘦肉型黑色母系品种,是由杜洛克猪、长白猪和东北民猪3个品种经过长时间杂交培育而成的一个培育品种。松辽黑猪具有繁殖率高、肉质优良、适应性强、耐粗饲等特性,同时松辽黑猪的肌内脂肪含量可达3.19%,猪肉品质极佳[12]。长白猪原产于丹麦, 是目前世界上使用最为广泛的瘦肉型猪种之一,具有生长速度快,瘦肉率高等特点。研究发现,二者在肉质方面存在较大差异[12,13]。因此,以松辽黑猪和长白猪作为试验对象来研究lncRNA与不同猪品种骨骼肌发育差异的关系,有利于研究lncRNA在猪肌肉发育过程中的分子机制。目前已有多项研究证实lncRNA在猪肌肉发育过程中发挥作用,但lncRNA在中外不同品种猪肌肉生长发育过程中的差异性以及参与的生物学信息还有待于深入研究。【拟解决的关键问题】采用高通量测序技术对其背最长肌转录组进行测序分析,筛选差异表达的mRNA和lncRNAs,并进行相关生物信息学的分析。最终筛选出关于猪肌肉发育和脂肪沉积的关键mRNA和lncRNAs,以加深对肌肉生长发育和脂肪沉积分子调控机制的了解,为国内外猪品种在肉质和生长性状方面形成差异的分子机理研究提供新的参考信息。1 材料与方法
1.1 试验材料
本研究所用的长白猪和松辽黑猪来自天津宁河原种猪场。所有动物均在统一条件下饲养管理,自由采食和饮水。饲养至体重为100kg左右时进行屠宰。屠宰在华都阳光食品公司进行。屠宰标准按照GB/T 17236-2008生猪屠宰操作规程进行[14]。屠宰后,立即将背最长肌组织移至液氮中进行冷冻保存,待提取RNA使用。1.2 总RNA提取和建库
采用Trizol法按照产品说明提取每个背最长肌组织的总RNA。用1%的琼脂糖凝胶电泳初步检测总RNA 的完整性以及有无DNA 污染。采用Nanodrop、Qubit 2.0和Aglient 2100方法检测各样品的浓度、纯度和完整性等。保证样品浓度≥500 ng·µL-1,28S:18S >1.0,RIN≥8。将每个品种的6个样品的总RNA等量混合后,进行混池建库,随后利用Illumina HiSeq 2500 (Illumina, America)平台进行双末端测序。文库的构建及测序在广州基迪奥生物科技有限公司进行。1.3 数据的质控、比对和组装
FastQC软件(http://www.bioinformatics.babraham. ac.uk/projects/fastqc/)用于对原始数据的质量控制,质控的标准为:(1)去除含adapter的reads;(2)去除含N比例大于10% 的reads;(3)去除低质量reads (质量值Q≤20的碱基数占整条read的50%以上)[15]。TopHat2将高质量的序列数据(clean reads)比对到猪的参考基因组Sus scrofa 11.1中[16]。Cufflinks流程用于数据的转录本组装、注释与合并[17]。HT-seq软件用于每个样本中基因表达量的计算[18]。1.4 差异表达基因的功能富集分析
基因本体(gene ontology, GO )数据库 (http://www. geneontology.org/) 被用于差异表达基因的GO功能富集分析,包括细胞组分,分子功能和生物学过程。超几何检验被用于P-value的计算其公式为$P=1-\sum^{m-1}_{i=0}\frac{(^{M}_{i})(^{N-M}_{n-i})}{(^{N}_{n})}$
式中:N为所有Unigene中具有GO注释的转录本数目;n为N中差异表达转录本的数目;M为所有Unigene中注释为某特定GO 条目的转录本数目;m为注释为某特定GO条目的差异表达转录本数目。FDR被用于校正原始的P value。FDR<0.05的GO terms被鉴定为显著富集的通路。京都基因和基因组百科全书(kyoto encyclopedia of genes and genomes, KEGG) 数据库 (https://www.genome.jp/kegg/) 被用于通路研究。同样,采用上述的方法和阈值鉴定差异表达基因的显著富集通路。
1.5 lncRNA的鉴别
首先,筛选cuffcompare转录本类别符号为“I”、“j”、“o”、“u”、“x”的转录本,其可能是新的lncRNA。进一步,为了剔除其中的小的非编码RNA,过滤掉其中外显子总长度小于200nt及只有1个或无外显子的转录本。将剩下的转录本通过CPAT(Computerized Progressive Attention Training )软件[19]进行lncRNA的初步预测。为了得到可靠的lncRNA,继而利用CPC(Cetylpyridinium Chloride)软件[20]对CPAT软件预测的结果进行进一步lncRNA的识别。Cufflinks用于对筛选出来的lncRNA进行定量分析[17]。1.6 基因及lncRNA的差异表达分析
采用FPKM(fragments per kb per million fragments)的方法校正基因的表达水平,其同时考虑到了测序深度和基因长度对基因计数的影响[21]。edgeR软件包被用于筛选差异表达基因[22],其标准为FDR<0.05且|log2FC|>1。1.7 差异表达lncRNAs的靶基因的预测
基于lncRNA的顺式作用和反式作用方式预测差异表达lncRNA的靶基因。在lncRNA基因组位置上下游100kb的基因认为是该lncRNA潜在的靶基因。利用RNAplex软件来预测lncRNA通过碱基互补配对的反式作用模式的潜在靶基因[23]。1.8 测序结果可靠性分析
为了验证测序结果的可靠性,对差异表达的mRNA和lncRNA进行验证。挑选10个差异表达的mRNA和10个差异表达的lncRNA,包括5个上调基因和5个下调基因,进行RT-PCR验证。RT-PCR所使用的cDNA由高通量测序使用的剩余的RNA样品反转录产生。设计所挑选基因的引物,使用甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH)基因作为内部对照,引物均由上海生工合成(表1)。进行RT-PCR,并使用2-ΔΔCt方法计算样品之间基因的相对表达水平。Table 1
表1
表1引物序列
Table 1
| 基因Gene | 上游引物(5′-3′)Forward primer | 下游引物(5′-3′)Reverse primer |
|---|---|---|
| HPSE | GCGTTCCTGTCCGTAACCAT | ATGCGGGAGACAAACCTCTG |
| BTK | TTTCAGCACCATCCCTGAGC | ACGGCGCATTCTTGTTTTGT |
| SFT2D2 | ACTGTCTTCCTCATGGGACC | GTGCAAAACACAACAGCACC |
| SMAD6 | CCCATCTTCGTCAACTCCCC | GCTCGAAGTCGAACACCTTG |
| FGF10 | AGATGTCCGCTGGAGAAAGC | TCCAGGATACTGTACGGGCA |
| TECRL | CAAAGCATGTGGGCCCTTTT | GTGGTCCAACTGACTTGTTGAC |
| PRKD2 | CTACAACCGCTCACTGGACAT | TGCGTTCTGGATCTGGTCAT |
| TRAPPC2 | ACCAGTTCATAGCACACGCT | GCCGAGACAAACCACTCATTG |
| HOXD1 | CGCACGAATTTCAGCACCAA | TCAGCTGCAATGAGTTGGCT |
| ATP2B3 | GGAGCAGAAGTTCACCGTCA | GGCAGCAGGTCTCCGTATTT |
| TCONS-OOO52690 | GGGAAATCCAGCATCGTGTG | CCACAAGGCACAGTTTTGGTC |
| TCONS-OOO59635 | AGCGTTTATTCCTCCCAGGC | AGGTACTTGTAGGGCCGGTA |
| TCONS-OOO35174 | CACCGTAGGGTCGTTGCA | CATCCTTCCTTCCGTCCAG |
| TCONS-OOO39610 | ACACCTTGGGACCCCAACTA | GGCCAGAATGACGGATGACA |
| TCONS-OOO59695 | CAGGGTTGTCCAGCCAATGT | TGCTGGGCTGTCATCCTCTT |
| TCONS-OOO16916 | ATTACTTTGCCCCGGAGGTCT | TAATTAGGGCTTCCCGGGTCT |
| TCONS-OOO46917 | TGCCCGATACTGTAATGCCA | CATTCCCCGAGCACGGATTC |
| TCONS-OOO67508 | CGTGGACGAAAAACGATCCG | CTTCACCTCGTTCAGGGCTT |
| TCONS-OOO36675 | GGTCTTCCAAATCGGTTGCC | ACAGGGGTTGCTTTGGTGTA |
| TCONS-OOO52912 | GGACCTGGACCCAATCAACT | TGATGATGGCACGGAATGGT |
| GAPDH | TATGATTCCACCCACGGCAA | CCATTTGATGTTGGCGGGAT |
新窗口打开|下载CSV
2 结果
2.1 测序数据的质量及比对信息
本研究通过Illumina HiSeqTM 2000测序平台对松辽黑猪和长白猪的背最长肌组织mRNA进行测序,每个样本平均得到约10G数据量。测序数据的Q20在96%以上,Q30在91%以上,测序质量良好。对原始数据质控后,平均每个样本得到18 224 475 795条150bp的clean reads(表2)。除去比对到核糖体RNA上的reads后,平均81.22% 的clean reads比对到猪的参考基因组中,其中唯一比对到基因组位置的为79.49%。各项数据显示测序数据的质量较好,可以进行后续的分析。Table 2
表2
表2质控前后样本测序数据的质量
Table 2
| 样本 Sample | Reads数目 No. | Q20(%) | Q30(%) | N(%) | GC含量 GC content (%) | |
|---|---|---|---|---|---|---|
| 质控前 Before QC | SL | 18817856700 | 96.92% | 92.24% | 0.01% | 49.83% |
| L | 20211575400 | 96.60% | 91.58% | 0.01% | 51.92% | |
| 质控后 After QC | SL | 17560551309 | 98.01% | 94.00% | 0.01% | 49.77% |
| L | 18888400280 | 97.82% | 93.52% | 0.01% | 51.26% |
新窗口打开|下载CSV
2.2 基因表达水平和差异分析
对转录本进行统计分析,松辽黑猪和长白猪得到的所有转录本数目分别是21 866个和21 972个,其中检测到新的转录本分别有7 846个和7 838个。对转录本的表达量统计发现,多数转录本的RPKM值在1 000以下,其中FPKM值在1以下转录本最多,为11 047个;FPKM值1—10之间的转录本为9 905个;FPKM值在10—100的转录本为2 915个;而FPKM值大于100的转录本只有363个,占总数的1.65%。同时,还发现两个样本的转录本表达丰度模式基本相同(图1)。图1
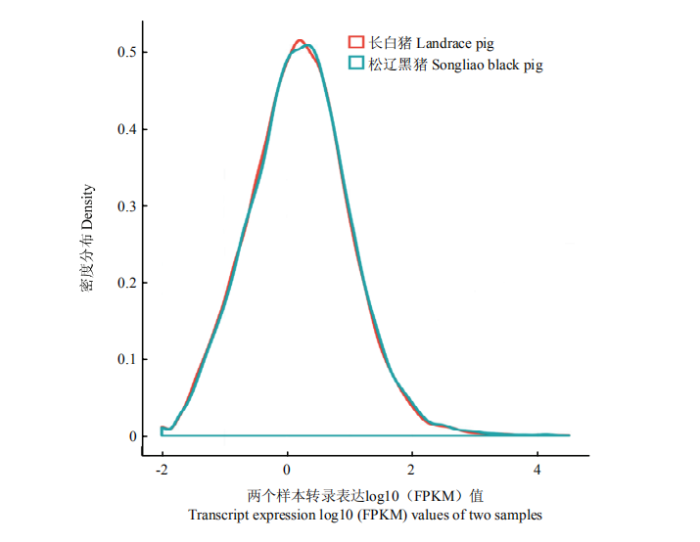 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1转录本表达量丰度分布函数图
横坐标为log10(FPKM)值,数值越高,表示转录本表达量越高;纵坐标为转录本的密度,即为对应横轴表达量的转录本数/检测已表达转录本的总数;分布曲线的峰值表示整个样本转录本表达量最集中的区域
Fig. 1Transcript distribution abundance distribution function map
The abscissa is the log10 (FPKM) value. The higher the value, the higher the transcript expression; the ordinate is the density of the transcript, which is the number of transcripts corresponding to the horizontal axis expression/detection of the total number of expressed transcripts; The peak of the distribution curve represents the region where the expression of the entire sample transcript is most concentrated
利用edgeR软件包来筛选差异表达基因,显著差异表达的条件为 FDR<0.05且| log2FC|>1 。结果显示两组之间一共有4 239个基因显著差异,相对于长白猪,松辽黑猪组中2 023个显著上调,2 216个显著下调(图2)。
图2
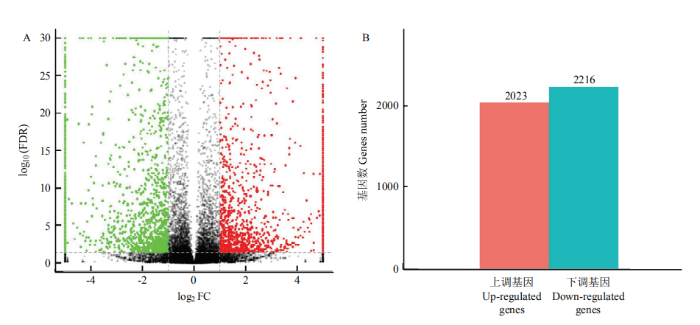 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2基因的差异表达分析
A图中横坐标表示样品间的差异倍数对数值,纵坐标表示两个样品的-log10(FDR)值;红色表示长白猪相对于松辽黑猪的样本表达量上调和绿色表示表达量下调,黑色的点标志没有差异。B图中横坐标表示成对比较的样品,纵坐标表示差异表达的基因数
Fig. 2Results of differential expression analysis of genes.
In the graph A, the abscissa indicates the logarithm of the difference between the samples, the ordinate indicates the -log10 (FDR) value of the two samples, and the red indicates the up-regulation of the sample expression and the green expression of the Landrace pig relative to the Songliao black pig. Down, there is no difference in the black dot mark. In the B diagram, the abscissa indicates the pairwise comparison of the samples, and the ordinate indicates the number of genes differentially expressed
2.3 差异表达基因功能分析
为了解差异表达基因涉及的生物学功能,使用超几何检验的方法对差异表达基因进行GO条目和KEGG通路的功能富集分析。富集GO条目的结果显示,差异表达基因主要分布在生物学过程、分子功能和细胞组成的三大类中的3 780个GO条目中,其中 1 314个是显著富集的(Q-value≥0.05)。主要参与的细胞膜的组成、跨膜运输、蛋白质结合、转运蛋白活性、能量代谢和氧化磷酸化等过程(表3)。对差异表达基因进行KEGG通路功能富集分析,共鉴定出25个显著的通路,其中包括生物素代谢、泛酸盐辅酶A生物合成、多聚糖生成、磷脂酰肌醇信号系统、肌动蛋白调控、溶酶体等通路(图3)。Table 3
表3
表3差异表达基因最显著富集的GO条目
Table 3
| 类型 Type | GO条目 GO entries | 基因数目 Number of genes | 校正后P值 Corrected P value |
|---|---|---|---|
| 细胞组成 Cell composition | 膜组分 Membrane part | 299 | 1×10-6 |
| 膜内组分 Intrinsic component of membrane | 249 | 1×10-6 | |
| 细胞器 Organelle | 1825 | 1×10-6 | |
| 细胞内部分 Intracellular part | 1890 | 1×10-6 | |
| 胞内 Intracellular | 2003 | 1×10-6 | |
| 被膜小泡 Membrane-bounded vesicle | 212 | 1×10-6 | |
| 包膜细胞器 Membrane-bounded organelle | 1491 | 1×10-6 | |
| 细胞器膜 Organelle membrane | 135 | 1×10-6 | |
| 胞内细胞器 Intracellular organelle | 1624 | 1×10-6 | |
| 质膜组分 Plasma membrane part | 68 | 1×10-6 | |
| 分子功能 Molecular function | 阳离子结合 Cation binding | 316 | 1×10-6 |
| 结合 Binding | 1791 | 1×10-6 | |
| 蛋白质结合 Protein binding | 705 | 1×10-6 | |
| 金属离子结合 Metal ion binding | 171 | 1×10-6 | |
| 转换金属离子结合 Transition metal ion binding | 128 | 1×10-6 | |
| 小分子结合(small molecule binding) | 181 | 1×10-6 | |
| 碳水化合物衍生物结合(carbohydrate derivative binding) | 164 | 1×10-6 | |
| 分子传感活性(molecular transducer activity) | 75 | 1×10-6 | |
| 嘌呤核苷酸结合 Purine nucleoside binding | 144 | 1×10-6 | |
| 核糖核苷结合 Ribonucleotide binding | 144 | 1×10-6 | |
| 生物过程 Biological process | 调控代谢过程 Regulation of metabolic process | 461 | 1×10-6 |
| 调控大分子代谢过程 Regulation of macromolecule metabolic process | 376 | 1×10-6 | |
| 生物过程的正调控 Positive regulation of biological process | 246 | 1×10-6 | |
| 调控细胞代谢过程 Regulation of cellular metabolic process | 312 | 1×10-6 | |
| 调控初级代谢过程 Regulation of primary metabolic process | 295 | 1×10-6 | |
| 细胞过程的正调控 Positive regulation of cellular process | 206 | 1×10-6 | |
| 基因表达调控 Regulation of gene expression | 275 | 1×10-6 | |
| 分子功能调控 Regulation of molecular function | 183 | 1×10-6 | |
| 调控刺激反应 Regulation of response to stimulus | 196 | 1×10-6 | |
| 调控蛋白质代谢过程 Regulation of protein metabolic process | 158 | 1×10-6 |
新窗口打开|下载CSV
图3
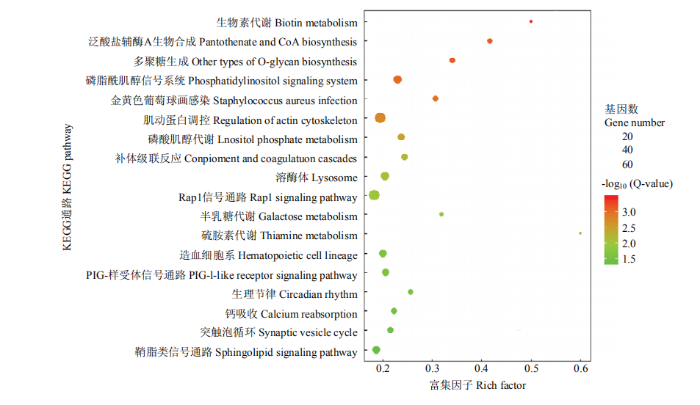 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3差异表达基因的KEGG通路功能富集分析结果
圆的大小和颜色分别代表富集通路和富集显着性的差异表达基因的量
Fig. 3Functional enrichment analysis of KEGG pathways of differentially expressed genes
The size and color of the circle represent the enrichment pathway and the amount of differentially expressed genes with significant enrichment, respectively
2.4 新lncRNA的鉴定及差异表达分析
通过cufflinks、CPAT和CPC流程鉴定转录本中的lncRNA,共得到885个新的lncRNA。所鉴定出的新lncRNA中,在两个组中都有表达的lncRNA有792个,只在松辽黑猪和长白猪中表达的基因分别为49个和44个 (图4-A)。同样利用edgeR分析软件筛选差异表达分析,结果显示松辽黑猪和长白猪之间一共有178个非编码基因显著差异,其中在松辽黑猪中84个显著上调,94个显著下调(图4-B)。图4
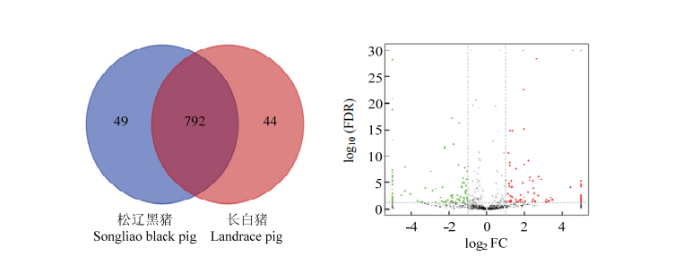 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4lncRNA的差异表达分析
A图表示鉴定出的新lncRNA,蓝色在松辽黑猪中表达的lncRNA,红色表示在长白猪中表达的lncRNA,中间的交集表示松辽黑猪和长白猪中均表达的lncRNA;B图表示松辽黑猪和长白猪之间差异表达的lncRNA,横坐标表示样品间的差异倍数对数值,纵坐标表示两个样品的-log10(FDR)值;红色表示长白猪相对于松辽黑猪的样本表达量上调和绿色表示表达量下调
Fig. 4Differential expression analysis results of lncRNA
A figure shows the identified new LNCRNA, blue shows the LNCRNA expressed in Songliao black pig, red shows the LNCRNA expressed in landrace pig, and the middle intersection shows the LNCRNA expressed in both Songliao black pig and landrace pig; B figure shows lncRNA of difference expression between Songliao black pig and landrace pig, horizontal coordinate shows logarithm of difference multiple between samples, vertical coordinate shows-log10(FDR) value of two samples; The red color showed that the expression of landrace pig was up-regulated and the green color showed down-regulated compared with Songliao black pig
2.5 lncRNA的靶基因预测
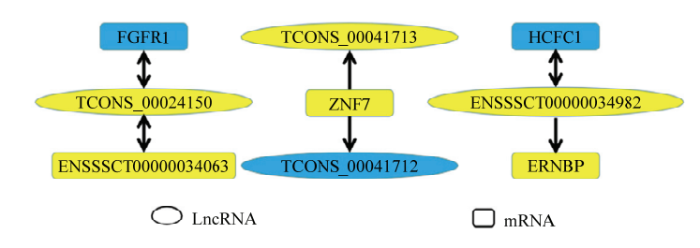
基于lncRNA的作用模式,通过cis作用和反义作用预测差异表达lncRNA的靶基因。根据lncRNA的在染色体上的位置,共在241个新预测到的lncRNA的上下游500kb内找到342个编码基因。同时,在26个已知的lncRNA中发现81个编码基因。使用RNAplex软件预测lncRNA的反义作用靶基因,共鉴定得到84个lncRNA与编码基因存在碱基的互补配对的关系。结合两组间差异表达分析和靶基因预测的结果发现,几对差异表达lncRNA和基因之间可能存在着潜在的调控关系。如新lncRNA TCONS_00041713和TCONS_00041712在ZNF7基因的上游;lncRNA ENSSSCT00000034982在ERNBP基因的下游且与HCFC1存在碱基互补配对的关系;新lncRNA TCONS_00024150分别与FGFR1和 ENSSSCT00000034063基因间存在碱基互补配对的关系(图5)。以上的这些潜在调控关系为研究lncRNA调控基因表达进而影响肉质品质提供新的素材。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5lncRNA和基因之间存在的潜在调控关系
不同颜色表示基因的表达水平,黄色表示基因在松辽黑猪中表达量高,蓝色表示基因在长白猪中的表达量高。箭头表示lncRNA与基因之间存在的潜在调控关系。单向箭头代表基因上下游关系,箭头指向方向表示位于下游的基因;双向箭头表示基因之间存在碱基互补配对关系
Fig. 5Potential regulatory relationships between lncRNA and genes
Different colors indicate the level of gene expression. Yellow indicates that gene expression is high in Songliao Black pig, and blue indicates that gene expression is high in Landrace pigs. Arrows indicate potential regulatory relationships between lncRNA and genes. One-way arrows represent upstream and downstream relationships of genes, arrows point to downstream genes, and two-way arrows represent complementary pairing relationships between genes
2.6 测序结果的验证
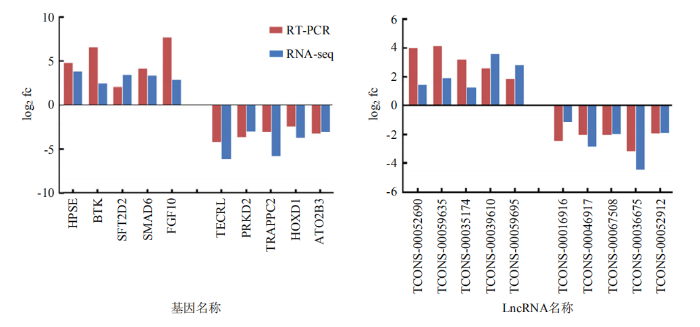
通过RT-PCR对随机挑选的10个差异表达mRNA和10个差异表达lncRNA进行验证。结果表明,随机挑选的10个mRNA的RT-PCR结果与转录组测序结果表达趋势相似(图6-A),同样随机挑选的lncRNA的验证结果也与转录组测序结果表达趋势相似(图6-B),证实了松辽黑猪和长白猪背最长肌组织转录组测序结果具有可靠性,可以用于后续的深入研究。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6差异表达基因的RT-PCR验证
通过RT-PCR验证松辽黑猪和长白猪背最长肌组织基因的差异表达
Fig. 6RT-PCR verification of differentially expressed genes
The differential expression of genes in the longissimus dorsi transcriptome of Songliao black pig and Landrace was verified by RT-PCR
3 讨论
本研究选用在肌肉品质和育肥期期间日增重方面存在较大差异的松辽黑猪和长白猪为对象,对其背最长肌进行RNA-seq测序,筛选与肌肉生长和脂肪沉积相关的基因。结合生物学功能分析发现了一些与松辽黑猪和长白猪背最长肌肌肉发育和脂肪沉积可能相关的mRNA和lncRNA。猪的脂肪性状直接影响猪肉品质,如肌间脂肪含量、胴体瘦肉率等。猪脂肪组织的沉积能力具有品种差异,且不同品种的猪脂肪沉积能力具有较大差异[24]。本研究中,通过功能富集分析发现了一些与脂类代谢差异性显著的信号通路和代谢途径,如磷脂酰肌醇信号通路、鞘脂信号通路、甘油磷脂代谢途径、醛固酮的合成与分泌,与之相关的基因包括二酰基甘油激酶α(diacylglycerol kinase α, DGKα)、磷脂酰肌醇-4-激酶α(phosphatidylinositol 4-kinase α, PI4Kα)、磷脂酰肌醇5-磷酸4激酶2α(phosphatidylinositol-5-phosphate 4-kinase type 2 α, PIP4K2α)、磷脂酶D1(phospholipase D1, PLD1)、脂肪酶F(lipase F, LIPE)、二酰基甘油脂酶α(diacylglycerol lipase α, DAGLα)、蛋白激酶D2(protein kinase D2, PRKD2)、脂质1(lipin 1, LPIN1)、3-磷酸甘油酰基转移酶4(glycerol-3-phosphate acyltransferase 4, GPAT4)。在筛选得到的差异表达基因中,发现一些基因同时参与不同信号通路和代谢途径,如DGKα,其在磷脂酰肌醇信号通路和甘油磷脂代谢中均有参与。DGKα属于二酰基甘油激酶家族,该酶类家族成员众多,主要作用为将1,2-二酰基甘油生成磷脂酸。DGKα是一种在含有特殊脂类和蛋白质分子的外泌体生成的过程中发挥重要调控作用的因子,其在脂类代谢活动中具有重要作用[25]。前人研究发现,DGKα作为LIPF的下游分子,而LIPF与食物中的甘油三酯在胃肠道中的消化过程相关[25]。因此可将DGKα基因作为影响猪脂类代谢活动重要候选基因之一。HE等研究发现在猪中LPIN1基因与肌内脂肪沉积及肌肉品质存在显著相关性,可影响肌内脂肪含量、瘦肉率和风味,故将LPIN1基因作为改善肌肉品质的候选基因之一[26]。这些基因现在已知的功能均与能量代谢和氧化磷酸化相关,但是在猪中影响组织脂类代谢的机制还有待深入研究。
育肥期期间日增重是猪的一种重要的经济性状,其受到肌肉发育直接影响。本研究发现了一些在松辽黑猪和长白猪之间显著差异的信号通路和代谢途径与肌肉发育有关,可能是影响育肥期期间日增重差异的因素。这些信号通路和代谢途径包括肌动蛋白调控过程、磷脂酰肌醇3-激酶/丝氨酸-苏氨酸蛋白激酶(phosphoinositide-3-kinase/serine-threonine kinase, PI3K- AKt)信号通路、单磷酸腺苷活化蛋白激酶(AMP- activated protein kinase, AMPK)信号通路,参与这些生物过程的基因包括成纤维因子1(fibroblast growth factor 1, FGF1)、成纤维因子10(fibroblast growth factor 10, FGF10)、成纤维细胞生长因子受体1(fibroblast growth factor receptor 1, FGFR1)、肌醇多磷酸-1-磷酸酶(inositol polyphosphate-1-phosphatase, INPP1)、磷脂酰肌醇-3-激酶调节亚基1(phosphoinositide-3-kinase regulatory subunit 1, PIK3R1)、血管生成素2(angiopoietin 2, ANGPT2)。研究已经证实,FGF1在细胞增殖和细胞分化等多种生物学过程中发挥作用[27];FGF10在多种细胞的增殖、迁移、分化和生存中起到重要的支持作用[28]。PIK3R1基因可编码磷脂酰肌醇3激酶(phosphoinositide-3-kinase, PIK3)的调节亚基p85α蛋白[29,30],而PIK3作用于多种细胞外因子的信号转导,在细胞的生长、增殖、分化和凋亡等多种生命活动中起到重要作用[31]。可发现这些基因多数与细胞增殖、生长和分化等生命活动相关。在功能方面,多数与肿瘤细胞相关,但与肌肉细胞增殖生长的相关报道很少。因此,这些基因可作为研究肌肉发育的候选基因进行深入研究和验证。
在生物学功能分析中,本研究发现生物素代谢过程最为显著。生物素,又称为维生素H、辅酶R,是一种在机体内生物活动过程中必不可少的辅酶因子[32]。GO功能分析发现全羧化酶合成酶(holocarboxylase synthetase, HLCS)和生物素酶(biotinidase, BTD)基因参与了生物素代谢过程。HLCS作用是能够催化生物素与羧化酶及组蛋白结合[33,34,35]。DO等在对猪饲料利用效率的研究中发现该基因与剩余采食量性状具有显著关联性,发现全羧化酶合成酶活性丧失时会对机体组织的葡萄糖、脂肪及蛋白质的代谢产生重要影响[36]。而在本研究中发现HLCS基因在两个品种猪背最长肌组织中表达显著差异,提示HLCS可能参与了肌肉组织的生物代谢活动,与猪生长性状具有潜在联系。BTD缺乏时会导致肠道对生物素的摄取能力下降,导致体内生物素缺乏,使依赖于生物素的各种羧化酶(丙酰辅酶A羧化酶、丙酮酰羧化酶、乙酰辅酶A 羧化酶和甲基巴豆酰辅酶A 羧化酶)的活性下降,影响糖类、脂肪及氨基酸代谢活动[37]。
在本研究中通过对差异表达的lncRNA进行靶基因预测和功能分析,3个差异表达lncRNA与基因之间可能存在着潜在的调控关系,分别是TCONS- 00024150、TCONS-00041713、TCONS-00041712。其中新lncRNA TCONS-00041713和TCONS-00041712位于锌指蛋白7(zinc finger protein 7, ZNF7)基因的上游。ZNF7属于锌指蛋白家族,前人研究发现,许多锌指蛋白家族成员在脂肪生成过程中起到关键的作用[38,39]。同时,在本研究发现新lncRNA TCONS- 00024150与FGFR1基因间存在碱基互补配对的关系,FGFR1在脂肪组织中最先被发现,其可能在血管新生、糖脂调节、新陈代谢等方面发挥作用[40]。本研究表明新lncRNAs可能参与了肌肉组织的脂类代谢活动,为解释猪品种间脂肪性状差异性提供新的思路,可为脂肪性状相关的基因研究奠定理论基础。
综上所述,通过对松辽黑猪和长白猪背最长肌之间差异表达基因的鉴定与分析,筛选出多个参与肌肉发育和脂类代谢的候选基因,其包括HLCS、BTD、DGKα、LPIN1、FGF1、FGF10、FGFR1和ZNF7基因。这些基因可能对研究松辽黑猪和长白猪两品种间肌肉发育和肌肉脂类代谢的差异性研究具有参考价值。
4 结论
本研究通过高通量测序技术对松辽黑猪和长白猪背最长肌肌肉组织进行RNA测序,通过基因比对分析,筛选出4 239个编码基因差异mRNA,其中2 023个显著上调,2 216个显著下调;与此同时,筛选出178个差异表达的lncRNAs,84个显著上调,94个显著下调。通过对差异表达基因进行GO功能分析和KEGG通路分析共筛选出了HLCS、BTD、DGKα、LPIN1、FGF1、FGF10、FGFR1和ZNF7等与脂类代谢和肌肉发育相关的基因,并且参与了相关的信号通路和生物代谢途径。结合差异表达基因分析和靶基因预测的结果发现,几对差异表达lncRNA和mRNA之间可能存在着潜在的调控关系。如新lncRNA TCONS_00041713和TCONS_00041712在ZNF7基因的上游;lncRNA ENSSSCT00000034982在ERNBP基因的下游且与HCFC1存在碱基互补配对的关系;新lncRNA TCONS_00024150分别与FGFR1和 ENSSSCT00000034063基因间存在碱基互补配对的关系。这些结果为了解松辽黑猪和长白猪之间肌内脂肪含量和肌肉发育性状差异的分子机制提供了新的参考。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[M]//SCRIVER C R, BEAUDET A L, SLY W S.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}