Effects of Histone Acetylation on Ganoderma lucidum Growth, Polysaccharide and Ganoderic Acid Biosynthesis
ZHANG ZongYuan1,2, JIANG YongMei1, ZHANG WenXian,11 College of Life Sciences, Fujian Normal University, Fuzhou 350108 2 Institute of Biology Co., Ltd., Henan Academy of Sciences, Zhengzhou 450008
Abstract 【Objective】Histone acetylation modification plays an important role in the growth, development and metabolic synthesis of fungi. Few epigenetic studies of higher medicinal fungi were reported at present. In this study, the effects of histone acetylation on the growth and development of Ganoderma lucidum (G. lucidum) and the synthesis of its main metabolites were studied by adding chemical epigenetic inhibitor octanedianiline hydroxamic acid (SAHA), which provided a theoretical basis for improving the biosynthesis of G. lucidum polysaccharides and Ganoderma acid (GA) by epigenetic means.【Method】A two-stage cultivation, liquid fermentation combined with static cultivation, was applied to culture G. lucidum. The cell was treated with different concentration of SAHA (0, 0.6, 60, 120, and 180 μmol?L -1) during the liquid static cultivation of G. lucidum. Biomass, sugar consumption, mycelial mat formation, mycelial morphology, sporulation and biosynthesis of GA and G.lucidum polysaccharides were measured or observed by conventional methods. Histone acetylation levels of G. luidum were examined by Western blot, the relative expression levels of polysaccharides biosynthesis genes (e.g. ugp, gls, and pgm), GA biosynthesis genes (e.g. hmg, sqs, se, and ls) and global regulator vet, LaeA gene were detected by qRT-PCR. 【Result】The acetylation level of histone H4 in G. lucidum treated with SAHA increased to 1.6 times as much as that under control group. SAHA inhibited the growth of G. lucidum mycelia and the production of pigments, and changed the morphology of mycelia. The formation of spores was also inhibited, and the higher the concentration of SAHA, the more obvious the inhibition degree. SAHA treatment significantly increased the yield of G. lucidum polysaccharides, up to 50%, and the biosynthesis of GA was inhibited, which decreased by 13%-27% compared with the control. The results of qRT-PCR analysis showed that the gene expression of the key enzymes in G.lucidum polysaccharides and GA synthesis were up-regulated in different degrees under SAHA treatment. The gene expression of the key enzymes in polysaccharides synthesis were increased by 1.5-3.5 times and that of the key enzymes in GA synthesis by 1.8-12.1 times. The expression of vet and LaeA genes, the global regulators, were inhibited, which was 11.3%-62.4% of the control group.【Conclusion】Histone acetylation could regulate the growth and development of G. lucidum through global regulatory factors, thus affecting the biosynthesis of GA, while histone acetylation also had an effect on G. lucidum polysaccharides biosynthesis. Keywords:Ganoderme lucidum;histone acetylation;ganoderic acid;Ganoderma lucidum polysaccharide;SAHA
PDF (1201KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张宗源, 蒋咏梅, 章文贤. 组蛋白乙酰化对灵芝生长、灵芝多糖和灵芝酸生物合成的影响[J]. 中国农业科学, 2020, 53(3): 632-641 doi:10.3864/j.issn.0578-1752.2020.03.014 ZHANG ZongYuan, JIANG YongMei, ZHANG WenXian. Effects of Histone Acetylation on Ganoderma lucidum Growth, Polysaccharide and Ganoderic Acid Biosynthesis[J]. Scientia Acricultura Sinica, 2020, 53(3): 632-641 doi:10.3864/j.issn.0578-1752.2020.03.014
A:灵芝组蛋白免疫印迹;B:灵芝组蛋白乙酰化水平比较。不同小写字母表示差异显著(P<0.05)。下同 Fig. 3Effect of SAHA on the level of histone acetylation of G. lucidum
A: The Western blotting of histone of G. lucidum. B: Detection of the level of histone acetylation in G. lucidum by Western blot assay. Different lowercase letters indicate significant differences (P<0.05). The same as below
A:液体静置培养6 d时0(Ⅰ)、120 μmol∙L-1(Ⅱ)、180 μmol∙L-1(Ⅲ)SAHA处理下灵芝菌丝体形态及培养9 d时0.09% DMSO(Ⅳ)、0(Ⅴ)、0.6 μmol∙L-1(Ⅵ)、60 μmol∙L-1(Ⅶ)、120 μmol∙L-1(Ⅷ)及180 μmol∙L-1(Ⅸ)SAHA处理下的灵芝孢子;B:不同处理下灵芝孢子数目 Fig. 4Effects of SAHA on aerial mycelia morphology and spore formation in liquid static culture of G. lucidum
A: The mycelia morphology on liquid surface were cultured on day 6 at 0 μmol∙L-1 (Ⅰ), 120 μmol∙L-1 (Ⅱ), 180 μmol∙L-1 (Ⅲ) SAHA and the mycelia with spores on liquid surface were cultured on day 9 at 0.09% DMSO (Ⅳ) and 0 μmol∙L-1 (Ⅴ), 0.6 μmol∙L-1 (Ⅵ), 60 μmol∙L-1 (Ⅶ), 120 μmol∙L-1 (Ⅷ), 180 μmol∙L-1 (Ⅸ) SAHA, respectively. B: Sporulation of G. lucidum after treatment with 0.09% DMSO and different concentration of SAHA
SHIAO MS, LEE KR, LIN LJ, WANG CT . Natural products and biological activities of the Chinese medicinal fungus Ganoderma lucidum. ACS Symposium Series (USA), 1994,547:342-354. [本文引用: 1]
SONEY, OKUDAR, WADAN, KISHIDAE, MISAKIA . Structures and antitumor activities of the polysaccharides isolated from fruiting body and the growing culture of mycelium of Ganoderma lucidum. Agricultural and Biological Chemistry, 1985,49(9):2641-2653. [本文引用: 1]
XU JW, ZHAOW, ZHONG JJ . Biotechnological production and application of ganoderic acids Applied Microbiology and Biotechnology, 2010,87(2):457-466. [本文引用: 1]
LIU GQ, ZHAOY, WANG XL, ZHU CY . Biosynthesis and fermentation control of polysaccharides from Ganoderma lucidum. Mycosystema, 2011,30(2):198-205. (in Chinese) [本文引用: 2]
NING YB, WANG HY, QIAOK, LIU XM, WANG KY . Induced resistance by polysaccharides isolated from Ganoderma lucidum in tomato against gray mold. Scientia Agricultura Sinica, 2016,49(11):2103-2112. (in Chinese) [本文引用: 1]
ZHANG ZX, WANG HY, WANG KY, WANGD, JIANG LL . Induction effect of sharp eyespot of wheat and the effect of wheat growth after Ganoderma lucidum polysaccharides (GLP) seed dressing. Scientia Agricultura Sinica, 2018,51(1):96-104. (in Chinese) [本文引用: 1]
FANG QH, ZHONG JJ . Two-stage culture process for improved production of ganoderic acid by liquid fermentation of higher fungus Ganoderma lucidum. Biotechnology Progress, 2002,18(1):51-54. [本文引用: 3]
KONG XJ, VAN DIEPENINGENA D, VAN DER LEE TA J, WAALWIJKC, XUJ S, XUJ, ZHANGH, CHENW Q, FENGJ . The Fusarium graminearum histone acetyltransferases are important for morphogenesis, DON biosynthesis, and pathogenicity. Frontiers in Microbiology, 2018,9:654. doi: 10.3389/fmicb.2018.00654. [本文引用: 1]
HEDTKEM, RAUSCHERS, ROHRIGJ, RODRIGUEZ-ROMEROJ, YU ZZ, FISCHERR . Light-dependent gene activation in Aspergillus nidulans is strictly dependent on phytochrome and involves the interplay of phytochrome and white collar-regulated histone H3 acetylation. Molecular Microbiology, 2015,97(4):733-745. doi: 10.1111/mmi.13062. [本文引用: 1]
BRENNAA, GRIMALDIB, FILETICIP, BALLARIOP . Physical association of the WC-1 photoreceptor and the histone acetyltransferase NGF-1 is required for blue light signal transduction in Neurospora crassa. Molecular Biology of the Cell, 2012,23(19):3863-3872. doi: 10.1091/mbc.E12-02-0142. [本文引用: 1]
CANOVASD, MARCOS AT, GACEKA, RAMOS MS, GUTIERREZG, REYES-DOMINGUEZY, STRAUSSJ . The histone acetyltransferase GcnE (GCN5) plays a central role in the regulation of Aspergillus asexual development. Genetics, 2014,197(4):1175-1189. doi: 10.1534/genetics.114.165688. [本文引用: 1]
TRIBUSM, BAUERI, GALEHRJ, RIESERG, TROJERP, BROSCHG, LOIDLP, HAASH, GRAESSLES . A novel motif in fungal class 1 histone deacetylases is essential for growth and development of Aspergillus. Molecular Biology of the Cell, 2010,21(2):345-353. doi: 10.1091/mbc.E09-08-0750. [本文引用: 1]
LAN HH, SUN RL, FANK, YANG KL, ZHANGF, NIE XY, WANG XN, ZHUANG ZH, WANG SH . The Aspergillus flavus histone acetyltransferase AflGcnE regulates morphogenesis, aflatoxin biosynthesis, and pathogenicity. Frontiers in Microbiology, 2016,7:e74030. doi: 10.3389/fmicb.2016.01324. [本文引用: 2]
ZHANGQ, CHEN LF, YUX, LIUH, AKHBERDIO, PANJ, ZHU XD . A B-type histone acetyltransferase HAT1 regulates secondary metabolism, conidiation, and cell wall integrity in the taxol-producing fungus Pestalotiopsis microspora. Journal of basic microbiology, 2016,56(12):1380-1391. [本文引用: 2]
TURNER EL, MALO ME, PISCLEVICH MG, DASH MD, DAVIES GF, ARNASON TG, HARKNESS TA A . The Saccharomyces cerevisiae anaphase-promoting complex interacts with multiple histone-modifying enzymes to regulate cell cycle progression. Eukaryotic Cell, 2010,9(10):1418-1431. doi: 10.1128/ec.00097-10. [本文引用: 1]
CAIQ, WANG JJ, SHAOW, YING SH, FENGM G . Rtt109-dependent histone H3 K56 acetylation and gene activity are essential for the biological control potential of Beauveria bassiana Pest Management Science2018, 74(11): 2626-2635. DOI: 10.1002/ps.5054. [本文引用: 1]
BASIMIAT, REZAEES, ZAMANIZADEH HR, MOUSAVIA . SAHA, histone deacetylase inhibitor causes reduction of aflatoxin production and conidiation in the Aspergillus flavus. General Pharmacology the Vascular System, 2013,30(3):357-366. [本文引用: 3]
YANG KL, ZHUANG ZH, ZHANGF, SONG FQ, ZHONGH, RAN FL, YUS, XU GP, LAN FX, WANG SH . Inhibition of aflatoxin metabolism and growth of Aspergillus flavus in liquid culture by a DNA methylation inhibitor. Food Additives & Contaminants: Part A, 2015,32(4):554-563. doi: 10.1080/19440049. 2014.972992. [本文引用: 1]
LIN ZP, SUN AC, LIU YA . Studies on the submerged cultivation of Ling Chin(Ganoderma sp.). Acta microbiologica Sinica, 1973,13(2):63-68. (in Chinese) [本文引用: 1]
WANGQ, LI YY, YANG XX, WANGL, JIH, WANGH . Submerged culture of Ganoderma lucidum and its application. Acta Edulis Fungicide, 1994,1(2):41-44. (in Chinese) [本文引用: 1]
LEE SY, KANG TS, LEE MC . Condition of exo-polysacchride production from submerged mycelial culture of Ganoderma lucidum by using air-lift fermenter system. Korean Journal of Biotechnology and Bioengineering, 1998,13:547-553. [本文引用: 1]
FANG QH, ZHONG JJ . Simultaneous production of polysaccharide and ganoderic acid by submerged fermentation of Ganoderma lucidum. Journal of East China University of Science and Technology (Natural Science Edition), 2001,27(3):254-257. (in Chinese) [本文引用: 1]
LIANG CX, LI YB, XU JW, WANG JL, MIAO XL, TANG YJ, GU TY, ZHONG JJ . Enhanced biosynthetic gene expressions and production of ganoderic acids in static liquid culture of Ganoderma lucidum under phenobarbital induction. Applied Microbiology & Biotechnology, 2010,86(5):1367-1374. [本文引用: 1]
XU YN, ZHONG JJ . Impacts of calcium signal transduction on the fermentation production of antitumor ganoderic acids by medicinal mushroom Ganoderma lucidum. Biotechnology Advances, 2012,30(6):1301-1308. [本文引用: 1]
ZHAOW, XU JW, ZHONG JJ . Enhanced production of ganoderic acids in static liquid culture of Ganoderma lucidum under nitrogen- limiting conditions. Bioresource Technology, 2011,102(17):8185-8190. [本文引用: 1]
ZHANG WX, ZHONG JJ . Effect of oxygen concentration in gas phase on sporulation and individual ganoderic acids accumulation in liquid static culture of Ganoderma lucidum. Journal of Bioscience and Bioengineering, 2010,109(1):37-40. [本文引用: 3]
JI SL, LIUR, REN MF, LI HJ, XU JW . Enhanced production of polysaccharide through the overexpression of homologous uridine diphosphate glucose pyrophosphorylase gene in a submerged culture of Lingzhi or Reishi Medicinal Mushroom,Ganoderma lucidum(Higher Basidiomycetes). International Journal of Medicinal Mushrooms, 2015,17(5):435-442. [本文引用: 2]
LIUK, LI HJ, ZHANG DH, YUE TH, LIN, XU JW . Heterologous expression of vitreoscilla hemoglobin in ganoderma lingzhi for increased exopolysaccharide production Acta Edulis Fungi, 2017,24(3):35-41.(in Chinese) [本文引用: 1]
WILLIAMS RB, HENRIKSON JC, HOOVER AR, LEE AE, CICHEWICZ RH . Epigenetic remodeling of the fungal secondary metabolome Organic & Biomolecular Chemistry, 2008,6(11):1895-1897. [本文引用: 1]
FISCH KM, GILLASPY AF, GIPSONM, HENRIKSON JC, HOOVER AR, JACKSONL, NAJAR FZW GELEH, CICHEWICZR H. , Chemical induction of silent biosynthetic pathway transcription in Aspergillus niger. Journal of Industrial Microbiology & Biotechnology, 2009,36(9):1199-1213. [本文引用: 1]
TANG YJ, ZHONG JJ . Role of oxygen supply in submerged fermentation of Ganoderma lucidum for production of Ganoderma polysaccharide and ganoderic acid. Enzyme and Microbial Technology, 2003,32:478-484. [本文引用: 1]
LAN LW . Study on the regulation of DNA methylation on ganoderic acids biosynthesis in Ganoderma lucidum [D]. Fuzhou: Fujian Normal University, 2016. ( in Chinese) [本文引用: 3]
HU BB, LIN LB, WEI YL, JI XL, ZHANGQ . An efficient fungal protein extraction method China Biotechnology, 2013,33(9):53-58. [本文引用: 1]
ZHANG WX, TANG YJ, ZHONG J J . Impact of oxygen level in gaseous phase on gene transcription and ganoderic acid biosynthesis in liquid static cultures of Ganoderma lucidum. Bioprocess & Biosystems Engineering, 2010,33(6):683-690. [本文引用: 1]
LIVAK KJ, SCHMITTGEN TD . Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCT method Methods, 2001,25(4):402-408. [本文引用: 1]
ZHANGH . The regulation of histone deacetylase Rpd3 on meiastatin biosynthesis and growth and development in Penicillium citrinum [D]. Chongqing: Southwest University, 2017. ( in Chinese) [本文引用: 1]
YE LQ . Regulation of Trichostatin A on toxin production and growth of Aspergillus flavus [D]. Fuzhou: Fujian Agriculture and Forestry University, 2015. ( in Chinese) [本文引用: 1]
LEEI, OH JH, SHWAB EK, DAGENAIS TR, ANDESD, KELLER NP . HdaA, a class 2 histone deacetylase of Aspergillus fumigatus, affects germination and secondary metabolite production. Fungal Genetics & Biology Fg & B, 2009,46(10):782. [本文引用: 1]
CHENH, YANG HL, LIU GQ . Biosynthesis and fermentation control of triterpenoids from Ganoderma lingzhi Mycosystema, 2015,34(1):1-9. (in Chinese) [本文引用: 1]
CALVO AM, WILSON RA, WOO BJ, KELLER NP . Relationship between secondary metabolism and fungal development Microbiology and Molecular Biology Reviews, 2002,66(3):447-459. [本文引用: 1]
KALE SP, MILDE LT, MARISAK, FRISVAD JC, KELLER NP, BOK JW . Requirement of LaeA for secondary metabolism and sclerotial production in Aspergillus flavus. Fungal Genetics & Biology, 2008,45(10):1422-1429. [本文引用: 1]
ATOUIA, KASTNERC, LAREY CM, THOKALAR, ETXEBESTEO, ESPESO EA, FISCHERR, CALVO AM . Cross-talk between light and glucose regulation controls toxin production and morphogenesis in Aspergillus nidulans. Fungal Genetics and Biology, 2010,47(12):962-972. [本文引用: 1]
CHETTRIP, CALVO AM, CARY JW, DHINGRAS, GUOY, MCDOUGAL RL, BRADSHAW RE . The veA gene of the pine needle pathogen Dothistroma septosporum regulates sporulation and secondary metabolism. Fungal Genetics and Biology, 2012,49(2):141-151. [本文引用: 1]
BAYRAM OS, BAYRAMO, VALERIUSO, PARK HS, IRNIGERS, GERKEJ, NIM, HAN KH, YU JH, BRAUS GH . LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity PLoS Genetics, 2010,6(12):e1001226. [本文引用: 1]
 ,11
,11 
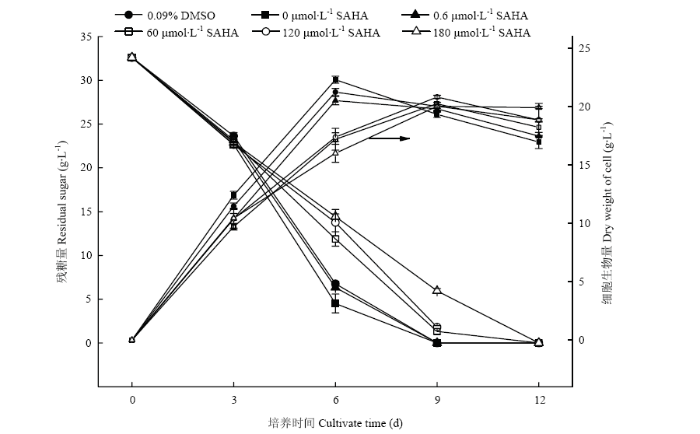 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT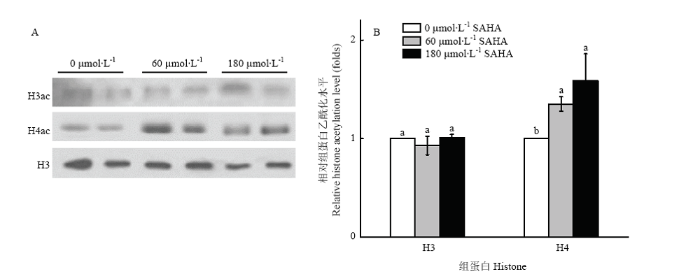 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT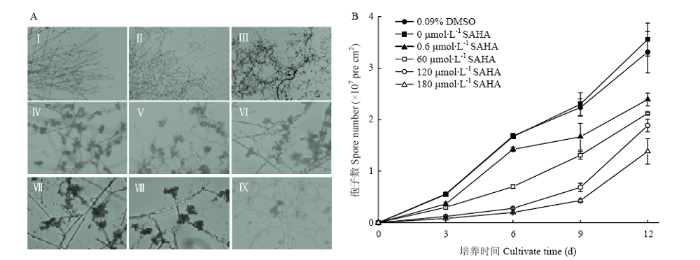 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT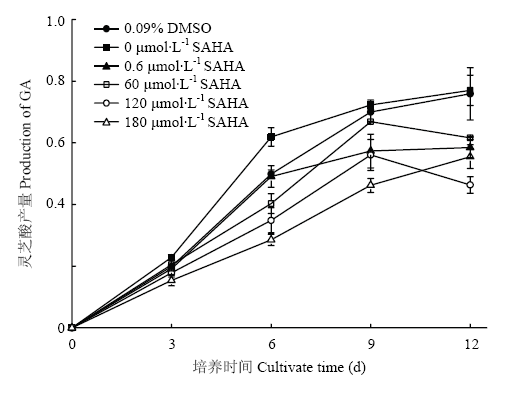 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT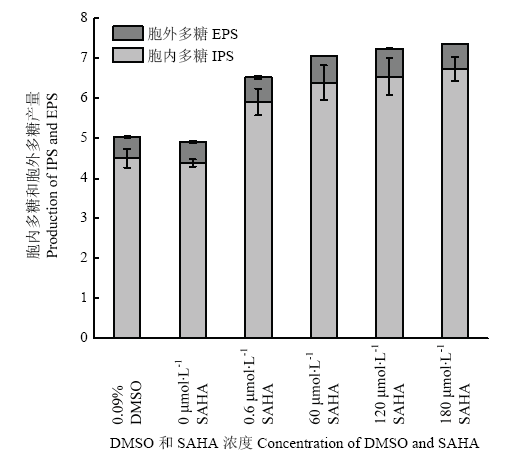 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT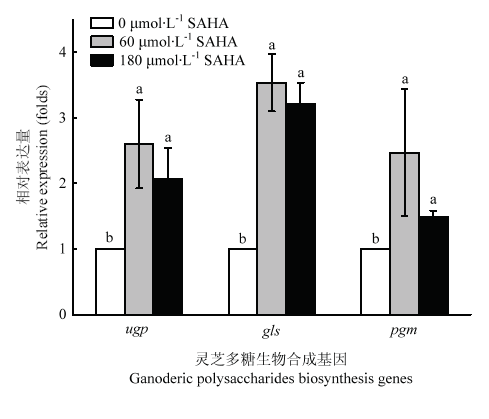 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT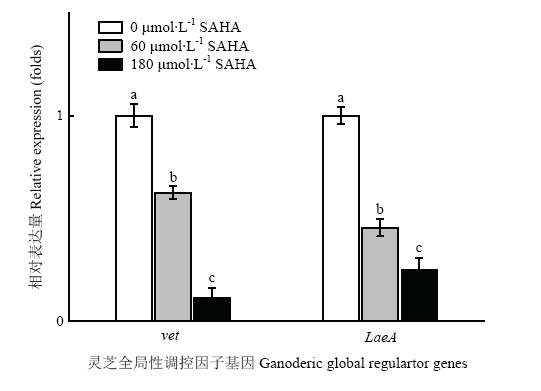 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}